
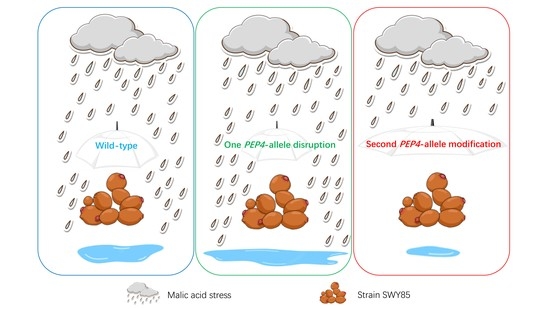
PEP4-Allele Modification Provides an Industrial Brewing Yeast with Malate Stress Tolerance
 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Chemicals
2.3. Acidification of Culture Medium
2.4. Assay of Cell Growth Inhibitory Rates
2.5. Spot Assay
2.6. Intracellular PrA Activity Assay
2.7. Measurement of Intracellular Trehalose Accumulation
2.8. Anaerobic Fermentations
2.9. Statistical Analysis
3. Results
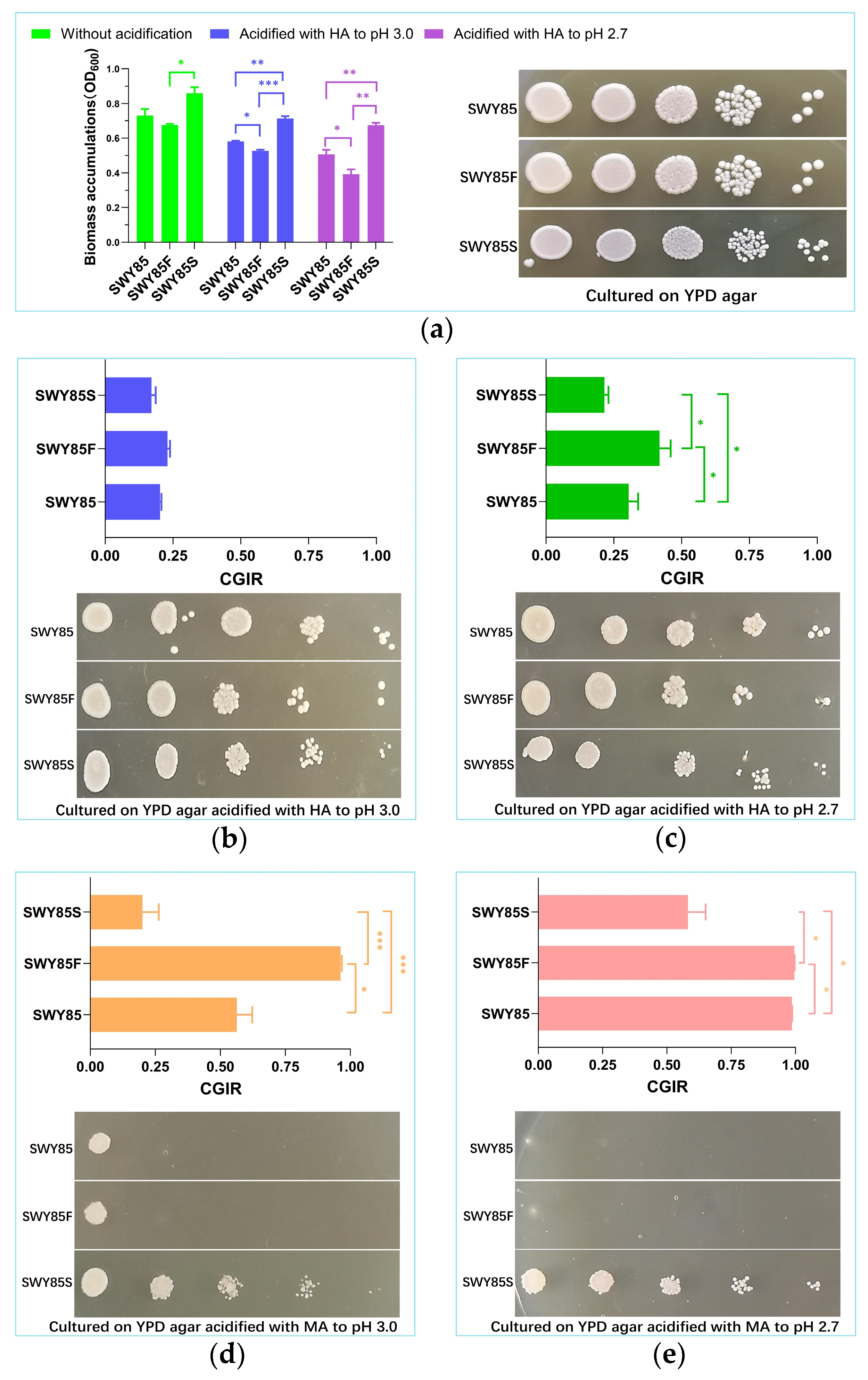
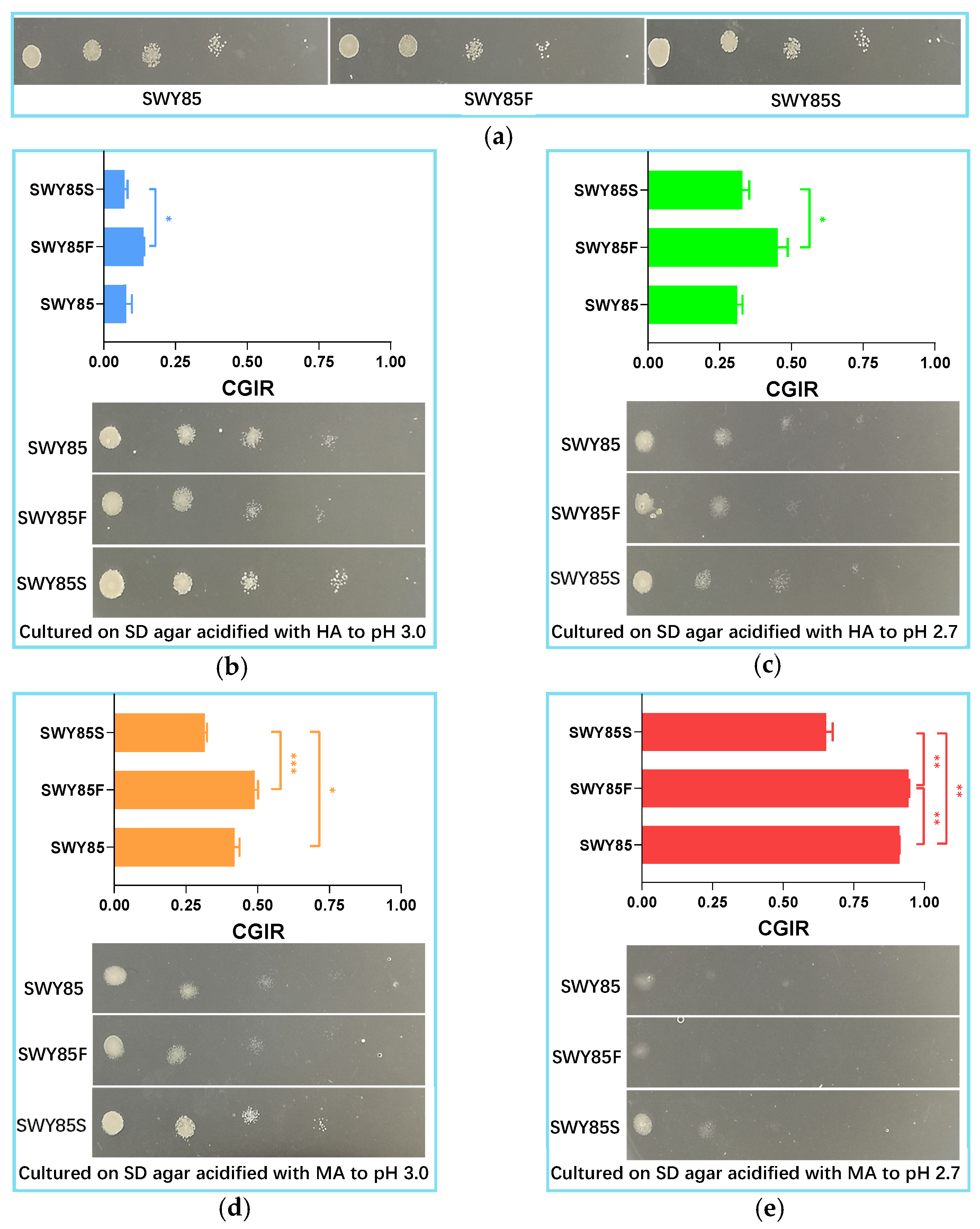
3.1. The Second PEP4-Allele Modification Facilitates Cell Growth under Malic Acid Stress
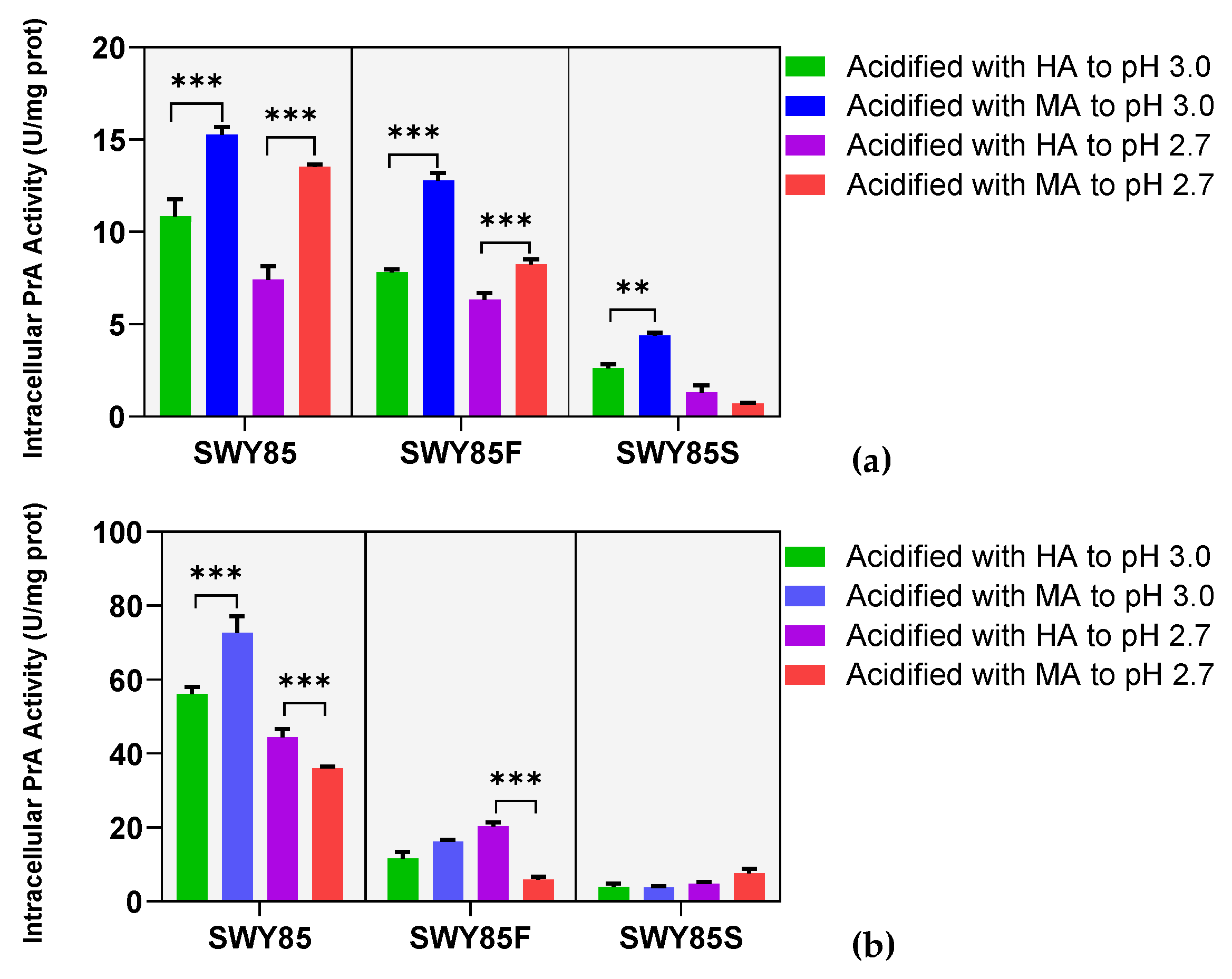
3.2. The Second PEP4-Allele Modification Stabilized Intracellular PrA Activity under Malate Anions
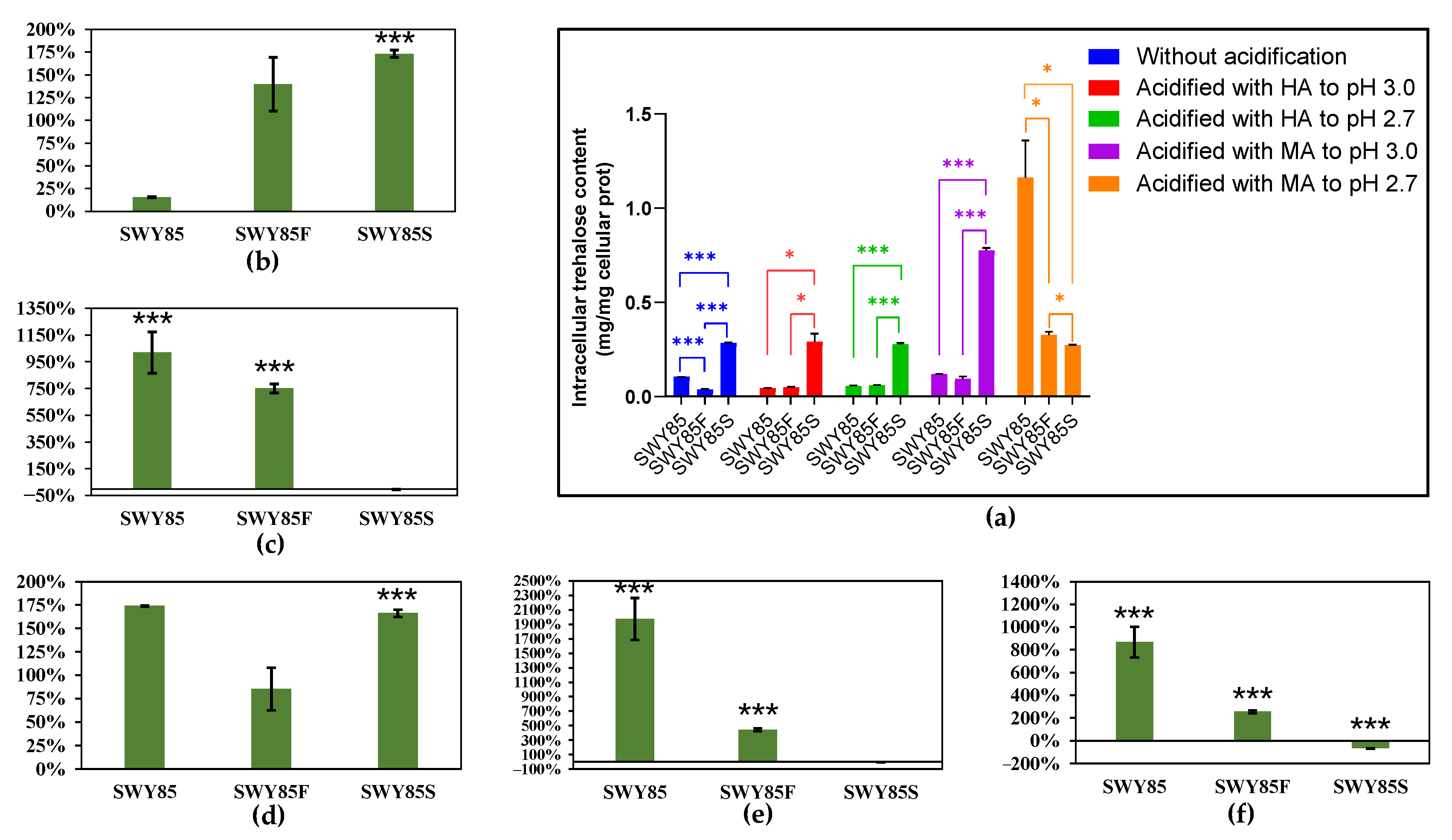
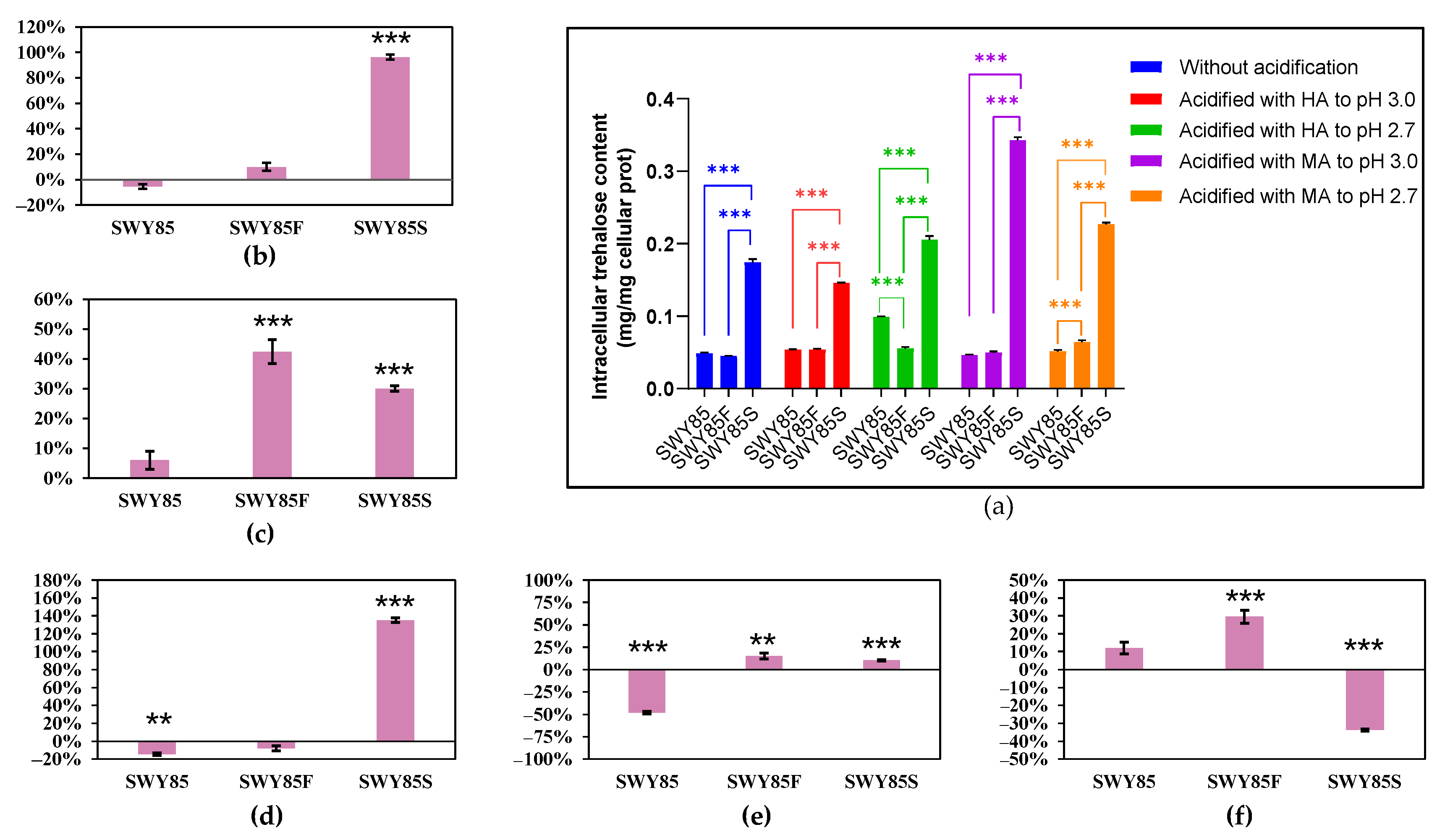
3.3. Opposing Effects on Intracellular Trehalose Accumulation between the PEP4-Allele Modification and Disruption
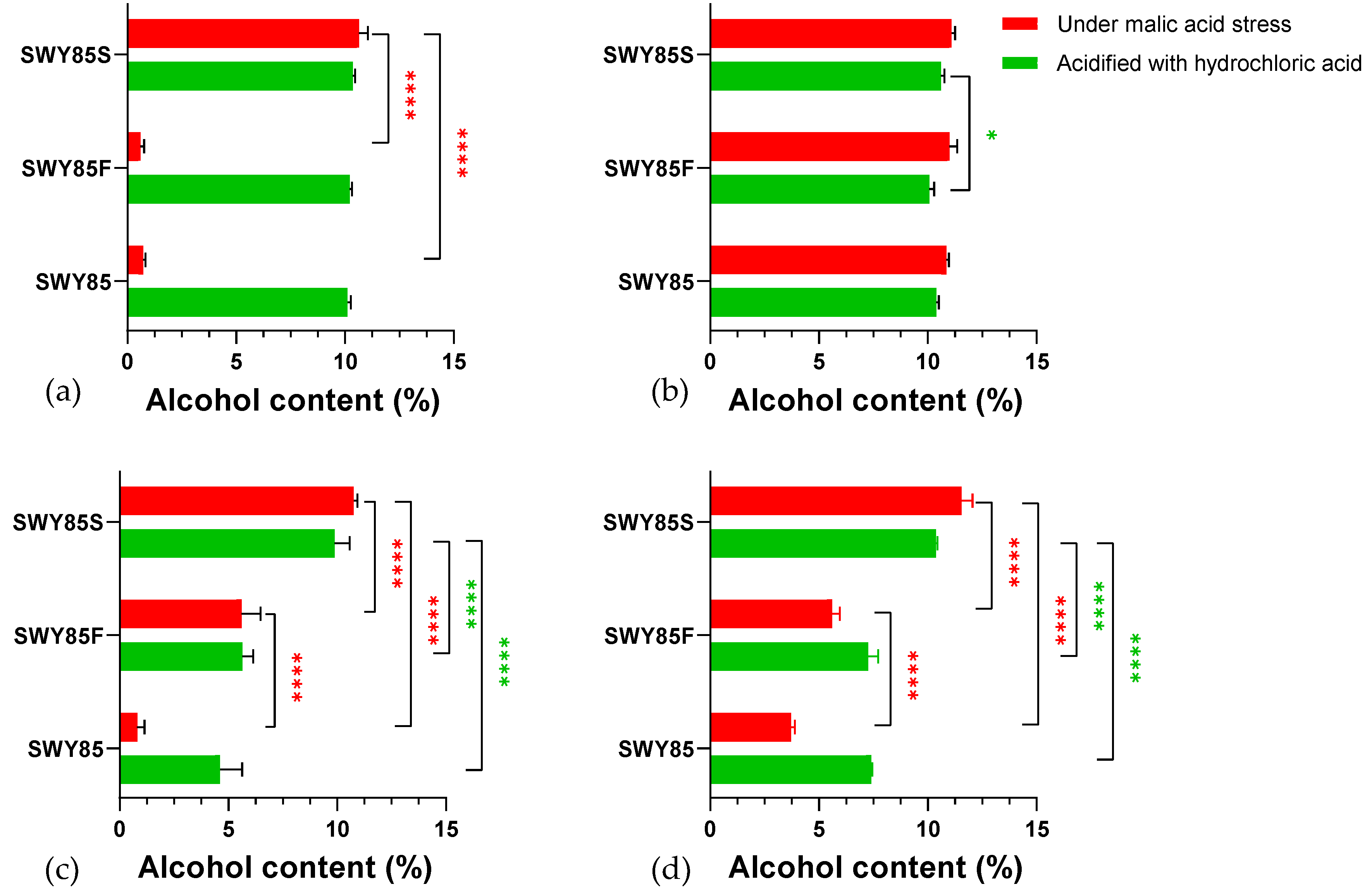
3.4. The PEP4-Allele Modification Maintains Anaerobic Glucose Consumption under Malic Acid Stress
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Attfield, P.V. Stress tolerance: The key to effective strains of industrial baker’s yeast. Nat. Biotechnol. 1997, 15, 1351–1357. [Google Scholar] [CrossRef] [PubMed]
- Betlej, G.; Bator, E.; Oklejewicz, B.; Potocki, L.; Górka, A.; Slowik-Borowiec, M.; Czarny, W.; Domka, W.; Kwiatkowska, A. Long-term adaption to high osmotic stress as a tool for improving enological characteristics in industrial wine yeast. Genes 2020, 11, 576. [Google Scholar] [CrossRef] [PubMed]
- Gibson, B.R.; Lawrence, S.J.; Leclaire, J.P.R.; Powell, C.D.; Smart, K.A. Yeast responses to stresses associated with industrial brewery handling. FEMS Microbiol. Rev. 2007, 31, 535–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, J.M.; Huang, X.W.; Zhang, L.M.; Zhao, N.; Yang, D.M.; Zhang, K.Q. Tolerance and stress response to ethanol in the yeast Saccharomyces cerevisiae. Appl. Microbiol. Biot. 2009, 85, 253–263. [Google Scholar] [CrossRef]
- Stanley, D.; Bandara, A.; Fraser, S.; Chambers, P.J.; Stanley, G.A. The ethanol stress response and ethanol tolerance of Saccharomyces cerevisiae. J. Appl. Microbiol. 2010, 109, 13–24. [Google Scholar] [CrossRef]
- Li, H.; Wang, H.L.; Du, J.; Du, G.; Zhan, J.C.; Huang, W.D. Trehalose protects wine yeast against oxidation under thermal stress. World J. Microb. Biot. 2010, 26, 969–976. [Google Scholar] [CrossRef]
- Kitichantaropas, Y.; Boonchird, C.; Sugiyama, M.; Kaneko, Y.; Harashima, S.; Auesukaree, C. Cellular mechanisms contributing to multiple stress tolerance in Saccharomyces cerevisiae strains with potential use in high-temperature ethanol fermentation. AMB Express 2016, 6, 107. [Google Scholar] [CrossRef] [Green Version]
- Auesukaree, C. Molecular mechanisms of the yeast adaptive response and tolerance to stresses. J. Biosci. Bioeng. 2017, 124, 133–142. [Google Scholar] [CrossRef]
- Narendranath, N.V.; Power, R. Relationship between pH and medium dissolved solids in terms of growth and metabolism of lactobacilli and Saccharomyces cerevisiae during ethanol production. Appl. Environ. Microb. 2005, 71, 2239–2243. [Google Scholar] [CrossRef] [Green Version]
- Attfield, P.V.; Kletsas, S. Hyperosmotic stress response by strains of bakers’ yeasts in high sugar concentration medium. Lett. Appl. Microbiol. 2000, 31, 323–327. [Google Scholar] [CrossRef]
- Mira, N.P.; Teixeira, M.C.; Sá-Correia, I. Adaptive response and tolerance to weak acids in Saccharomyces cerevisiae: A genome-wide view. Omics A J. Integr. Biol. 2010, 14, 525–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardona, F.; Carrasco, P.; Pérez-Ortín, J.E.; lí del Olmo, M.; Aranda, A. A novel approach for the improvement of stress resistance in wine yeasts. Int. J. Food. Microbiol. 2007, 114, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Giannattasio, S.; Guaragnella, N.; Ždralević, M.; Marra, E. Molecular mechanisms of Saccharomyces cerevisiae stress adaptation and programmed cell death in response to acetic acid. Front. Microbiol. 2013, 4, 33. [Google Scholar] [CrossRef] [Green Version]
- Meijnen, J.P.; Randazzo, P.; Foulquié-Moreno, M.R.; Van Den Brink, J.; Vandecruys, P.; Stojiljkovic, M.; Dumortier, F.; Zalar, P.; Boekhout, T.; Gunde-Cimerman, N.; et al. Polygenic analysis and targeted improvement of the complex trait of high acetic acid tolerance in the yeast Saccharomyces cerevisiae. Biotechnol. Biofuels 2016, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- Samanfar, B.; Shostak, K.; Moteshareie, H.; Hajikarimlou, M.; Shaikho, S.; Omidi, K.; Hooshyar, M.; Burnside, D.; Márquez, I.G.; Kazmirchuk, T.; et al. The sensitivity of the yeast, Saccharomyces cerevisiae, to acetic acid is influenced by DOM34 and RPL36A. PeerJ 2017, 5, e4037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stojiljkovic, M.; Foulquié-Moreno, M.R.; Thevelein, J.M. Polygenic analysis of very high acetic acid tolerance in the yeast Saccharomyces cerevisiae reveals a complex genetic background and several new causative alleles. Biotechnol. Biofuels 2020, 13, 126. [Google Scholar] [CrossRef] [PubMed]
- Mitsui, R.; Yamada, R.; Matsumoto, T.; Yoshihara, S.; Tokumoto, H.; Ogino, H. Construction of lactic acid-tolerant Saccharomyces cerevisiae by using CRISPR-Cas-mediated genome evolution for efficient d-lactic acid production. Appl. Microbiol. Biot. 2020, 104, 9147–9158. [Google Scholar] [CrossRef]
- Yamada, R.; Kumata, Y.; Mitsui, R.; Matsumoto, T.; Ogino, H. Improvement of lactic acid tolerance by cocktail δ-integration strategy and identification of the transcription factor PDR3 responsible for lactic acid tolerance in yeast Saccharomyces cerevisiae. World J. Microb. Biot. 2021, 37, 19. [Google Scholar] [CrossRef]
- Baldi, N.; de Valk, S.C.; Sousa-Silva, M.; Casal, M.; Soares-Silva, I.; Mans, R. Evolutionary engineering reveals amino acid substitutions in Ato2 and Ato3 that allow improved growth of Saccharomyces cerevisiae on lactic acid. FEMS Yeast Res. 2021, 21, foab033. [Google Scholar] [CrossRef]
- Stratford, M.; Nebe-von-Caron, G.; Steels, H.; Novodvorska, M.; Ueckert, J.; Archer, D.B. Weak-acid preservatives: pH and proton movements in the yeast Saccharomyces cerevisiae. Int. J. Food Microbiol. 2013, 161, 164–171. [Google Scholar] [CrossRef]
- Geoghegan, I.A.; Stratford, M.; Bromley, M.; Archer, D.B.; Avery, S.V. Weak acid resistance A (WarA), a novel transcription factor required for regulation of weak-acid resistance and spore-spore heterogeneity in Aspergillus niger. Msphere 2020, 5, e00685-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndukwe, J.K.; Aliyu, G.O.; Onwosi, C.O.; Chukwu, K.O.; Ezugworie, F.N. Mechanisms of weak acid-induced stress tolerance in yeasts: Prospects for improved bioethanol production from lignocellulosic biomass. Process Biochem. 2020, 90, 118–130. [Google Scholar] [CrossRef]
- Volschenk, H.; Viljoen, M.; Grobler, J.; Petzold, B.; Bauer, F.; Subden, R.E.; Young, R.A.; Lonvaud, A.; Denayrolles, M.; van Vuuren, H.J. Engineering pathways for malate degradation in Saccharomyces cerevisiae. Nat. Biotechnol. 1997, 15, 253–257. [Google Scholar] [CrossRef]
- Gurban, A.M.; Prieto-Simón, B.; Marty, J.L.; Noguer, T. Malate biosensors for the monitoring of malolactic fermentation: Different approaches. Anal. Lett. 2006, 39, 1543–1558. [Google Scholar] [CrossRef]
- Fletcher, E.; Feizi, A.; Bisschops, M.M.; Hallström, B.M.; Khoomrung, S.; Siewers, V.; Nielsen, J. Evolutionary engineering reveals divergent paths when yeast is adapted to different acidic environments. Metab. Eng. 2017, 39, 19–28. [Google Scholar] [CrossRef]
- Côrte-Real, M.; Leao, C. Transport of malic acid and other dicarboxylic acids in the yeast Hansenula anomala. Appl. Environ. Microb. 1990, 56, 1109–1113. [Google Scholar] [CrossRef] [Green Version]
- Zelle, R.M.; De Hulster, E.; Van Winden, W.A.; De Waard, P.; Dijkema, C.; Winkler, A.A.; Geertman, J.M.A.; Van Dijken, J.P.; Pronk, J.T.; Van Maris, A.J. Malic acid production by Saccharomyces cerevisiae: Engineering of pyruvate carboxylation, oxaloacetate reduction, and malate export. Appl. Environ. Microb. 2008, 74, 2766–2777. [Google Scholar] [CrossRef] [Green Version]
- Volschenk, H.; Van Vuuren, H.J.; Viljoen-Bloom, M. Malic acid in wine: Origin, function and metabolism during vinification. S. Afr. J. Enol. Vitic. 2006, 27, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Kunkee, R.E. Some roles of malic acid in the malolactic fermentation in wine making. FEMS Microbiol. Lett. 1991, 88, 55–71. [Google Scholar] [CrossRef]
- van den Hazel, H.B.; Kielland-Brandt, M.C.; Winther, J.R. Biosynthesis and function of yeast vacuolar proteases. Yeast 1996, 12, 1–16. [Google Scholar] [CrossRef]
- Wolff, A.M.; Din, N.; Petersen, J.G.L. Vacuolar and extracellular maturation of Saccharomyces cerevisiae proteinase A. Yeast 1996, 12, 823–832. [Google Scholar] [CrossRef]
- Zhang, H.B.; Shao, F.F.; Cong, J.H.; Huang, Y.; Chen, M.F.; He, W.X.; Zhang, T.; Liu, L.Y.; Yao, M.Z.; Gwabin, H.; et al. Modification of the second PEP4-allele enhances citric acid stress tolerance during cultivation of an industrial rice wine yeast strain with one PEP4-allele disrupted. LWT Food Sci. Technol. 2021, 152, 112286. [Google Scholar] [CrossRef]
- Parr, C.L.; Keates, R.A.; Bryksa, B.C.; Ogawa, M.; Yada, R.Y. The structure and function of Saccharomyces cerevisiae proteinase A. Yeast 2007, 24, 467–480. [Google Scholar] [CrossRef]
- Callis, J. Regulation of protein degradation. Plant Cell 1995, 7, 845–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teichert, U.; Mechler, B.; Müller, H.; Wolf, D.H. Lysosomal (vacuolar) proteinases of yeast are essential catalysts for protein degradation, differentiation, and cell survival. J. Biol. Chem. 1989, 264, 16037–16045. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.B.; Zhang, H.F.; Chen, Q.H.; Ruan, H.; Fu, M.L.; He, G.Q. Effects of proteinase A on cultivation and viability characteristics of industrial Saccharomyces cerevisiae WZJ. Zhejiang Univ. Sci. B 2009, 10, 769–776. [Google Scholar] [CrossRef] [Green Version]
- Mahmud, S.A.; Nagahisa, K.; Hirasawa, T.; Yoshikawa, K.; Ashitani, K.; Shimizu, H. Effect of trehalose accumulation on response to saline stress in Saccharomyces cerevisiae. Yeast 2009, 26, 17–30. [Google Scholar] [CrossRef] [Green Version]
- Caspeta, L.; Castillo, T.; Nielsen, J. Modifying yeast tolerance to inhibitory conditions of ethanol production processes. Front. Bioeng. Biotech. 2015, 3, 184. [Google Scholar] [CrossRef] [Green Version]
- Tesnière, C. Importance and role of lipids in wine yeast fermentation. Appl. Microbiol. Biot. 2019, 103, 8293–8300. [Google Scholar] [CrossRef]
- Zhang, M.M.; Xiong, L.; Tang, Y.J.; Mehmood, M.A.; Zhao, Z.K.; Bai, F.W.; Zhao, X.Q. Enhanced acetic acid stress tolerance and ethanol production in Saccharomyces cerevisiae by modulating expression of the de novo purine biosynthesis genes. Biotechnol. Biofuels 2019, 12, 116. [Google Scholar] [CrossRef]
- Xu, K.; Qin, L.; Bai, W.; Wang, X.; Li, F.; Ren, S.; Gao, X.; Chen, B.; Tong, Y.; Li, J.; et al. Multilevel defense system (MDS) relieves multiple stresses for economically boosting ethanol production of industrial Saccharomyces cerevisiae. ACS Energy Lett. 2020, 5, 572–582. [Google Scholar] [CrossRef]
- Jones, E.W. The synthesis and function of proteases in Saccharomyces: Genetic approaches. Annu. Rev. Genet. 1984, 18, 233–270. [Google Scholar] [CrossRef] [PubMed]
- Hemmings, B.A.; Zubenko, G.S.; Hasilik, A.; Jones, E.W. Mutant defective in processing of an enzyme located in the lysosome-like vacuole of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1981, 78, 43–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, E.W.; Zubenko, G.S.; Parker, R.R. PEP4 gene function is required for expression of several vacuolar hydrolases in Saccharomyces cerevisiae. Genetics 1982, 102, 665–677. [Google Scholar] [CrossRef] [PubMed]
- Zubenko, G.S.; Park, F.J.; Jones, E.W. Mutations in PEP4 locus of Saccharomyces cerevisiae block final step in maturation of two vacuolar hydrolases. Proc. Natl. Acad. Sci. USA 1983, 80, 510–514. [Google Scholar] [CrossRef] [Green Version]
- Ammerer, G.; Hunter, C.P.; Rothman, J.H.; Saari, G.C.; Valls, L.A.; Stevens, T.H. PEP4 gene of Saccharomyces cerevisiae encodes proteinase A, a vacuolar enzyme required for processing of vacuolar precursors. Mol. Cell. Biol. 1986, 6, 2490–2499. [Google Scholar] [CrossRef]
- Woolford, C.A.; Daniels, L.B.; Park, F.J.; Jones, E.W.; Van Arsdell, J.N.; Innis, M.A. The PEP4 gene encodes an aspartyl protease implicated in the posttranslational regulation of Saccharomyces cerevisiae vacuolar hydrolases. Mol. Cell. Biol. 1986, 6, 2500–2510. [Google Scholar] [CrossRef] [Green Version]
- Hegemann, J.H.; Heick, S.B. Delete and repeat: A comprehensive toolkit for sequential gene knockout in the budding yeast Saccharomyces cerevisiae. In Strain Engineering Methods and Protocols; Humana Totowa: Paramus, NJ, USA, 2011; pp. 189–206. [Google Scholar] [CrossRef]
- Chen, D.C.; Wang, B.D.; Chou, P.Y.; Kuo, T.T. Asparagine as a nitrogen source for improving the secretion of mouse α-amylase in Saccharomyces cerevisiae protease A-deficient strains. Yeast 2000, 16, 207–217. [Google Scholar] [CrossRef]
- Komeda, T.; Sakai, Y.; Kato, N.; Kondo, K. Construction of Protease-deficient Candida boidinii Strains Useful for Recombinant Protein Production: Cloning and Disruption of Proteinase A Gene (PEP4) and Proteinase B Gen. Biosci. Biotech. Bioch. 2002, 66, 628–631. [Google Scholar] [CrossRef]
- Zhang, H.B.; Ruan, H.; Li, W.F.; Zhang, W.; Su, Z.R.; He, G.Q.; Chen, Q.H. Construction of recombinant industrial S. cerevisiae strain with barley lipid-transfer protein 1 secretion capability and lower PrA activity. Eur. Food Res. Technol. 2011, 233, 707–716. [Google Scholar] [CrossRef]
- Lu, J.; Dong, J.; Wu, D.; Chen, Y.; Guo, X.; Shi, Y.; Sun, X.; Xiao, D. Construction of recombinant industrial brewer’s yeast with lower diacetyl production and proteinase A activity. Eur. Food Res. Technol. 2012, 235, 951–961. [Google Scholar] [CrossRef]
- Wu, M.; Shen, Q.; Yang, Y.; Zhang, S.; Qu, W.; Chen, J.; Sun, H.; Chen, S. Disruption of YPS1 and PEP4 genes reduces proteolytic degradation of secreted HSA/PTH in Pichia pastoris GSJ. Ind. Microbiol. Biot. 2013, 40, 589–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Guan, X.; Wang, Y.; Li, L.; Wu, D.; Chen, Y.; Pei, H.; Xiao, D. Reduction of biogenic amines production by eliminating the PEP4 gene in Saccharomyces cerevisiae during fermentation of Chinese rice wine. Food Chem. 2015, 178, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Alugoju, P.; Janardhanshetty, S.S.; Subaramanian, S.; Periyasamy, L.; Dyavaiah, M. Quercetin protects yeast Saccharomyces cerevisiae pep4 mutant from oxidative and apoptotic stress and extends chronological lifespan. Curr. Microbiol. 2018, 75, 519–530. [Google Scholar] [CrossRef]
- Chen, D.C.; Chen, S.Y.; Gee, M.F.; Pan, J.T.; Kuo, T.T. A variant of Saccharomyces cerevisiae pep4 strain with improved oligotrophic proliferation, cell survival and heterologous secretion of α-amylase. Appl. Microbiol. Biot. 1999, 51, 185–192. [Google Scholar] [CrossRef]
- Giannattasio, S.; Guaragnella, N.; Corte-Real, M.; Passarella, S.; Marra, E. Acid stress adaptation protects Saccharomyces cerevisiae from acetic acid-induced programmed cell death. Gene 2005, 354, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Kawahata, M.; Masaki, K.; Fujii, T.; Iefuji, H. Yeast genes involved in response to lactic acid and acetic acid: Acidic conditions caused by the organic acids in Saccharomyces cerevisiae cultures induce expression of intracellular metal metabolism genes regulated by Aft1p. FEMS Yeast Res. 2006, 6, 924–936. [Google Scholar] [CrossRef] [Green Version]
- Abbott, D.A.; Knijnenburg, T.A.; De Poorter, L.M.; Reinders, M.J.; Pronk, J.T.; Van Maris, A.J. Generic and specific transcriptional responses to different weak organic acids in anaerobic chemostat cultures of Saccharomyces cerevisiae. Fems Yeast Res. 2007, 7, 819–833. [Google Scholar] [CrossRef] [Green Version]
- Ullah, A.; Orij, R.; Brul, S.; Smits, G.J. Quantitative analysis of the modes of growth inhibition by weak organic acids in Saccharomyces cerevisiae. Appl. Environ. Microb. 2012, 78, 8377–8387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Amore, T.; Crumplen, R.; Stewart, G.G. The involvement of trehalose in yeast stress tolerance. J. Ind. Microbiol. 1991, 7, 191–195. [Google Scholar] [CrossRef]
- Van Dijck, P.; Colavizza, D.; Smet, P.; Thevelein, J.M. Differential importance of trehalose in stress resistance in fermenting and nonfermenting Saccharomyces cerevisiae cells. Appl. Environ. Microb. 1995, 61, 109–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshiyama, Y.; Tanaka, K.; Yoshiyama, K.; Hibi, M.; Ogawa, J.; Shima, J. Trehalose accumulation enhances tolerance of Saccharomyces cerevisiae to acetic acid. J. Biosci. Bioeng. 2015, 119, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.C. A possible role of trehalose in osmotolerance and ethanol tolerance in Saccharomyces cerevisiae. FEMS Microbiol. Lett. 1997, 152, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, S.A.; Hirasawa, T.; Shimizu, H. Differential importance of trehalose accumulation in Saccharomyces cerevisiae in response to various environmental stresses. J. Biosci. Bioeng. 2010, 109, 262–266. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Hu, W.; Lu, Y.; Shen, C.; Yao, H.; Yang, X.; Yao, X.; Lu, T.; Hu, B. PEP4-Allele Modification Provides an Industrial Brewing Yeast with Malate Stress Tolerance. Fermentation 2023, 9, 378. https://doi.org/10.3390/fermentation9040378
Zhang H, Hu W, Lu Y, Shen C, Yao H, Yang X, Yao X, Lu T, Hu B. PEP4-Allele Modification Provides an Industrial Brewing Yeast with Malate Stress Tolerance. Fermentation. 2023; 9(4):378. https://doi.org/10.3390/fermentation9040378
Chicago/Turabian StyleZhang, Hongbo, Wanting Hu, Yuhang Lu, Chi Shen, Hongfei Yao, Xiaomei Yang, Xiaomin Yao, Tianyi Lu, and Baowei Hu. 2023. "PEP4-Allele Modification Provides an Industrial Brewing Yeast with Malate Stress Tolerance" Fermentation 9, no. 4: 378. https://doi.org/10.3390/fermentation9040378