
Molecular and Technological Characterization of Saccharomyces cerevisiae from Sourdough

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
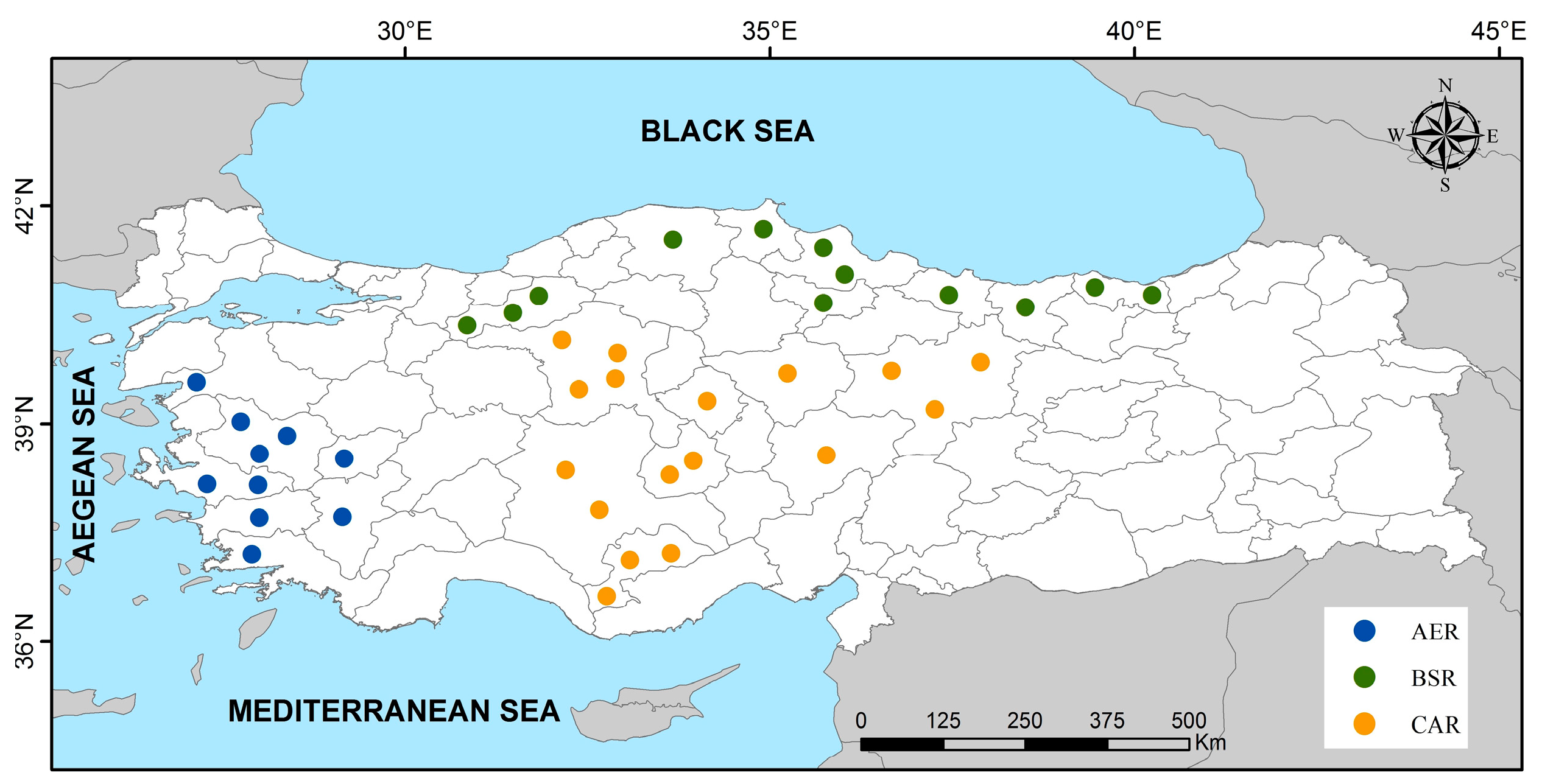
2.1. Yeast Isolates
2.2. Molecular Analyses
2.2.1. DNA Extraction
2.2.2. Molecular Confirmation of the Isolates
2.2.3. ISSR Analyses
2.2.4. Statistical Evaluation of Molecular Analyses
2.3. Technological Characterization
2.3.1. Growth at Different pH Values, NaCl Concentrations and Temperatures
2.3.2. Resistance to Lactic Acid and Acetic Acid
2.3.3. Fermentation Rate
2.3.4. Leavening Activity
2.3.5. Statistical Analyses
2.4. Trait-Loci Associations
3. Results
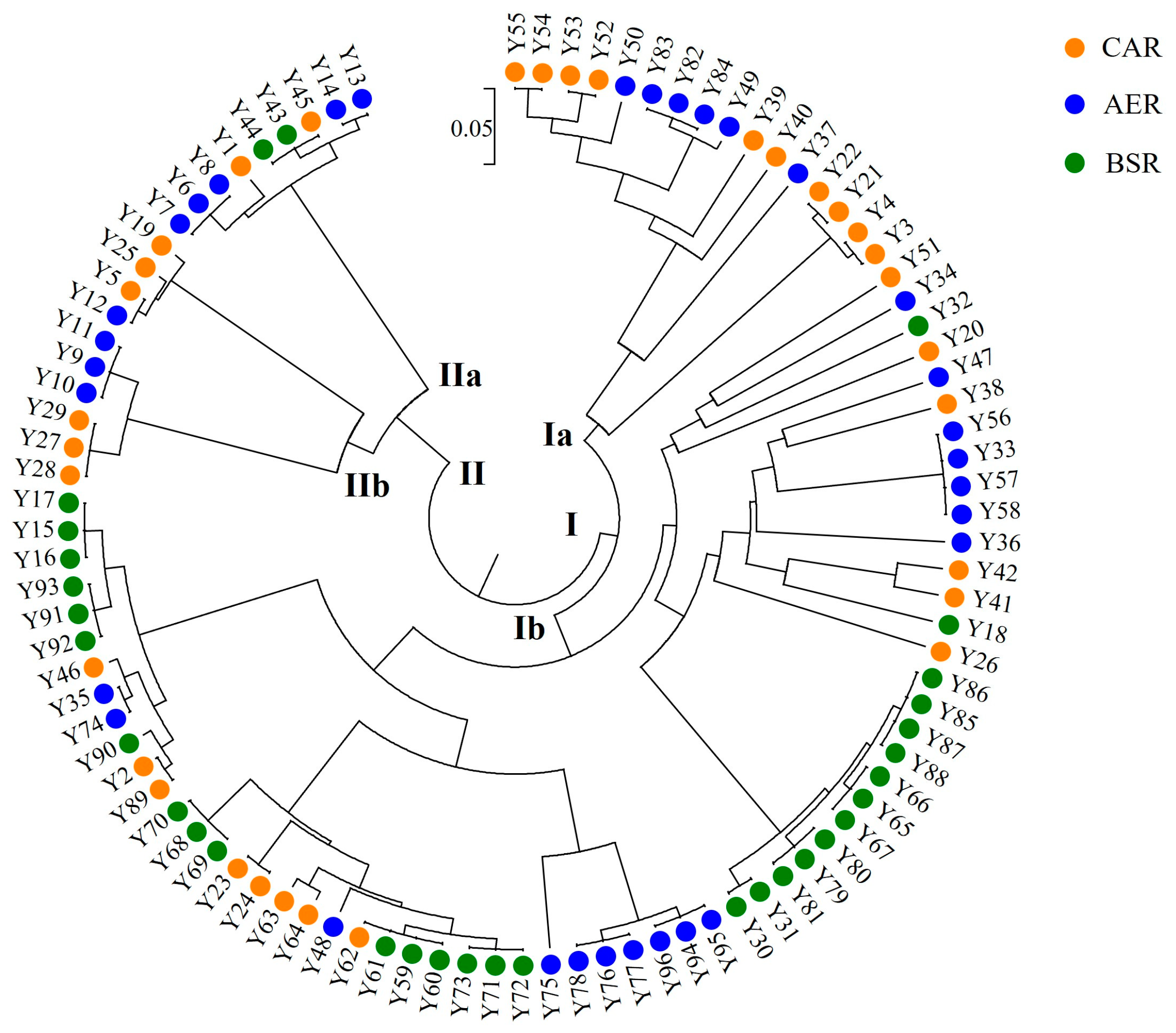
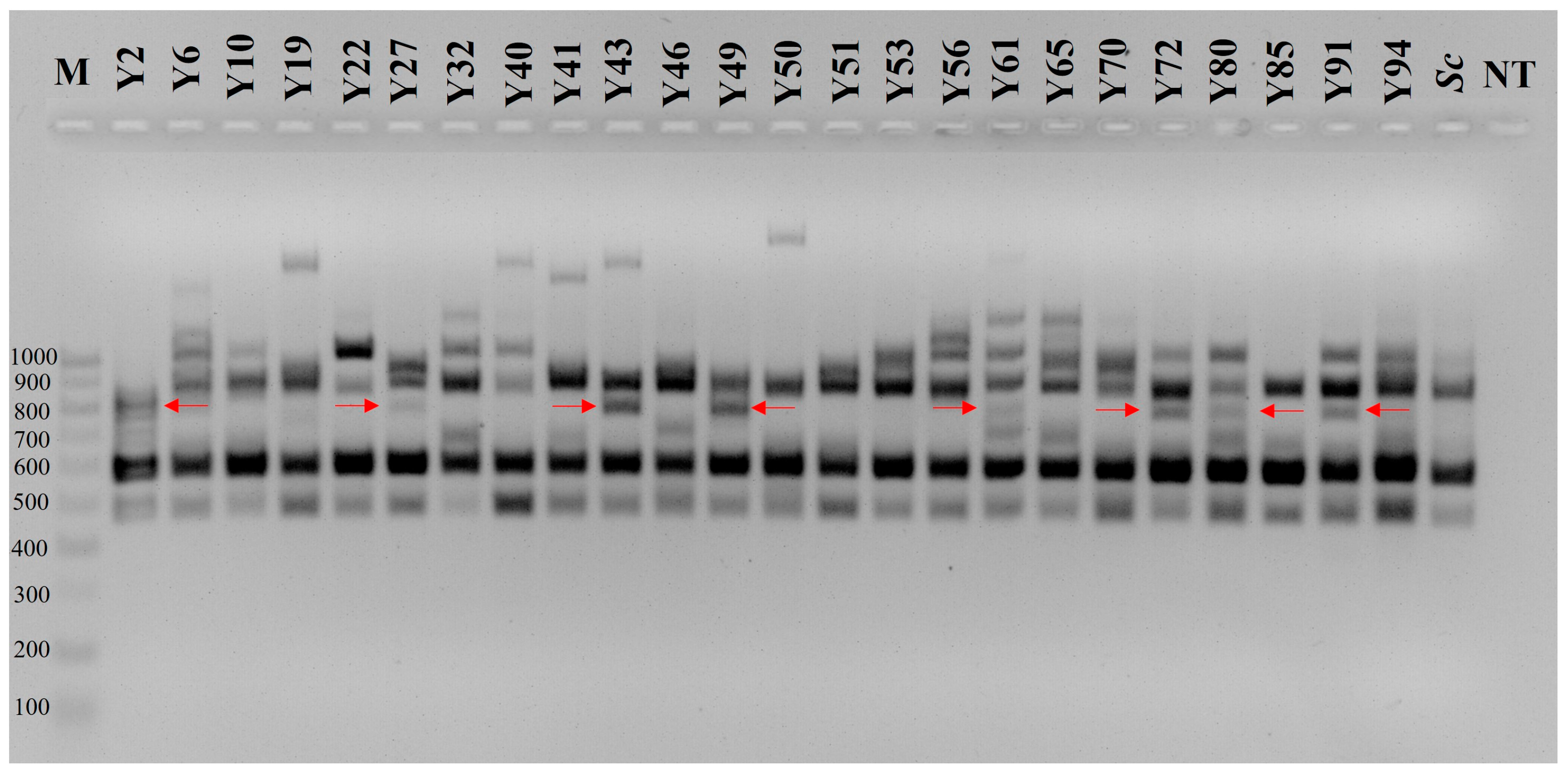
3.1. Intraspecific Genetic Variation
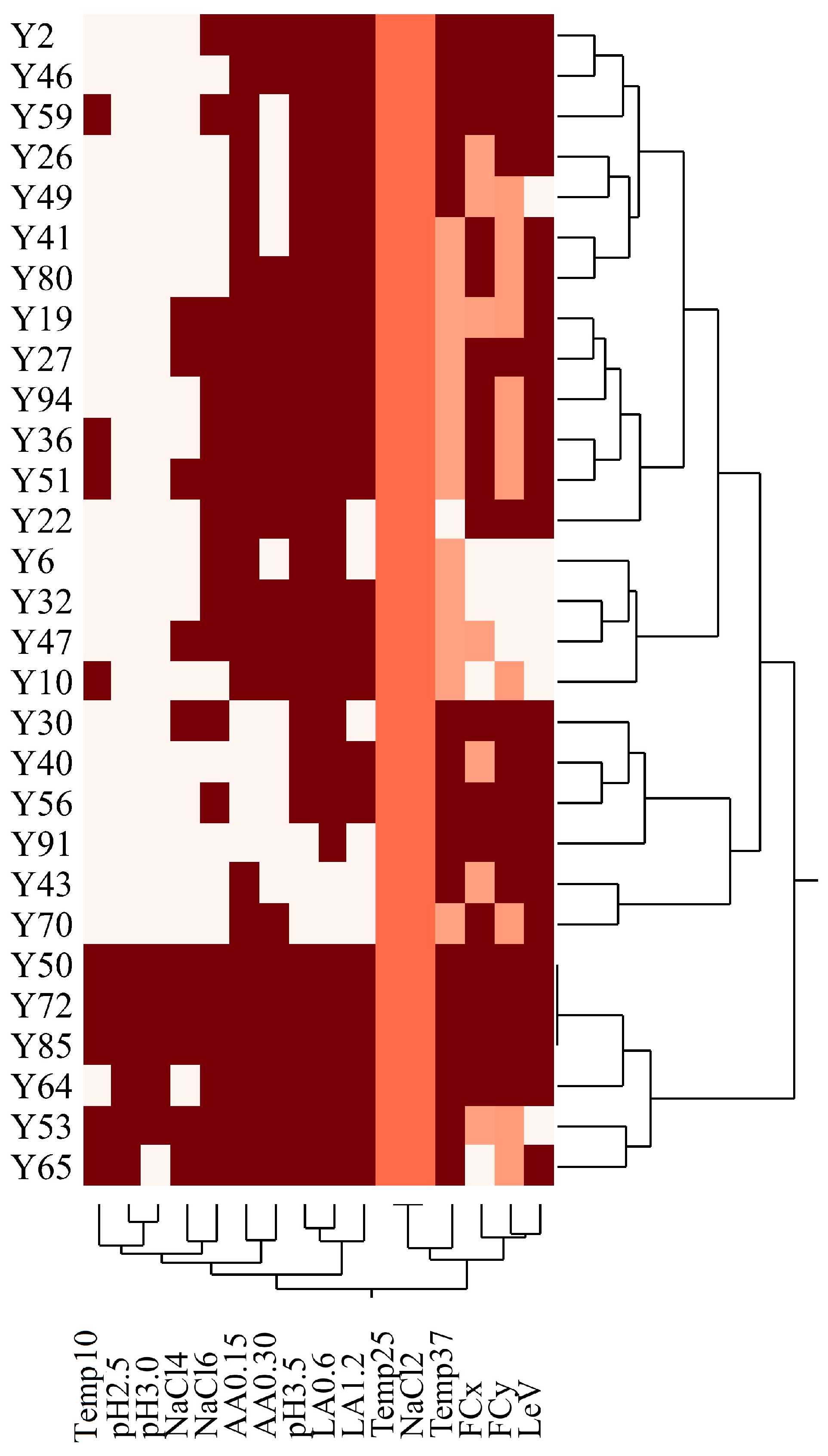
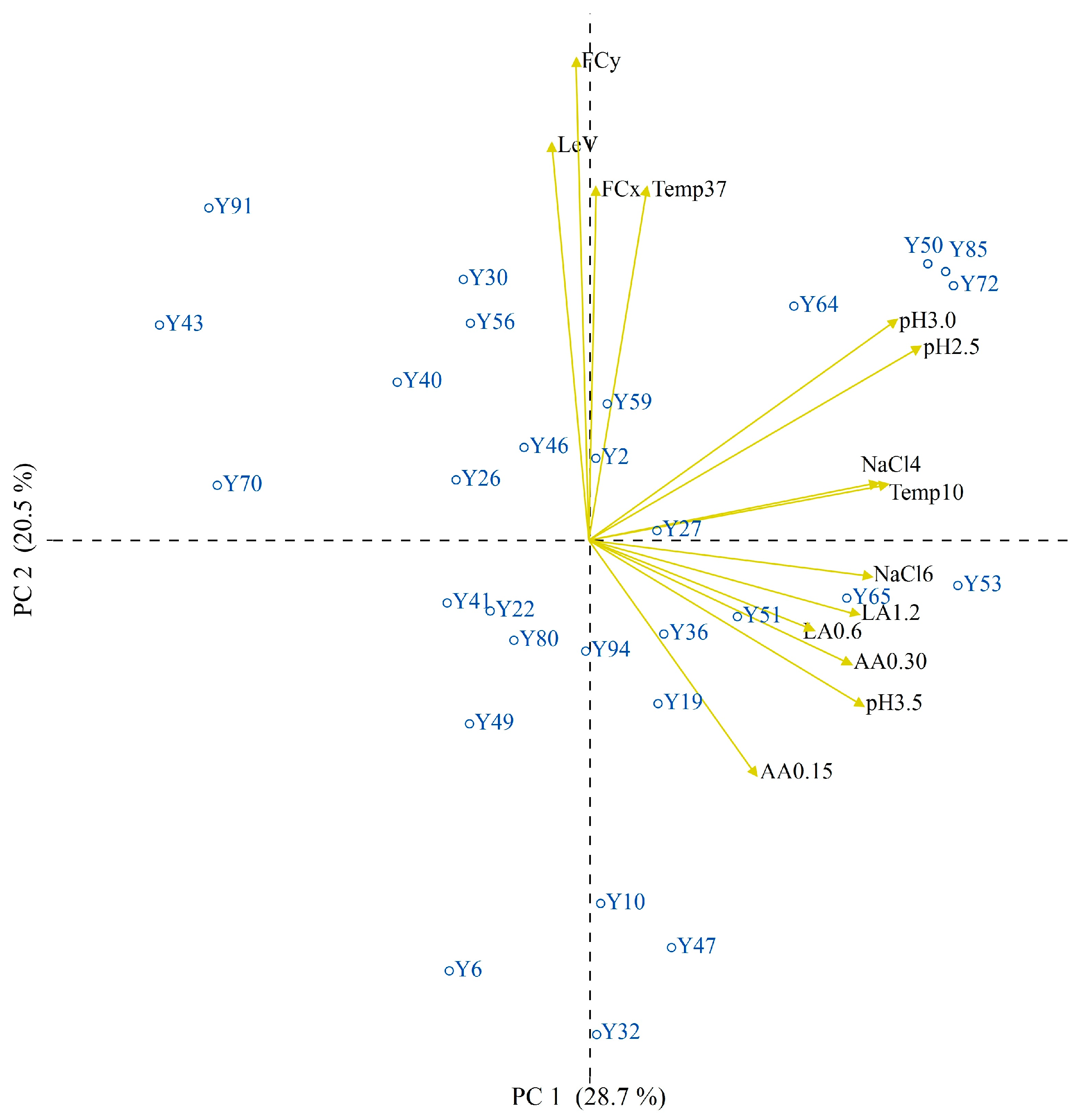
3.2. Technological Characterization
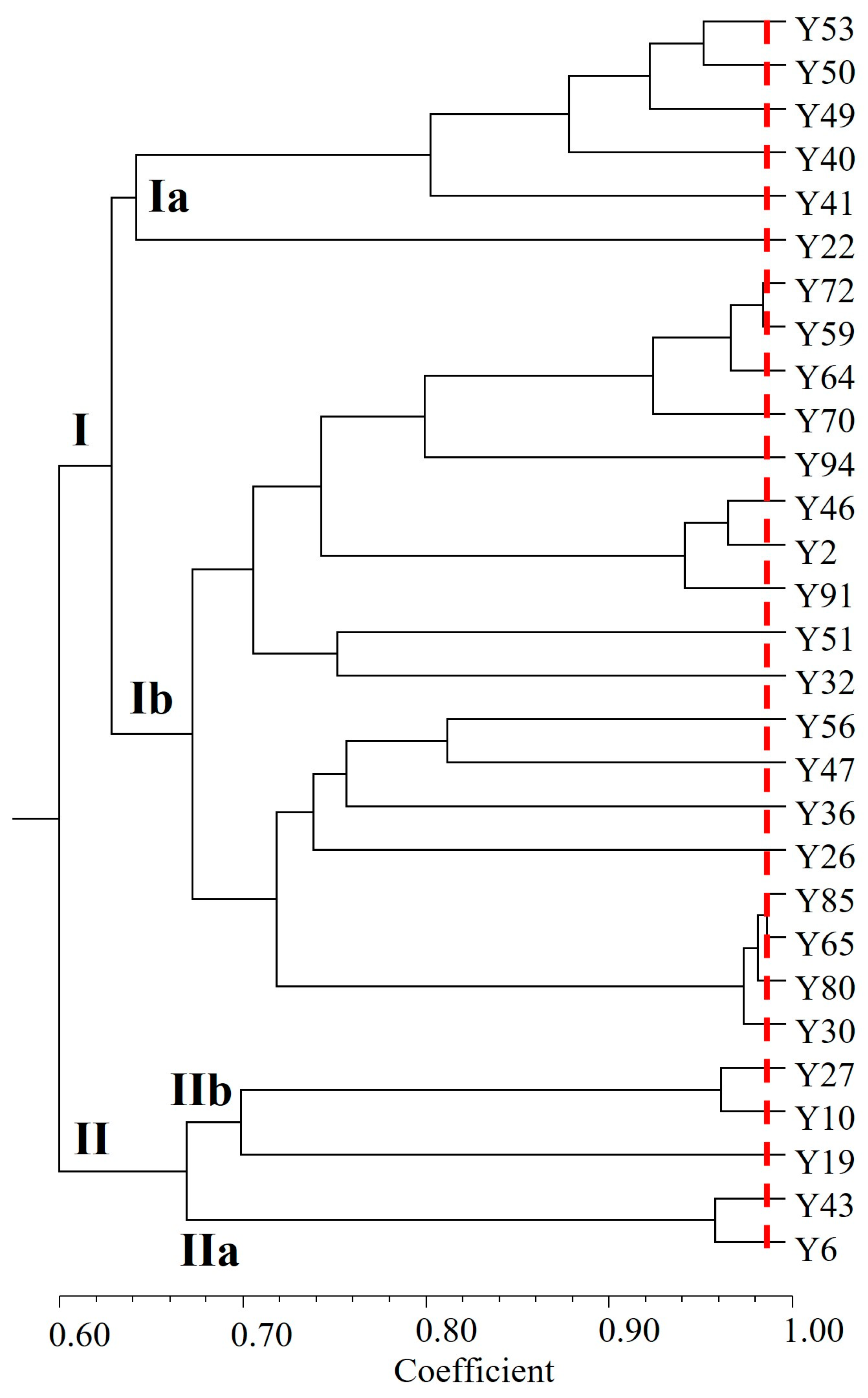
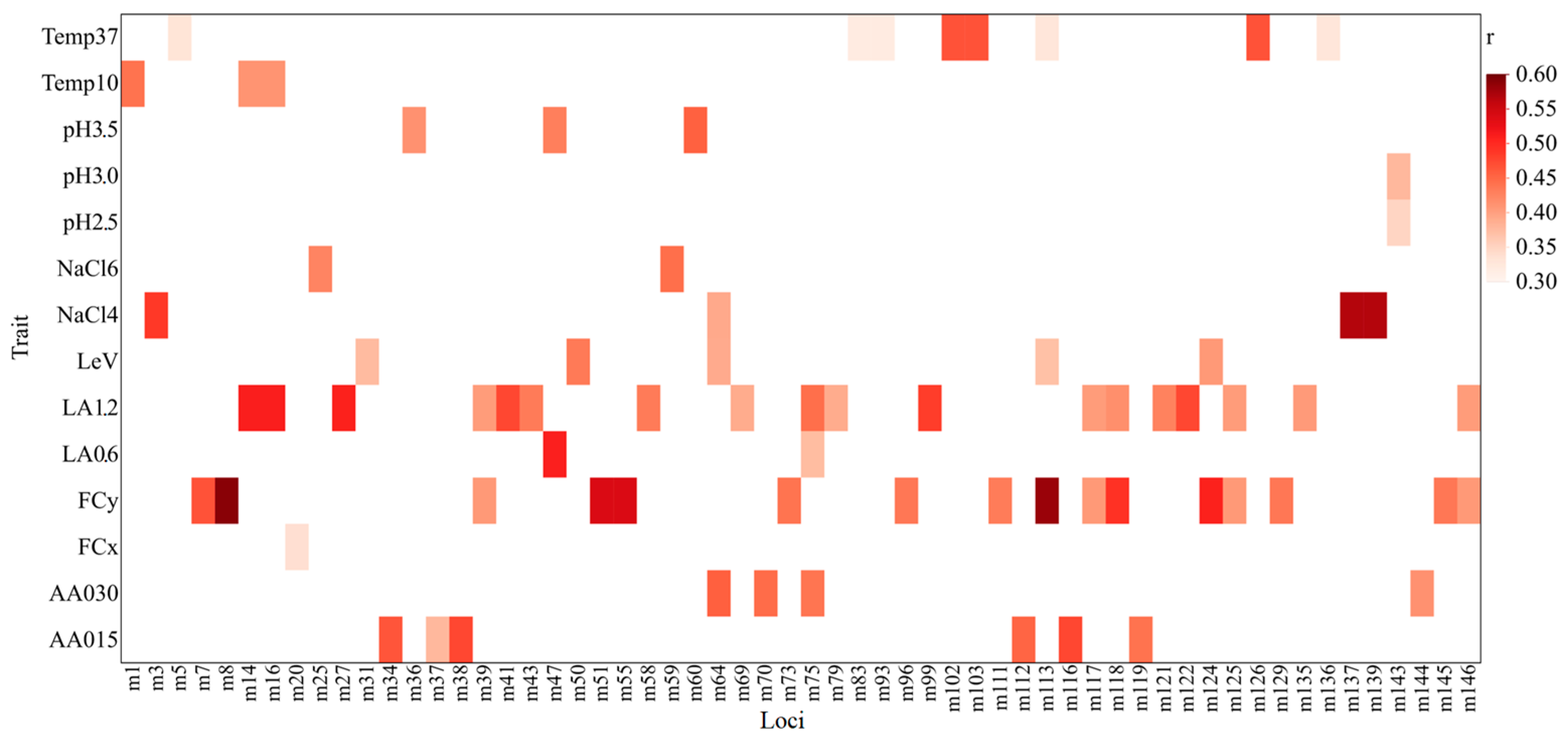
3.3. Trait-Loci Associations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Vuyst, L.; Neysens, P. The Sourdough Microflora: Biodiversity and Metabolic Interactions. Trends Food Sci. Technol. 2005, 16, 43–56. [Google Scholar] [CrossRef]
- De Vuyst, L.; Harth, H.; Van Kerrebroeck, S.; Leroy, F. Yeast Diversity of Sourdoughs and Associated Metabolic Properties and Functionalities. Int. J. Food Microbiol. 2016, 239, 26–34. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Van Kerrebroeck, S.; Harth, H.; Huys, G.; Daniel, H.M.; Weckx, S. Microbial Ecology of Sourdough Fermentations: Diverse or uniform? Food Microbiol. 2014, 37, 11–29. [Google Scholar] [CrossRef] [PubMed]
- Aydın, F.; Özer, G.; Alkan, M.; Çakır, İ. Start Codon Targeted (SCoT) Markers for the Assessment of Genetic Diversity in Yeast Isolated from Turkish Sourdough. Food Microbiol. 2022, 107, 104081. [Google Scholar] [CrossRef]
- Boyaci-Gunduz, C.P.; Erten, H. Predominant Yeasts in the Sourdoughs Collected from Some Parts of Turkey. Yeast 2020, 37, 449–466. [Google Scholar] [CrossRef]
- Pino, A.; Russo, N.; Solieri, L.; Sola, L.; Caggia, C.; Randazzo, C.L. Microbial Consortia Involved in Traditional Sicilian Sourdough: Characterization of Lactic Acid Bacteria and Yeast Populations. Microorganisms 2022, 10, 283. [Google Scholar] [CrossRef] [PubMed]
- Valmorri, S.; Tofalo, R.; Settanni, L.; Corsetti, A.; Suzzi, G. Yeast Microbiota Associated with Spontaneous Sourdough Fermentations in the Production of Traditional Wheat Sourdough Breads of the Abruzzo Region (Italy). Antonie Leeuwenhoek 2010, 97, 119–129. [Google Scholar] [CrossRef]
- Yang, H.; Liu, T.; Zhang, G.; He, G. Intraspecific Diversity and Fermentative Properties of Saccharomyces cerevisiae from Chinese Traditional Sourdough. LWT-Food Sci. Technol. 2020, 124, 109195. [Google Scholar] [CrossRef]
- Arora, K.; Ameur, H.; Polo, A.; Di Cagno, R.; Rizzello, C.G.; Gobbetti, M. Thirty Years of Knowledge on Sourdough Fermentation: A Systematic Review. Trends Food Sci. Technol. 2021, 108, 71–83. [Google Scholar] [CrossRef]
- Birch, A.N.; Petersen, M.A.; Arneborg, N.; Hansen, A.S. Influence of Commercial Baker’s Yeasts on Bread Aroma Profiles. Food Res. Int. 2013, 52, 160–166. [Google Scholar] [CrossRef]
- Lampignano, V.; Laverse, J.; Mastromatteo, M.; Del Nobile, M.A. Microstructure, Textural and Sensorial Properties of Durum Wheat Bread as Affected by Yeast Content. Food Res. Int. 2013, 50, 369–376. [Google Scholar] [CrossRef]
- Huys, G.; Daniel, H.-M.; De Vuyst, L. Taxonomy and Biodiversity of Sourdough Yeasts and Lactic Acid Bacteria. In Handbook on Sourdough Biotechnology; Springer: New York, NY, USA, 2013; pp. 105–154. [Google Scholar] [CrossRef]
- Aydın, F.; Özer, G.; Alkan, M.; Çakır, İ. The Utility of iPBS Retrotransposons Markers to Analyze Genetic Variation in Yeast. Int. J. Food Microbiol. 2020, 325, 108647. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Gómez, F.; Romero-Gil, V.; Bautista-Gallego, J.; Garrido-Fernández, A.; Arroyo-López, F.N. Multivariate Analysis to Discriminate Yeast Strains with Technological Applications in Table Olive Processing. World J. Microbiol. Biotechnol. 2012, 28, 1761–1770. [Google Scholar] [CrossRef] [PubMed]
- Bhagyawant, S.S. RAPD-SCAR markers: An Interface Tool for Authentication of Traits. J. Biosci. Med. 2016, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Luan, C.; Li, X.; Zheng, G.; Yao, J.; Wang, J. ISSR Fingerprint Analysis and SCAR Marker of 23 Strains of Saccharomyces cerevisiae. Food Sci. 2014, 35, 163–167. [Google Scholar] [CrossRef]
- Palla, M.; Blandino, M.; Grassi, A.; Giordano, D.; Sgherri, C.; Quartacci, M.F.; Reyneri, A.; Agnolucci, M.; Giovannetti, M. Characterization and Selection of Functional Yeast Strains during Sourdough Fermentation of Different Cereal Wholegrain Flours. Sci. Rep. 2020, 10, 12856. [Google Scholar] [CrossRef]
- Liu, J.; Li, X.; Liu, Y.; Xing, C.; Xie, Y.; Cai, G.; Lu, J. Evaluation of Genetic Diversity and Development of Core Collections of Industrial Brewing Yeast Using ISSR Markers. Arch. Microbiol. 2021, 203, 1001–1008. [Google Scholar] [CrossRef]
- Aydın, F.; Özer, G.; Alkan, M.; Çakır, İ. Genetic Diversity and Population Structure of Saccharomyces cerevisiae Isolated from Turkish Sourdough by iPBS-Retrotransposons Markers. Arch. Microbiol. 2022, 204, 693. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.M.; Sarla, N.; Siddiq, E.A. Inter Simple Sequence Repeat (ISSR) Polymorphism and its Application in Plant Breeding. Euphytica 2002, 128, 9–17. [Google Scholar] [CrossRef]
- Gallardo, G.; Ruiz-Moyano, S.; Hernández, A.; Benito, M.J.; Córdoba, M.G.; Pérez-Nevado, F.; Martín, A. Application of ISSR-PCR for Rapid Strain Typing of Debaryomyces hansenii Isolated from Dry-Cured Iberian Ham. Food Microbiol. 2014, 42, 205–211. [Google Scholar] [CrossRef]
- Agarbati, A.; Marini, E.; Galli, E.; Canonico, L.; Ciani, M.; Comitini, F. Characterization of Wild Yeasts Isolated from Artisan Dairies in the Marche Region, Italy, for Selection of Promising Functional Starters. LWT-Food Sci. Technol. 2021, 139, 110531. [Google Scholar] [CrossRef]
- Merchán, A.V.; Benito, M.J.; Galván, A.I.; de Herrera, S.R.M.S. Identification and Selection of Yeast with Functional Properties for Future Application in Soft Paste Cheese. LWT-Food Sci. Technol. 2020, 124, 109173. [Google Scholar] [CrossRef]
- Nova, M.X.V.; Schuler, A.R.P.; Brasileiro, B.T.R.V.; Morais Jr, M.A. Yeast Species Involved in Artisanal Cachaça Fermentation in Three Stills with Different Technological Levels in Pernambuco, Brazil. Food Microbiol. 2009, 26, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Moyano, S.; Esperilla, A.; Hernández, A.; Benito, M.J.; Casquete, R.; Martín-Vertedor, D.; Pérez-Nevado, F. Application of ISSR-PCR as a Rapid Method for Clustering and Typing of Yeasts Isolated from Table Olives. LWT-Food Sci. Technol. 2019, 109, 250–254. [Google Scholar] [CrossRef]
- Hammadi, S.Y.; Hussein, A.S.; Majeed, D.M.; Dheeb, B.I.; Ismail, E.N. RAPD and ISSR Analyses of Saccharomyces cerevisiae Isolates from Different Sources. J. Biotech. Res. Center. 2018, 12, 40–50. [Google Scholar] [CrossRef]
- Pathania, N.; Kanwar, S.S.; Jhang, T.; Koundal, K.R.; Sharma, T.R. Application of Different Molecular Techniques for Deciphering Genetic Diversity among Yeast Isolates of Traditional Fermented Food Products of Western Himalayas. World J. Microbiol. Biotechnol. 2010, 26, 1539–1547. [Google Scholar] [CrossRef]
- Josepa, S.; Guillamon, J.M.; Cano, J. PCR Differentiation of Saccharomyces cerevisiae from Saccharomyces bayanus/Saccharomyces pastorianus Using Specific Primers. FEMS Microbiol. Lett. 2000, 193, 255–259. [Google Scholar] [CrossRef]
- Prevost, A.; Wilkinson, M.J. A New System of Comparing PCR Primers Applied to ISSR Fingerprinting of Potato Cultivars. Theor. Appl. Genet. 1999, 98, 107–112. [Google Scholar] [CrossRef]
- Roldàn-Ruiz, I.; Dendauw, J.; Van Bockstaele, E.; Depicker, A.; De Loose, M. AFLP Markers Reveal High Polymorphic Rates in Ryegrasses (Lolium spp.). Mol. Breed. 2000, 6, 125–134. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H.; et al. Package ‘Vegan’. Community Ecol. Package Version 2013, 2, 1–295. [Google Scholar]
- RStudio Team. RStudio: Integrated Development for R; RStudio: Boston, MA, USA, 2020. [Google Scholar]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research-an Update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Yeh, F.; Yang, R.-C.; Boyle, T. POPGENE Version1. 32, Microsoft Window-Base Software for Population Genetic Analysis: A Quick User’s Guide. University of Alberta; Center for International Forestry Research: Edmonton, AB, Canada, 1999. [Google Scholar]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of Molecular Variance Inferred from Metric Distances Among DNA Haplotypes: Application to Human Mitochondrial DNA Restriction Data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef]
- Earl, D.A. Structure Harvester: A Website and Program for Visualizing STRUCTURE Output and Implementing the Evanno Method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the Number of Clusters of Individuals Using the Software Structure: A Simulation Study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Bevilacqua, A.; Perricone, M.; Cannarsi, M.; Corbo, M.R.; Sinigaglia, M. Technological and Spoiling Characteristics of the Yeast Microflora Isolated from Bella di Cerignola Table Olives. Int. J. Food Sci. Technol. 2009, 44, 2198–2207. [Google Scholar] [CrossRef]
- Perricone, M.; Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M. Technological Characterization and Probiotic Traits of Yeasts Isolated from Altamura Sourdough to Select Promising Microorganisms as Functional Starter Cultures for Cereal-Based Products. Food Microbiol. 2014, 38, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Coello, M.S.; Perez, A.B.; Iranzo, J.U.; Alvarez, P.M. Characteristics of Wines Fermented with Different Saccharomyces cerevisiae Strains Isolated from the La Mancha Region. Food Microbiol. 1999, 16, 563–573. [Google Scholar] [CrossRef]
- Jolliffe, I.T. Principal Component Analysis for Special Types of data. In Principal Component Analysis; Springer: New York, NY, USA, 2002; pp. 338–372. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for Association Mapping of Complex Traits in Diverse Samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Shi, A.; Buckley, B.; Mou, B.; Motes, D.; Morris, J.B.; Ma, J.; Xiong, H.; Qin, J.; Yang, W.; Chitwood, J.; et al. Association Analysis of Cowpea Bacterial Blight Resistance in USDA Cowpea Germplasm. Euphytica 2016, 208, 143–155. [Google Scholar] [CrossRef]
- Lattanzi, A.; Minervini, F.; Di Cagno, R.; Diviccaro, A.; Antonielli, L.; Cardinali, G.; Cappelle, S.; De Angelis, M.; Gobbetti, M. The Lactic Acid Bacteria and Yeast Microbiota of Eighteen Sourdoughs Used for the Manufacture of Traditional Italian Sweet Leavened Baked Goods. Int. J. Food Microbiol. 2013, 163, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Minervini, F.; Di Cagno, R.; Lattanzi, A.; De Angelis, M.; Antonielli, L.; Cardinali, G.; Cappelle, S.; Gobbetti, M. Lactic Acid Bacterium and Yeast Microbiotas of 19 Sourdoughs Used for Traditional/Typical Italian Breads: Interactions Between Ingredients and Microbial Species Diversity. Appl. Environ. Microbiol. 2012, 78, 1251–1264. [Google Scholar] [CrossRef] [Green Version]
- Vigentini, I.; Antoniani, D.; Roscini, L.; Comasio, A.; Galafassi, S.; Picozzi, C.; Corte, L.; Compagno, C.; Dal Bello, F.; Cardinali, G.; et al. Candida milleri Species Reveals Intraspecific Genetic and Metabolic Polymorphisms. Food Microbiol. 2014, 42, 72–81. [Google Scholar] [CrossRef]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a Genetic Linkage Map in Man Using Restriction Fragment Length Polymorphisms. Am. J. Hum. Genet. 1980, 32, 314. [Google Scholar] [PubMed]
- Barut, M.; Nadeem, M.A.; Karaköy, T.; Baloch, F.S. DNA Fingerprinting and Genetic Diversity Analysis of World Quinoa Germplasm Using iPBS-Retrotransposon Marker System. Turk. J. Agric. For. 2020, 44, 479–491. [Google Scholar] [CrossRef]
- Bigey, F.; Segond, D.; Friedrich, A.; Guezenec, S.; Bourgais, A.; Huyghe, L.; Aiger, N.; Nidelet, T.; Sicard, D. Evidence for Two Main Domestication Trajectories in Saccharomyces cerevisiae Linked to Distinct Bread-Making Processes. Curr. Biol. 2021, 31, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Börlin, M.; Venet, P.; Claisse, O.; Salin, F.; Legras, J.L.; Masneuf-Pomarede, I. Cellar-Associated Saccharomyces cerevisiae Population Structure Revealed High-Level Diversity and Perennial Persistence at Sauternes Wine Estates. Appl. Environ. Microbiol. 2016, 82, 2909–2918. [Google Scholar] [CrossRef] [Green Version]
- El Mousadik, A.; Petit, R.J. Chloroplast DNA Phylogeography of the Argan Tree of Morocco. Mol. Ecol. 1996, 5, 547–555. [Google Scholar] [CrossRef]
- Gayevskiy, V.; Klaere, S.; Knight, S.; Goddard, M.R. ObStruct: A Method to Objectively Analyse Factors Driving Population Structure Using Bayesian Ancestry Profiles. PLoS ONE 2014, 9, e85196. [Google Scholar] [CrossRef] [Green Version]
- Steensels, J.; Snoek, T.; Meersman, E.; Nicolino, M.P.; Voordeckers, K.; Verstrepen, K.J. Improving Industrial Yeast Strains: Exploiting Natural and Artificial Diversity. FEMS Microbiol. Rev. 2014, 38, 947–995. [Google Scholar] [CrossRef] [Green Version]
- Andrade, M.J.; Rodríguez, M.; Casado, E.M.; Bermúdez, E.; Córdoba, J.J. Differentiation of Yeasts Growing on Dry-Cured Iberian Ham by Mitochondrial DNA Restriction Analysis, RAPD-PCR, and Their Volatile Compounds Production. Food Microbiol. 2009, 26, 578–586. [Google Scholar] [CrossRef]
- Martínez, C.; Cosgaya, P.; Vásquez, C.; Gac, S.; Ganga, A. High Degree of Correlation Between Molecular Polymorphism and Geographic Origin of Wine Yeast Strains. J. Appl. Microbiol. 2007, 103, 2185–2195. [Google Scholar] [CrossRef]
- Nikolaou, E.; Andrighetto, C.; Lombardi, A.; Litopoulou-Tzanetaki, E.; Tzanetakis, N. Heterogeneity in Genetic and Phenotypic Characteristics of Saccharomyces cerevisiae Strains Isolated from Red and White Wine Fermentations. Food Control 2007, 18, 1458–1465. [Google Scholar] [CrossRef]
- Palla, M.; Agnolucci, M.; Calzone, A.; Giovannetti, M.; Di Cagno, R.; Gobbetti, M.; Rizzello, C.G.; Pontonio, E. Exploitation of Autochthonous Tuscan Sourdough Yeasts as Potential Starters. Int. J. Food Microbiol. 2019, 302, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Pulvirenti, A.; Rainieri, S.; Boveri, S.; Giudici, P. Optimizing the Selection Process of Yeast Starter Cultures by Preselecting Strains Dominating Spontaneous Fermentations. Can. J. Microbiol. 2009, 55, 326–332. [Google Scholar] [CrossRef]
- Arici, M.; Ozulku, G.; Yildirim, R.M.; Sagdic, O.; Durak, M.Z. Biodiversity and technological Properties of Yeasts from Turkish Sourdough. Food Sci. Biotechnol. 2018, 27, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Heitmann, M.; Zannini, E.; Arendt, E. Impact of Saccharomyces cerevisiae Metabolites Produced During Fermentation on Bread Quality Parameters: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1152–1164. [Google Scholar] [CrossRef] [PubMed]
- Gabriela, C.G.; Daniela, V. The Influence of Different forms of Backery Yeast Saccharomyces cerevisie Type Strain on the Concentration of Individual Sugars and their Utilization during Fermentation. Rom. Biotechnol. Lett. 2010, 15, 5418. [Google Scholar]
- Tokashiki, T.; Yamamoto, H.; Watanabe, H.; Nakajima, R.; Shima, J. A Functional Compound Contained in Sugar Cane Molasses Enhances the Fermentation Ability of Baker’s Yeast in High-Sugar Dough. J. Gen. Appl. Microbiol. 2011, 57, 303–307. [Google Scholar] [CrossRef] [Green Version]
- Altay, F.; Karbancıoglu-Güler, F.; Daskaya-Dikmen, C.; Heperkan, D. A Review on Traditional Turkish Fermented Non-Alcoholic Beverages: Microbiota, Fermentation Process and Quality Characteristics. Int. J. Food Microbiol. 2013, 167, 44–56. [Google Scholar] [CrossRef] [Green Version]
- Kahve, H.I.; Akbulut, M.; Coklar, H. Identification and Technological Characterization of Endogenous Yeast Isolated from Fermented Black Carrot Juice, Shalgam. LWT-Food Sci. Technol. 2022, 154, 112823. [Google Scholar] [CrossRef]
- Kamali, M.; Samsampour, D.; Bagheri, A.; Mehrafarin, A.; Homaei, A. Association Analysis and Evaluation of Genetic Diversity of Teucrium stocksianum Boiss. Populations Using ISSR Markers. Genet. Resour. Crop Evol. 2023, 70, 691–709. [Google Scholar] [CrossRef]
- Mohammadi, A.R.; Darvishzadeh, R.; Alijanpour, A.; Razi, M. Toward the Identification of Molecular Markers Associated with Phytochemical Traits in the Iranian Sumac (Rhus coriaria L.) Population. Food Sci. Nutr. 2021, 9, 3142–3154. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ISSR ID | Primer Sequence (5′–3′) | Tm (°C) | TB | PB | PIC | RP |
|---|---|---|---|---|---|---|
| ISSR1 | ARRTYCAGCAGCAGCAG | 50.0 | 29 | 23 | 0.66 | 2.72 |
| ISSR2 | GTGGTGGTGGTGGTG | 55.0 | 11 | 5 | 0.76 | 0.27 |
| ISSR3 | AGAGAGAGAGAGAAGAGAGT | 50.4 | 9 | 6 | 0.58 | 0.73 |
| ISSR4 | HVHTGTGTGTGTGTG | 45.6 | 30 | 25 | 0.79 | 3.25 |
| ISSR5 | AGAGAGAGAGAGAGAGVC | 50.8 | 25 | 23 | 0.44 | 1.46 |
| ISSR6 | ACACACACACACAGACYG | 54.0 | 10 | 7 | 0.59 | 0.86 |
| ISSR7 | AGAGAGAGAGAGAGAGYG | 51.4 | 23 | 20 | 0.74 | 2.16 |
| ISSR8 | GACAGACAGACAGACA | 52.0 | 17 | 14 | 0.58 | 1.31 |
| Quantitative Analyses a | Codes | |||
|---|---|---|---|---|
| b GI < 25 | FCx < 3.75 | FCy < 0.80 | VI = 0 | 0 |
| 25 < GI < 75 | 3.75 < FCx < 4.0 | 0.80 < FCy < 1.00 | VI < 50 | 1 |
| GI > 75 | FCx > 4.0 | FCy > 1.0 | VI > 50 | 2 |
| Population | Na ** | Ne | I | h | PPL (%) |
|---|---|---|---|---|---|
| CAR * (n = 32) | 1.60 ± 0.05 | 1.37 ± 0.03 | 0.33 ± 0.02 | 0.22 ± 0.01 | 69.48 |
| BSR (n = 32) | 1.56 ± 0.06 | 1.40 ± 0.03 | 0.33 ± 0.02 | 0.22 ± 0.01 | 68.18 |
| AER (n = 32) | 1.32 ± 0.06 | 1.31 ± 0.03 | 0.28 ± 0.02 | 0.19 ± 0.01 | 53.25 |
| Average (n = 96) | 1.50 ± 0.03 | 1.36 ± 0.02 | 0.31 ± 0.01 | 0.21 ± 0.01 | 63.64 ± 5.21 |
| Source | d-f | SS | MS | Est. var. | % | FST | p |
|---|---|---|---|---|---|---|---|
| AP * | 2 | 230.375 | 115.188 | 3.075 | 15 | 0.155 | 0.001 |
| AIWP | 93 | 1562.500 | 16.801 | 16.801 | 85 | ||
| Total | 95 | 1792.875 | 19.876 | 100 |
| No | a Temperature (°C) | NaCl | pH | LAR (%) | AAR (%) | FCx | FCy | Lev | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 10 | 37 | 4 | 6 | 2.5 | 3.0 | 3.5 | 0.60 | 1.20 | 0.15 | 0.30 | ||||
| Y2 | 25–75 | >75 | 25–75 | 25–75 | <25 | 25–75 | >75 | >75 | >75 | >75 | >75 | >4.0 | >1.0 | >50 |
| Y6 | 25–75 | 25–75 | 25–75 | 25–75 | <25 | 25–75 | >75 | >75 | 25–75 | >75 | 25–75 | 3.5–3.75 | <0.80 | <50 |
| Y10 | 25–75 | 25–75 | 25–75 | <25 | <25 | 25–75 | >75 | >75 | >75 | >75 | >75 | 3.5–3.75 | 0.80–1.00 | <50 |
| Y19 | 25–75 | 25–75 | >75 | 25–75 | <25 | 25–75 | >75 | >75 | >75 | >75 | >75 | 3.75–4.0 | 0.80–1.00 | >50 |
| Y22 | 25–75 | 25–75 | 25–75 | 25–75 | <25 | 25–75 | >75 | >75 | 25–75 | >75 | >75 | >4.0 | >1.0 | >50 |
| Y26 | 25–75 | >75 | 25–75 | <25 | <25 | 25–75 | >75 | >75 | >75 | >75 | 25–75 | 3.75–4.0 | >1.0 | >50 |
| Y27 | 25–75 | 25–75 | >75 | 25–75 | <25 | 25–75 | >75 | >75 | >75 | >75 | >75 | >4.0 | >1.0 | >50 |
| Y30 | 25–75 | >75 | >75 | 25–75 | <25 | 25–75 | >75 | >75 | 25–75 | 25–75 | 25–75 | >4.0 | >1.0 | >50 |
| Y32 | 25–75 | 25–75 | 25–75 | 25–75 | <25 | 25–75 | >75 | >75 | >75 | >75 | >75 | 3.5–3.75 | <0.80 | <50 |
| Y36 | >75 | 25–75 | 25–75 | 25–75 | <25 | 25–75 | >75 | >75 | >75 | >75 | >75 | >4.0 | 0.80–1.00 | >50 |
| Y40 | 25–75 | >75 | 25–75 | <25 | <25 | 25–75 | >75 | >75 | >75 | 25–75 | 25–75 | 3.75–4.0 | >1.0 | >50 |
| Y41 | 25–75 | 25–75 | 25–75 | <25 | <25 | 25–75 | >75 | >75 | >75 | >75 | 25–75 | >4.0 | 0.80–1.00 | >50 |
| Y43 | 25–75 | >75 | 25–75 | <25 | <25 | 25–75 | 25–75 | 25–75 | 25–75 | >75 | 25–75 | 3.75–4.0 | >1.0 | >50 |
| Y46 | 25–75 | >75 | 25–75 | <25 | <25 | 25–75 | >75 | >75 | >75 | >75 | >75 | >4.0 | >1.0 | >50 |
| Y47 | 25–75 | 25–75 | >75 | 25–75 | <25 | 25–75 | >75 | >75 | >75 | >75 | >75 | 3.5–3.75 | <0.80 | <50 |
| Y49 | 25–75 | >75 | 25–75 | <25 | <25 | 25–75 | >75 | >75 | >75 | >75 | 25–75 | 3.5–3.75 | 0.80–1.00 | <50 |
| Y50 | 25–75 | >75 | >75 | 25–75 | 25–75 | >75 | >75 | >75 | >75 | >75 | >75 | >4.0 | >1.0 | >50 |
| Y51 | >75 | 25–75 | >75 | 25–75 | <25 | 25–75 | >75 | >75 | >75 | >75 | >75 | >4.0 | 0.80–1.00 | >50 |
| Y53 | >75 | >75 | >75 | 25–75 | 25–75 | >75 | >75 | >75 | >75 | >75 | >75 | 3.5–3.75 | 0.80–1.00 | <50 |
| Y56 | 25–75 | >75 | 25–75 | 25–75 | <25 | 25–75 | >75 | >75 | >75 | 25–75 | 25–75 | >4.0 | >1.0 | >50 |
| Y59 | >75 | >75 | 25–75 | 25–75 | <25 | 25–75 | >75 | >75 | >75 | >75 | 25–75 | >4.0 | >1.0 | >50 |
| Y64 | 25–75 | >75 | 25–75 | 25–75 | 25–75 | >75 | >75 | >75 | >75 | >75 | >75 | >4.0 | >1.0 | >50 |
| Y65 | >75 | >75 | >75 | 25–75 | 25–75 | 25–75 | >75 | >75 | >75 | >75 | >75 | 3.5–3.75 | 0.80–1.00 | >50 |
| Y70 | 25–75 | 25–75 | 25–75 | <25 | <25 | 25–75 | 25–75 | 25–75 | 25–75 | >75 | >75 | >4.0 | 0.80–1.00 | >50 |
| Y72 | >75 | >75 | >75 | 25–75 | 25–75 | >75 | >75 | >75 | 25–75 | >75 | >75 | >4.0 | >1.0 | >50 |
| Y80 | 25–75 | 25–75 | 25–75 | <25 | <25 | 25–75 | >75 | >75 | >75 | >75 | >75 | >4.0 | 0.80–1.00 | >50 |
| Y85 | >75 | >75 | >75 | 25–75 | 25–75 | >75 | >75 | >75 | >75 | >75 | >75 | >4.0 | >1.0 | >50 |
| Y91 | 25–75 | >75 | 25–75 | <25 | <25 | 25–75 | 25–75 | >75 | 25–75 | 25–75 | 25–75 | >4.0 | >1.0 | >50 |
| Y94 | 25–75 | 25–75 | 25–75 | 25–75 | <25 | 25–75 | >75 | >75 | >75 | >75 | >75 | >4.0 | 0.80–1.00 | >50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aydın, F.; Günen, T.U.; Kahve, H.İ.; Güler, E.; Özer, G.; Aktepe, Y.; Çakır, İ. Molecular and Technological Characterization of Saccharomyces cerevisiae from Sourdough. Fermentation 2023, 9, 329. https://doi.org/10.3390/fermentation9040329
Aydın F, Günen TU, Kahve Hİ, Güler E, Özer G, Aktepe Y, Çakır İ. Molecular and Technological Characterization of Saccharomyces cerevisiae from Sourdough. Fermentation. 2023; 9(4):329. https://doi.org/10.3390/fermentation9040329
Chicago/Turabian StyleAydın, Furkan, Tacettin Utku Günen, Halil İbrahim Kahve, Emrah Güler, Göksel Özer, Yeşim Aktepe, and İbrahim Çakır. 2023. "Molecular and Technological Characterization of Saccharomyces cerevisiae from Sourdough" Fermentation 9, no. 4: 329. https://doi.org/10.3390/fermentation9040329