

Effects of Four Critical Gene Deletions in Saccharomyces cerevisiae on Fusel Alcohols during Red Wine Fermentation

1
School of Biology and Biological Engineering, South China University of Technology, Guangzhou 510006, China
2
College of Food Science, South China Agricultural University, Guangzhou 510642, China
*
Author to whom correspondence should be addressed.
Fermentation 2023, 9(4), 379; https://doi.org/10.3390/fermentation9040379
Submission received: 9 March 2023
/
Revised: 10 April 2023
/
Accepted: 13 April 2023
/
Published: 14 April 2023
(This article belongs to the Special Issue Advances in Beverages, Food, Yeast and Brewing Research 2.0)
Abstract
:Excessive fusel alcohols in red wine will bring an uncomfortable bitterness and generate an intoxicating effect, which affects the quality and attractivity of the red wine. In order to achieve better regulation of fusel alcohols in red wine, strains with LEU1 and PDC5 deletions were constructed, and seven engineered yeast strains based on THI3 and BAT2 deletions were applied to red wine fermentation to dissect the effects of four critical genes on fusel alcohols during wine fermentation. The fermentation results of these recombinant strains showed that the deletion of THI3 increased the contents of n-propanol, isobutanol, and isoamyl alcohol by 48.46%, 42.01%, and 7.84%, respectively; the deletion of BAT2 decreased isoamyl alcohol and isobutanol by 32.81% and 44.91%; the deletion of PDC5 and LEU1 decreased isoamyl alcohol by 40.21% and 68.28%, while increased isobutanol by 24.31% and 142%, respectively; the deletion of THI3 exerted a negative influence on the reduction of isoamyl alcohol caused by BAT2 or PDC5 deletion; the deletion of THI3 and PDC5 had a synergistic effect on the increase of isobutanol, while BAT2 and PDC5 deletion presented no additive property to the decrease of isoamyl alcohol. Hence, it is concluded that either BAT2, PDC5, or LEU1 deletion can effectively decrease fusel alcohols, especially isoamyl alcohol, which provides an important reference for the control of fusel alcohols in red wine.
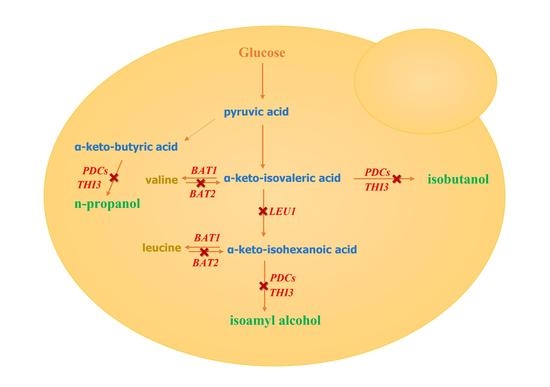
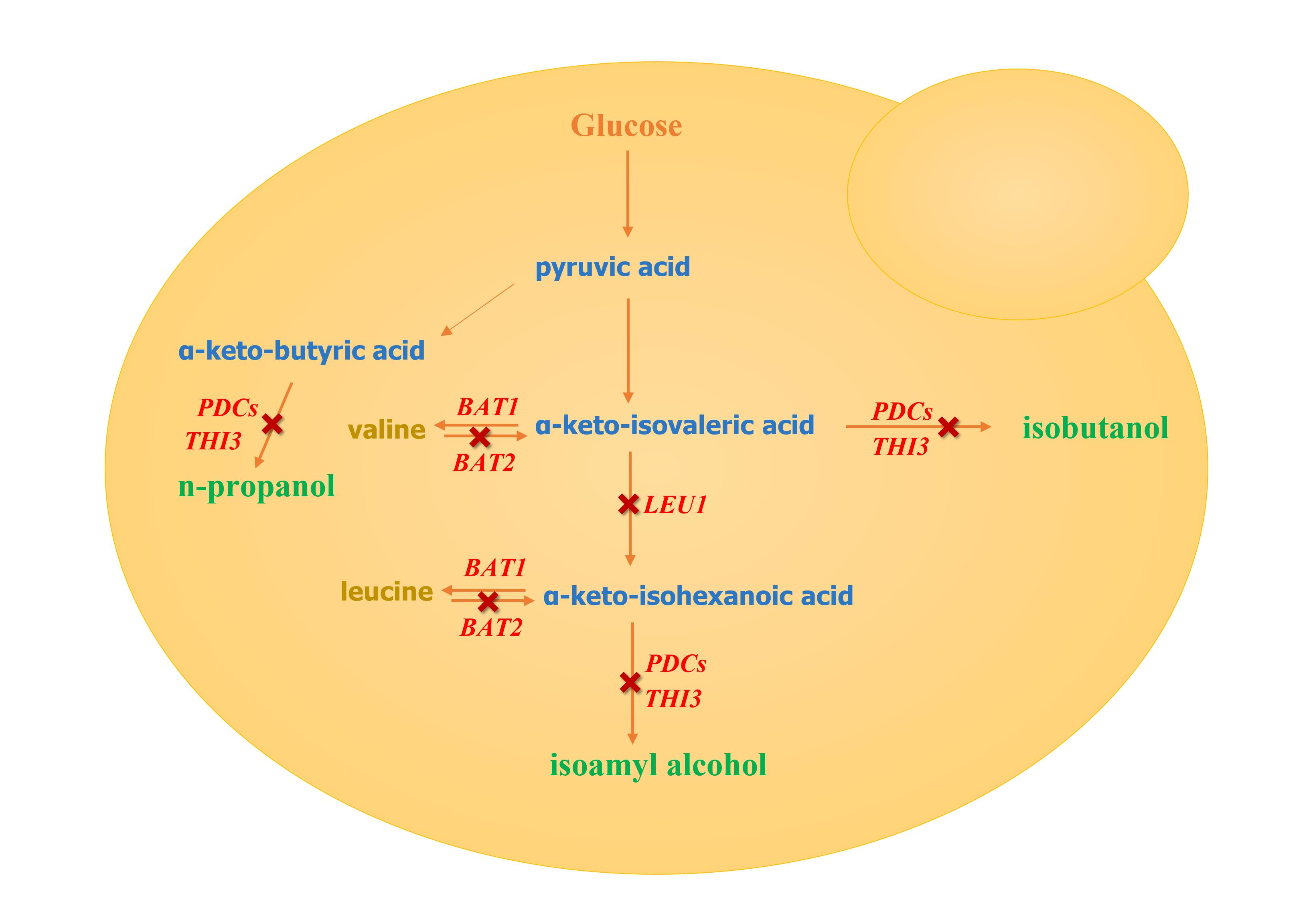
1. Introduction
Red wine is popular with all kinds of people for its attractive aroma and excellent health value [1]. Fusel alcohols are an influential factor in the mellow, plump, and coordinated fragrance of wine [2]. However, excessive fusel alcohols will also bring an uncomfortable bitterness and meanwhile generate an intoxicating effect. Among fusel alcohols, the n-propanol, isobutanol, and isoamyl alcohol are of the most concern in red wine [3]. Some studies show that the intoxication effect of these three fusel alcohols in alcoholic beverages deepens with the increase of the carbon chain [4], and the threshold of bitterness decreases successively [5,6]. However, currently, the content of isoamyl alcohol in red wine is much higher than those of the other two fusel alcohols. For example, the total fusel alcohols in Cabernet Sauvignon dry red wine were about 300–400 mg/L, and the isoamyl alcohol content accounts for about 69% of the total fusel alcohols [7]. Therefore, controlling the content of fusel alcohols, especially isoamyl alcohol, is critical for promoting the quality and attractiveness of red wine.
The fusel alcohols in red wine mainly originate from the growth and metabolism of yeast at the early stage of fermentation, generated by the Ehrlich pathway and the biosynthetic pathway [8,9]. The α-keto acids are important intermediates from amino acids and pyruvate to fusel alcohols and the intersection of the two pathways [10]. The branched-chain amino acid transferase encoded by BAT2 is in control of the conversion of amino acids to α-keto acids [11], thus being considered a rate-limiting enzyme of the Ehrlich pathway [12]. With the deletions of two BAT2 genes and the overexpression of one BAT1 gene in Saccharomyces cerevisiae, in Ma’s study, the isobutanol and isoamyl alcohol were reduced by 39.41% and 37.18%, respectively, during red wine fermentation [2]. The α-keto acids can also be produced from glucose via pyruvate [13] and α-isopropylmalate dehydrogenase encoded by LEU1 controls the conversion of α-keto-isovaleric acid to α-keto-isohexanoic acid [14]. Li constructed the recombinant yeast strain with LEU1 deletion and the isoamyl alcohol decreased by 33.74% during simulated alcoholic fermentation [15]. The decarboxylase encoded by THI3 or PDCs controls the flow of α-keto acid to fusel alcohols [16,17], however, the presence of isoenzymes makes their effect on fusel alcohols uncertain. Dickinson reported that the deletion of THI3 resulted in a 94% reduction in isoamyl alcohol when leucine was the only nitrogen source [18], while some studies have shown that THI3 deletion has no impact on the production of isoamyl alcohol [19,20]. PDC5 is a minor decarboxylase gene during glucose fermentation where the expressive intensity is approximately one-sixth that of PDC1 [21,22]. However, the effect of PDC5 deletion on isoamyl alcohol has not been investigated. In addition, the effect of the single-gene deletion LEU1 or THI3 on fusel alcohols in red wine is unknown; neither is the effect of some double gene deletions (BAT2, THI3, and PDC5) on fusel alcohols. In view of this, the genetic analysis of these genes is of great significance for the control of fusel alcohols in red wine fermentation.
In our previous studies, it was proven that the deletion of THI3 or BAT2 in S. cerevisiae can reduce fusel alcohols in grain brewing. THI3 deletion decreased isoamyl alcohol and isobutanol by 17.20% and 23.43% in wort fermentation [23]; BAT2 deletion decreased isoamyl alcohol and isobutanol by 18.12% and 35.21%, respectively in Chinese rice wine [24]. In addition, the amino nitrogen content of fermentation raw materials is also an important factor of fusel alcohols [25], and grape juice is lower in amino nitrogen than grain [26,27,28]. In this paper, we continued to delete LEU1 and PDC5 in the metabolic pathway of fusel alcohols, and the expression levels of LEU1, PDC5, and PDC1 in seven modified yeast strains, including THI3 and BAT2 deletions, were determined. Then, they were applied to red wine fermentation to dissect the effects of four critical genes on fusel alcohols, and the effects of these recombinant strains on fermentation performance and the physicochemical indices of red wine were investigated as well.
2. Materials and Methods
2.1. Strains and Media
All strains and plasmids used in this study are listed in Table S1 (Supporting Information). The E. coli DH5α with pUG6 or pSH65 was cultured in LB broth containing 100 μg/mL ampicillin under 37 °C and at 200 rpm. The S. cerevisiae strains were cultivated at 30 °C in a YPD medium, and 200 μg/mL of G418 (Geneticin, Shanghai, China) and 300 μg/mL of Zeocin (Promega, Carlsbad, CA, USA) were used to screen positive yeast transformants with KanMX and Zeocin resistance, respectively. The expression of Cre in yeast transformants was induced by the YPG medium. The hybridized diploid strains were verified using the MacConkey medium. All media formulations were derived from the previous report [23].
2.2. Design and Synthesis of Primers
According to the sequences of LEU1 on genomes of S. cerevisiae S288c published by NCBI, primer pairs LEU1-F1/R1 and LEU1-F2/R2 were designed to amplify the left and right homology arms, respectively. The primer pairs L-loxP-F/R were designed to amplify loxP-KanMX-loxP from the plasmid pUG6. The partially overlapping bases were introduced to the 5′ of primer pairs for the overlap extension PCR to obtain the gene disruption cassette, and the positive clones were identified by primers LEU1-A/D. The primer design of the PDC5 disruption cassette was similar to LEU1. All the primer pairs used in this study were synthesized by Shanghai Biological Engineering Technology Co., Ltd. (Shanghai, China)., as listed in Table S2 (Supporting Information).
2.3. Construction of the Disruption Cassette
The LEU1 disruption cassette was synthesized by the overlap extension PCR [29]. First, the left homology arm (240 bp) and the right homology arm (334 bp) were synthesized from the genome of the wild-type yeast strain XF1 using the primer pairs LEU1-F1/R1 and LEU1-F2/R2. Then, the G418 resistant marker loxP-KanMX-loxP (1738 bp) was amplified by primer pairs L-loxP-F/R, using pUG6 plasmid as the template. Finally, the left homology arm and the right homology arm were connected with loxP-KanMX-loxP by the overlap extension PCR to construct the disruption cassette (2262 bp). Similarly, the PDC5 disruption cassette (2034 bp) was constructed through the same process according to the primers shown in Table S2 (Supporting Information).
2.4. Yeast Transformation and Screening
First, the LEU1 and PDC5 disruption cassettes were transferred into the haploid S. cerevisiae XFa7 and XFα6, respectively by electrotransformation [30], and transformed yeast was plated on YPD (200 μg/mL G418) and incubated at 30 °C for 48 h. Then, the positive yeast transformants were selected using the colony PCR [31] with the primer pairs LEU1-A/D, and PDC5-A/D, respectively. In addition, using the electrotransformation to remove the selected marker KanMX [32], the pSH65 plasmid was transformed into haploid yeast strains with LEU1 and PDC5 deletions, and finally, the recombinant yeast strains XFa7-P/XFα6-P and XFa7-L/XFα6-L were obtained. Similarly, the PDC5 disruption cassette was transferred into the haploid yeast strains XFa7-T/XFα6-T and XFa7-B/XFα6-B to construct XFa7-TP/XFα6-TP and XFa7-BP/XFα6-BP, respectively.
2.5. Construction of the Diploid S. cerevisiae
The diploid S. cerevisiae was constructed through the hybridization of α-type and a-type haploid recombinant yeasts. First, 100 μL of α-type and 100 μL of a-type haploid recombinant yeasts were pipetted into 1 mL of YPD liquid medium, and the mixed solution was shaken and then cultured at 30 °C and 100 rpm for 16 h. When a dumbbell-shaped yeast fusion morphology was observed, the culture was plated on YPD and incubated there at 30 °C for 48 h to select the diploid yeast strains. After that, the diploid strains were verified on the MacConkey plate to observe the sporulation production through microscope (Murzider, Dongguan, China) examination.
2.6. Real-Time Quantitative PCR
RNA was extracted using the Yeast Total RNA Extraction Kit (Sangon, Guangzhou, China), and then reversely transcribed to cDNA by Evo M-MLV RT Premix (Accyrate, Changsha, China). The cDNA was used for Real-Time Quantitative PCR by SYBR Green Premix Pro Taq HS qPCR Kit (Accyrate, Changsha, China), and the expression levels of LEU1, PDC5, and PDC1 of different strains were calculated using the 2−ΔΔCt method with ACT1 as a reference gene. The primers are listed in Table S2 (Supporting Information).
2.7. Fermentation Analysis of Recombinant Yeasts in Red Wine Fermentation
The Cabernet Sauvignon grapes (Qingdao, Shandong, China) were harvested in September 2021, with the sugar content being 20° Brix and a pH being 3.28. The grapes were rinsed with water and destemmed, and then crushed using a screw press (Joyoung, Guangzhou, China). The grape mash was macerated for 16 h at 12 °C and then the pomace was discarded to obtain the juice. After that, the grape juice was divided into small portions, with 50 mL for each portion poured into 100 mL conical flasks, and then sealed using rubber bands and plastic bags. The yeast strains were precultured in 10 mL of YPD media at 30 °C and 200 rpm for 12 h, and then 1 mL of yeast cells was inoculated into 20 mL of the YPD media for synchronous culture for 16 h. Finally, 2 mL of yeast cells (6 × 106 CFU/mL) were added to the grape juice and fermented at 25 °C for 4 days. When the fermentation was completed, the yeast sludge was discarded after standing and clarification, and then the obtained red wines were stored at 4 °C. The weight loss of CO2 was measured by an analytical balance (Mettler-Toledo, Columbus, OH, USA) every 12 h, and the fusel alcohol content and physicochemical indices of red wines were determined after the fermentation.
2.8. Analytical Methods
The contents of fusel alcohols and ethanol in the red wine were measured by the internal standard method using a modified gas chromatography (GC, Jingwei-Analytical, Tengzhou, China) process based on the previous report [23]. N-Butyl acetate was used as the internal standard for the fusel alcohols. The red wine was collected and centrifuged at 10,000× g for 3 min, and then the supernatant was filtered through a 0.22 μm syringe filter. The assay sample consisted of 980 μL of wine and 20 μL of N-Butyl acetate (17.66 mg/mL). Nitrogen was used as the carrier gas with a flow rate of 1.5 mL min−1. The temperature of the FID and injector were 230 °C and 220 °C, respectively, and the injection volume was 0.4 μL. The column temperature started to increase from 60 °C, with 60 °C to 120 °C at the rate of 10 °C min−1, and 120 °C to 200 °C at the rate of 30 °C min−1, and it was kept at 200 °C for 3 min. In addition, the ethanol was determined with acetone as the internal standard, and the assay sample consisted of 800 μL of H2O, 100 μL of wine, and 20 μL of acetone (7.32 mg/mL). The column temperature was first kept at 50 °C for 4 min, then rose from 50 °C to 200 °C at the rate of 30 °C min−1, and finally held at 200 °C for 3 min. As for other physicochemical indices of the red wine, the pH was measured by a pH meter (Mettler-Toledo, Columbus, OH, USA); the residual sugar was quantified by the 3,5-Dinitrosalicylic acid (DNS) method; the total acid of the red wine was determined with NaOH (0.05 mol/L) by an automatic potentiometric titrator (Rex, Columbus, OH, USA).
2.9. Statistical Analysis
All experiments were performed in three biological replicates and the results were reported as mean ± standard deviation (SD). The significance of the differences was analyzed by Student’s t-test, and the differences were considered statistically significant at p < 0.05.
3. Results
3.1. Construction of Recombinant Yeast Strains
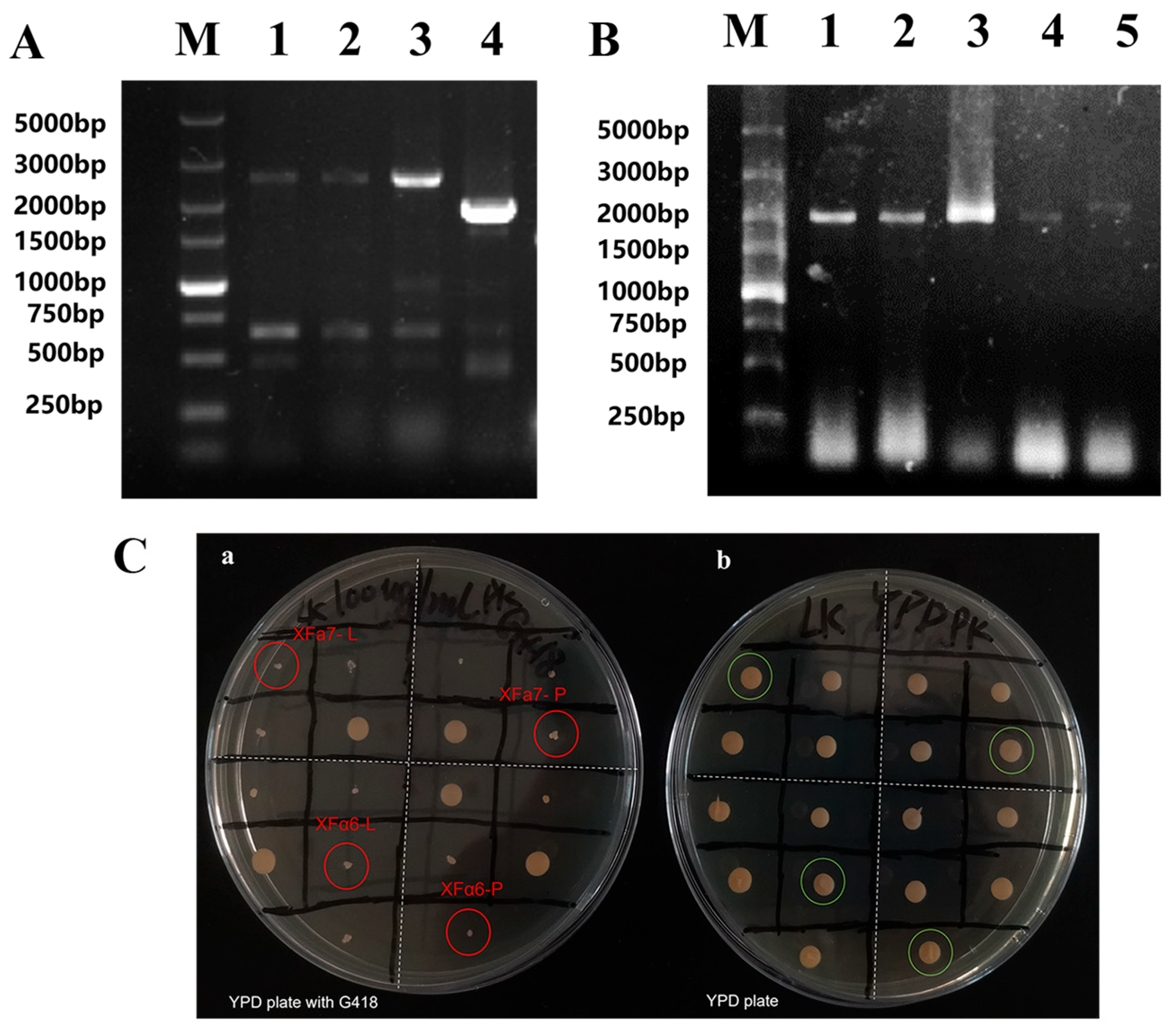
The construction results of the haploid recombinant yeast strains are revealed in Figure 1. The LEU1 and PDC5 genes of the haploid strains XFa7 and XFα6 were replaced by the corresponding gene disruption cassettes, respectively. The bands with 2505 bp were obtained from the LEU1 deletion haploid yeast by primer pairs LEU1-A/D, while the band with 1891 bp was obtained from the wild-type strain XF1 (Figure 1A). Similarly, the bands with 2092 bp were obtained from the PDC5 deletion haploid yeast by primer pairs PDC5-A/D, while the band with 2345 bp was obtained from XF1 (Figure 1B). Moreover, the replica plates showed that the resistance markers of the haploid recombinant strains were successfully removed (Figure 1C). These data indicate the successful construction of the haploid yeast strains with the LEU1 and PDC5 deletions, respectively. The results of hybridization and the sporulation verification of haploid recombinant strains are shown in Figure S1 (Supporting Information). Based on the haploid strains with BAT2 or THI3 deletion, double gene-deletion strains XF-BP and XF-TP were successfully constructed with the same process as XF-P.
3.2. Relative Expression Levels of LEU1, PDC5, and PDC1
To further confirm whether LEU1 and PDC5 are expressed after deletion and the expression of the major decarboxylase gene PDC1 in the different deletion strains. The relative expression levels of LEU1, PDC5, and PDC1 in seven recombinant strains were measured and shown in Figure S2 (Supporting Information). It can be observed that no expression of the corresponding gene was detected in either the PDC5 or LEU1 deletion strains, and all the deletions of THI3, PDC5, and BAT2 caused a downregulation expression of LEU1. In addition, it was found that PDC5 or LEU1 deletion did not affect the expression level of PDC1, while THI3 deletion significantly enhanced the expression levels of PDC5 and PDC1, by 4.19 fold and 9.95 fold, respectively.
3.3. Comparison of Fermentation Performance of Different Recombinant Strains
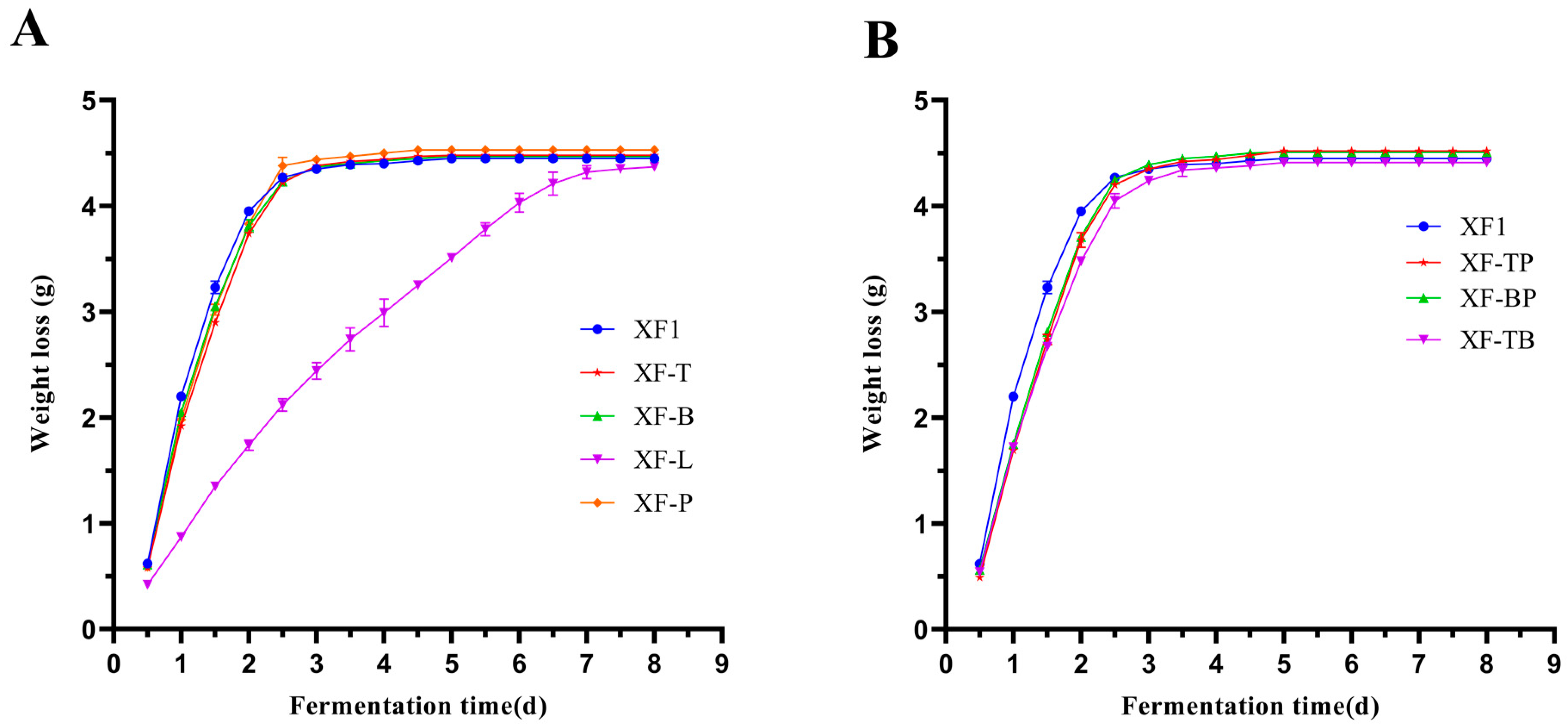
The wild-type strain XF1 and seven recombinant strains were inoculated in grape juice for fermentation. The weight loss of CO2 can reflect the fermentation rates of the yeast. Therefore, the accumulation of CO2 was determined every 12 h and shown in Figure 2. The fermentation rates of XF-T, XF-B, and XF-P were similar and slightly lower than that of XF1 (Figure 2A). It was worth mentioning that the fermentation rate of XF-L was remarkably affected, and the fermentation period was extended to 7 d. However, the final CO2 production of different recombinant strains was similar to that of XF1, indicating that these gene deletions had little influence on the fermentation degree.
The CO2 productions of the double gene-deletion strains are shown in Figure 2B. It can be observed that the fermentation rates of XF-BP and XF-TP were similar, slightly lower than those of the single gene-deletion strains. The XF-TB had a relatively poor fermentation rate, suggesting that the deletion of THI3 and BAT2 slightly weakened the fermentation performance of the yeast. The CO2 production of the recombinant strains was almost the same as that of XF1 after 72 h, indicating that most of these gene deletions had little influence on the fermentation period.
After fermentation, the alcohol content, pH, residual sugar, and total acid were determined. As shown in Table 1, all these data of the red wine fermented by the recombinant strains were similar to those by XF1, indicating that these gene deletions had no impact on the physicochemical indices of the red wine.
3.4. Effects of BAT2, THI3, and PDC5 Deletion on the Production of Fusel Alcohols
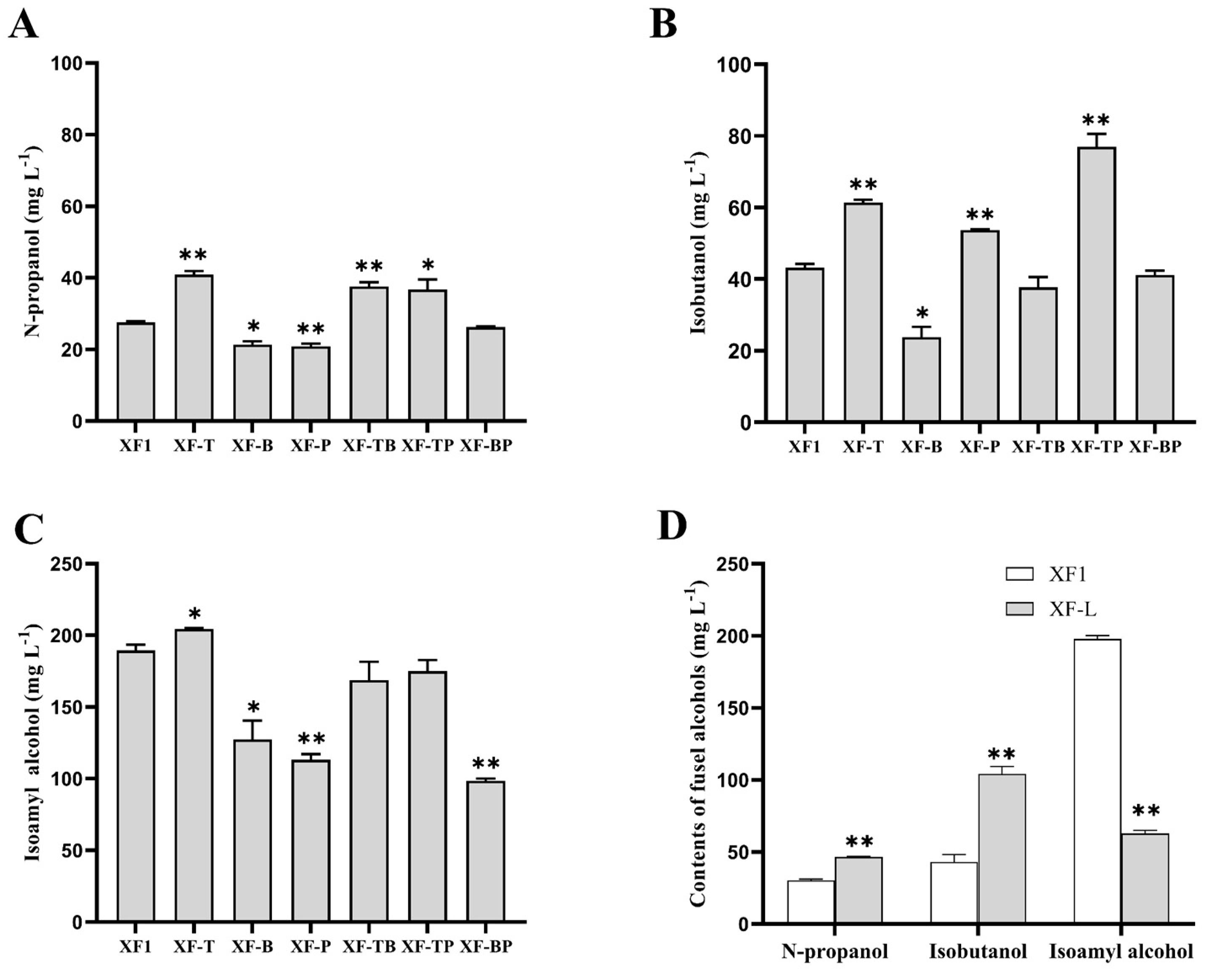
The contents of n-propanol, isobutanol, and isoamyl alcohol in the recombinant strains and the wild-type strain XF1 were detected to investigate the effects of BAT2, THI3, and PDC5 deletions on the production of fusel alcohols and analyze the interactions among these genes. Between the recombinant strains and XF1, remarkable differences were noticed in the fusel alcohol contents (Figure 3).
As shown in Figure 3A, the content of n-propanol in the single gene-deletion strain XF-T increased by 48.46% to 40.9 mg/L compared with that of the wild-type strain XF1. Moreover, the double gene-deletion strains XF-TB and XF-TP also increased by 36.30% and 33.21%, respectively in n-propanol content with THI3 deletion. This implies that THI3 deletion had a positive effect on the increase of n-propanol. The n-propanol content of strains XF-B and XF-P decreased by 22.32% and 24.31% (i.e., 21.8 mg/L and 20.4 mg/L), respectively, compared to XF1, while the n-propanol content of the double gene-deletion strain XF-BP appeared to be the same as that of XF1.
The isobutanol contents of strains with different gene deletions are shown in Figure 3B. The isobutanol contents of strains XF-T and XF-P increased by 42.01% and 24.31% to 61.35 mg/L and 53.7 mg/L, respectively, compared to that of XF1, and the double gene-deletion strain XF-TP even increased by 78.13% (i.e., 76.95 mg/L), indicating that the deletions of THI3 and PDC5 have a synergistic effect on the increase in isobutanol. The isobutanol contents of strains XF-B decreased by 44.91% (i.e., 23.8 mg/L), while the double gene-deletion strain XF-TB and XF-BP showed no significant difference from XF1 in isobutanol content. It suggests that the increase in isobutanol caused by THI3 or PDC5 deletion can be eliminated by BAT2 deletion.
The isoamyl alcohol content of strain XF-T slightly increased by 7.84% to 204.35 mg/L compared to the wild-type strain XF1 (Figure 3C), while those of the strains XF-B and XF-P significantly decreased by 32.80% (i.e., 127.35 mg/L) and 40.21% (i.e., 113.30 mg/L), respectively. However, the content of isoamyl alcohol in the double gene-deletion strains XF-TB and XF-TP showed no significant difference from that of strain XF1, indicating that the deletion of THI3 had a negative influence on the reduction of isoamyl alcohol caused by the deletion of the other two genes. Meanwhile, the double gene-deletion strain XF-BP decreased the isoamyl alcohol by 47.97% (i.e., 87.6 mg/L), illustrating that BAT2 and PDC5 deletions had no additive effect to the decrease of isoamyl alcohol.
3.5. Effect of LEU1 Deletion on the Production of Fusel Alcohols
The effect of LEU1 deletion on the production of fusel alcohols was investigated individually in red wine fermentation due to the extension of the fermentation period. The data on fermentation samples analyzed by GC are presented in Figure 3D. It can be seen that the production of isoamyl alcohol by recombinant XF-L declined to 62.75 mg/L, which is a 68.28% decrease compared with that of the wild-type strain XF1. Moreover, the contents of n-propanol and isobutanol produced by XF-L were 46.5 mg/L and 104.2 mg/L, respectively, approximately 53.21% and 142% higher than those produced by XF1.
3.6. Comparison of Total Fusel Alcohols of Different Recombinant Strains
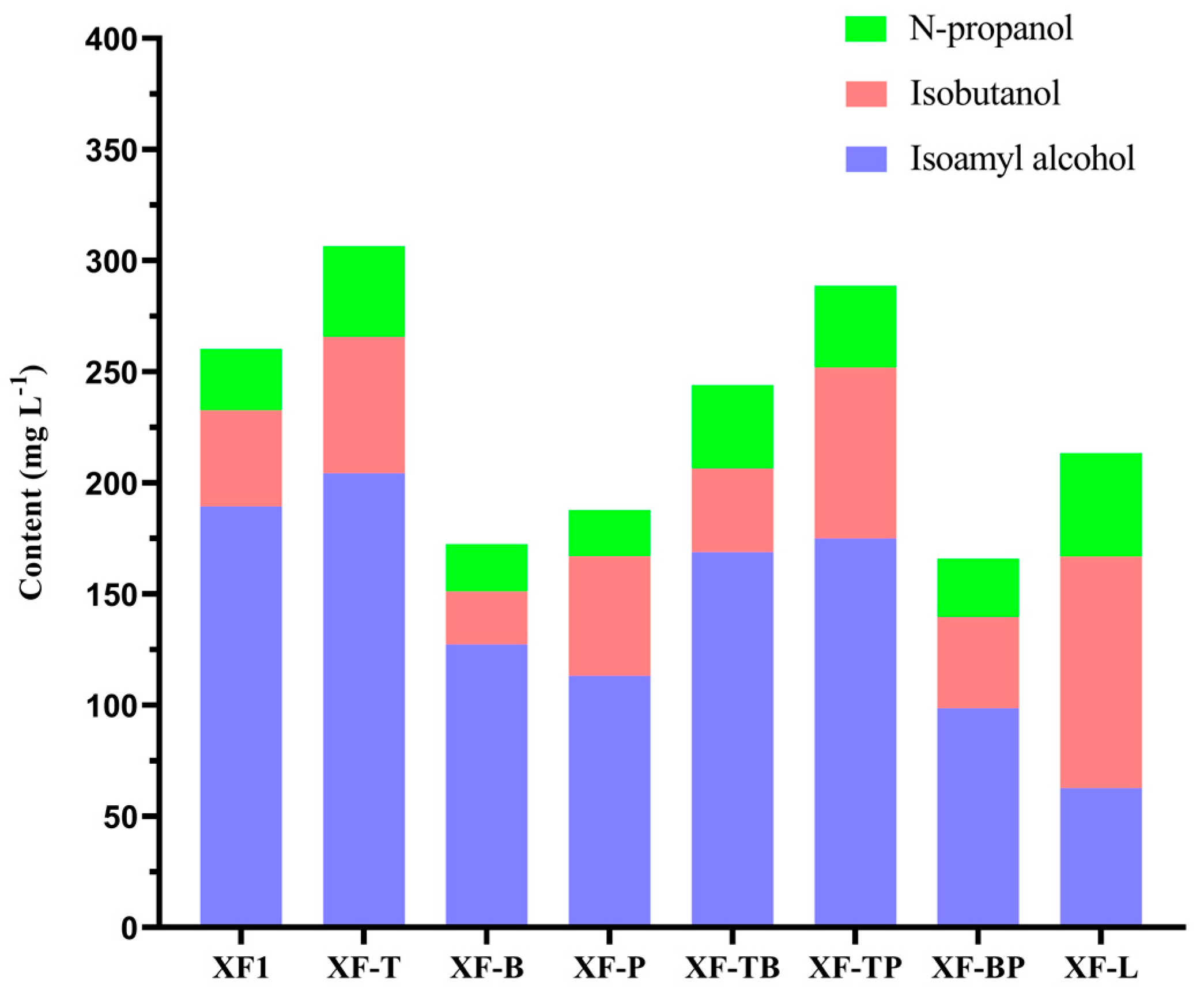
The total fusel alcohol content was defined by the sum of n-propanol, isobutanol, and isoamyl alcohol. Figure 4 shows the fusel alcohol content of seven recombinant yeast strains and the wild-type strain XF1 in the form of an accumulative bar diagram. It can be observed that the total fusel alcohol content of the XF1 strain was 260.25 mg/L; the total fusel alcohol contents of XF-T and XF-TP increased by 17.81% and 10.91%, respectively; the total fusel alcohol contents of XF-B, XF-P, and XF-BP decreased by 33.70%, 27.82%, and 36.23%, respectively. In terms of the composition of the fusel alcohols, XF-B had a good reduction effect on isobutanol and isoamyl alcohol, indicating that BAT2 deletion possesses a high application value in red wine industrial production. The strains XF-P and XF-L regulated isoamyl alcohol to isobutanol at different levels, suggesting that PDC5 or LEU1 deletion can also have a positive effect on the decrease in bitterness and intoxicating effect. The deletions of BAT2 and PDC5 could slightly neutralize the difference among the constituents of fusel alcohol based on further decreasing the total fusel alcohol content. In this study, different gene deletion strategies led to various changes in fusel alcohols, which provides a significant reference for the control of fusel alcohols in red wine production.
4. Discussion
The fusel alcohols produced by yeast have an essential effect on the flavor and taste of red wine [33], and the genes THI3, BAT2, PDC5, and LEU1 play a critical role in different nodes on the metabolic pathway of fusel alcohols [34]. Therefore, the genetic analysis of these genes is of great significance for controlling the content of fusel alcohols in red wine. This study, for the first time, showed that the deletion of THI3 could increase n-propanol, isobutanol, and isoamyl alcohol in red wine fermentation. The possible reason might be that THI3 deletion enhanced the expression levels of PDC1 and PDC5 [35], which also played a role in the transformation of α-keto acid to fusel alcohols. However, some studies have shown that THI3 deletion can reduce fusel alcohols in low glucose and high amino nitrogen conditions. It can be inferred that the pyruvate decarboxylase-like enzyme encoded by THI3 tends to use the α-keto acid derived from the Ehrlich pathway as the substrate [12,18,36].
The branched-chain amino acid transferase encoded by BAT2 is a rate-limiting enzyme in the Ehrlich pathway. In this study, the deletion of BAT2 caused a significant decrease in isobutanol and isoamyl alcohol, which is consistent with previous studies [2,37]. When BAT2 and THI3 were deleted, the isoamyl alcohol content was similar to that of the wild-type strain. However, the decreased effect on isoamyl alcohol of BAT2 deletion was more significant than the increased effect of THI3 deletion. We speculated that double gene deletions might break the balance of leucine conversion with α-keto-isohexanoic acid, leading to a slight decrease in BAT1 expression [11]. Therefore, part of the α-keto acids that should be converted into amino acids was diverted to isoamyl alcohol.
It was also found for the first time that PDC5 deletion led to a significant decrease in isoamyl alcohol and an increase in isobutanol, indicating that PDC5 mainly controlled the decarboxylation of α-keto-isohexanoic acid to isoamyl alcohol. The deletion of THI3 and PDC5 synergically increased the isobutanol content, while isoamyl alcohol showed no significant change, which might be due to the different substrate specificity of the two decarboxylases: the decarboxylase encoded by PDC1 tends to produce isobutanol while that encoded by PDC5 is more likely to produce isoamyl alcohol. In addition, the deletion of either BAT2 or PDC5 could decrease isoamyl alcohol, though the deletions of BAT2 and PDC5 showed no additive effect on the reduction of isoamyl alcohol. This is due to the pyruvate decarboxylase encoded by PDC5 also using α-keto-isohexanoic acid from the amino acid metabolic pathway as a substrate [16].
The deletion of LEU1 interfered with the contribution of the biosynthetic pathway to isoamyl alcohol and turned the original strain into a leucine-nutrient-deficient strain [38], thus resulting in a significant decrease in isoamyl alcohol, which directly confirmed that 75% of fusel alcohols came from the biosynthetic pathway in alcoholic fermentation [39]. At the same time, the fermentation performance of the strain was severely affected as a result of the blocking of the synthesis pathway of leucine, though this effect was only reflected in the extension of the fermentation period. Similarly, in Li’s study, the growth performance of the LEU1 deletion strain was also affected, and the decreased effect of isoamyl alcohol in the simulated alcohol fermentation was lower than that in red wine fermentation due to the rich amino nitrogen [15].
5. Conclusions
With the appropriate contents of fusel alcohols, the coordinated aroma and mellow taste can be achieved in red wine, while the excessive contents will bring an uncomfortable bitterness and an intoxicating effect. The isoamyl alcohol is of the highest content among the fusel alcohols and it contributes the most to the bitterness and intoxicating effect in red wine. Based on the results of this study, either BAT2, PDC5, or LEU1 deletion can effectively decrease isoamyl alcohol. Notably, LEU1 deletion can sharply decrease isoamyl alcohol, with little change in total fusel alcohol. Hence, it might be able to reduce the bitterness and the intoxicating effect without affecting the flavor and taste of red wine. However, as the safety of genetically modified organisms for food and the environment is uncertain, these recombinant strains can only be applied in a few licensed countries and regions. Despite the limitation, this study still provides a worthy insight for the regulation of fusel alcohols.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fermentation9040379/s1, Figure S1: Hybridization acquisition and verification of recombinant diploid strain; Figure S2: Relative expression level of LEU1, PDC5, and PDC1 in different recombinant strains. Table S1: Strains and plasmids used in this study; Table S2: List of the Primer sequences used in the current study.
Author Contributions
Conceptualization, T.Y.; methodology, T.Y. and Z.W.; software, T.Y.; validation, T.Y., Z.W. and J.H.; formal analysis, T.Y.; investigation, T.Y. and Z.W.; resources, T.Y. and Z.W.; data curation, T.Y. and H.Z.; writing—original draft preparation, T.Y.; writing—review and editing, T.Y.; H.Z., J.H. and S.Z.; visualization, T.Y.; supervision, S.Z.; project administration, S.Z.; funding acquisition, S.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (grant number 52070079).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chang, K.J.; Liz Thach, M.; Olsen, J. Wine and health perceptions: Exploring the impact of gender, age and ethnicity on consumer perceptions of wine and health. Wine Econ. Policy 2016, 5, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Huang, S.; Du, L.; Tang, P.; Xiao, D. Reduced Production of Higher Alcohols by Saccharomyces cerevisiae in Red Wine Fermentation by Simultaneously Overexpressing BAT1 and Deleting BAT2. J. Agric. Food Chem. 2017, 65, 6936–6942. [Google Scholar] [CrossRef] [PubMed]
- Connell, D.W.; Strauss, C.R. Major constituents of fusel oils distilled from Australian grape wines. J. Sci. Food Agric. 1974, 25, 31–44. [Google Scholar] [CrossRef]
- Xie, J.; Tian, X.; He, S.; Wei, Y.; Peng, B.; Wu, Z. Evaluating the Intoxicating Degree of Liquor Products with Combinations of Fusel Alcohols, Acids, and Esters. Molecules 2018, 23, 1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.; Zeng, X. Effect of activated carbon on bitterness substances of litchi spirit. Sci. Technol. Food Ind. 2012, 4, 291–293. [Google Scholar]
- Luo, Y.; Kong, L.; Xue, R.; Wang, W.; Xia, X. Bitterness in alcoholic beverages: The profiles of perception, constituents, and contributors. Trends Food Sci. Technol. 2020, 96, 222–232. [Google Scholar] [CrossRef]
- Qi, X.; Shi, X.; Wang, J.; Ma, Y. Determination of Methanol, Ethyl Acetate and Higher Alcohols Content in Cabernet Sauvignon Dry Red Wine. Liquor-Mak. Sci. Technol. 2018, 3, 98–101. [Google Scholar] [CrossRef]
- Eden, A.; Van Nedervelde, L.; Drukker, M.; Benvenisty, N.; Debourg, A. Involvement of branched-chain amino acid aminotransferases in the production of fusel alcohols during fermentation in yeast. Appl. Microbiol. Biotechnol. 2001, 55, 296–300. [Google Scholar] [CrossRef]
- Hazelwood, L.A.; Daran, J.M.; van Maris, A.J.A.; Pronk, J.T.; Dickinson, J.R. The Ehrlich Pathway for Fusel Alcohol Production: A Century of Research on Saccharomyces cerevisiae Metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Liu, L.; Wang, Y.; Wang, X.; Xiao, D. Higher alcohols metabolism by Saccharomyces cerevisiae: A mini review. Sheng Wu Gong Cheng Xue Bao 2021, 37, 429–447. [Google Scholar] [CrossRef]
- Colón, M.; Hernández, F.; López, K.; Quezada, H.; González, J.; López, G.; Aranda, C.; González, A. Saccharomyces cerevisiae Bat1 and Bat2 Aminotransferases Have Functionally Diverged from the Ancestral-Like Kluyveromyces lactis Orthologous Enzyme. PLoS ONE 2011, 6, e16099. [Google Scholar] [CrossRef] [Green Version]
- Styger, G.; Jacobson, D.; Prior, B.A.; Bauer, F.F. Genetic analysis of the metabolic pathways responsible for aroma metabolite production by Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2013, 97, 4429–4442. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, J.R.; Norte, V. A study of branched-chain amino acid aminotransferase and isolation of mutations affecting the catabolism of branched-chain amino acids in Saccharomyces cerevisiae. FEBS Lett. 1993, 326, 29–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skala, J.; Capieaux, E.; Balzi, E.; Chen, W.N.; Goffeau, A. Complete sequence of the Saccharomyces cerevisiae LEU1 gene encoding isopropylmalate isomerase. Yeast 1991, 7, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, S.; Wang, J.; Zhang, C.; Shi, Y.; Guo, X.; Chen, Y.; Xiao, D. Genetic engineering to alter carbon flux for various higher alcohol productions by Saccharomyces cerevisiae for Chinese Baijiu fermentation. Appl. Microbiol. Biotechnol. 2018, 102, 1783–1795. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, J.R.; Harrison, S.J.; Hewlins, M.J. An investigation of the metabolism of valine to isobutyl alcohol in Saccharomyces cerevisiae. J. Biol. Chem. 1998, 273, 25751–25756. [Google Scholar] [CrossRef] [Green Version]
- Romagnoli, G.; Luttik, M.A.H.; Kötter, P.; Pronk, J.T.; Daran, J.M. Substrate specificity of thiamine pyrophosphate-dependent 2-oxo-acid decarboxylases in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2012, 78, 7538–7548. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, J.R.; Salgado, L.E.J.; Hewlins, M.J.E. The Catabolism of Amino Acids to Long Chain and Complex Alcohols in Saccharomyces cerevisiae. J. Biol. Chem. 2003, 278, 8028–8034. [Google Scholar] [CrossRef] [Green Version]
- Hao, X.; Xiao, D.; Zhang, C. Effect of YDL080C gene deletion on higher alcohols production in Saccharomyces cerevisiae haploids. Wei Sheng Wu Xue Bao = Acta Microbiol. Sin. 2010, 50, 1030–1035. [Google Scholar]
- Li, T.; Sun, J.; Wu, D.; Li, X.M.; Xie, G.F.; Lu, J. Effect of YDL080C and LEU2 gene knockout on isoamyl alcohol production in industrial yellow rice wine yeast. Sci. Technol. Food Ind. 2015, 36, 1891193. [Google Scholar]
- Hohmann, S.; Cederberg, H. Autoregulation may control the expression of yeast pyruvate decarboxylase structural genes PDC1 and PDC5. Eur. J. Biochem. 1990, 188, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, S. PDC6, a weakly expressed pyruvate decarboxylase gene from yeast, is activated when fused spontaneously under the control of the PDC1 promoter. Curr. Genet. 1991, 20, 373–378. [Google Scholar] [CrossRef]
- Wu, L.; Wen, Y.; Chen, W.; Yan, T.; Tian, X.; Zhou, S. Simultaneously deleting ADH2 and THI3 genes of Saccharomyces cerevisiae for reducing the yield of acetaldehyde and fusel alcohols. FEMS Microbiol. Lett. 2021, 368, fnab094. [Google Scholar] [CrossRef]
- Chen, W.; Zhou, S.; Yan, T.; Liang, S. Construction of THI3/BAT2 Gene-Deleted Saccharomyces cerevisiae and Its Application in Preparing Chinese Rice Wine. Mod. Food Sci. Technol. 2022, 11, 55–62. [Google Scholar]
- Wang, Y.; Sun, Z.; Zhang, C.; Zhang, Q.; Guo, X.; Xiao, D. Comparative transcriptome analysis reveals the key regulatory genes for higher alcohol formation by yeast at different α-amino nitrogen concentrations. Food Microbiol. 2021, 95, 103713. [Google Scholar] [CrossRef] [PubMed]
- Mataffo, A.; Scognamiglio, P.; Dente, A.; Strollo, D.; Colla, G.; Rouphael, Y.; Basile, B. Foliar Application of an Amino Acid-Enriched Urea Fertilizer on ‘Greco’ Grapevines at Full Veraison Increases Berry Yeast-Assimilable Nitrogen Content. Plants 2020, 9, 619. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.L.; Budde, A.D. Effect of Reducing and Oxidizing Agents and pH on Malt Endoproteolytic Activities and Brewing Mashes. J. Agric. Food Chem. 2003, 51, 7504–7512. [Google Scholar] [CrossRef]
- Okuda, M.; Miyamoto, M.; Joyo, M.; Takahashi, K.; Goto-Yamamoto, N.; Iida, S.; Ishii, T. The relationship between rice protein composition and nitrogen compounds in sake. J. Biosci. Bioeng. 2016, 122, 70–78. [Google Scholar] [CrossRef]
- González, C.; Perdomo, G.; Tejera, P.; Brito, N.; Siverio, J.M. One-step, PCR-mediated, gene disruption in the yeast Hansenula polymorpha. Yeast 1999, 15, 1323–1329. [Google Scholar] [CrossRef]
- Meilhoc, E.; Teissie, J. Electrotransformation of Saccharomyces cerevisiae. In Electroporation Protocols: Microorganism, Mammalian System, and Nanodevice; Li, S., Chang, L., Teissie, J., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2020; pp. 187–193. [Google Scholar]
- Azevedo, F.; Pereira, H.; Johansson, B. Colony PCR. In PCR: Methods and Protocols; Domingues, L., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2017; pp. 129–139. [Google Scholar]
- Güldener, U.; Heck, S.; Fiedler, T.; Beinhauer, J.; Hegemann, J.H. A New Efficient Gene Disruption Cassette for Repeated Use in Budding Yeast. Nucleic Acids Res. 1996, 24, 2519–2524. [Google Scholar] [CrossRef] [Green Version]
- de-la-Fuente-Blanco, A.; Sáenz-Navajas, M.P.; Ferreira, V. Levels of higher alcohols inducing aroma changes and modulating experts’ preferences in wine model solutions. Aust. J. Grape Wine Res. 2017, 23, 162–169. [Google Scholar] [CrossRef]
- Sun, Z.; Zhou, B.; Wang, M.; Wang, Y.; Xing, S.; Guo, X.; Xiao, D. Construction of industrial brewing yeast for fermentation under high temperature and high gravity condition. Sheng Wu Gong Cheng Xue Bao 2019, 35, 522–534. [Google Scholar] [PubMed]
- Brion, C.; Ambroset, C.; Delobel, P.; Sanchez, I.; Blondin, B. Deciphering regulatory variation of THI genes in alcoholic fermentation indicate an impact of Thi3p on PDC1 expression. BMC Genom. 2014, 15, 1085. [Google Scholar] [CrossRef] [Green Version]
- Styger, G.; Jacobson, D.; Bauer, F.F. Identifying genes that impact on aroma profiles produced by Saccharomyces cerevisiae and the production of higher alcohols. Appl. Microbiol. Biotechnol. 2011, 91, 713–730. [Google Scholar] [CrossRef] [Green Version]
- Lilly, M.; Bauer, F.F.; Styger, G.; Lambrechts, M.G.; Pretorius, I.S. The effect of increased branched-chain amino acid transaminase activity in yeast on the production of higher alcohols and on the flavour profiles of wine and distillates. FEMS Yeast Res. 2006, 6, 726–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, Y.P.; Schimmel, P. Yeast LEU1. Repression of mRNA levels by leucine and relationship of 5′-noncoding region to that of LEU2. J. Biol. Chem. 1984, 259, 3714–3719. [Google Scholar] [CrossRef]
- Mangas, J.J.; Cabranes, C.; Moreno, J.; Gomis, D.B. Influence of Cider-Making Technology on Cider Taste. LWT Food Sci. Technol. 1994, 27, 583–586. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Construction results of haploid recombinant yeast strains. (A) PCR verification of recombinant yeasts with LEU1 deletion. M, DL 5000 DNA Marker; lanes 1−3, recombinant strain bands: 2505 bp; lanes 4, wild-type strain band: 1891 bp. (B) PCR verification of recombinant yeasts with PDC5 deletion. Lanes 1−4, recombinant strain bands 2092 bp. lanes 5, wild-type strain band: 2345 bp. (C) Verification of the removal of resistance markers in haploid recombinant strains.
Figure 1.
Construction results of haploid recombinant yeast strains. (A) PCR verification of recombinant yeasts with LEU1 deletion. M, DL 5000 DNA Marker; lanes 1−3, recombinant strain bands: 2505 bp; lanes 4, wild-type strain band: 1891 bp. (B) PCR verification of recombinant yeasts with PDC5 deletion. Lanes 1−4, recombinant strain bands 2092 bp. lanes 5, wild-type strain band: 2345 bp. (C) Verification of the removal of resistance markers in haploid recombinant strains.

Figure 2.
Comparison of fermentation rates of different recombinant strains and the wild-type strain XF1. (A) The CO2 production of single gene-deletion strains and XF1. (B) The CO2 production of double gene-deletion strains and XF1.
Figure 2.
Comparison of fermentation rates of different recombinant strains and the wild-type strain XF1. (A) The CO2 production of single gene-deletion strains and XF1. (B) The CO2 production of double gene-deletion strains and XF1.

Figure 3.
Contents of fusel alcohols produced by the recombinant strains and the wild-type strain XF1. (A) N-propanol content of recombinant strains and XF1. (B) Isobutanol content of recombinant strains and XF1. (C) Isoamyl alcohol content of recombinant strains and XF1. (D) Contents of fusel alcohols produced by recombinant strains XF-L and the wild-type strain XF1. The significant difference between the recombinant strains and XF1 was verified by Student’s t-test (* p < 0.05, ** p < 0.01).
Figure 3.
Contents of fusel alcohols produced by the recombinant strains and the wild-type strain XF1. (A) N-propanol content of recombinant strains and XF1. (B) Isobutanol content of recombinant strains and XF1. (C) Isoamyl alcohol content of recombinant strains and XF1. (D) Contents of fusel alcohols produced by recombinant strains XF-L and the wild-type strain XF1. The significant difference between the recombinant strains and XF1 was verified by Student’s t-test (* p < 0.05, ** p < 0.01).

Figure 4.
Content of total fusel alcohols in the recombinant strains and wild-type strain XF1.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Physicochemical indices of red wines fermented by different strains.
| Strains | Ethanol (%vol) | pH Value | Residual Sugar (g·L−1) | Total Acid (g·L−1) |
|---|---|---|---|---|
| XF1 | 10.43 ± 0.21 | 3.28 ± 0.01 | 0.77 ± 0.11 | 8.63 ± 0.24 |
| XF-T | 10.45 ± 0.26 | 3.27 ± 0.02 | 0.83 ± 0.08 | 8.75 ± 0.18 |
| XF-B | 10.98 ± 0.33 | 3.30 ± 0.01 | 0.82 ± 0.13 | 8.63 ± 0.27 |
| XF-P | 10.64 ± 0.18 | 3.28 ± 0.01 | 0.87 ± 0.19 | 9.00 ± 0.33 |
| XF-L | 10.97 ± 0.25 | 3.27 ± 0.01 | 0.79 ± 0.15 | 8.75 ± 0.13 |
| XF-TP | 10.78 ± 0.17 | 3.32 ± 0.01 | 0.78 ± 0.06 | 8.38 ± 0.29 |
| XF-TB | 10.76 ± 0.24 | 3.31 ± 0.02 | 0.83 ± 0.17 | 8.63 ± 0.15 |
| XF-BP | 10.75 ± 0.22 | 3.32 ± 0.01 | 0.86 ± 0.12 | 8.50 ± 0.21 |
Values are expressed as means ± standard deviations from three independent tests (p > 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yan, T.; Wang, Z.; Zhou, H.; He, J.; Zhou, S. Effects of Four Critical Gene Deletions in Saccharomyces cerevisiae on Fusel Alcohols during Red Wine Fermentation. Fermentation 2023, 9, 379. https://doi.org/10.3390/fermentation9040379
AMA Style
Yan T, Wang Z, Zhou H, He J, Zhou S. Effects of Four Critical Gene Deletions in Saccharomyces cerevisiae on Fusel Alcohols during Red Wine Fermentation. Fermentation. 2023; 9(4):379. https://doi.org/10.3390/fermentation9040379
Chicago/Turabian StyleYan, Tongshuai, Zexiang Wang, Haoyang Zhou, Jiaojiao He, and Shishui Zhou. 2023. "Effects of Four Critical Gene Deletions in Saccharomyces cerevisiae on Fusel Alcohols during Red Wine Fermentation" Fermentation 9, no. 4: 379. https://doi.org/10.3390/fermentation9040379
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.