
Combined Ensiling of Tropical Beans and Sugarcane Stalks: Effects on Their Secondary Metabolites

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Procedure to Prepare Laboratory Silage
2.3. Sampling and Chemical Analysis of Fresh and Ensiled Forage
2.4. Proximate Chemical Analysis
2.5. Plant Secondary Metabolites
- -
- Trypsin inhibitor
- -
- L-canavanine
- -
- Total tannins
- -
- Phytic acid
- -
- L-Dopa
2.6. Calculations
2.7. In Vitro Cellulase Digestibility of Organic Matter
2.8. Statistical Analysis
3. Results
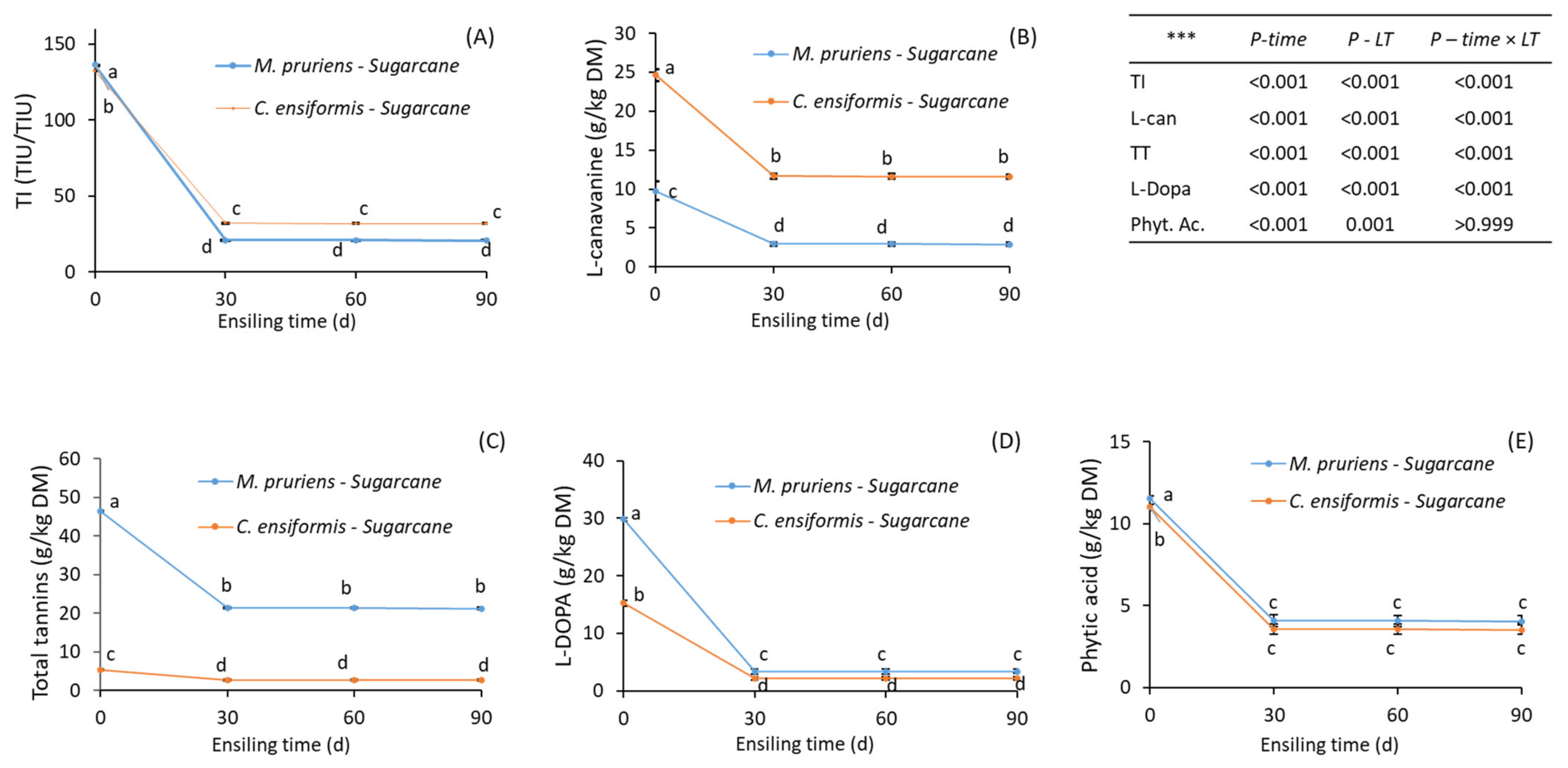
3.1. Chemical Characteristics and Plant Secondary Metabolites (PSM) of Fresh and Ensiled Feed
Ensilability
3.2. In Vitro Cellulase Digestibility of Organic Matter
4. Discussion
4.1. Effect of Ensiling Process on the Ensilability
4.2. Effect of Ensiling Process on the Chemical Composition and Digestibility
4.3. Effect of Ensiling Process on the PSM Concentrations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Ojeda, F. Harvesting and ensiling techniques. In Proceedings of the FAO Electronic Conference on Tropical Silage, Silage Making in the Tropics with Particular Emphasis on Smallholders, Rome, Italy, 1 September–15 December 2000; pp. 133–140. [Google Scholar]
- Lima-Orozco, R.; Van Daele, I.; Álvarez-Hernández, U.; Fievez, V. Combination of the underutilised legumes Canavalia ensiformis (L.) D.C. and Mucuna pruriens (L.) D.C. with sorghum: Integrated assessment of their potential as conserved ruminant feed. Cuba. J. Agric. Sci. 2016, 50, 99–103. [Google Scholar]
- Lima-Orozco, R.; Arce-González, M.A.; Bello-Morales, I.; Artiles-Ortega, E.; Fievez, V. Assessment of quality and rumen degradability of mixed silages of sugarcane tops with whole plants of Tithonia diversifolia (Hemsl.) Gray in combination with molasses, fungi and lactobacilli. Adv. Anim. Biosci. 2018, 8, 519. [Google Scholar]
- Lima Orozco, R.; Bello Morales, I.; Arce González, M.A.; Artiles Ortega, E.; Fievez, V. Assessment of quality and rumen degradability of mixed silages of sugarcane tops with Marabú forage. In Proceedings of the TROPENTAG 2019: Conference on International Research on Food Security, Natural Resource Management and Rural Development, Kassel, Germany, 18–20 September 2019; p. 211. [Google Scholar]
- Clavero, T.; Razz, R. The performance of goats browsing Leucaena leucocephala in the semi arid areas of northwest Venezuela. Rev. Cient. Fac. Cienc. Vet. Univ. Zulia 2003, 13, 460–463. [Google Scholar]
- Lima-Orozco, R.; Castro-Alegría, A.; Fievez, V. Ensiled sorghum and soybean as ruminant feed in the tropics, with emphasis on Cuba. Grass Forage Sci. 2013, 68, 20–32. [Google Scholar] [CrossRef]
- Lima-Orozco, R.; Van Daele, I.; Álvarez-Hernández, U.; Fievez, V. Combined conservation of jack bean and velvet bean with sorghum: Evaluation of lab-scale silages and in vitro assessment of their nutritive value. J. Agric. Sci. 2014, 152, 967–980. [Google Scholar] [CrossRef]
- Díaz, M.F.; González, A.; Padilla, C.; Curbelo, F. Comportamiento de la producción de forrajes y granos de Canavalia ensiformis, Lablab purpureus y Stizolobium niveum en siembras de septiembre. Rev. Cuba. Cienc. Agrícola 2003, 37, 65–71. [Google Scholar]
- Kambashi, B.; Picron, P.; Boudry, C.; Théwis, A.; Kiatoko, H.; Bindelle, J. Nutritive value of tropical forage plants fed to pigs in the Western provinces of the Democratic Republic of the Congo. Anim. Feed. Sci. Technol. 2014, 191, 47–56. [Google Scholar] [CrossRef]
- González Díaz, L.A.; Hoedtke, S.; Castro, A.; Zeynerb, A. In vitro ensilability of Jack bean (Canavalia ensiformis) and Cowpea (Vigna unguiculata) grains sole or mixed with sorghum (Sorghum bicolor) grains: An alternative for low input pig feeding systems. In Proceedings of the TROPENTAG “Conference on International Research on Food Security, Natural Resource Management and Rural Development”, Hamburg, Germany, 6–8 October 2009. [Google Scholar]
- Hai, P.V.; Everts, H.; Tien, D.V.; Schonewille, J.T.; Hendriks, W.H. Feeding Chromonaela odorata during pregnancy to goat dams affects acceptance of this feedstuff by their offspring. Appl. Anim. Behav. Sci. 2012, 137, 30–35. [Google Scholar] [CrossRef]
- Kalač, P.; Price, K.R.; Fenwick, G.R. Changes in saponin content and composition during the ensilage of alfalfa (Medicago sativa L.). Food Chem. 1996, 56, 377–380. [Google Scholar] [CrossRef]
- Abbasi, M.; Rouzbehan, Y.; Rezaei, J.; Jacobsen, S.-E. The effect of lactic acid bacteria inoculation, molasses, or wilting on the fermentation quality and nutritive value of amaranth (Amaranthus hypochondriaus) silage. J. Anim. Sci. 2018, 96, 3983–3992. [Google Scholar] [CrossRef] [Green Version]
- Fan, Z.; Chen, K.; Ban, L.; Mao, Y.; Hou, C.; Li, J. Silage fermentation: A potential biological approach for the long-term preservation and recycling of polyphenols and terpenes in Globe artichoke (Cynara scolymus L.) by-products. Molecules 2020, 25, 3302. [Google Scholar] [CrossRef] [PubMed]
- Titterton, M.; Bareeba, F. Grass and Legume Silages in the Tropics; ‘t Mannetje, L., Ed.; FAO Plant Production and Protection Paper; Food and Agriculture Organization (FAO): Rome, Italy, 2000; pp. 43–50. [Google Scholar]
- Jaurena, G.; Pichard, G. Contribution of storage and structural polysaccharides to the fermentation process and nutritive value of lucerne ensiled alone or mixed with cereal grains. Anim. Feed. Sci. Technol. 2001, 92, 159–173. [Google Scholar] [CrossRef]
- Lima, R.; Díaz, R.F.; Castro, A.; Hoedtke, S.; Fievez, V. Multifactorial models to assess responses to sorghum proportion, molasses and bacterial inoculant on in vitro quality of sorghum–soybean silages. Anim. Feed. Sci. Technol. 2011, 164, 161–173. [Google Scholar] [CrossRef]
- Ajayi, F.; Akande, S.; Adegbite, A.; Idowu, B. Assessment of seven under-utilized grain legume foliages as feed resources for ruminants. Livest. Res. Rural Dev. 2009, 21, 149–156. [Google Scholar]
- Chikagwa-Malunga, S.K.; Adesogan, A.T.; Sollenberger, L.E.; Badinga, L.K.; Szabo, N.J.; Littell, R.C. Nutritional characterization of Mucuna pruriens: 1. Effect of maturity on the nutritional quality of botanical fractions and the whole plant. Anim. Feed. Sci. Technol. 2009, 148, 34–50. [Google Scholar] [CrossRef]
- Suárez-Benítez, O.J.; Jorge-Suarez, H.; Delgado-Mora, I.; Casanova-Cosió, E.; Álvarez-González, Y.; Moreno-Lorenzo, X.; González-Rodríguez, J.M.; González-Marrero, A.N. Composición química y fenológica de ocho cultivares de caña de azúcar (Saccharum sp.) para su recomendación en la alimentación de rumiantes. ICIDCA Sobre Deriv. Caña Azúcar 2018, 52, 7–12. [Google Scholar]
- McCune, N.M.; González, Y.R.; Alcántara, E.A.; Martínez, O.F.; Fundora, C.O.; Arzola, N.C.; Cairo, P.C.; D’Haese, M.; DeNeve, S.; Hernández, F.G. Global questions, local answers: Soil management and sustainable intensification in diverse socioeconomic contexts of cuba. J. Sustain. Agric. 2011, 35, 650–670. [Google Scholar] [CrossRef]
- Lima Orozco, R.; González Díaz, L.A. Lecture: Ensiling as method to reduce Antinutritional factors (ANFs). Master of Nutrition and Rural Development, 2010.
- Marin-Cárdenas, A.; García, A.; Herrera, N.; Gómez, L. Registro de Marca BIOPRANAL y Diseño para Distinguir Productos en las Clases 5 y 31. Solicitud de Marca # 2004-0226. Resolución # 750/2005. 22 February 2005. [Google Scholar]
- Lima, R.; Lourenço, M.; Díaz, R.F.; Castro, A.; Fievez, V. Effect of combined ensiling of sorghum and soybean with or without molasses and lactobacilli on silage quality and in vitro rumen fermentation. Anim. Feed. Sci. Technol. 2010, 155, 122–131. [Google Scholar] [CrossRef]
- Broderick, G.A.; Kang, J.H. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef]
- Castro-Montoya, J.; De Campeneere, S.; Van Ranst, G.; Fievez, V. Interactions between methane mitigation additives and basal substrates on in vitro methane and VFA production. Anim. Feed. Sci. Technol. 2012, 176, 47–60. [Google Scholar] [CrossRef]
- A.O.A.C. Official Methods of Analysis of AOAC International, 18th ed.; Horwitz, W., Ed.; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- EC. European Union Official Diary. Reglament (CE) no152/2009. 2009; 54. [Google Scholar]
- Al-Rabadi, G.J.S.; Gilbert, R.G.; Gidley, M.J. Effect of particle size on kinetics of starch digestion in milled barley and sorghum grains by porcine alpha-amylase. J. Cereal Sci. 2009, 50, 198–204. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Makkar, H.P.S.; Siddhuraju, P.; Becker, K. Plant Secondary Metabolites; Humana Press: Totowa, NJ, USA, 2007. [Google Scholar]
- Doss, A.; Pugalenthi, M.; Vadivel, V.; Subhashini, G.; Subash, A.R. Effects of processing technique on the nutritional composition and antinutrients content of under—Utilized food legume Canavalia ensiformis L. DC. Int. Food Res. J. 2011, 18, 965–970. [Google Scholar]
- De Boever, J.L.; Cottyn, B.G.; Buysse, F.X.; Wainman, F.W.; Vanacker, J.M. The use of an enzymatic technique to predict digestibility, metabolizable and net energy of compound feedstuffs for ruminants. Anim. Feed. Sci. Technol. 1986, 14, 203–214. [Google Scholar] [CrossRef]
- SPSS. Software for Windows, Release 21.0, Inc.; Property of IBM Corporation: Chicago, IL, USA, 2012. [Google Scholar]
- Tukey, J.W. Comparing Individual Means in the Analysis of Variance. Biometrics 1949, 5, 99–114. [Google Scholar] [CrossRef]
- Ojeda, F.; Cáceres, O.; Esperance, M. Sistema de evaluación para ensilajes tropicales. In Conservación de Forrajes; López Tapanes, R.V., Pereira Pérez, N., Loidi Ramos, R., Pérez Navarrete, A., Eds.; Pueblo y Educación: Ciudad de La Habana, Cuba, 1991; pp. 15–65. [Google Scholar]
- Umaña, R.; Staples, C.R.; Bates, D.B.; Wilcox, C.J.; Mahanna, W.C. Effects of a microbial inoculant and(or) sugarcane molasses on the fermentation, aerobic stability, and digestibility of bermudagrass ensiled at two moisture contents. J. Anim. Sci. 1991, 69, 4588–4601. [Google Scholar] [CrossRef] [Green Version]
- Filya, I.; Ashbell, G.; Hen, Y.; Weinberg, Z.G. The effect of bacterial inoculants on the fermentation and aerobic stability of whole crop wheat silage. Anim. Feed. Sci. Technol. 2000, 88, 39–46. [Google Scholar] [CrossRef]
- Filya, I. The effect of Lactobacillus buchneri and Lactobacillus plantarum on the fermentation, aerobic stability, and ruminal degradability of low dry matter corn and sorghum silages. J. Dairy Sci. 2003, 86, 3575–3581. [Google Scholar] [CrossRef] [Green Version]
- Danner, H.; Holzer, M.; Mayrhuber, E.; Braun, R. Acetic acid increases stability of silage under aerobic conditions. Appl. Environ. Microbiol. 2003, 69, 562–567. [Google Scholar] [CrossRef] [Green Version]
- Njidda, A.A.; Nasiru, A. In vitro gas production and dry matter digestibility of tannin-containing forges of semi-arid region of North-Eastern Nigeria. Pak. J. Nutr. 2010, 9, 60–66. [Google Scholar] [CrossRef] [Green Version]
- Chikagwa-Malunga, S.K.; Adesogan, A.T.; Salawu, M.B.; Szabo, N.J.; Littell, R.C.; Kim, S.C.; Phatak, S.C. Nutritional characterization of Mucuna pruriens: 2. In vitro ruminal fluid fermentability of Mucuna pruriens, Mucuna l-dopa and soybean meal incubated with or without l-dopa. Anim. Feed. Sci. Technol. 2009, 148, 51–67. [Google Scholar] [CrossRef]
- Huisden, C.M.; Szabo, N.J.; Ogunade, I.M.; Adesogan, A.T. Mucuna pruriens detoxification: Effects of ensiling duration and particle size. Anim. Feed. Sci. Technol. 2014, 198, 20–27. [Google Scholar] [CrossRef]
- Moore, K.J.; Jung, H.-J.G. Lignin and fiber digestion. Rangel. Ecol. Manag./J. Range Manag. Arch. 2001, 54, 420–430. [Google Scholar] [CrossRef]
- McDonald, P.; Henderson, A.; Heron, S. The Biochemistry of Silage, 2nd ed.; Chalcombe Publications: Aberystwyth, UK, 1991. [Google Scholar]
- Makkar, H.P.S.; Francis, G.; Becker, K. Bioactivity of phytochemicals in some lesser-known plants and their effects and potential applications in livestock and aquaculture production systems. Anim. Int. J. Anim. Biosci. 2007, 1, 1371–1391. [Google Scholar] [CrossRef] [Green Version]
- Estell, R. Coping with shrub secondary metabolites by ruminants. Small Rumin. Res. 2010, 94, 1–9. [Google Scholar] [CrossRef]
- Agbede, J.O.; Aletor, V.A. Chemical characterization and protein quality evaluation of leaf protein concentrates from Glyricidia sepium and Leucaena leucocephala. Int. J. Food Sci. Technol. 2004, 39, 253–261. [Google Scholar] [CrossRef]
- Belmar, R.; Nava-Montero, R.; Sandoval-Castro, C.; McNab, J.M. Jack bean (Canavalia ensifomis L. DC) in poultry diets: Antinutritional factors and detoxification studies—A review. World’s Poult. Sci. J. 1999, 55, 37–59. [Google Scholar] [CrossRef]
- Janardhanan, K.; Gurumoorthi, P.; Pugalenthi, M. Nutritional potential of five accessions of a South Indian tribal pulse, Mucuna pruriens var utilis I. The effect of processing methods on the content of l-dopa, phytic acid, and oligosaccharides. Trop. Subtrop. Agroecosyst. 2003, 1, 141–152. [Google Scholar]
- Lackey, J.A. A revised classification of the tribe Phaseoleae (Leguminosae: Papilionoideae), and its relation to canavanine distribution. Bot. J. Linn. Soc. 1977, 74, 163–178. [Google Scholar] [CrossRef]
- Udoh, P.; Ekpenyong, J. Effect of Mucuna urens (horse eye bean) on the gonads of male guinea-pigs. Phytother. Res. 2001, 15, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Martens, S.D.; Hoedtke, S.; Avila, P.; Heinritz, S.N.; Zeyner, A. Effect of ensiling treatment on secondary compounds and amino acid profile of tropical forage legumes, and implications for their pig feeding potential. J. Sci. Food Agric. 2014, 94, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Ehsen, S.; Qasim, M.; Abideen, Z.; Rizvi, R.F.; Gul, B.; Ansari, R.; Khan, M.A. Secondary metabolites as anti-nutritional factors in locally used halophytic forage/fodder. Pak. J. Bot. 2016, 48, 629–636. [Google Scholar]
- Njidda, A. Chemical composition, fibre fraction and anti-nutritional substances of semi-arid browse forages of North-Eastern Nigeria. Niger. J. Basic Appl. Sci. 2010, 18, 181–188. [Google Scholar] [CrossRef] [Green Version]
- González Díaz, L.A.; Hoedtke, S.; Castro, A.; Zeynerb, A. Effect of ensilage on L-canavanine reduction in soaked grains of Canavalia ensiformis sole or mixed with Sorghum bicolor. In Proceedings of the Gesellschaft für Ernährungsphysiologie: Proceedings of the Society of Nutrition Physiology; DLG-Verlags GmbH: Frankfurt, Germany, 2010. [Google Scholar]
- González, L.A.; Hoedtke, S.; Castro, A.; Zeynerb, A. Effect of ensiling soaked cowpea (Vigna unguiculata) grains sole or mixed with sorghum (Sorghum bicolor) on the reduction of selected ant-nutritional factors. In 14th Conference European Society of Veterinary Comparative Nutrition (ESVCN); Liesegang, A., Bühler, K., Wanner, M., Eds.; Studentendruckerei University of Zurich: Zurich, Switzerland, 2010; p. S49. [Google Scholar]
- Tao, Y.; Niu, D.; Li, F.; Zuo, S.; Sun, Q.; Xu, C. Effects of Ensiling Oxytropis glabra with Whole-Plant Corn at Different Proportions on Fermentation Quality, Alkaloid Swainsonine Content, and Lactic Acid Bacteria Populations. Animals 2020, 10, 1733. [Google Scholar] [CrossRef]
- Pagoada, H. Tecnología de Producción Para la Yuca (Manihot esculenta Crantz) Fermentada. Master’s Thesis, Universidad Central “Marta Abreu” de Las Villas, Santa Clara, Cuba, 2010. [Google Scholar]
- Weinberg, Z.G.; Muck, R.E. New trends and opportunities in the development and use of inoculants for silage. FEMS Microbiol. Rev. 1996, 19, 53–68. [Google Scholar] [CrossRef]
- Batra, A.; Saxena, R.K. Potential tannase producers from the genera Aspergillus and Penicillium. Process Biochem. 2005, 40, 1553–1557. [Google Scholar] [CrossRef]
- Aguilar-Zárate, P.; Belmares-Cerda, R.; Aguilar, C.; Cruz-Hernández, M.; Montañez, J. Bacterial tannases: Production, properties and applications. Rev. Mex. Ing. Quím. 2014, 13, 63–74. [Google Scholar]
- Wang, N.; Xiong, Y.; Wang, X.; Guo, L.; Lin, Y.; Ni, K.; Yang, F. Effects of Lactobacillus plantarum on Fermentation Quality and Anti-Nutritional Factors of Paper Mulberry Silage. Fermentation 2022, 8, 144. [Google Scholar] [CrossRef]
- Aharwar, A.; Parihar, D.K. Tannases: Production, properties, applications. Biocatal. Agric. Biotechnol. 2018, 15, 322–334. [Google Scholar] [CrossRef]
- Xu, D.; Ding, Z.; Wang, M.; Bai, J.; Ke, W.; Zhang, Y.; Guo, X. Characterization of the microbial community, metabolome and biotransformation of phenolic compounds of sainfoin (Onobrychis viciifolia) silage ensiled with or without inoculation of Lactobacillus plantarum. Bioresour. Technol. 2020, 316, 123910. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ni, Y.; Zhang, P.; Liang, X.; Fang, S. Acidic natural deep eutectic solvents as dual solvents and catalysts for the solubilization and deglycosylation of soybean isoflavone extracts: Genistin as a model compound. Food Chem. 2023, 406, 134999. [Google Scholar] [CrossRef] [PubMed]
- Olukomaiya, O.O.; Adiamo, O.Q.; Fernando, W.C.; Mereddy, R.; Li, X.; Sultanbawa, Y. Effect of solid-state fermentation on proximate composition, anti-nutritional factor, microbiological and functional properties of lupin flour. Food Chem. 2020, 315, 126238. [Google Scholar] [CrossRef] [PubMed]
- El-Shinnawy, M.; Emara, M.; Motawe, H.; Fatma, M.S.; El-Shinnawy, A. Effect of two kinds of bacteria inoculants on preservation and nutritive values of vine broad bean silages, compared with clover hay. In Proceedings of the 4th Scientific Conference of Animal Wealth Research in the Middle East and North Africa, Foreign Agricultural Relations (FAR), Giza, Egypt, 3–5 October 2011; pp. 457–474. [Google Scholar]


{kind=link}
| Parameters | Canavalia ensiformis | Mucuna pruriens | Saccharum officinarum |
|---|---|---|---|
| Proximate chemical analysis (g/kg DM unless stated otherwise) | |||
| Dry Matter (g/kg FM) † | 870 (±2) | 873 (±2) | 280 (±2) |
| Organic Matter | 963 (±2) | 945 (±2) | 950 (±2) |
| Crude Protein | 320 (±2) | 282 (±3) | 30 (±2) |
| Crude Fiber | 90.0 (±0.2) | 70.0 (±0.4) | 280.0 (±0.7) |
| Ether Extract | 29.0 (±0.5) | 42.0 (±1.0) | nd ‡ |
| Starch | 365 (±1) | 298 (±3.0) | nd |
| Calculated WSC 1 | 20 (±2) | 127 (±7) | 380 (±1) |
| Fiber fractions | |||
| Neutral Detergent Fiber | 229 (±1) | 196 (±2) | 540 (±1) |
| Acid Detergent Fiber | 140 (±1) | 113 (±2) | 340 (±2) |
| Cellulose | 123 (±2) | 94 (±2) | 270 (±2) |
| Lignin | 17.0 (±0.1) | 19.0 (±0.6) | 70.0 (±0.3) |
| Plant secondary metabolites (g/kg DM unless stated otherwise) | |||
| Trypsin inhibitor (TIU/g DM) | 220.9 (±0.3) | 227.1 (±0.3) | nd |
| L-canavanine | 41.1 (±1.3) | 16.3 (±2.1) | nd |
| Total tannins | 9.1 (±0.2) | 77.4 (±0.1) | nd |
| L-dopa | 25.5 (±0.9) | 49.9 (±0.2) | nd |
| Phytic acid | 18.4 (±0.3) | 19.3 (±0.2) | nd |
| Parameters | Ensiling Time (Days) | p-Value † | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 30 | 60 | 90 | ||||||||
| Ce-SS | Mp-SS | Ce-SS | Mp-SS | Ce-SS | Mp-SS | Ce-SS | Mp-SS | Ensiling Time | LT | Ensiling Time × LT | |
| Proximate chemical analysis (g/kg DM unless stated otherwise) | |||||||||||
| Dry Matter (g/kg FM ‡) | 472 (±1) x | 473 (±0) x | 437 (±1) y | 438 (±1) y | 435 (±2) by | 438 (±2) ay | 434 (±2) y | 436 (±2) y | <0.001 | 0.014 | 0.789 |
| Organic Matter | 965 (±1) x | 964 (±1) x | 958 (±1) y | 960 (±1) xy | 955 (±2) yz | 958 (±2) y | 952 (±3) z | 955 (±3) y | <0.001 | 0.050 | 0.378 |
| Crude Protein | 205 (±2) a | 182 (±2) b | 204 (±2) a | 181 (±2) b | 203 (±2) a | 181 (±2) b | 203 (±2) a | 180 (±2) b | 0.397 | <0.001 | 0.992 |
| Crude Fiber | 167 (±1) ax | 121 (±0) bx | 166 (±1) ay | 120 (±0) by | 166 (±1) ay | 120 (±0) byz | 165 (±1) ay | 120 (±0) bz | 0.040 | <0.001 | 0.774 |
| Ether Extract | 16.1 (±1.0) b | 25.4 (±0.6) a | 16.0 (±1.0) b | 25.3 (±0.6) a | 16.0 (±1.0) b | 25.2 (±0.6) a | 15.9 (±1.0) b | 25.1 (±0.7) a | 0.984 | <0.001 | 0.984 |
| Starch | 217 (±1) a | 180 (±2) b | 215 (±1) a | 179 (±2) b | 214 (±1) a | 178 (±2) b | 214 (±2) a | 178 (±2) b | 0.098 | <0.001 | 0.983 |
| Fiber fractions | |||||||||||
| Neutral Detergent Fiber | 352 (±1) ax | 330 (±1) bx | 350 (±1) axy | 329 (±1) by | 349 (±2) axy | 328 (±2) by | 348 (±2) ay | 327 (±2) by | 0.006 | <0.001 | 0.943 |
| Acid Detergent Fiber | 231 (±2) a | 165 (±1) b | 229 (±2) a | 164 (±1) b | 228 (±2) a | 164 (±1) b | 228 (±2) a | 163 (±1) b | 0.145 | <0.001 | 0.835 |
| Cellulose | 183 (±2) a | 132 (±1) b | 182 (±2) a | 132 (±1) b | 181 (±2) a | 132 (±1) b | 181 (±2) a | 131 (±1) b | 0.491 | <0.001 | 0.916 |
| Lignin | 38.6 (±0.5) a | 31.3 (±0.4) b | 38.3 (±0.5) a | 31.2 (±0.4) b | 38.1 (±0.5) a | 31.1 (±0.4) b | 38.1 (±0.6) a | 31.0 (±0.4) b | 0.533 | <0.001 | 0.954 |
| Plant secondary metabolites (% disappeared, expressed relative to the original amount) | |||||||||||
| Trypsin Inhibitor | 75.8 (±0.25) b | 84.7 (±0.2) a | 75.9 (±0.3) b | 84.7 (±0.2) a | 75.9 (±0.3) b | 84.8 (±0.3) a | 0.744 | <0.001 | 0.976 | ||
| L-canavanine | 52.7 (±1.98) b | 69.7 (±4.9) a | 52.8 (±1.9) b | 69.8 (±4.9) a | 52.9 (±1.9) b | 69.9 (±4.9) a | 0.995 | <0.001 | 0.999 | ||
| Total tannins | 49.6 (±1.28) b | 53.9 (±0.3) a | 49.7 (±1.4) b | 54.0 (±0.3) a | 49.9 (±1.3) b | 54.1 (±0.3) a | 0.886 | <0.001 | 0.998 | ||
| L-dopa | 85.7 (±1.60) b | 88.7 (±1.4) a | 85.7 (±1.6) b | 88.7 (±1.4) a | 85.8 (±1.6) b | 88.8 (±1.4) a | 0.996 | 0.001 | >0.999 | ||
| Phytic acid | 67.6 (±2.30) a | 64.7 (±2.6) b | 67.7 (±2.3) a | 64.8 (±2.6) b | 67.8 (±2.2) a | 64.9 (±2.7) b | 0.989 | 0.025 | >0.999 | ||
| Parameters § | Ensiling Time (Days) | p-Value ‡ | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 30 | 60 | 90 | |||||||
| Ce-SS | Mp-SS | Ce-SS | Mp-SS | Ce-SS | Mp-SS | Ensiling Time | LT | Ensiling Time × LT | |
| pH | 4.41 (±0.14) bc | 4.43 (±0.10) c | 4.26 (±0.04) b | 4.08 (±0.06) a | 4.29 (±0.02) bc | 4.10 (±0.07) a | <0.001 | <0.001 | 0.005 |
| NH3-N/N (g/100 g) | 5.91 (±0.03) i | 5.30 (±0.04) j | 5.92 (±0.03) i | 5.30 (±0.05) j | 5.92 (±0.03) i | 5.30 (±0.05) j | 0.995 | <0.001 | 0.956 |
| Lactic acid (mg/g FM) | 31.4 (±0.2) j | 33.1 (±0.3) i | 31.3 (±0.2) j | 33.0 (±0.3) i | 31.2 (±0.2) j | 32.9 (±0.3) i | 0.281 | <0.001 | 0.999 |
| Acetic acid (mg/g FM) | 27.0 (±0.2) i | 21.3 (±0.1) j | 26.9 (±0.1) i | 21.2 (±0.1) j | 26.8 (±0.2) i | 21.2 (±0.1) j | 0.075 | <0.001 | 0.962 |
| Ethanol (mg/g FM) | 7.26 (±0.27) i | 6.33 (±0.04) jy | 7.40 (±0.16) i | 6.46 (±0.10) jy | 7.57 (±0.17) i | 6.60 (±0.11) x | <0.001 | <0.001 | 0.968 |
| L/Fa | 0.54 (±0.23) j | 0.61 (±0.29) i | 0.54 (±0.23) j | 0.61 (±0.29) i | 0.54 (±0.23) j | 0.61 (±0.29) i | >0.999 | 0.001 | >0.999 |
| Parameters | Ensiling Time (Days) | p Value ‡ | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 30 | 60 | 90 | ||||||||
| Ce-SS | Mp-SS | Ce-SS | Mp-SS | Ce-SS | Mp-SS | Ce-SS | Mp-SS | Ensiling Time | LT | Ensiling Time × LT | |
| CDOMD (g/kg DM) | 767 (±1) ax | 742 (±2) b | 764 (±1) ay | 740 (±2) b | 763 (±1) ayz | 739 (±2) b | 762 (±1) az | 738 (±1) b | <0.001 | 0.001 | 0.827 |
| ME (MJ/kg DM) | 11.1 (±0.0) ax | 11.0 (±0.0) bx | 11.0 (±0.0) ay | 10.9 (±0.0) by | 10.9 (±0.0) yz | 10.8 (±0.0) yz | 10.8 (±0.0) z | 10.8 (±0.0) z | <0.001 | <0.001 | 0.446 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Artiles-Ortega, E.; Andrade-Yucailla, V.; Medina-López, B.; de la Fe-Rodríguez, P.Y.; Acosta-Lozano, N.; Fievez, V.; Lima-Orozco, R. Combined Ensiling of Tropical Beans and Sugarcane Stalks: Effects on Their Secondary Metabolites. Fermentation 2023, 9, 310. https://doi.org/10.3390/fermentation9030310
Artiles-Ortega E, Andrade-Yucailla V, Medina-López B, de la Fe-Rodríguez PY, Acosta-Lozano N, Fievez V, Lima-Orozco R. Combined Ensiling of Tropical Beans and Sugarcane Stalks: Effects on Their Secondary Metabolites. Fermentation. 2023; 9(3):310. https://doi.org/10.3390/fermentation9030310
Chicago/Turabian StyleArtiles-Ortega, Einar, Verónica Andrade-Yucailla, Beatriz Medina-López, Pedro Yoelvys de la Fe-Rodríguez, Néstor Acosta-Lozano, Veerle Fievez, and Raciel Lima-Orozco. 2023. "Combined Ensiling of Tropical Beans and Sugarcane Stalks: Effects on Their Secondary Metabolites" Fermentation 9, no. 3: 310. https://doi.org/10.3390/fermentation9030310