
Fruit Beer with the Bisucciu Sardinian Apricot Cultivar (Prunus armeniaca L.): A Technological and Analytical Approach
 , , , , , , , , ,
, , , , , , , , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sampling and Preparation of Apricot Puree
2.2. Strains
Starter Preparation
2.3. Beer Production
2.4. Microbiological Analysis
2.4.1. Microbiological Analysis of Fermented Apricot Puree
2.4.2. Growth Kinetics during Fermentation of Beer Wort
2.4.3. Microbial Strains Identification by microFlex™ Mass Spectrometry (MALDI Biotyper, Bruker Daltonik GmbH)
2.5. Standard Quality Attributes of Beers
2.6. Volatiles Profile of Fermented Wort and Beer
2.7. NMR Analysis
2.7.1. Sample Preparation for NMR Analysis
2.7.2. NMR Measurements
2.7.3. NMR Quantification of Metabolites
2.8. Sensorial Analysis
3. Results
3.1. Beer Production
3.1.1. Identification of Yeast Strain in Wort
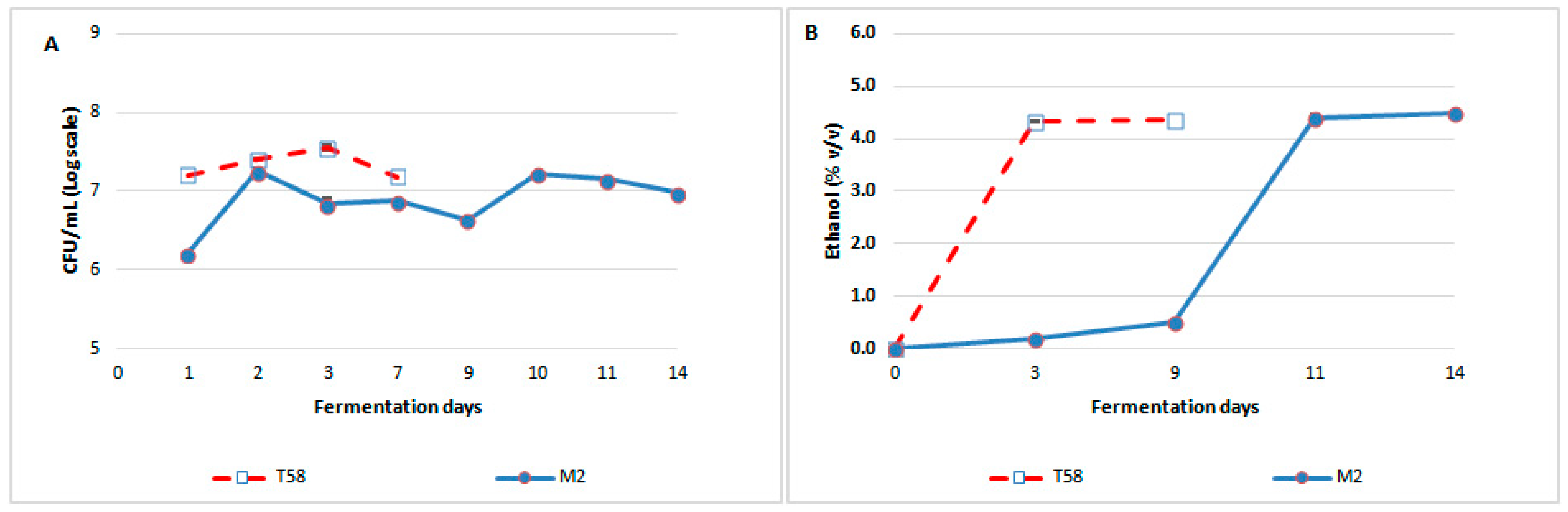
3.1.2. Viable Population Kinetics during Fermentation
3.1.3. Ethanol Production Kinetics during Fermentation
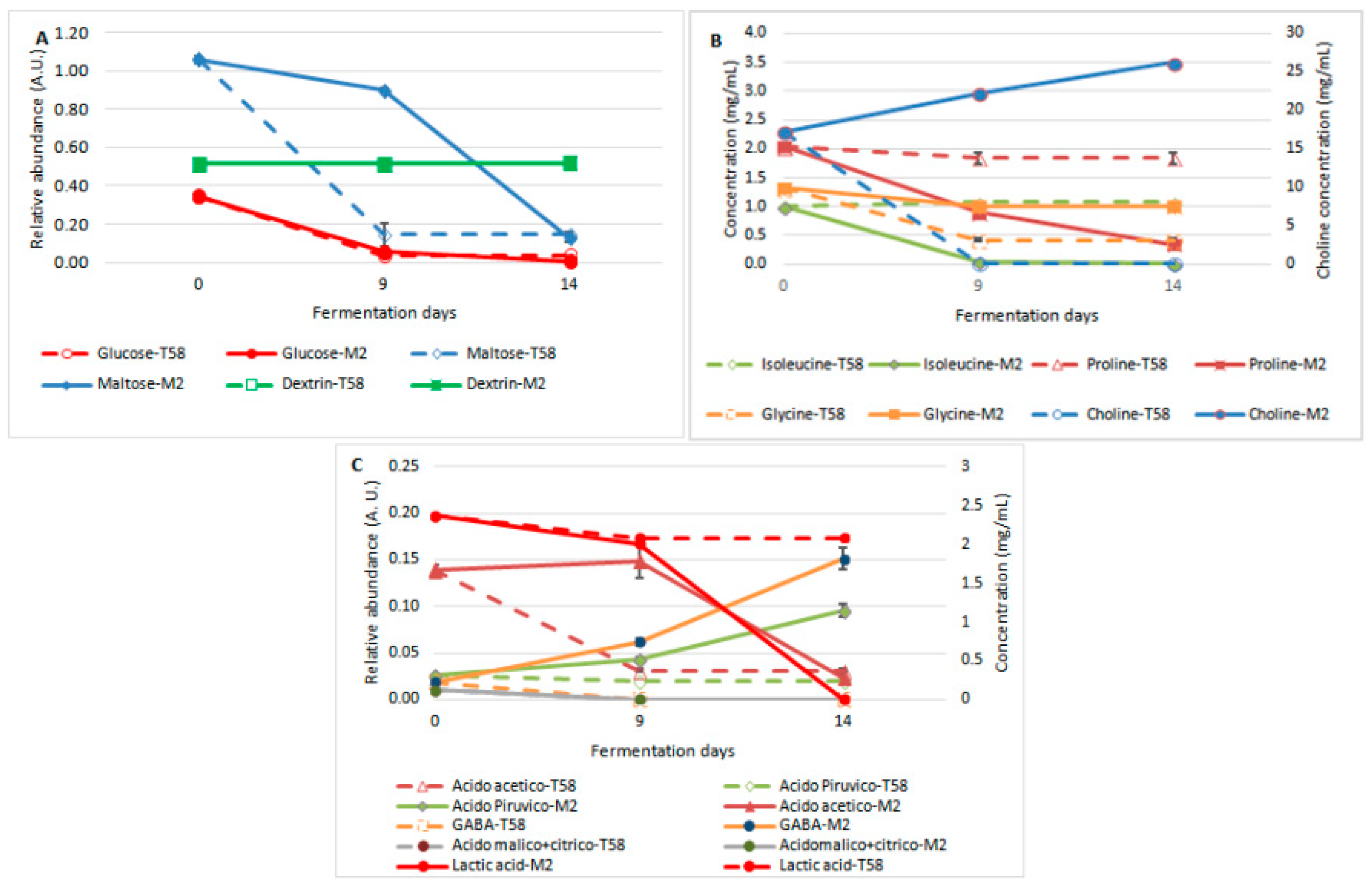
3.1.4. Kinetics of the Carbohydrates, Amino Acids and Acids Concentrations during Fermentation
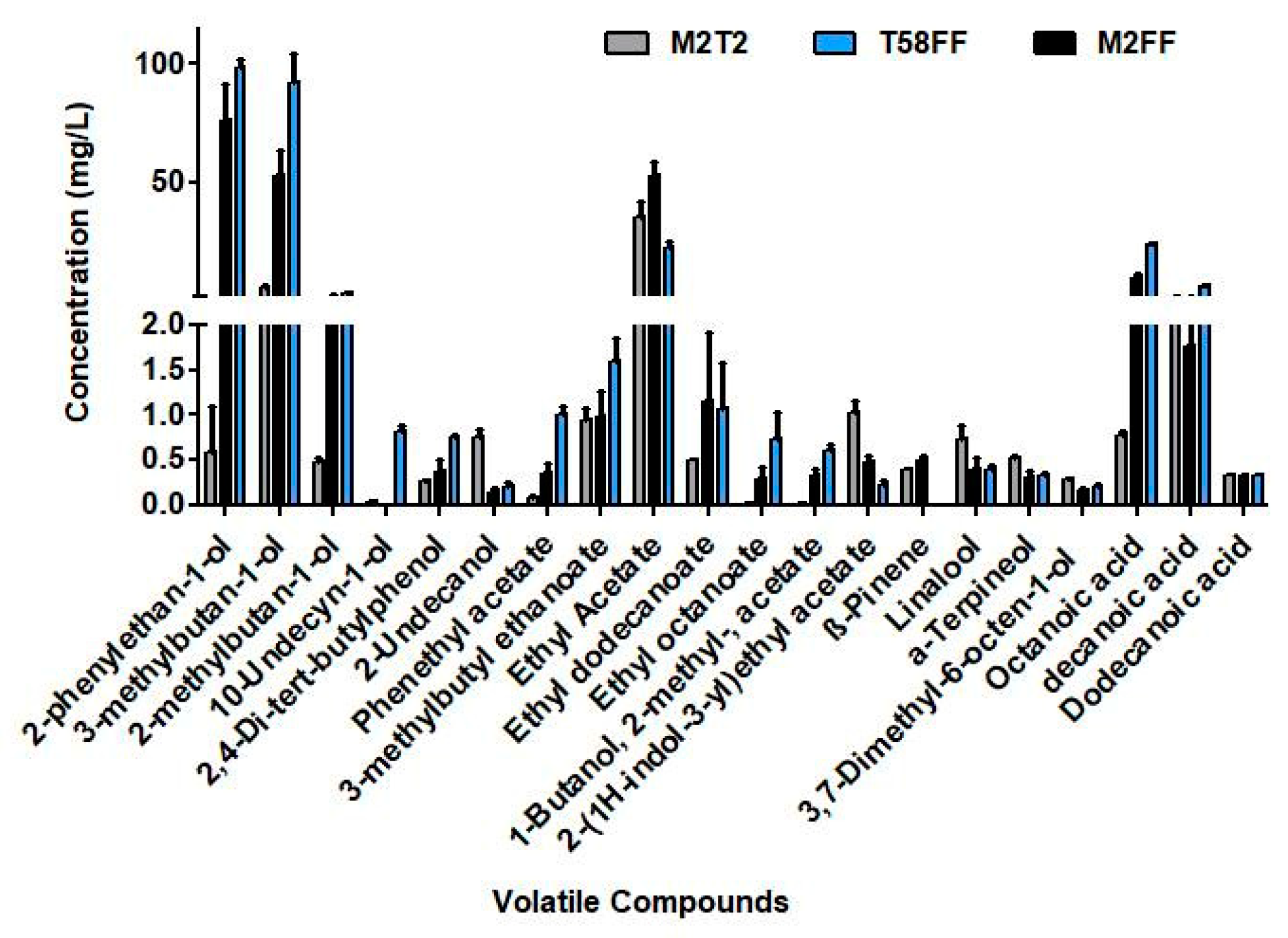
3.1.5. Kinetics of the Volatiles Organic Compounds during Fermentation
3.2. Characterization of Beers
3.2.1. Technological Parameters
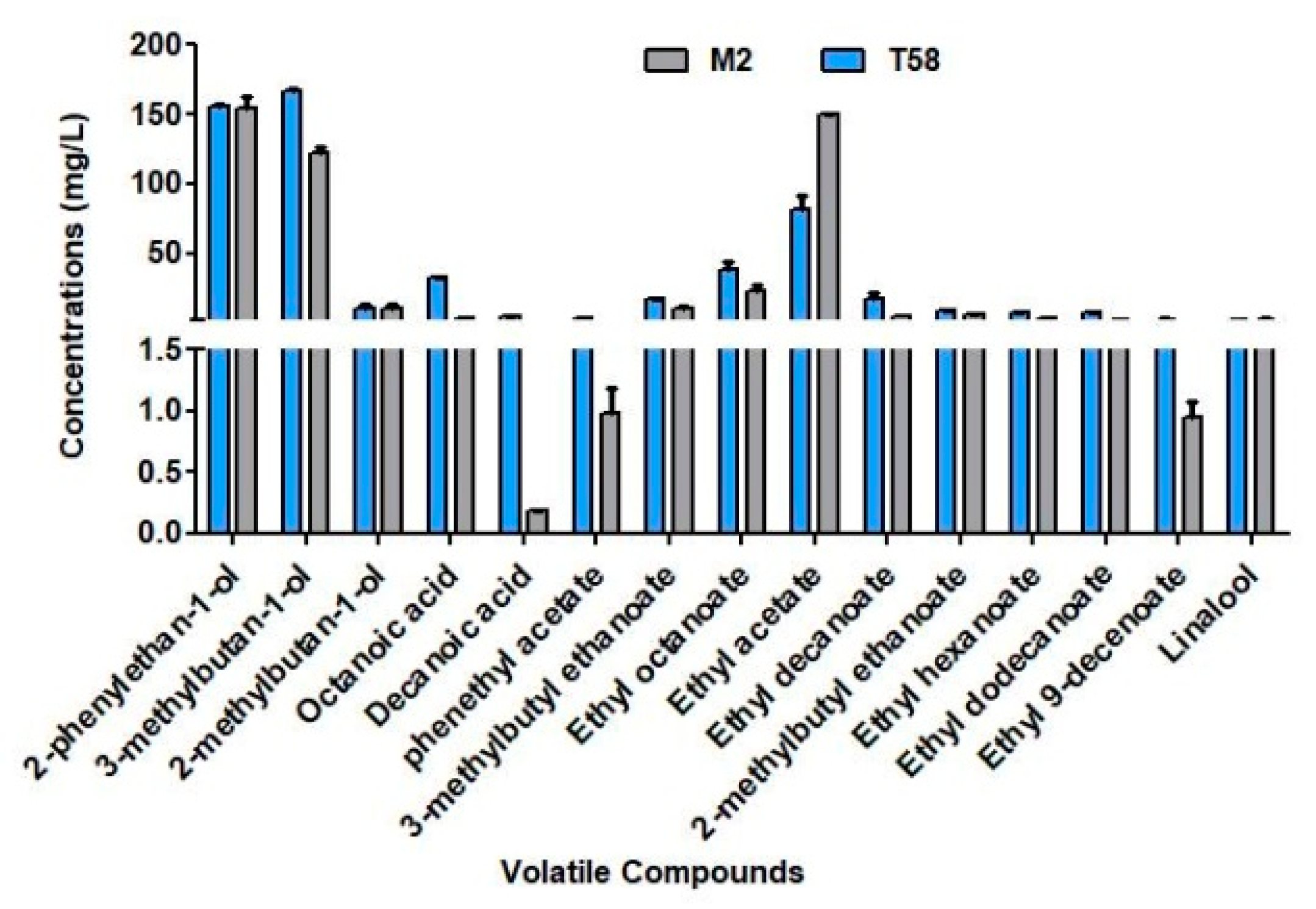
3.2.2. Volatiles Profile
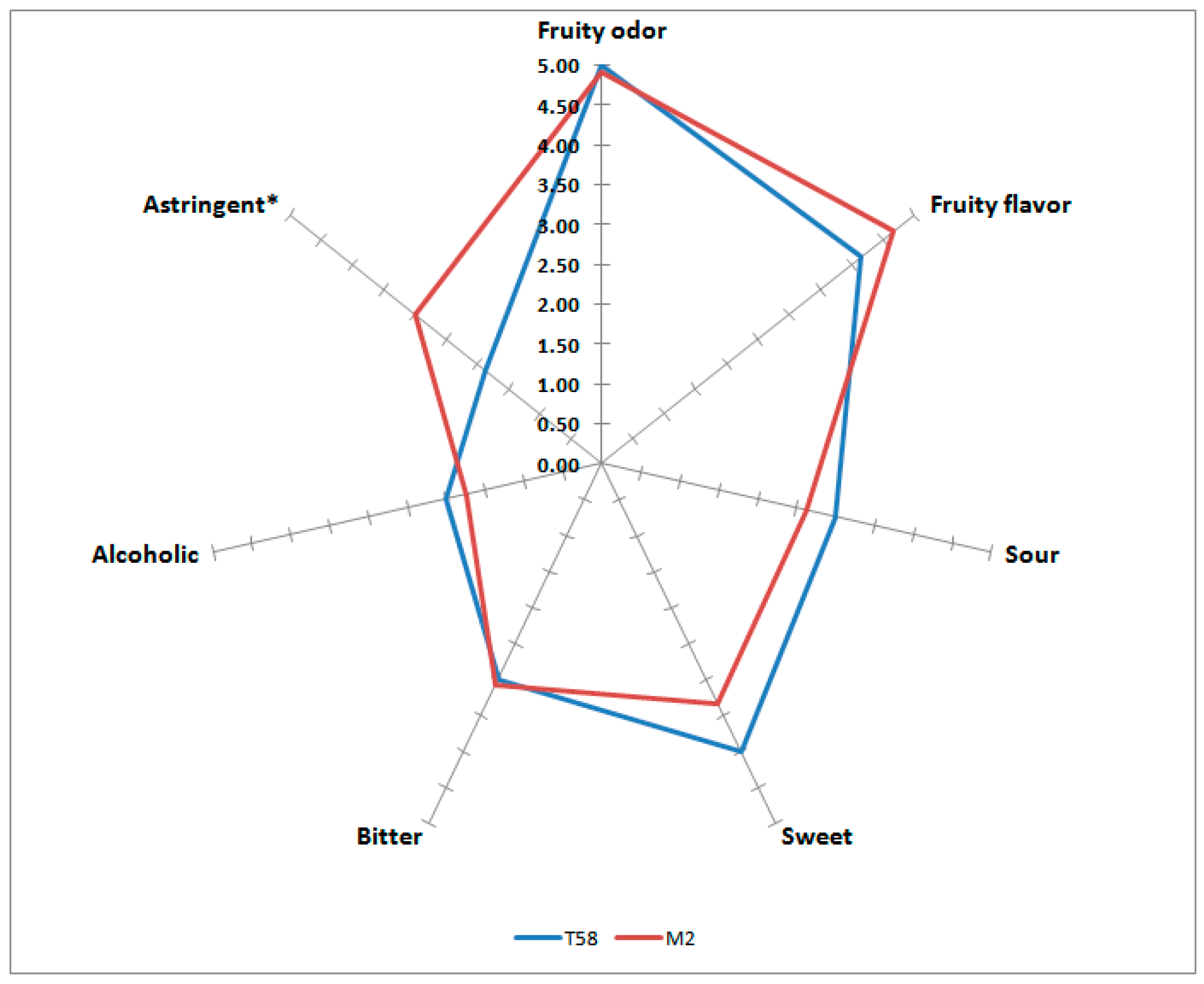
3.2.3. Sensorial Characterization
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caon, G.; Morrone, M.; Feistauer, L.; Sganzerla, D.; Moreira, J.C.F. Moderate beer consumption promotes silymarin-like redox status without affecting the liver integrity in vivo. Food Biosci. 2021, 43, 101307. [Google Scholar] [CrossRef]
- Martínez, A.; Vegara, S.; Martí, N.; Valero, M.; Saura, D. Physicochemical characterization of special persimmon fruit beers using bohemian pilsner malt as a base. J. Inst. Brew. 2017, 123, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Aquilani, B.; Laureti, T.; Poponi, S.; Secondi, L. Beer choice and consumption determinants when craft beers are tasted: An exploratory study of consumer preferences. Food Qual. Prefer. 2015, 41, 214–224. [Google Scholar] [CrossRef]
- Mongelli, A.; Rodolfi, M.; Ganino, T.; Marieschi, M.; Caligiani, A.; Dall’Asta, C.; Bruni, R. Are Humulus lupulus L. ecotypes and cultivars suitable for the cultivation of aromatic hop in Italy? A phytochemical approach. Ind. Crops Prod. 2016, 83, 693–700. [Google Scholar] [CrossRef]
- Forteschi, M.; Porcu, M.C.; Fanari, M.; Zinellu, M.; Secchi, N.; Buiatti, S.; Passaghe, P.; Bertoli, S.; Pretti, L. Quality assessment of Cascade Hop (Humulus lupulus L.) grown in Sardinia. Eur. Food Res. Technol. 2019, 245, 863–871. [Google Scholar] [CrossRef]
- Alfeo, V.; Jaskula-Goiris, B.; Venora, G.; Schimmenti, E.; Aerts, G.; Todaro, A. Screening of durum wheat landraces (Triticum turgidum subsp. durum) for the malting suitability. J. Cereal Sci. 2018, 83, 101–109. [Google Scholar] [CrossRef]
- Benedetti, P.; Salvi, S.; Giomo, A.; Van Deun, R.; Boselli, E.; Frega, N.G. Taste-active Components of Beers from Emmer Wheat (Triticum dicoccum) Malt. Sci. Agric. Bohem. 2016, 47, 82–89. [Google Scholar] [CrossRef] [Green Version]
- Denby, C.M.; Li, R.A.; Vu, V.T.; Costello, Z.; Lin, W.; Chan, L.J.G.; Williams, J.; Donaldson, B.; Bamforth, C.W.; Petzold, C.J.; et al. Industrial brewing yeast engineered for the production of primary flavor determinants in hopped beer. Nat. Commun. 2018, 9, 965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducruet, J.; Rebenaque, P.; Diserens, S.; Kosinska-Cagnazzo, A.; Heritier, I.; Andlauer, W. Amber ale beer enriched with goji berries—The effect on bioactive compound content and sensorial properties. Food Chem. 2017, 226, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Mayer, H.; Ceccaroni, D.; Marconi, O.; Sileoni, V.; Perretti, G.; Fantozzi, P. Development of an all rice malt beer: A gluten free alternative. LWT-Food Sci. Technol. 2016, 67, 67–73. [Google Scholar] [CrossRef]
- De Simone, N.; Russo, P.; Tufariello, M.; Fragasso, M.; Solimando, M.; Capozzi, V.; Grieco, F.; Spano, G. Autochthonous Biological Resources for the Production of Regional Craft Beers: Exploring Possible Contributions of Cereals, Hops, Microbes, and Other Ingredients. Foods 2021, 10, 1831. [Google Scholar] [CrossRef] [PubMed]
- Strong, G.; England, K. BJCP Beer Style Guidelines; Beer Judge Certification Program: Minneapolis, MN, USA, 2021. [Google Scholar]
- Garavaglia, C. The Emergence of Italian Craft Breweries and the Development of Their Local Identity. In The Geography of Beer: Culture and Economics; Hoalst-Pullen, N., Patterson, M.W., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 135–147. [Google Scholar]
- Matraxia, M.; Alfonzo, A.; Prestianni, R.; Francesca, N.; Gaglio, R.; Todaro, A.; Alfeo, V.; Perretti, G.; Columba, P.; Settanni, L.; et al. Non-conventional yeasts from fermented honey by-products: Focus on Hanseniaspora uvarum strains for craft beer production. Food Microbiol. 2021, 99, 103806. [Google Scholar] [CrossRef] [PubMed]
- Iattici, F.; Catallo, M.; Solieri, L. Designing New Yeasts for Craft Brewing: When Natural Biodiversity Meets Biotechnology. Beverages 2020, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Marongiu, A.; Zara, G.; Legras, J.L.; Del Caro, A.; Mascia, I.; Fadda, C.; Budroni, M. Novel starters for old processes: Use of Saccharomyces cerevisiae strains isolated from artisanal sourdough for craft beer production at a brewery scale. J. Ind. Microbiol. Biotechnol. 2015, 42, 85–92. [Google Scholar] [CrossRef]
- Mascia, I.; Fadda, C.; Dostalek, P.; Karabin, M.; Zara, G.; Budroni, M. Is it possible to create an innovative craft durum wheat beer with sourdough yeasts? A case study. J. Inst. Brew. 2015, 121, 283–286. [Google Scholar] [CrossRef]
- Catallo, M.; Nikulin, J.; Johansson, L.; Krogerus, K.; Laitinen, M.; Magalhães, F.; Piironen, M.; Mikkelson, A.; Randazzo, C.L.; Solieri, L.; et al. Sourdough derived strains of Saccharomyces cerevisiae. and their potential for farmhouse ale brewing. J. Inst. Brew. 2020, 126, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Pires, E.J.; Teixeira, J.A.; Brányik, T.; Vicente, A.A. Yeast: The soul of beer’s aroma—A review of flavour-active esters and higher alcohols produced by the brewing yeast. Appl. Microbiol. Biotechnol. 2014, 98, 1937–1949. [Google Scholar] [CrossRef] [Green Version]
- Holt, S.; Mukherjee, V.; Lievens, B.; Verstrepen, K.J.; Thevelein, J.M. Bioflavoring by non-conventional yeasts in sequential beer fermentations. Food Microbiol. 2018, 72, 55–66. [Google Scholar] [CrossRef]
- Serra Colomer, M.; Funch, B.; Forster, J. The raise of Brettanomyces yeast species for beer production. Curr. Opin. Biotechnol. 2019, 56, 30–35. [Google Scholar] [CrossRef]
- Tataridis, P.; Kanelis, A.; Logotetis, S.; Nerancis, E. Use of non-Saccharomyces Torulaspora delbrueckii yeast strains in winemaking and brewing. Zb. Matice Srp. Za Prir. Nauk. 2013, 124, 415–426. [Google Scholar] [CrossRef]
- Larroque, M.N.; Carrau, F.; Farina, L.; Boido, E.; Dellacassa, E.; Medina, K. Effect of Saccharomyces and non-Saccharomyces native yeasts on beer aroma compounds. Int. J. Food Microbiol. 2021, 337, 108953. [Google Scholar] [CrossRef] [PubMed]
- Andrés-Iglesias, C.; Montero, O.; Sancho, D.; Blanco, C.A. New trends in beer flavour compound analysis. J. Sci. Food Agric. 2015, 95, 1571–1576. [Google Scholar] [CrossRef] [PubMed]
- Braga, C.M.; Zielinski, A.A.F.; da Silva, K.M.; de Souza, F.K.F.; Pietrowski, G.d.A.M.; Couto, M.; Granato, D.; Wosiacki, G.; Nogueira, A. Classification of juices and fermented beverages made from unripe, ripe and senescent apples based on the aromatic profile using chemometrics. Food Chem. 2013, 141, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Fancello, F.; Bianco, A.; Niccolai, M.; Zara, G.; Coronas, R.; Serra, E.; D’Hallewin, G.; Valentoni, A.; Santoru, A.; Pretti, L.; et al. Fruit Microbial Communities of the Bisucciu Sardinian Apricot Cultivar (Prunus armeniaca L.) as a Reservoir of New Brewing Starter Strains. Fermentation 2022, 8, 364. [Google Scholar] [CrossRef]
- European Brewery Convention; Fachverlag, H.C. (Eds.) Analytica-Microbiologica-EBC, 5th ed.; EBC: Nürnberg, Germany, 2006. [Google Scholar]
- Fanari, M.; Forteschi, M.; Sanna, M.; Piu, P.P.; Porcu, M.C.; d’Hallewin, G.; Secchi, N.; Zinellu, M.; Pretti, L. Pilot plant production of craft fruit beer by using Ohmic treated fruit puree. J. Food Process. Preserv. 2019, 44, e14339. [Google Scholar] [CrossRef]
- Kleinová, J.; Klejdus, B. Determination of volatiles in beer using solid-phase microextraction in combination with gas chromatography/mass spectrometry. Czech J. Food Sci. 2014, 32, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Bourbon-Melo, N.; Palma, M.; Rocha, M.P.; Ferreira, A.; Bronze, M.R.; Elias, H.; Sá-Correia, I. Use of Hanseniaspora guilliermondii and Hanseniaspora opuntiae to enhance the aromatic profile of beer in mixed-culture fermentation with Saccharomyces cerevisiae. Food Microbiol. 2021, 95, 103678. [Google Scholar] [CrossRef]
- Nord, L.I.; Vaag, P.; Duus, J.Ø. Quantification of Organic and Amino Acids in Beer by 1H NMR Spectroscopy. Anal. Chem. 2004, 76, 4790–4798. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2017, 46, D608–D617. [Google Scholar] [CrossRef]
- Jacob, D.; Deborde, C.; Lefebvre, M.; Maucourt, M.; Moing, A. NMRProcFlow: A graphical and interactive tool dedicated to 1D spectra processing for NMR-based metabolomics. Metabolomics 2017, 13, 36. [Google Scholar] [CrossRef] [Green Version]
- Bharti, S.K.; Roy, R. Quantitative 1H NMR spectroscopy. TrAC Trends Anal. Chem. 2012, 35, 5–26. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, H.; Sidel, J.L. (Eds.) Index. In Sensory Evaluation Practices, 3rd ed.; Academic Press: San Diego, CA, USA, 2004; pp. 365–374. [Google Scholar]
- Saerens, S.M.G.; Delvaux, F.; Verstrepen, K.J.; Van Dijck, P.; Thevelein, J.M.; Delvaux, F.R. Parameters affecting ethyl ester production by Saccharomyces cerevisiae during fermentation. Appl. Environ. Microbiol. 2008, 74, 454–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verstrepen, K.J.; Derdelinckx, G.; Dufour, J.P.; Winderickx, J.; Thevelein, J.M.; Pretorius, I.S.; Delvaux, F.R. Flavour-active esters: Adding fruitiness to beer. J. Biosci. Bioeng. 2003, 96, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Younis, O.S.; Stewart, G.G. Sugar uptake and subsequent ester and higher alcohol production by Saccharomyces cerevisiae. J. Inst. Brew. 1998, 104, 255–264. [Google Scholar] [CrossRef]
- Basso, R.F.; Alcarde, A.R.; Portugal, C.B. Could non-Saccharomyces yeasts contribute on innovative brewing fermentations? Food Res. Int. 2016, 86, 112–120. [Google Scholar] [CrossRef]
- Zeisel, S.H. Choline: An important nutrient in brain development, liver function and carcinogenesis. J. Am. Coll. Nutr. 1992, 11, 473–481. [Google Scholar] [CrossRef]
- Moretti, A.; Paoletta, M.; Liguori, S.; Bertone, M.; Toro, G.; Iolascon, G. Choline: An Essential Nutrient for Skeletal Muscle. Nutrients 2020, 12, 2144. [Google Scholar] [CrossRef]
- López-Sobaler, A.M.; Lorenzo Mora, A.M.; Salas González, M.D.; Suárez, A.P.; Aparicio, A.; Ortega, R.M. Importance of choline in cognitive function. Nutr. Hosp. 2021, 37, 18–23. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Compound | Linear Fit | R2 Value | Odor Threshold (mg/L) | Odor Characteristics |
|---|---|---|---|---|---|
| Alcohols | 2-phenylethan-1-ol | y = 0.0873x + 0.4306 | 0.9657 | 0.10 | Rose, floral |
| Alcohols | 3-methylbutan-1-ol | y = 0.0324x + 0.234 | 0.9834 | 40.00 | Whiskey, solvent |
| Esters | 3-methylbutyl ethanoate | y = 0.5164x + 0.5359 | 0.963 | 1.20 | Fruity, banana |
| Esters | Phenylethyl acetate | y = 4.9085x + 0.7028 | 0.9786 | 3.80 | Floral, honey |
| Esters | Ethyl acetate | y = 0.0453x + 0.2779 | 0.9833 | 21.00 | Ethery, fruity, sweet |
| Parameter | T58 | M2 |
|---|---|---|
| Alcohol (% v/v) | 4.9 ± 0.0 | 5.0 ± 0.0 |
| Density (g/cm3) | 1.0137 ± 0.0000 | 1.0126 ± 0.0000 |
| Re (Real extract) (% w/w) | 5.7 ± 0.0 | 5.5 ± 0.0 |
| Ae (App. extract) (% w/w) | 4.0 ± 0.0 | 3.7 ± 0.0 |
| Oe (Original extract) (% w/w) | 13.1 ± 0.0 | 13.0 ± 0.0 |
| RDF (Real Deg. of Ferm.) | 58 ± 0 | 59 ± 0 |
| Foam stability (s/3 cm) | 203 ± 4 | 313 ± 8 |
| Color (EBC unit) | 10 ± 0 | 11 ± 0 |
| pH | 4.1 ± 0.0 | 4.1 ± 0.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valentoni, A.; Melis, R.; Sanna, M.; Porcu, M.C.; Rodolfi, M.; Braca, A.; Bianco, A.; Zara, G.; Budroni, M.; Anedda, R.; et al. Fruit Beer with the Bisucciu Sardinian Apricot Cultivar (Prunus armeniaca L.): A Technological and Analytical Approach. Fermentation 2023, 9, 305. https://doi.org/10.3390/fermentation9030305
Valentoni A, Melis R, Sanna M, Porcu MC, Rodolfi M, Braca A, Bianco A, Zara G, Budroni M, Anedda R, et al. Fruit Beer with the Bisucciu Sardinian Apricot Cultivar (Prunus armeniaca L.): A Technological and Analytical Approach. Fermentation. 2023; 9(3):305. https://doi.org/10.3390/fermentation9030305
Chicago/Turabian StyleValentoni, Antonio, Riccardo Melis, Manuela Sanna, Maria Cristina Porcu, Margherita Rodolfi, Angela Braca, Angela Bianco, Giacomo Zara, Marilena Budroni, Roberto Anedda, and et al. 2023. "Fruit Beer with the Bisucciu Sardinian Apricot Cultivar (Prunus armeniaca L.): A Technological and Analytical Approach" Fermentation 9, no. 3: 305. https://doi.org/10.3390/fermentation9030305