

Bioaugmentation Strategies for Enhancing Methane Production from Shrimp Processing Waste through Anaerobic Digestion
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Substrate Characteristics
2.2. Microbial Sources
2.3. Set-Up of Anaerobic Digestion Experiment
2.4. Biogas and Organic Acid Detection
2.5. Microbial Community Analysis
2.6. Statistical Analysis
3. Results and Discussion
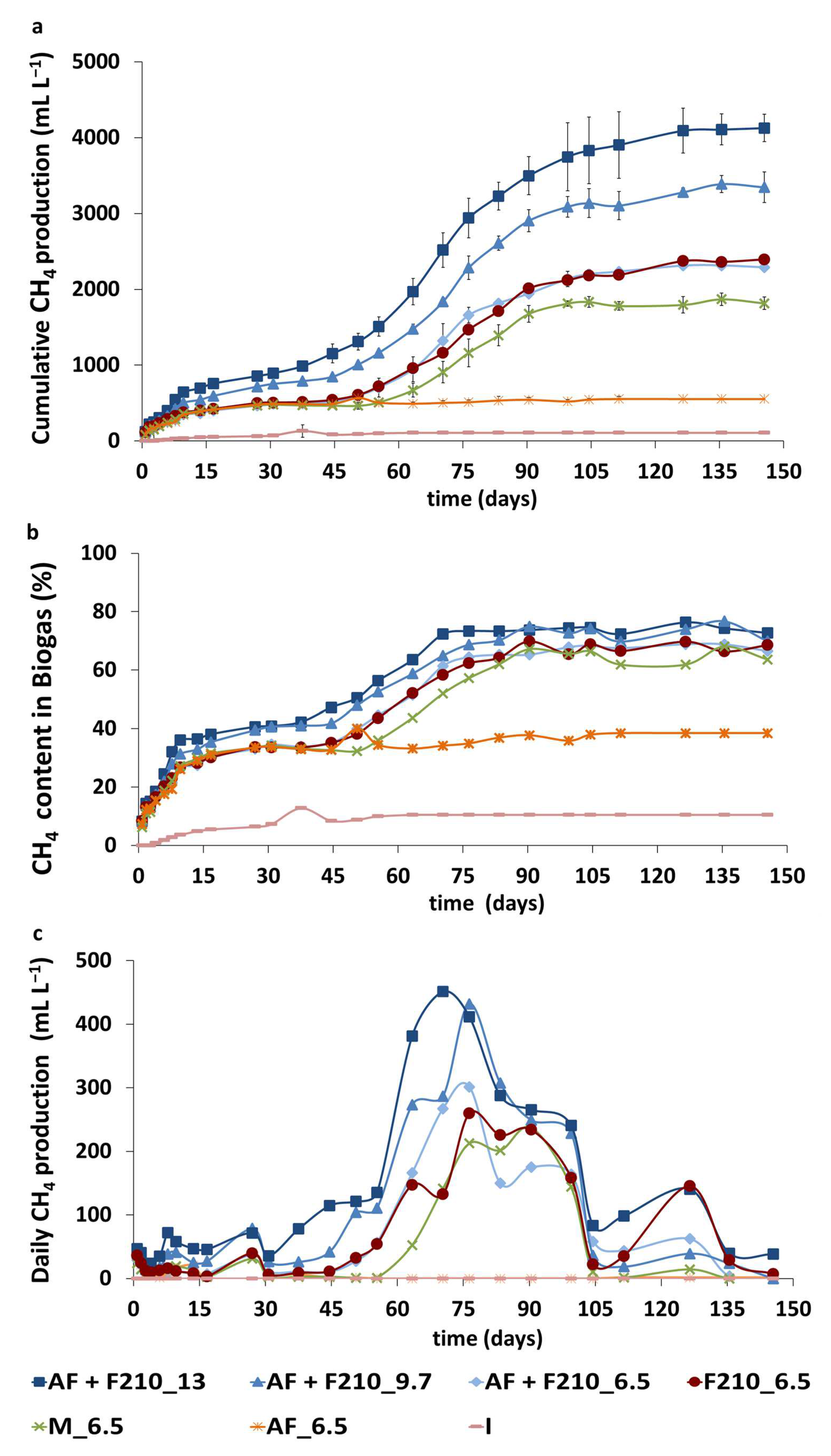
3.1. Biogas Production
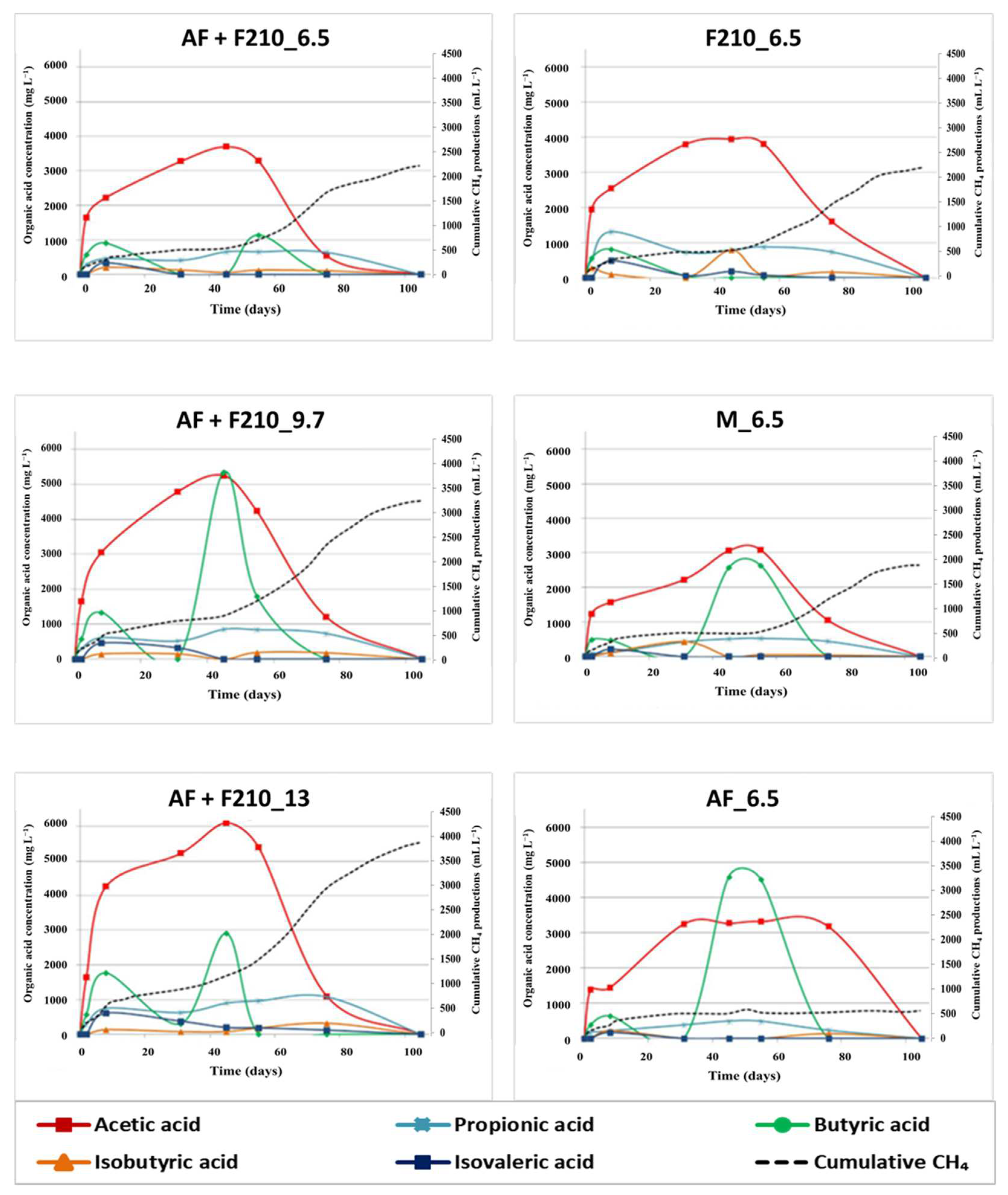
3.2. Organic Acid Measurements
3.3. SPW Removal and Composition
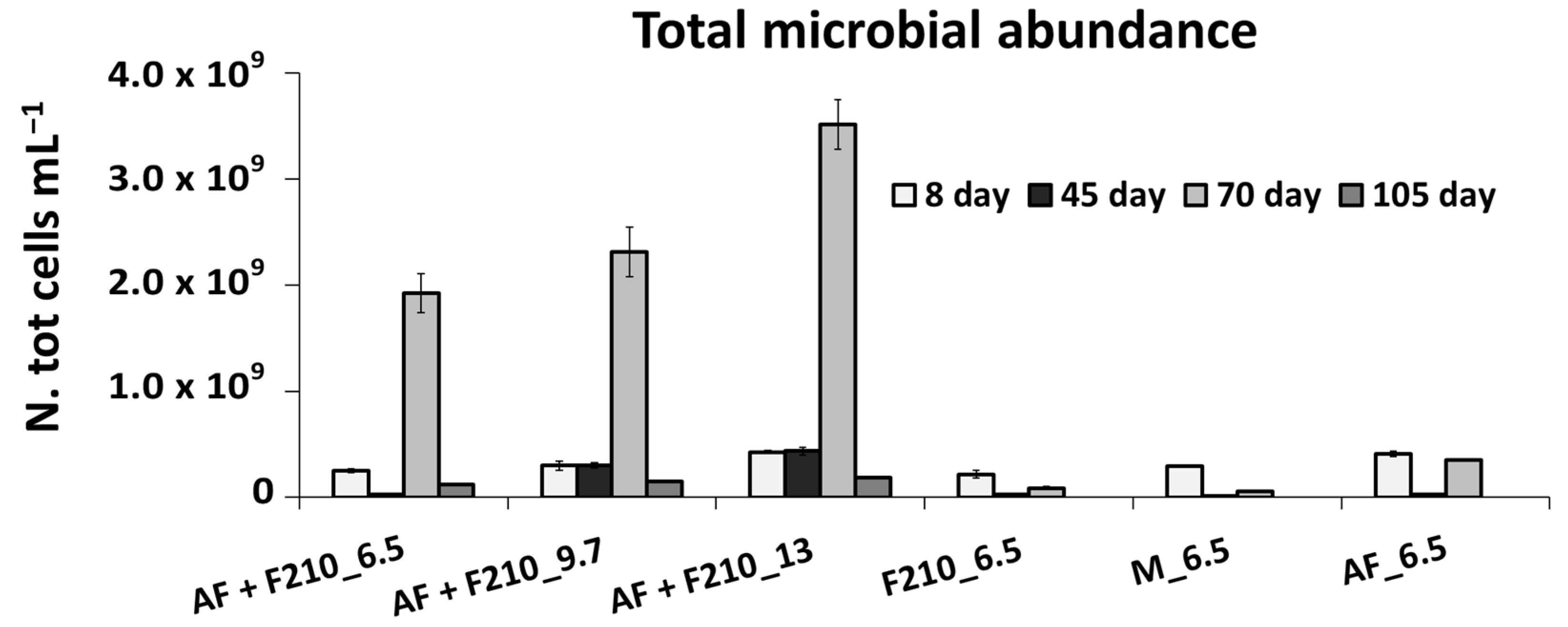
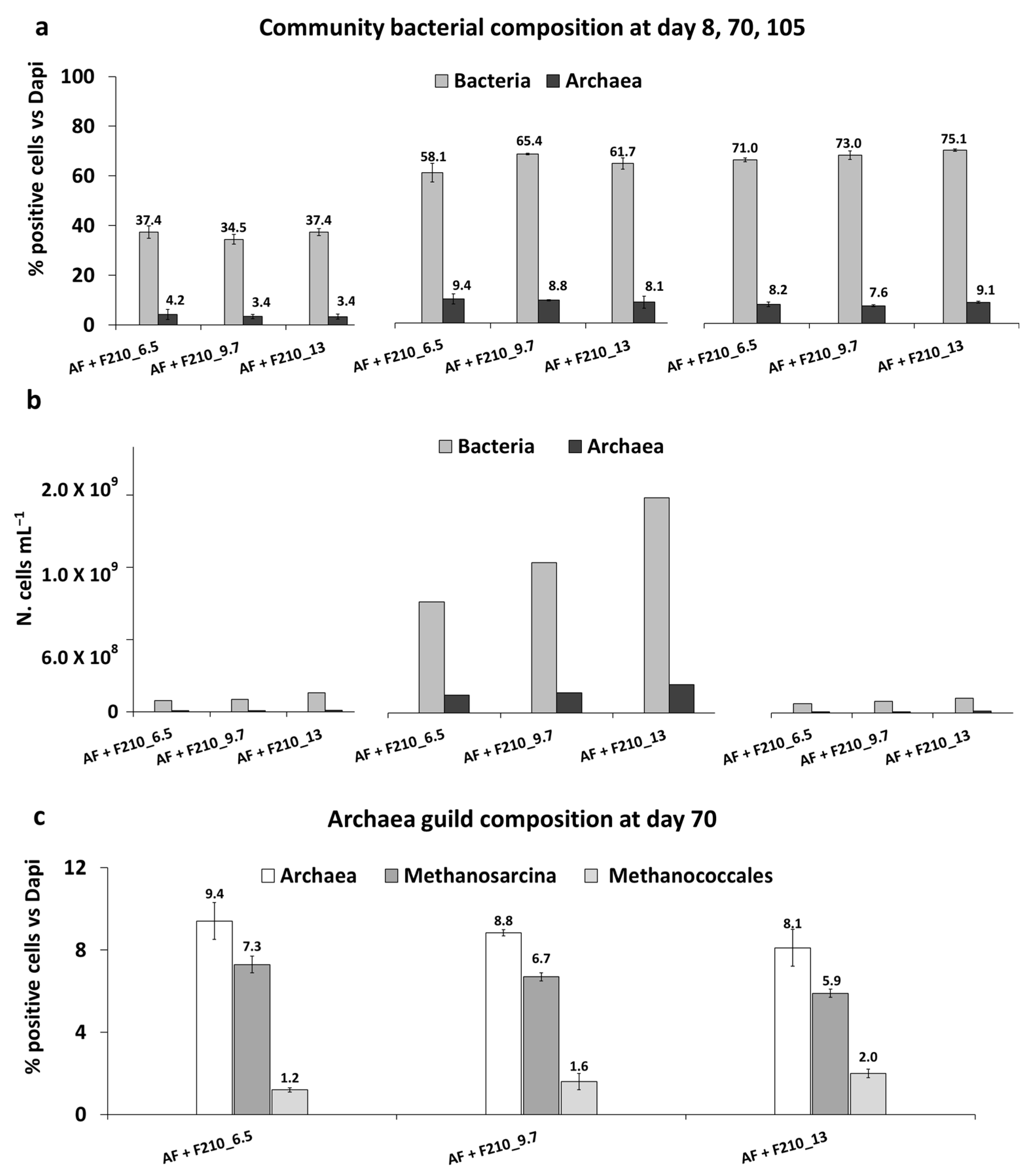
3.4. Microbiological Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. Brief to the State of World Fisheries and Aquaculture; FAO, Fisheries and Aquaculture Department: Rome, Italy, 2022; ISBN 9789251363645. [Google Scholar]
- Mathew, G.M.; Mathew, D.C.; Sukumaran, R.K.; Sindhu, R.; Huang, C.C.; Binod, P.; Sirohi, R.; Kim, S.H.; Pandey, A. Sustainable and eco-friendly strategies for shrimp shell valorization. Environ. Pollut. 2020, 267, 115656. [Google Scholar] [CrossRef] [PubMed]
- Kandra, P.; Challa, M.M.; Kalangi Padma Jyothi, H. Efficient use of shrimp waste: Present and future trends. Appl. Microbiol. Biotechnol. 2012, 93, 17–29. [Google Scholar] [PubMed]
- FAO. Increased Production of Farmed Shrimp Leads to Improved International Trade. GLOBEFISH-Analysis Andinformation on World Fish Trade. 2017. Available online: https://www.fao.org/in-action/globefish/market-reports/resource-detail/en/c/989543/ (accessed on 5 April 2023).
- Mao, X.; Guo, N.; Sun, J.; Xue, C. Comprehensive utilization of shrimp waste based on biotechnological methods: A review. J. Clean. Prod. 2017, 143, 814–823. [Google Scholar]
- Suryawanshi, N.; Jujjavarapu, S.E.; Ayothiraman, S. Marine shell industrial wastes–an abundant source of chitin and its derivatives: Constituents, pretreatment, fermentation, and pleiotropic applications-a revisit. Int. J. Environ. Sci. Technol. 2019, 16, 3877–3898. [Google Scholar]
- Nirmal, N.P.; Santivarangkna, C.; Rajput, M.S.; Benjakul, S. Trends in shrimp processing waste utilization: An industrial prospective. Trends Food Sci. Technol. 2020, 103, 20–35. [Google Scholar]
- Ali, G.; Sharma, M.; Salama, E.S.; Ling, Z.; Li, X. Applications of chitin and chitosan as natural biopolymer: Potential sources, pretreatments, and degradation pathways. Biomass Convers. Biorefinery 2022, 1–15. [Google Scholar] [CrossRef]
- Digman, B.; Kim, D.-S. Alternative energy from food processing wastes. Environ. Prog. 2008, 27, 524–537. [Google Scholar]
- Xu, F.; Li, Y.; Ge, X.; Yang, L.; Li, Y. Anaerobic digestion of food waste—Challenges and opportunities. Bioresour. Technol. 2018, 247, 1047–1058. [Google Scholar] [CrossRef]
- Dollhofer, V.; Podmirseg, S.M.; Callaghan, T.M.; Griffith, G.W.; Fliegerová, K. Anaerobic fungi and their potential for biogas production. In Biogas Science and Technology; Springer: Cham, Switzerland, 2015; pp. 41–61. ISBN 9783319219936. [Google Scholar]
- Young, D.; Dollhofer, V.; Callaghan, T.M.; Reitberger, S.; Lebuhn, M.; Benz, J.P. Isolation, identification and characterization of lignocellulolytic aerobic and anaerobic fungi in one- and two-phase biogas plants. Bioresour. Technol. 2018, 268, 470–479. [Google Scholar] [CrossRef]
- Cheng, Y.; Shi, Q.; Sun, R.; Liang, D.; Li, Y.; Li, Y.; Jin, W.; Zhu, W. The biotechnological potential of anaerobic fungi on fiber degradation and methane production. World J. Microbiol. Biotechnol. 2018, 34, 1–8. [Google Scholar] [CrossRef]
- Lembo, G.; Rosa, S.; Miritana, V.M.; Marone, A.; Massini, G.; Fenice, M.; Signorini, A. Thermophilic anaerobic digestion of second cheese whey: Microbial community response to H2 addition in a partially immobilized anaerobic hybrid reactor. Processes 2021, 9, 43. [Google Scholar] [CrossRef]
- Atelge, M.R.; Krisa, D.; Kumar, G.; Eskicioglu, C.; Nguyen, D.D.; Chang, S.W.; Atabani, A.E.; Al-Muhtaseb, A.H.; Unalan, S. Biogas Production from Organic Waste: Recent Progress and Perspectives. Waste Biomass Valorization 2020, 11, 1019–1040. [Google Scholar] [CrossRef]
- Ellacuriaga, M.; García-Cascallana, J.; Gómez, X. Biogas Production from Organic Wastes: Integrating Concepts of Circular Economy. Fuels 2021, 2, 144–167. [Google Scholar] [CrossRef]
- Kundu, D.; Dutta, D.; Samanta, P.; Dey, S.; Sherpa, K.C.; Kumar, S.; Dubey, B.K. Valorization of wastewater: A paradigm shift towards circular bioeconomy and sustainability. Sci. Total Environ. 2022, 848, 157709. [Google Scholar] [CrossRef]
- Ivanovs, K.; Spalvins, K.; Blumberga, D. Approach for modelling anaerobic digestion processes of fish waste. Energy Procedia 2018, 147, 390–396. [Google Scholar] [CrossRef]
- Choe, U.; Mustafa, A.M.; Lin, H.; Xu, J.; Sheng, K. Effect of bamboo hydrochar on anaerobic digestion of fish processing waste for biogas production. Bioresour. Technol. 2019, 283, 340–349. [Google Scholar] [CrossRef]
- Chairattanawat, C.; Yulisa, A.; Hwang, S. Effect of fish waste augmentation on anaerobic co-digestion of sludge with food waste. Bioresour. Technol. 2022, 361, 127731. [Google Scholar] [CrossRef] [PubMed]
- Ferdeș, M.; Zăbavă, B.Ș.; Paraschiv, G.; Ionescu, M.; Dincă, M.N.; Moiceanu, G. Food Waste Management for Biogas Production in the Context of Sustainable Development. Energies 2022, 15, 6268. [Google Scholar] [CrossRef]
- Zappi, M.E.; Revellame, E.; Fortela, D.L.; Hernandez, R.; Gang, D.; Holmes, W.; Sharp, W.; Picou-Mikolajczyk, A.; Nigam, K.D.P.; Bajpai, R. Evaluation of the Potential to Produce Biogas and Other Energetic Coproducts Using Anaerobic Digestion of Wastewater Generated at Shrimp Processing Operations. Ind. Eng. Chem. Res. 2019, 58, 15930–15944. [Google Scholar] [CrossRef]
- European Commission. Direttiva (UE) 2018/2001 del Parlamento Europeo e del Consiglio, dell’11 Dicembre 2018, Sulla Promozione Dell’uso Dell’energia da Fonti Rinnovabili (Rifusione) (Testo Rilevante ai fini del SEE)2018/2001 del Parlamento Europeo e del Consiglio 2018. 2022. Available online: http://data.europa.eu/eli/dir/2018/2001/oj (accessed on 1 April 2023).
- Coppola, D.; Lauritano, C.; Esposito, F.P.; Riccio, G.; Rizzo, C.; de Pascale, D. Fish Waste: From Problem to Valuable Resource. Mar. Drugs 2021, 19, 116. [Google Scholar] [CrossRef]
- Caruso, G.; Floris, R.; Serangeli, C.; Di Paola, L. Fishery Wastes as a Yet Undiscovered Treasure from the Sea: Biomolecules Sources, Extraction Methods and Valorization. Mar. Drugs 2020, 18, 622. [Google Scholar] [CrossRef] [PubMed]
- De Boer, W.; Gerards, S.; Klein Gunnewiek, P.J.A.; Modderman, R. Response of the chitinolytic microbial community to chitin amendments of dune soils. Biol. Fertil. Soils 1999, 29, 170–177. [Google Scholar] [CrossRef]
- Ghosh, S.; Chowdhury, R.; Bhattacharya, P. Mixed consortia in bioprocesses: Role of microbial interactions. Appl. Microbiol. Biotechnol. 2016, 100, 4283–4295. [Google Scholar] [CrossRef] [PubMed]
- Gorrasi, S.; Izzo, G.; Massini, G.; Signorini, A.; Barghini, P.; Fenice, M. From polluting seafood wastes to energy. production of hydrogen and methane from raw chitin material by a two-phase process. J. Environ. Prot. Ecol. 2014, 15, 526–536. [Google Scholar]
- Izzo, G.; Rosa, S.; Massing, G.; Patriarca, C.; Fenice, M.; Fiocchetti, F.; Marone, A.; Varrone, C.; Signorini, A. From hypertrophic lagoons to bioenergy production. J. Environ. Prot. Ecol. 2014, 15, 537–546. [Google Scholar]
- Di Bonito, R.; Marone, A.; Massini, G.; Patriarca, C.; Rosa, S.; Signorini, A.; Varrone, C.; Viola, C.; Izzo, G. Characterization by length heterogeneity (LH)-PCR of a hydrogen-producing community obtained in dark fermentation using coastal lake sediment as an inoculum. Energy. Sustain. Soc. 2013, 3, 3. [Google Scholar] [CrossRef]
- Ferraro, A.; Dottorini, G.; Massini, G.; Mazzurco Miritana, V.; Signorini, A.; Lembo, G.; Fabbricino, M. Combined bioaugmentation with anaerobic ruminal fungi and fermentative bacteria to enhance biogas production from wheat straw and mushroom spent straw. Bioresour. Technol. 2018, 260, 364–373. [Google Scholar] [CrossRef]
- Marone, A.; Varrone, C.; Fiocchetti, F.; Giussani, B.; Izzo, G.; Mentuccia, L.; Rosa, S.; Signorini, A. Optimization of substrate composition for biohydrogen production from buffalo slurry co-fermented with cheese whey and crude glycerol, using microbial mixed culture. Int. J. Hydrogen Energy 2015, 40, 209–218. [Google Scholar] [CrossRef]
- Ferraro, A.; Massini, G.; Mazzurco Miritana, V.; Rosa, S.; Signorini, A.; Fabbricino, M. A novel enrichment approach for anaerobic digestion of lignocellulosic biomass: Process performance enhancement through an inoculum habitat selection. Bioresour. Technol. 2020, 313, 123703. [Google Scholar] [CrossRef]
- American Public Health Association. Standard Methods for the Examination of Water and Wastewater, 9th ed.; American Public Health Association: Washington, DC, USA, 1995. [Google Scholar]
- Miller, T.L.; Wolin, M.J. Formation of hydrogen and formate by Ruminococcus albus. J. Bacteriol. 1973, 116, 836–846. [Google Scholar] [CrossRef]
- Martínez-Sibaja, A.; Alvarado-Lassman, A.; Astorga-Zaragoza, C.M.; Adam-Medina, M.; Posada-Gómez, R.; Rodríguez-Jarquin, J.P. Volumetric gas meter for laboratory-scale anaerobic bioreactors. Meas. J. Int. Meas. Confed. 2011, 44, 1801–1805. [Google Scholar] [CrossRef]
- Logan, B.E.; Oh, S.E.; Kim, I.S.; Van Ginkel, S. Biological hydrogen production measured in batch anaerobic respirometers. Environ. Sci. Technol. 2002, 36, 2530–2535. [Google Scholar] [CrossRef] [PubMed]
- Pernthaler, J.; Glöckner, F.O.; Schönhuber, W.; Amann, R. Fluorescence in situ hybridization with rRNA-targeted oligonucleotide probes. Methods Microbiol. 2001, 30, 207–226. [Google Scholar] [CrossRef]
- Amann, R.I.; Ludwig, W.; Schleifer, K.H. Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol. Rev. 1995, 59, 143–169. [Google Scholar] [CrossRef]
- Barra Caracciolo, A.; Giuliano, G.; Grenni, P.; Cremisini, C.; Ciccoli, R.; Ubaldi, C. Effect of urea on degradation of terbuthylazine in soil. Environ. Toxicol. Chem. 2005, 24, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Barra Caracciolo, A.; Grenni, P.; Cupo, C.; Rossetti, S. In situ analysis of native microbial communities in complex samples with high particulate loads. FEMS Microbiol. Lett. 2005, 253, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Greuter, D.; Loy, A.; Horn, M.; Rattei, T. ProbeBase-an online resource for rRNA-targeted oligonucleotide probes and primers: New features 2016. Nucleic Acids Res. 2016, 44, D586–D589. [Google Scholar] [CrossRef]
- Ali, G.; Ling, Z.; Saif, I.; Usman, M.; Jalalah, M.; Harraz, F.A.; Al-Assiri, M.S.; Salama, E.S.; Li, X. Biomethanation and microbial community response during agricultural biomass and shrimp chaff digestion. Environ. Pollut. 2021, 278, 116801. [Google Scholar] [CrossRef]
- Haitjema, C.H.; Solomon, K.V.; Henske, J.K.; Theodorou, M.K.; O’Malley, M.A. Anaerobic gut fungi: Advances in isolation, culture, and cellulolytic enzyme discovery for biofuel production. Biotechnol. Bioeng. 2014, 111, 1471–1482. [Google Scholar] [CrossRef]
- Theuerl, S.; Kohrs, F.; Benndorf, D.; Maus, I.; Wibberg, D.; Schlüter, A.; Kausmann, R.; Heiermann, M.; Rapp, E.; Reichl, U.; et al. Community shifts in a well-operating agricultural biogas plant: How process variations are handled by the microbiome. Appl. Microbiol. Biotechnol. 2015, 99, 7791–7803. [Google Scholar] [CrossRef]
- Nkemka, V.N.; Gilroyed, B.; Yanke, J.; Gruninger, R.; Vedres, D.; McAllister, T.; Hao, X. Bioaugmentation with an anaerobic fungus in a two-stage process for biohydrogen and biogas production using corn silage and cattail. Bioresour. Technol. 2015, 185, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Adnan, A.I.; Ong, M.Y.; Nomanbhay, S.; Chew, K.W.; Show, P.L. Technologies for biogas upgrading to biomethane: A review. Bioengineering 2019, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.B.; Jensen, B.; Ottosen, L.D.M.; Kofoed, M.V.W. Integrating H2 injection and reactor mixing for low-cost H2 gas-liquid mass transfer in full-scale in situ biomethanation. Biochem. Eng. J. 2021, 166, 107869. [Google Scholar] [CrossRef]
- O’Flaherty, V.; Collins, G.; Mahony, T. The microbiology and biochemistry of anaerobic bioreactors with relevance to domestic sewage treatment. Rev. Environ. Sci. Biotechnol. 2006, 5, 39–55. [Google Scholar] [CrossRef]
- Krakat, N.; Westphal, A.; Schmidt, S.; Scherer, P. Anaerobic digestion of renewable biomass: Thermophilic temperature governs methanogen population dynamics. Appl. Environ. Microbiol. 2010, 76, 1842–1850. [Google Scholar] [CrossRef]
- Čater, M.; Zorec, M.; Marinšek Logar, R. Methods for Improving Anaerobic Lignocellulosic Substrates Degradation for Enhanced Biogas Production. Springer Sci. Rev. 2014, 2, 51–61. [Google Scholar] [CrossRef]
- Ali Shah, F.; Mahmood, Q.; Maroof Shah, M.; Pervez, A.; Ahmad Asad, S. Microbial ecology of anaerobic digesters: The key players of anaerobiosis. Sci. World J. 2014, 2014, 183752. [Google Scholar] [CrossRef]
- Brookman, J.L.; Mennim, G.; Trinci, A.P.J.; Theodorou, M.K.; Tuckwell, D.S. Identification and characterization of anaerobic gut fungi using molecular methodologies based on ribosomal ITS1 and 18S rRNA. Microbiology 2000, 146, 393–403. [Google Scholar] [CrossRef]
- Demirel, B.; Yenigün, O. Changes in microbial ecology in an anaerobic reactor. Bioresour. Technol. 2006, 97, 1201–1208. [Google Scholar] [CrossRef]
- De Vrieze, J.; Hennebel, T.; Boon, N.; Verstraete, W. Methanosarcina: The rediscovered methanogen for heavy duty biomethanation. Bioresour. Technol. 2012, 112, 1–9. [Google Scholar] [CrossRef]
- Smith, K.S.; Ingram-Smith, C. Methanosaeta, the forgotten methanogen? Trends Microbiol. 2007, 15, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Whitman, W.B. Metabolic, phylogenetic, and ecological diversity of the methanogenic archaea. Ann. N. Y. Acad. Sci. 2008, 1125, 171–189. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, H.; Qiu, Y.; Ren, L.; Jiang, B. Microbial characteristics in anaerobic digestion process of food waste for methane production–A review. Bioresour. Technol. 2018, 248, 29–36. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total solid (gL−1) | 87.7 ± 0.7 |
| Volatile Solid (gL−1) | 53.5 ± 0.8 |
| Organic Carbon (%) | 27.3 ± 3.0 |
| Total nitrogen (%) | 6.6 ± 0.7 |
| Microbial Sources | Acronym | AF | F210 | M | SPW g L−1 |
|---|---|---|---|---|---|
| Volume (%) | |||||
| AF + F210 +M | AF + F210_6.5 | 8 | 8 | 20 | 6.5 |
| AF + F210 +M | AF + F210_9.7 | 8 | 8 | 20 | 9.7 |
| AF + F210 +M | AF + F210_13 | 8 | 8 | 20 | 13 |
| F210 + M | F210_6.5 | - | 16 | 20 | 6.5 |
| M | M_6.5 | - | - | 20 | 6.5 |
| AF +M | AF_6.5 | 16 | - | 20 | 6.5 |
| M without SPW | I | - | - | 20 | - |
| TS Removal (%) | VS Removal (%) | |
|---|---|---|
| SPW | / | / |
| AF + F210_6.5 | 97.8 | 99.4 |
| AF + F210_9.7 | 97.4 | 99.3 |
| AF + F210_13 | 97.0 | 99.0 |
| F210 | 97.8 | 99.5 |
| M | 97.7 | 99.4 |
| AF | 97.7 | 99.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazzurco Miritana, V.; Gaetani, A.; Signorini, A.; Marone, A.; Massini, G. Bioaugmentation Strategies for Enhancing Methane Production from Shrimp Processing Waste through Anaerobic Digestion. Fermentation 2023, 9, 401. https://doi.org/10.3390/fermentation9040401
Mazzurco Miritana V, Gaetani A, Signorini A, Marone A, Massini G. Bioaugmentation Strategies for Enhancing Methane Production from Shrimp Processing Waste through Anaerobic Digestion. Fermentation. 2023; 9(4):401. https://doi.org/10.3390/fermentation9040401
Chicago/Turabian StyleMazzurco Miritana, Valentina, Alessia Gaetani, Antonella Signorini, Antonella Marone, and Giulia Massini. 2023. "Bioaugmentation Strategies for Enhancing Methane Production from Shrimp Processing Waste through Anaerobic Digestion" Fermentation 9, no. 4: 401. https://doi.org/10.3390/fermentation9040401