
Populations of Saccharomyces cerevisiae in Vineyards: Biodiversity and Persistence Associated with Terroir

Abstract
:1. Introduction
2. Materials and Methods
2.1. Vineyard Selection
2.2. Sampling, Yeast Isolation, and Identification
2.3. S. cerevisiae Molecular Characterization and Data Analysis
2.4. Genetic and Ecological Diversity Analysis of S. cerevisiae Grape Populations
- Average genetic diversity (h): intra-population genetic diversity calculated as average diversity per locus.
- Coefficient of interpopulation genetic diversity (Gst): equivalent to Wright’s Fst [36], it quantifies the level of differentiation among populations considering the total genetic diversity of the populations compared (Ht) and the genetic diversity of each population (Hs).
- Nei’s coefficient of genetic identity among populations [37]: it considers all isolates from every population. Another dendrogram was constructed by UPGMA from these identity coefficients.
3. Results
3.1. Vineyard S. cerevisiae Populations
3.2. Analysis of S. cerevisiae Population Genetic Diversity
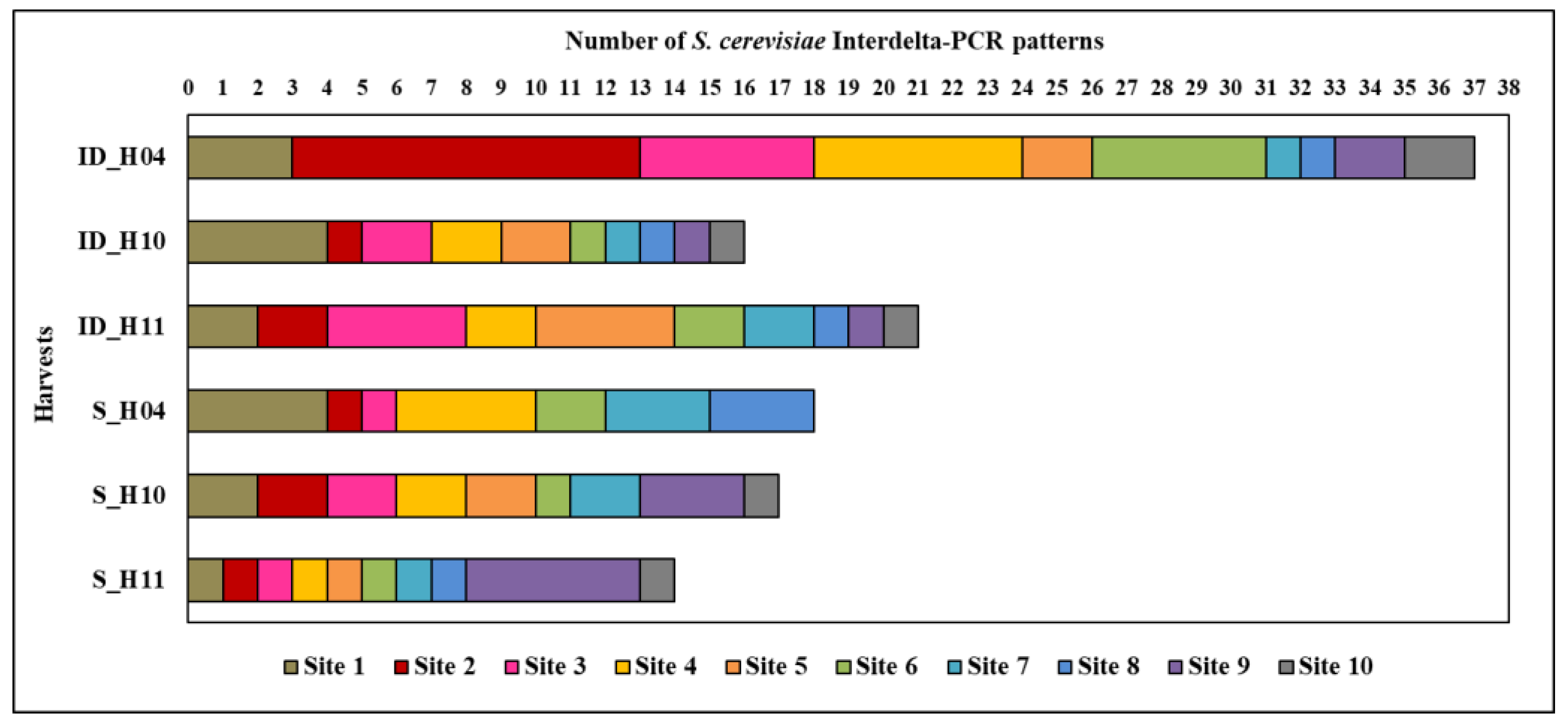
3.3. Analysis of S. cerevisiae Population Ecological Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- International Organization of Vine and Wine. Definition of Vitivinicultural “Terroir”. In Wine Resolution OIV/VITI 333/2010; International Organization of Vine: Dijon, France, 2010. [Google Scholar]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud, A. Handbook of Enology. The Microbiology of Wine and Vinifications, 2nd ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2006; Volume 1, ISBN 978-0-470-01036-5. [Google Scholar]
- Fleet, G.H. Wine Yeasts for the Future. FEMS Yeast Res. 2008, 8, 979–995. [Google Scholar] [CrossRef] [Green Version]
- De Celis, M.; Ruiz, J.; Martín-Santamaría, M.; Alonso, A.; Marquina, D.; Navascués, E.; Gómez-Flechoso, M.Á.; Belda, I.; Santos, A. Diversity of Saccharomyces cerevisiae Yeasts Associated to Spontaneous and Inoculated Fermenting Grapes from Spanish Vineyards. Lett. Appl. Microbiol. 2019, 68, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Mercado, L.; Dalcero, A.; Masuelli, R.; Combina, M. Diversity of Saccharomyces Strains on Grapes and Winery Surfaces: Analysis of Their Contribution to Fermentative Flora of Malbec Wine from Mendoza (Argentina) during Two Consecutive Years. Food Microbiol. 2007, 24, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Santamaría, P.; López, R.; del Patrocinio Garijo, M.; Escribano, R.; González-Arenzana, L.; López-Alfaro, I.; Rosa Gutiérrez, A. Biodiversity of Saccharomyces cerevisiae Yeasts in Spontaneous Alcoholic Fermentations: Typical Cellar or Zone Strains? In Advances in Grape and Wine Biotechnology; Morata, A., Loira, I., Eds.; IntechOpen: London, UK, 2019; ISBN 978-1-78984-612-6. [Google Scholar]
- Cordero-Bueso, G.; Arroyo, T.; Serrano, A.; Valero, E. Influence of Different Floor Management Strategies of the Vineyard on the Natural Yeast Population Associated with Grape Berries. Int. J. Food Microbiol. 2011, 148, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Schuller, D.; Alves, H.; Dequin, S.; Casal, M. Ecological Survey of Saccharomyces Cerevisiae Strains from Vineyards in the Vinho Verde Region of Portugal. FEMS Microbiol. Ecol. 2005, 51, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Schuller, D.; Casal, M. The Genetic Structure of Fermentative Vineyard-Associated Saccharomyces Cerevisiae Populations Revealed by Microsatellite Analysis. Antonie Leeuwenhoek 2007, 91, 137–150. [Google Scholar] [CrossRef] [Green Version]
- Vigentini, I.; De Lorenzis, G.; Fabrizio, V.; Valdetara, F.; Faccincani, M.; Panont, C.A.; Picozzi, C.; Imazio, S.; Failla, O.; Foschino, R. The Vintage Effect Overcomes the Terroir Effect: A Three Year Survey on the Wine Yeast Biodiversity in Franciacorta and Oltrepò Pavese, Two Northern Italian Vine-Growing Areas. Microbiology 2015, 161, 362–373. [Google Scholar] [CrossRef] [Green Version]
- Gayevskiy, V.; Goddard, M.R. Geographic Delineations of Yeast Communities and Populations Associated with Vines and Wines in New Zealand. ISME J. 2012, 6, 1281–1290. [Google Scholar] [CrossRef] [Green Version]
- Knight, S.; Goddard, M.R. Quantifying Separation and Similarity in a Saccharomyces Cerevisiae Metapopulation. ISME J. 2015, 9, 361–370. [Google Scholar] [CrossRef] [Green Version]
- DeLong, D.C. Defining Biodiversity. Wildl. Soc. Bull. 1996, 24, 738–749. [Google Scholar]
- Hughes, J.B.; Bohannan, B.J.M. Application of Ecological Diversity Statistics in Microbial Ecology. In Molecular Microbial Ecology Manual; Kluwer Academic Publishers: Amsterdam, The Netherlands, 2004; Volume 1, pp. 1321–1344. ISBN 1-4020-2176-3. [Google Scholar]
- Daly, A.; Baetens, J.; De Baets, B. Ecological Diversity: Measuring the Unmeasurable. Mathematics 2018, 6, 119. [Google Scholar] [CrossRef] [Green Version]
- Carmona-Galindo, V.D.; Carmona, T.V. La diversidad de los análisis de diversidad. Bioma 1. 2013, 2013, 20–28. [Google Scholar]
- Gotelli, N.J.; Chao, A. Measuring and Estimating Species Richness, Species Diversity, and Biotic Similarity from Sampling Data. In Encyclopedia of Biodiversity; Elsevier: Amsterdam, The Netherlands, 2013; pp. 195–211. ISBN 978-0-12-384720-1. [Google Scholar]
- Hughes, J.B.; Hellmann, J.J. The Application of Rarefaction Techniques to Molecular Inventories of Microbial Diversity. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2005; Volume 397, pp. 292–308. ISBN 978-0-12-182802-8. [Google Scholar]
- Instituto Nacional de Vitivinicultura. Informe de Variedades Malbec; Instituto Nacional de Vitivinicultura: Mendoza, Argentina, 2022. [Google Scholar]
- Catania, C.D.; Tonietto, J.; Avagnina De Del Monte, S.; Martín Uliarte, E.; Del Monte, R.F. El Clima Vitícola de las Regiones Productoras de uvas para Vinos de Argentina; Embrapa Uva e Vinho: Bento Gonçalves, Brazil, 2007; ISBN 978-85-89921-04-6. [Google Scholar]
- Combina, M.; Mercado, L.; Borgo, P.; Elia, A.; Jofre, V.; Ganga, A.; Martinez, C.; Catania, C. Yeasts Associated to Malbec Grape Berries from Mendoza, Argentina. J. Appl. Microbiol. 2005, 98, 1055–1061. [Google Scholar] [CrossRef] [PubMed]
- Combina, M.; Elía, A.; Mercado, L.; Catania, C.; Ganga, A.; Martinez, C. Dynamics of Indigenous Yeast Populations during Spontaneous Fermentation of Wines from Mendoza, Argentina. Int. J. Food Microbiol. 2005, 99, 237–243. [Google Scholar] [CrossRef]
- Mercado, L.; Jubany, S.; Gaggero, C.; Masuelli, R.W.; Combina, M. Molecular Relationships Between Saccharomyces Cerevisiae Strains Involved in Winemaking from Mendoza, Argentina. Curr. Microbiol. 2010, 61, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Mercado, L.; Sturm, M.E.; Rojo, M.C.; Ciklic, I.; Martínez, C.; Combina, M. Biodiversity of Saccharomyces Cerevisiae Populations in Malbec Vineyards from the “Zona Alta Del Río Mendoza” Region in Argentina. Int. J. Food Microbiol. 2011, 151, 319–326. [Google Scholar] [CrossRef]
- González, M.L.; Sturm, M.E.; Lerena, M.C.; Rojo, M.C.; Chimeno, S.V.; Combina, M.; Mercado, L.A. Persistence and Reservoirs of Saccharomyces Cerevisiae Biodiversity in Different Vineyard Niches. Food Microbiol. 2020, 86, 103328. [Google Scholar] [CrossRef]
- Santos, J.A.; Fraga, H.; Malheiro, A.C.; Moutinho-Pereira, J.; Dinis, L.-T.; Correia, C.; Moriondo, M.; Leolini, L.; Dibari, C.; Costafreda-Aumedes, S.; et al. A Review of the Potential Climate Change Impacts and Adaptation Options for European Viticulture. Appl. Sci. 2020, 10, 3092. [Google Scholar] [CrossRef]
- Pallmann, C.L.; Brown, J.A.; Olineka, T.L.; Cocolin, L.; Mills, D.A.; Bisson, L.F. Use of WL Medium to Profile Native Flora Fermentations. Am. J. Enol. Vitic. 2001, 52, 198–203. [Google Scholar] [CrossRef]
- International Organisation of Vine and Wine (2022) Compendium of International Methods of Wine and Must Analysis, OIV, Paris, France, ISBN: 978-2-85038-052-5 Volumes I and II ISBN: 978-2-85038-054-9 vol II. Available online: https://www.oiv.int/es/standards/compendium-of-international-methods-of-wine-and-must-analysis (accessed on 15 December 2022).
- Fleet, G.H.; Heard, G.M. Yeasts-Growth during Fermentation. In Wine Microbiology and Biotechnology; Harwood Academic Publishers: Chur, Switzerland, 1993; pp. 42–43. [Google Scholar]
- Hoffman, C.S.; Winston, F. A Ten-Minute DNA Preparation from Yeast Efficiently Releases Autonomous Plasmids for Transformation of Escherichia Coli. Gene 1987, 57, 267–272. [Google Scholar] [CrossRef]
- Legras, J.-L.; Karst, F. Optimisation of Interdelta Analysis for Saccharomyces Cerevisiae Strain Characterisation. FEMS Microbiol. Lett. 2003, 221, 249–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavel, A.B.; Vasile, C.I. PyElph—A Software Tool for Gel Images Analysis and Phylogenetics. BMC Bioinform. 2012, 13, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Rambaut, A. FigTree–Tree Figure Drawing Tool Version v.1.4.4. Institute of Evolutionary Biology, University of Edinburgh: Edinburgh. 2018. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 7 December 2021).
- Yeh, F.C.; Boyle, T.J.B. Population Genetic Analysis of Codominant and Dominant Markers and Quantitative Traits. Belg. J. Bot. 1997, 129, 157–163. [Google Scholar]
- Wright, S. Evolution and the Genetics of Populations; University of Chicago Press: Chicago, IL, USA, 1978; Volume 4, ISBN 0-226-91052-0. [Google Scholar]
- Nei, M. Analysis of Gene Diversity in Subdivided Populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [Green Version]
- Tristezza, M.; Vetrano, C.; Bleve, G.; Spano, G.; Capozzi, V.; Logrieco, A.; Mita, G.; Grieco, F. Biodiversity and Safety Aspects of Yeast Strains Characterized from Vineyards and Spontaneous Fermentations in the Apulia Region, Italy. Food Microbiol. 2013, 36, 335–342. [Google Scholar] [CrossRef]
- Colwell, R.K. Estimates: Statistical Estimation of Species Richness and Shared Species from Samples. Version 9. User’s Guide and Application. 2013. Available online: http://purl.oclc.org/estimates (accessed on 2 December 2022).
- Börlin, M.; Venet, P.; Claisse, O.; Salin, F.; Legras, J.-L.; Masneuf-Pomarede, I. Cellar-Associated Saccharomyces Cerevisiae Population Structure Revealed High-Level Diversity and Perennial Persistence at Sauternes Wine Estates. Appl. Environ. Microbiol. 2016, 82, 2909–2918. [Google Scholar] [CrossRef] [Green Version]
- Granchi, L.; Ganucci, D.; Buscioni, G.; Mangani, S.; Guerrini, S. The Biodiversity of Saccharomyces Cerevisiae in Spontaneous Wine Fermentation: The Occurrence and Persistence of Winery-Strains. Fermentation 2019, 5, 86. [Google Scholar] [CrossRef] [Green Version]
- Bisson, L.F.; Karpel, J.E.; Ramakrishnan, V.; Joseph, L. Functional Genomics of Wine Yeast Saccharomyces Cerevisiae. In Advances in Food and Nutrition Research; Elsevier: Amsterdam, The Netherlands, 2007; Volume 53, pp. 65–121. ISBN 978-0-12-373729-8. [Google Scholar]
- Schuller, D.; Cardoso, F.; Sousa, S.; Gomes, P.; Gomes, A.C.; Santos, M.A.S.; Casal, M. Genetic Diversity and Population Structure of Saccharomyces Cerevisiae Strains Isolated from Different Grape Varieties and Winemaking Regions. PLoS ONE 2012, 7, e32507. [Google Scholar] [CrossRef] [Green Version]
- Bisson, L.F. Geographic Origin and Diversity of Wine Strains of Saccharomyces. Am. J. Enol. Vitic. 2012, 63, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Castrillo, D.; Neira, N.; Blanco, P. Saccharomyces Cerevisiae Strain Diversity Associated with Spontaneous Fermentations in Organic Wineries from Galicia (NW Spain). Fermentation 2020, 6, 89. [Google Scholar] [CrossRef]
- Raymond Eder, M.L.; Reynoso, C.; Lauret, S.C.; Rosa, A.L. Isolation and Identification of the Indigenous Yeast Population during Spontaneous Fermentation of Isabella (Vitis labrusca L.) Grape Must. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Setati, M.E.; Jacobson, D.; Andong, U.-C.; Bauer, F. The Vineyard Yeast Microbiome, a Mixed Model Microbial Map. PLoS ONE 2012, 7, e52609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drumonde-Neves, J.; Franco-Duarte, R.; Vieira, E.; Mendes, I.; Lima, T.; Schuller, D.; Pais, C. Differentiation of Saccharomyces Cerevisiae Populations from Vineyards of the Azores Archipelago: Geography vs Ecology. Food Microbiol. 2018, 74, 151–162. [Google Scholar] [CrossRef]
- Vigentini, I.; Maghradze, D.; Petrozziello, M.; Bonello, F.; Mezzapelle, V.; Valdetara, F.; Failla, O.; Foschino, R. Indigenous Georgian Wine-Associated Yeasts and Grape Cultivars to Edit the Wine Quality in a Precision Oenology Perspective. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šuranská, H.; Vránová, D.; Omelková, J. Isolation, Identification and Characterization of Regional Indigenous Saccharomyces Cerevisiae Strains. Braz. J. Microbiol. 2016, 47, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Tra Bi, C.Y.; N’guessan, F.K.; Kouakou, C.A.; Jacques, N.; Casaregola, S.; Djè, M.K. Identification of Yeasts Isolated from Raffia Wine (Raphia Hookeri) Produced in Côte d’Ivoire and Genotyping of Saccharomyces Cerevisiae Strains by PCR Inter-Delta. World J. Microbiol. Biotechnol. 2016, 32, 125. [Google Scholar] [CrossRef]
- Granchi, L.; Ganucci, D.; Viti, C.; Giovannetti, L.; Vincenzini, M. Saccharomyces Cerevisiae Biodiversity in Spontaneous Commercial Fermentations of Grape Musts with ÔadequateÕ and ÔinadequateÕ Assimilable-Nitrogen Content. Lett. Appl. Microbiol. 2003, 36, 54–58. [Google Scholar] [CrossRef]
- Cubillos, F.A.; Vásquez, C.; Faugeron, S.; Ganga, A.; MartÃ-nez, C. Self-Fertilization Is the Main Sexual Reproduction Mechanism in Native Wine Yeast Populations: Sexual Reproduction Mechanisms in Wine Yeast Populations. FEMS Microbiol. Ecol. 2009, 67, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Vezinhet, F.; Hallet, J.; Valade, M.; Poulard, A. Ecological Survey of Wine Yeast Strains by Molecular Methods of Identification. Am. J. Enol. Vitic. 1992, 43, 83–86. [Google Scholar] [CrossRef]
- Alexandre, H. Wine Yeast Terroir: Separating the Wheat from the Chaff—for an Open Debate. Microorganisms 2020, 8, 787. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| V | H | VIS | NI | MP | B (%) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| ID | H04 | 1 | 11 | 1 (45.45%) | 2 (36.36%) | 3 (18.18%) | 27.27 | 31.32 | ||
| 2 | 12 | 4 (8.33%) | 5 (8.33%) | 6 (8.33%) | 7 (8.33%) | 8 (16.67%) | 83.33 | |||
| 9 (8.33%) | 10 (8.33%) | 11 (16.67%) | 12 (8.33%) | 13 (8.33%) | ||||||
| 3 | 8 | 1 (25%) | 14 (12.5%) | 15 (12.5%) | 16 (25%) | 25 (25%) | 62.50 | |||
| 4 | 9 | 1 (22.22%) | 17 (11.11%) | 18 (11.11%) | 19 (11.11%) | 20 (33.33%) | 66.67 | |||
| 22 (11.11%) | ||||||||||
| 5 | 7 | 1 (71.43%) | 25 (28.57%) | 28.57 | ||||||
| 6 | 8 | 20 (12.5%) | 21 (25%) | 23 (12.5%) | 24 (12.5%) | 25 (37.5%) | 62.50 | |||
| 7 | 8 | 26 (100%) | 12.50 | |||||||
| 8 | 7 | 26 (100%) | 14.29 | |||||||
| 9 | 7 | 24 (42.86%) | 25 (57.14%) | 28.57 | ||||||
| 10 | 6 | 20 (66.67%) | 25 (33.33%) | 66.67 | ||||||
| H10 | 1 | 13 | 27 (69%) | 28 (7.7%) | 29 (15.4%) | 30 (7.7%) | 30.77 | 11.29 | ||
| 2 | 20 | 30 (100%) | 5.00 | |||||||
| 3 | 14 | 31 (92.9%) | 32 (7.1%) | 14.29 | ||||||
| 4 | 10 | 33 (90%) | 34 (10%) | 20.00 | ||||||
| 5 | 11 | 35 (45.5%) | 36 (54.5%) | 18.18 | ||||||
| 6 | 10 | 37 (100%) | 10.00 | |||||||
| 7 | 12 | 38 (100%) | 8.33 | |||||||
| 8 | 11 | 30 (100%) | 9.09 | |||||||
| 9 | 13 | 39 (100%) | 7.69 | |||||||
| 10 | 10 | 40 (100%) | 10.00 | |||||||
| H11 | 1 | 15 | 44 (33.3%) | 45 (66.7%) | 13.33 | 8.44 | ||||
| 2 | 17 | 43 (88.2%) | 46 (5.9%) | 47 (5.9%) | 17.65 | |||||
| 3 | 15 | 41 (6.7%) | 48 (6.7%) | 49 (20%) | 50 (66.7%) | 26.67 | ||||
| 4 | 15 | 43 (80%) | 51 (20%) | 6.67 | ||||||
| 5 | 15 | 42 (6.7%) | 52 (66.7%) | 53 (20%) | 54 (6.7%) | 26.67 | ||||
| 6 | 15 | 55 (81.3%) | 56 (18.7%) | 13.33 | ||||||
| 7 | 15 | 57 (80%) | 58 (20%) | 13.33 | ||||||
| 8 | 15 | 49 (100%) | 6.67 | |||||||
| 9 | 15 | 43 (100%) | 6.67 | |||||||
| 10 | 15 | 42 (100%) | 6.67 | |||||||
| Subtotal | 359 | 58 | 16.15 | |||||||
| S | H04 | 1 | 6 | 1 (50%) | 2 (8.35%) | 3 (33.33%) | 6 (8.35%) | 66.67 | 14.29 | |
| 2 | 6 | 2 (100%) | 16.67 | |||||||
| 3 | 6 | 2 (100%) | 16.67 | |||||||
| 4 | 13 | 3 (61.54%) | 4 (23.07%) | 5 (7.69%) | 6 (7.69%) | 30.77 | ||||
| 5 | a | |||||||||
| 6 | 7 | 3 (85.71%) | 7 (14.29%) | 28.57 | ||||||
| 7 | 6 | 4 (16.67%) | 8 (50%) | 9 (33.34%) | 50.00 | |||||
| 8 | 5 | 3 (60%) | 9 (20%) | 10 (20%) | 60.00 | |||||
| 9 | ||||||||||
| 10 | ||||||||||
| H10 | 1 | 14 | 11 (92.9%) | 12 (7.1%) | 14.29 | 7.22 | ||||
| 2 | 14 | 11 (85.7%) | 12 (14.3%) | 14.29 | ||||||
| 3 | 14 | 11 (78.6%) | 13 (21.4%) | 14.29 | ||||||
| 4 | 14 | 11 (64.3%) | 13 (35.7%) | 14.29 | ||||||
| 5 | 9 | 11 (77.8%) | 13 (22.2%) | 22.22 | ||||||
| 6 | 9 | 11 (100%) | 11.11 | |||||||
| 7 | 11 | 11 (81.8%) | 14 (18.2%) | 18.18 | ||||||
| 8 | ||||||||||
| 9 | 6 | 13 (66.7%) | 15 (16.7%) | 16 (16.7%) | 16.67 | |||||
| 10 | 6 | 17 (100%) | ||||||||
| H11 | 1 | 15 | 26 (100%) | 6.67 | 7.38 | |||||
| 2 | 15 | 24 (100%) | 6.67 | |||||||
| 3 | 15 | 24 (100%) | 6.67 | |||||||
| 4 | 15 | 21 (100%) | 6.67 | |||||||
| 5 | 15 | 21 (100%) | 6.67 | |||||||
| 6 | 15 | 21 (100%) | 6.67 | |||||||
| 7 | 14 | 23 (100%) | 7.14 | |||||||
| 8 | 15 | 18 (100%) | 6.67 | |||||||
| 9 | 15 | 11 (20%) | 19 (26.7%) | 20 (20%) | 22 (13.3%) | 23 (20%) | 33.33 | |||
| 10 | 15 | 25 (100%) | 6.67 | |||||||
| Subtotal | 295 | 26 | 9.15 | |||||||
| Total | 654 | 84 | 12.84 | |||||||
| Populations Studied | L | P (%) | h |
|---|---|---|---|
| ID_H04 | 34 | 89.47 | 0.2475 (0.1745) |
| ID_H10 | 33 | 86.84 | 0.2668 (0.1727) |
| ID_H11 | 34 | 89.47 | 0.2473 (0.1699) |
| S_H04 | 28 | 73.68 | 0.2493 (0.1883) |
| S_H10 | 26 | 68.42 | 0.0967 (0.1081) |
| S_H11 | 35 | 92.11 | 0.3133 (0.1818) |
| ID_H04 | ID_H10 | ID_H11 | S_H04 | S_H10 | S_H11 | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NI | S | H′ | J | NI | S | H′ | J | NI | S | H′ | J | NI | S | H′ | J | NI | S | H′ | J | NI | S | H′ | J |
| 8.3 | 3.9 (0.8) | 1.1 (0.7) | 0.8 | 12.4 | 1.6 (0.4) | 0.2 (0.3) | 0.5 | 15.4 | 2.2 (0.5) | 0.5 (0.3) | 0.6 | 7.0 | 2.6 (0.7) | 0.7 (0.5) | 0.7 | 10.8 | 1.9 (0.8) | 0.4 (0.3) | 0.7 | 14.9 | 1.4 (0.4) | 0.2 (0.5) | 0.5 |
| 16.6 | 7.1 (1.4) | 1.6 (0.5) | 0.8 | 24.8 | 3.3 (0.8) | 0.9 (0.2) | 0.8 | 30.8 | 4.4 (1.0) | 1.1 (0.3) | 0.8 | 14.0 | 4.6 (0.9) | 1.2 (0.4) | 0.8 | 21.6 | 3.0 (1.1) | 0.7 (0.3) | 0.6 | 29.8 | 2.7 (0.8) | 0.8 (0.4) | 0.8 |
| 24.9 | 9.5 (1.9) | 1.9 (0.4) | 0.8 | 37.2 | 5.0 (1.2) | 1.3 (0.2) | 0.8 | 46.2 | 6.5 (1.4) | 1.5 (0.3) | 0.8 | 21.0 | 6.0 (1.1) | 1.4 (0.3) | 0.8 | 32.3 | 3.7 (1.2) | 0.8 (0.3) | 0.6 | 44.7 | 3.9 (1.1) | 1.2 (0.4) | 0.8 |
| 33.2 | 11.7 (2.3) | 2.1 (0.4) | 0.8 | 49.6 | 6.4 (1.5) | 1.6 (0.2) | 0.8 | 61.6 | 8.3 (1.7) | 1.7 (0.3) | 0.8 | 28.0 | 7.3 (1.2) | 1.5 (0.2) | 0.8 | 43.1 | 4.5 (1.4) | 0.9 (0.2) | 0.6 | 59.6 | 4.8 (1.4) | 1.3 (0.3) | 0.9 |
| 41.5 | 14.3 (2.7) | 2.2 (0.3) | 0.8 | 62.0 | 7.6 (1.9) | 1.7 (0.2) | 0.9 | 77.0 | 10.0 (2.1) | 1.9 (0.2) | 0.8 | 74.5 | 5.9 (1.7) | 1.5 (0.3) | 0.9 | ||||||||
| 49.8 | 17.1 (3.1) | 2.4 (0.3) | 0.8 | 74.4 | 8.9 (2.2) | 1.9 (0.2) | 0.9 | 92.4 | 11.6 (2.4) | 2.0 (0.2) | 0.8 | 35.0 | 8.4 (1.3) | 1.6 (0.2) | 0.8 | 53.9 | 4.9 (1.5) | 0.9 (0.2) | 0.5 | 89.4 | 7.1 (2.0) | 1.7 (0.2) | 0.9 |
| 58.1 | 19 (3.5) | 2.4 (0.3) | 0.8 | 86.8 | 10.2 (2.5) | 2.0 (0.2) | 0.9 | 107.8 | 13.5 (2.7) | 2.1 (0.2) | 0.8 | 42.0 | 9.3 (1.4) | 1.7 (0.1) | 0.7 | 64.7 | 5.6 (1.6) | 0.9 (0.2) | 0.5 | 104.3 | 7.9 (2.2) | 1.8 (0.2) | 0.9 |
| 66.4 | 21.5 (3.8) | 2.5 (0.2) | 0.8 | 99.2 | 11.5 (2.8) | 2.1 (0.1) | 0.9 | 123.2 | 15.0 (3.0) | 2.2 (0.2) | 0.8 | 49 | 10 (1.5) | 1.7 | 0.7 | 75.4 | 6.1 (1.7) | 0.9 (0.1) | 0.5 | 119.2 | 9.0 (2.4) | 1.9 (0.2) | 0.9 |
| 74.7 | 23.4 (4.2) | 2.6 (0.2) | 0.8 | 111.6 | 12.7 (3.1) | 2.2 (0.1) | 0.9 | 138.6 | 16.7 (3.3) | 2.3 (0.1) | 0.8 | 86.2 | 6.6 (1.8) | 1.0 (0.1) | 0.5 | 134.1 | 9.9 (2.7) | 2.0 (0.1) | 0.9 | ||||
| 83 | 26 (4.5) | 2.7 | 0.8 | 124 | 14 (3.4) | 2.3 | 0.9 | 154 | 18 (3.6) | 2.4 | 0.8 | 97 | 7 (1.9) | 1.0 | 0.5 | 149 | 11 (2.9) | 2.1 | 0.9 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González, M.L.; Chimeno, S.V.; Sturm, M.E.; Becerra, L.M.; Lerena, M.C.; Rojo, M.C.; Combina, M.; Mercado, L.A. Populations of Saccharomyces cerevisiae in Vineyards: Biodiversity and Persistence Associated with Terroir. Fermentation 2023, 9, 292. https://doi.org/10.3390/fermentation9030292
González ML, Chimeno SV, Sturm ME, Becerra LM, Lerena MC, Rojo MC, Combina M, Mercado LA. Populations of Saccharomyces cerevisiae in Vineyards: Biodiversity and Persistence Associated with Terroir. Fermentation. 2023; 9(3):292. https://doi.org/10.3390/fermentation9030292
Chicago/Turabian StyleGonzález, Magalí Lucía, Selva Valeria Chimeno, María Elena Sturm, Lucía Maribel Becerra, María Cecilia Lerena, María Cecilia Rojo, Mariana Combina, and Laura Analía Mercado. 2023. "Populations of Saccharomyces cerevisiae in Vineyards: Biodiversity and Persistence Associated with Terroir" Fermentation 9, no. 3: 292. https://doi.org/10.3390/fermentation9030292