
Assessment of Starters of Lactic Acid Bacteria and Killer Yeasts: Selected Strains in Lab-Scale Fermentations of Table Olives (Olea europaea L.) cv. Leccino
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Olive Samples
2.2. Microbial Strains and Starter Production
2.3. Experimental Set Up
2.4. Microbiological Analysis
2.5. Physico-Chemical Analysis
2.5.1. Sugars and Alcohols
2.5.2. Organic Acids
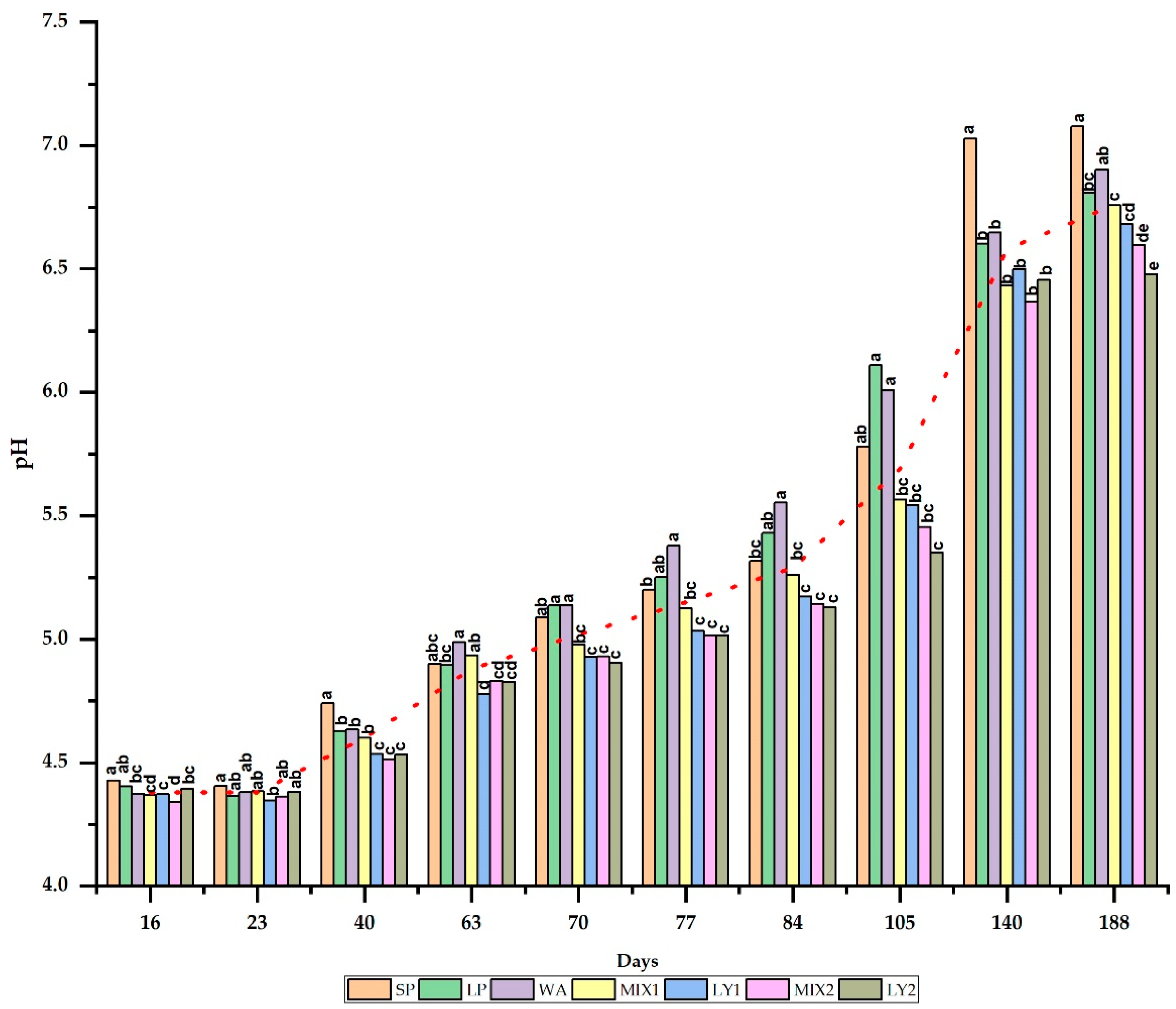
2.5.3. pH
2.5.4. Phenolic Compounds
2.6. Rheological Properties of Olive
2.6.1. Texture Profile Analysis (TPA)
- Hardness: Maximum force of the first compression.
- Springiness: Ratio between the distance traveled during the first compression cycle and the distance during the second cycle. It is the rate at which a deformed sample returns to its original size.
- Cohesiveness: Ratio between the area (work) during the second compression and the area (work) during the first compression. It is the degree to which a product can be deformed before it breaks.
- Gumminess: Hardness × cohesiveness. It is the energy required to disintegrate the product to the state ready for swallowing.
- Chewiness: Hardness × cohesiveness × springiness. It is the number of chews needed to masticate the product until it is ready for swallowing.
2.6.2. Puncture Test
2.7. Chemicals
2.8. Statistical Analyses
3. Results
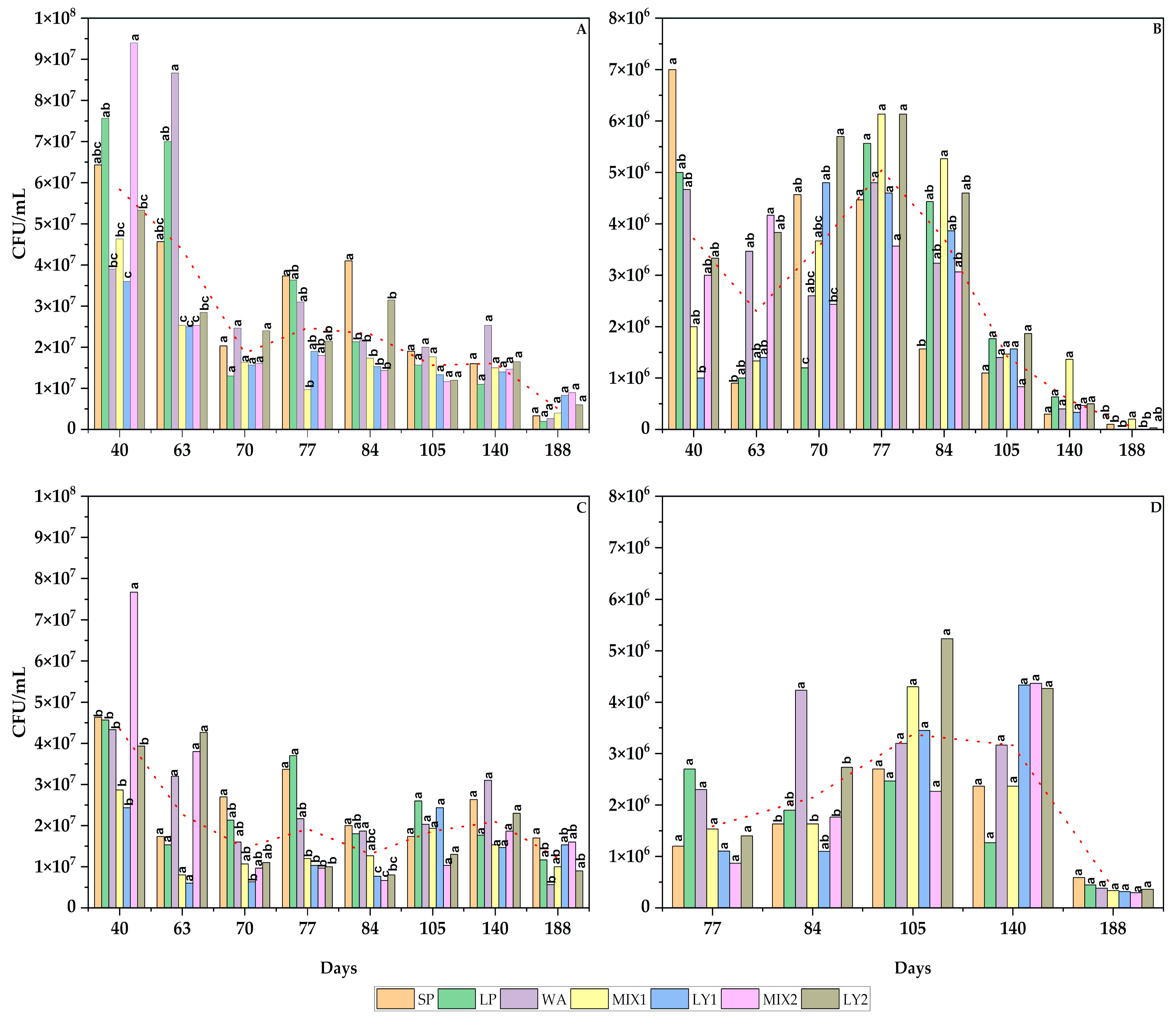
3.1. Microbial Analysis
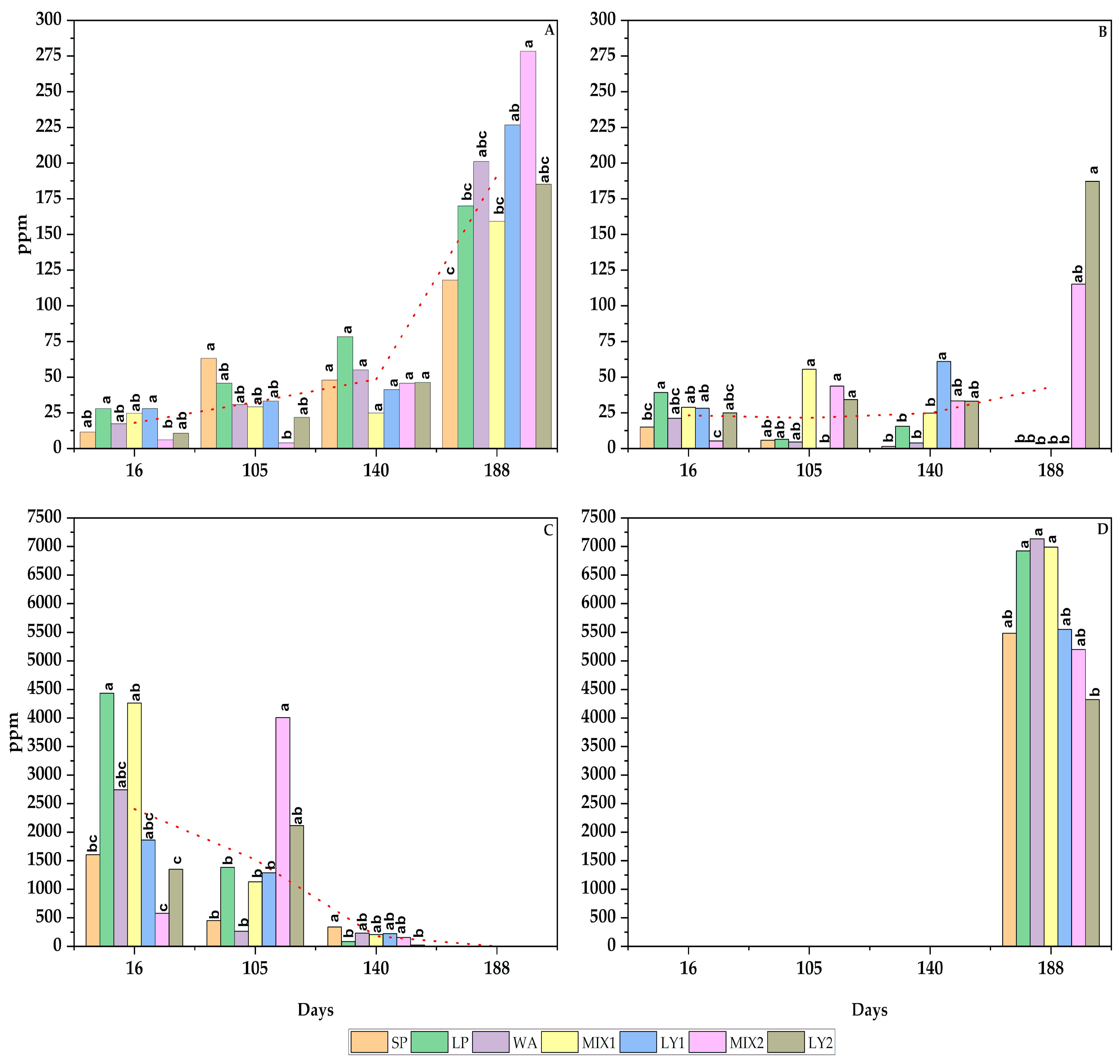
3.2. Physico-Chemical Analysis
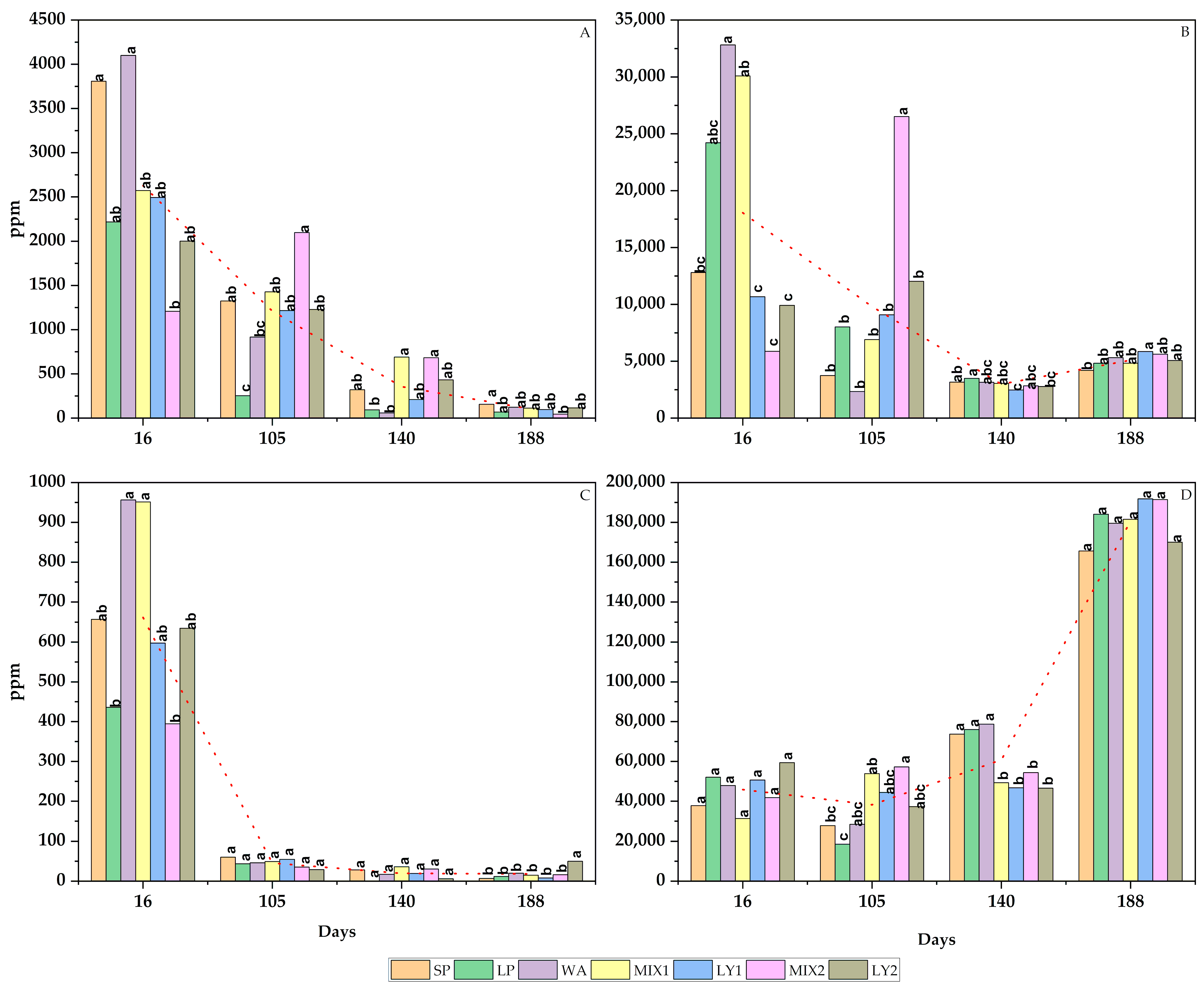
3.2.1. Sugars and Alcohols
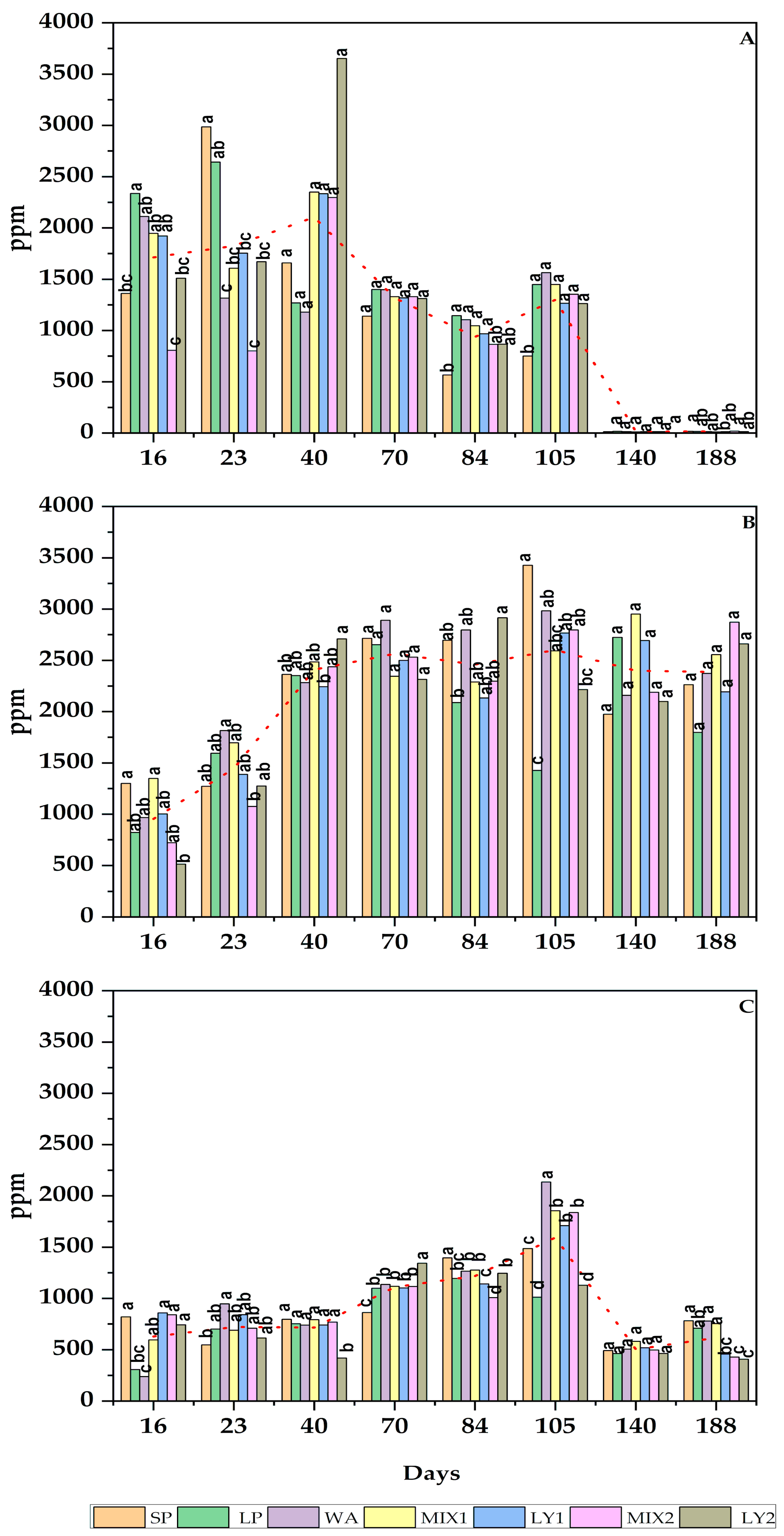
3.2.2. Organic Acids
3.2.3. pH
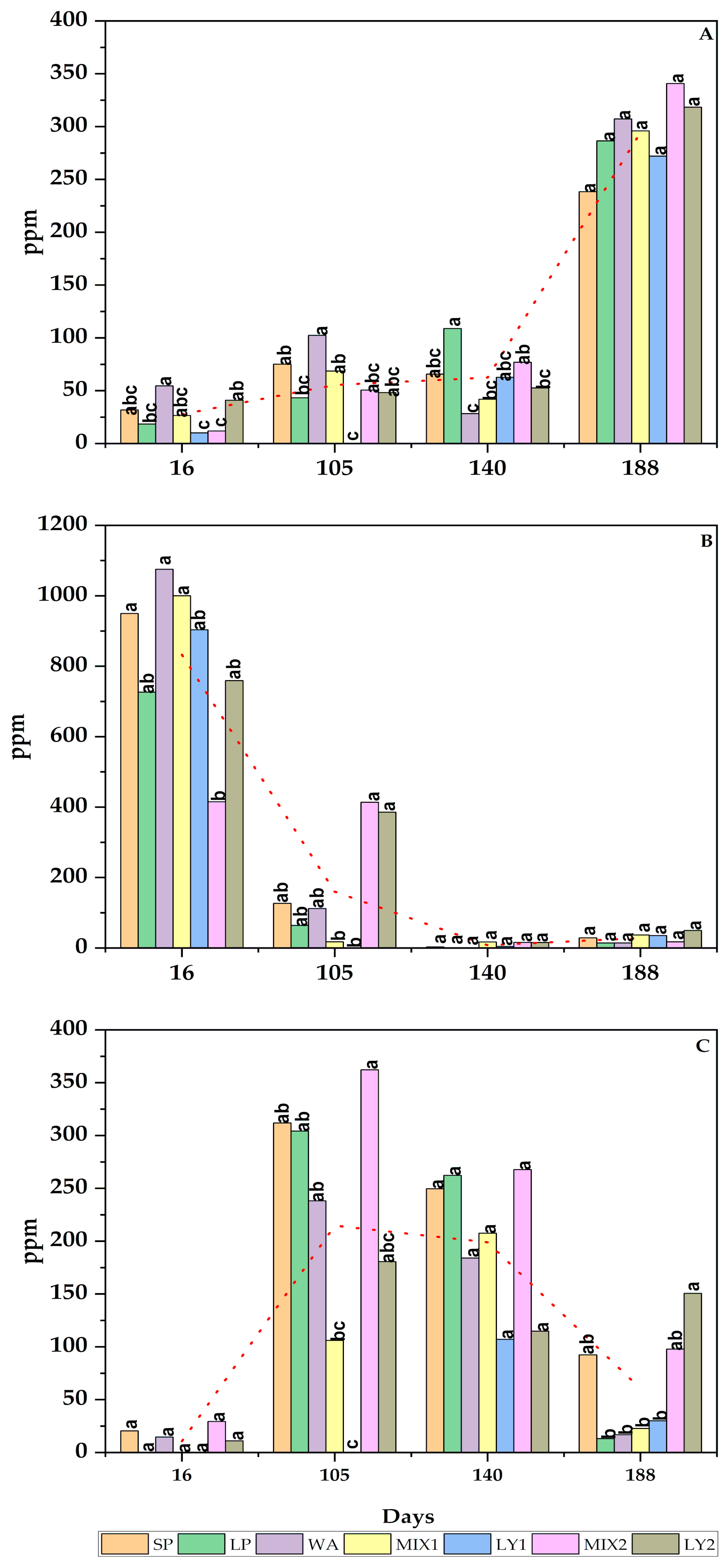
3.2.4. Phenolic Compounds
3.3. Textural Proprieties
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marsilio, V.; Lanza, B.; Pozzi, N. Progress in table olive debittering: Degradation in vitro of oleuropein and its derivatives by Lactobacillus plantarum. JAOCS J. Am. Oil Chem. Soc. 1996, 73, 593–597. [Google Scholar] [CrossRef]
- Ambra, R.; Natella, F.; Bello, C.; Lucchetti, S.; Forte, V.; Pastore, G. Phenolics fate in table olives (Olea europaea L. cv. Nocellara del Belice) debittered using the Spanish and Castelvetrano methods. Food Res. Int. 2017, 100, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Li, Y.; Zheng, M.; Xi, X.; Zhang, X.; Han, C. Structure properties, acquisition protocols, and biological activities of oleuropein aglycone. Front. Chem. 2018, 6, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanza, B.; Zago, M.; Di Marco, S.; Di Loreto, G.; Cellini, M.; Tidona, F.; Bonvini, B.; Bacceli, M.; Simone, N. Single and multiple inoculum of lactiplantibacillus plantarum strains in table olive lab-scale fermentations. Fermentation 2020, 6, 126. [Google Scholar] [CrossRef]
- Marković, A.K.; Torić, J.; Barbarić, M.; Brala, C.J. Hydroxytyrosol, tyrosol and derivatives and their potential effects on human health. Molecules 2019, 24, 2001. [Google Scholar] [CrossRef] [Green Version]
- Panagou, E.Z.; Schillinger, U.; Franz, C.M.A.P.; Nychas, G.J.E. Microbiological and biochemical profile of cv. Conservolea naturally black olives during controlled fermentation with selected strains of lactic acid bacteria. Food Microbiol. 2008, 25, 348–358. [Google Scholar] [CrossRef]
- Charoenprasert, S.; Mitchell, A. Factors influencing phenolic compounds in table olives (Olea europaea). J. Agric. Food Chem. 2012, 60, 7081–7095. [Google Scholar] [CrossRef]
- Grounta, A.; Doulgeraki, A.I.; Nychas, G.J.E.; Panagou, E.Z. Biofilm formation on Conservolea natural black olives during single and combined inoculation with a functional Lactobacillus pentosus starter culture. Food Microbiol. 2016, 56, 35–44. [Google Scholar] [CrossRef]
- Campus, M.; Degirmencioglu, N.; Comunian, R. Technologies and trends to improve table olive quality and safety. Front. Microbiol. 2018, 9, 617. [Google Scholar] [CrossRef] [Green Version]
- Bonatsou, S.; Paramithiotis, S.; Panagou, E.Z. Evolution of yeast consortia during the fermentation of Kalamata natural black olives upon two initial acidification treatments. Front. Microbiol. 2018, 8, 2673. [Google Scholar] [CrossRef]
- Marsilio, V.; Campestre, C.; Lanza, B.; De Angelis, M. Sugar and polyol compositions of some European olive fruit varieties (Olea europaea L.) suitable for table olive purposes. Food Chem. 2001, 72, 485–490. [Google Scholar] [CrossRef]
- Lanza, B.; Di Marco, S.; Bacceli, M.; Di Serio, M.G.; Di Loreto, G.; Cellini, M.; Simone, N. Lactiplantibacillus plantarum used as single, multiple, and mixed starter combined with Candida boidinii for table olive fermentations: Chemical, textural, and sensorial characterization of final products. Fermentation 2021, 7, 239. [Google Scholar] [CrossRef]
- Arroyo-López, F.N.; Romero-Gil, V.; Bautista-Gallego, J.; Rodríguez-Gómez, F.; Jiménez-Díaz, R.; García-García, P.; Querol, A.; Garrido-Fernández, A. Yeasts in table olive processing: Desirable or spoilage microorganisms? Int. J. Food Microbiol. 2012, 160, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Porru, C.; Rodríguez-Gómez, F.; Benítez-Cabello, A.; Jiménez-Díaz, R.; Zara, G.; Budroni, M.; Mannazzu, I.; Arroyo-López, F.N. Genotyping, identification and multifunctional features of yeasts associated to Bosana naturally black table olive fermentations. Food Microbiol. 2018, 69, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Camiolo, S.; Porru, C.; Benítez-Cabello, A.; Rodríguez-Gómez, F.; Calero-Delgado, B.; Porceddu, A.; Budroni, M.; Mannazzu, I.; Jiménez-Díaz, R.; Arroyo-López, F.N. Genome overview of eight Candida boidinii strains isolated from human activities and wild environments. Stand. Genomic Sci. 2017, 12, 70. [Google Scholar] [CrossRef] [Green Version]
- Hernández, A.; Martín, A.; Aranda, E.; Pérez-Nevado, F.; Córdoba, M.G. Identification and characterization of yeast isolated from the elaboration of seasoned green table olives. Food Microbiol. 2007, 24, 346–351. [Google Scholar] [CrossRef]
- Garrido-Fernández, A.; Fernandez Diez, M.J.; Adams, M.R. Table Olives: Production and Processing, 1st ed.; Chapman & Hall: London, UK, 1997. [Google Scholar]
- Chytiri, A.; Tasioula-Margari, M.; Bleve, G.; Kontogianni, V.G.; Kallimanis, A.; Kontominas, M.G. Effect of different inoculation strategies of selected yeast and LAB cultures on Conservolea and Kalamàta table olives considering phenol content, texture, and sensory attributes. J. Sci. Food Agric. 2019, 100, 926–935. [Google Scholar] [CrossRef]
- Blana, V.A.; Grounta, A.; Tassou, C.C.; Nychas, G.J.E.; Panagou, E.Z. Inoculated fermentation of green olives with potential probiotic Lactobacillus pentosus and Lactobacillus plantarum starter cultures isolated from industrially fermented olives. Food Microbiol. 2014, 38, 208–218. [Google Scholar] [CrossRef]
- De Castro, A.; Montaño, A.; Casado, F.J.; Sánchez, A.H.; Rejano, L. Utilization of Enterococcus casseliflavus and Lactobacillus pentosus as starter cultures for Spanish-style green olive fermentation. Food Microbiol. 2002, 19, 637–644. [Google Scholar] [CrossRef]
- Liu, G.L.; Chi, Z.; Wang, G.-Y.; Wang, Z.P.; Li, Y.; Chi, Z.-M. Yeast killer toxins, molecular mechanisms of their action and their applications. Crit. Rev. Biotechnol. 2015, 35, 222–234. [Google Scholar] [CrossRef]
- El-Banna, A.A.; El-Sahn, M.A.; Shehata, M.G. Yeasts Producing Killer Toxins: An Overview. Alexandria J. Food Sci. Technol. 2011, 8, 41–53. [Google Scholar] [CrossRef]
- Mannazzu, I.; Domizio, P.; Carboni, G.; Zara, S.; Zara, G.; Comitini, F.; Budroni, M.; Ciani, M. Yeast killer toxins: From ecological significance to application. Crit. Rev. Biotechnol. 2019, 39, 603–617. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Cortés, F.; Martinez-Calvo, J.; Badenes, M.L.; Bleiholder, H.; Hack, H.; Llacer, G.; Meier, U. Phenological growth stages of olive trees (Olea europaea). Ann. Appl. Biol. 2002, 140, 151–157. [Google Scholar] [CrossRef]
- Fibiani, M.; Paolo, D.; Leteo, F.; Campanelli, G.; Picchi, V.; Bianchi, G.; Lo Scalzo, R. Influence of year, genotype and cultivation system on nutritional values and bioactive compounds in tomato (Solanum lycopersicum L.). Food Chem. 2022, 389, 133090. [Google Scholar] [CrossRef]
- Bleve, G.; Tufariello, M.; Durante, M.; Grieco, F.; Ramires, F.A.; Mita, G.; Tasioula-Margari, M.; Logrieco, A.F. Physico-chemical characterization of natural fermentation process of Conservolea and Kalamàta table olives and developement of a protocol for the pre-selection of fermentation starters. Food Microbiol. 2015, 46, 368–382. [Google Scholar] [CrossRef]
- Bleve, G.; Tufariello, M.; Durante, M.; Perbellini, E.; Ramires, F.A.; Grieco, F.; Cappello, M.S.; de Domenico, S.; Mita, G.; Tasioula-Margari, M.; et al. Physico-chemical and microbiological characterization of spontaneous fermentation of Cellina di Nardò and Leccino table olives. Front. Microbiol. 2014, 5, 570. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Kim, Y.M.; Rhee, H.S.; Lee, M.W.; Park, J.M. Bioethanol production from mannitol by a newly isolated bacterium, Enterobacter sp. JMP3. Bioresour. Technol. 2013, 135, 199–206. [Google Scholar] [CrossRef]
- Russo, A.; Migliorini, M.; Cecchi, L.; Cherubini, C.; Giusti, M.; Zanoni, B.; Perri, E.; Nardi, M.; Sindona, G. Identification and assay of carbohydrates in olive drupes by cesium attachment electrospray tandem mass spectrometry (ESI-MS/MS). J. Mass Spectrom. 2012, 47, 1242–1246. [Google Scholar] [CrossRef]
- Brooks, W.F., Jr. The Enterobacteriaceae: A Preliminary Study of New Methods of Identification: Rapid Biochemical Testing and Salt Tolerance. Master Thesis, Medical University of South Carolina, Charleston, SC, USA, 1970. Available online: https://medica-musc.researchcommons.org/theses/167 (accessed on 20 January 2023).
- Zarour, K.; Vieco, N.; Pérez-Ramos, A.; Nácher-Vázquez, M.; Mohedano, M.L.; Lopez, P. Food Ingredients Synthesized by Lactic Acid Bacteria. Microbial Production of Food Ingredients and Additives. In Handbook of Food Bioengineering, Microbial Production of Food Ingredients and Additives; Holban, A.M., Grumezescu, A.M., Eds.; Elsevier Inc.: Amsterdam, The Netherlands; Academic Press: Cambridge, MA, USA, 2017; pp. 89–124. [Google Scholar]
- Bevilacqua, A.; Campaniello, D.; Speranza, B.; Sinigaglia, M.; Corbo, M.R. Survival of Listeria monocytogenes and Staphylococcus aureus in synthetic brines. Studying the effects of salt, temperature and sugar through the approach of the design of experiments. Front. Microbiol. 2018, 9, 240. [Google Scholar] [CrossRef] [Green Version]
- Massenti, R.; Scalisi, A.; Marra, F.P.; Caruso, T.; Marino, G.; Lo Bianco, R. Physiological and Structural Responses to Prolonged Water Deficit in Young Trees of Two Olive Cultivars. Plants 2022, 11, 1695. [Google Scholar] [CrossRef]
- Benítez-Cabello, A.; Ramiro-García, J.; Romero-Gil, V.; Medina, E.; Arroyo-López, F.N. Fungal biodiversity in commercial table olive packages. Food Microbiol. 2022, 107, 104082. [Google Scholar] [CrossRef] [PubMed]
- Emmanouilidou, M.G.; Koukourikou-Petridou, M.; Gerasopoulos, D.; Kyriacou, M.C. Evolution of physicochemical constitution and cultivar-differential maturity configuration in olive (Olea europaea L.) fruit. Sci. Hortic. 2020, 272, 109516. [Google Scholar] [CrossRef]
- Behera, B.C. Citric acid from Aspergillus niger: A comprehensive overview. Crit. Rev. Microbiol. 2020, 46, 727–749. [Google Scholar] [CrossRef] [PubMed]
- Bavaro, S.; Susca, A.; Frisvad, J.C.; Tufariello, M.; Chytiri, A.; Perrone, G.; Mita, G.; Logrieco, A.F.; Bleve, G. Isolation, characterization, and selection of molds associated to fermented black table olives. Front. Microbiol. 2017, 8, 1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonatsou, S.L.; Iliopoulos, V.; Mallouchos, A.; Gogou, E.; Oikonomopoulou, V.; Krokida, M.; Taoukis, P.; Panagou, E.Z. Effect of osmotic dehydration of olives as pre-fermentation treatment and partial substitution of sodium chloride by monosodium glutamate in the fermentation profile of Kalamata natural black olives. Food Microbiol. 2017, 63, 72–83. [Google Scholar] [CrossRef]
- Lanza, B. Abnormal fermentations in table-olive processing: Microbial origin and sensory evaluation. Front. Microbiol. 2013, 4, 91. [Google Scholar] [CrossRef] [Green Version]
- Servili, M.; Settanni, L.; Veneziani, G.; Esposto, S.; Massitti, O.; Taticchi, A.; Urbani, S.; Montedoro, G.F.; Corsetti, A. The use of Lactobacillus pentosus 1MO to shorten the debittering process time of black table olives (Cv. Itrana and Leccino): A pilot-scale application. J. Agric. Food Chem. 2006, 54, 3869–3875. [Google Scholar] [CrossRef] [PubMed]
- Ranalli, A.; Marchegiani, D.; Contento, S.; Girardi, F.; Nicolosi, M.P.; Brullo, M.D. Variations of iridoid oleuropein in Italian olive varieties during growth and maturation. Eur. J. Lipid Sci. Technol. 2009, 111, 678–687. [Google Scholar] [CrossRef]
- Perpetuini, G.; Caruso, G.; Urbani, S.; Schirone, M.; Esposto, S.; Ciarrocchi, A.; Prete, R.; Garcia-Gonzalez, N.; Battistelli, N.; Gucci, R.; et al. Changes in polyphenolic concentrations of table olives (cv. Itrana) produced under different irrigation regimes during spontaneous or inoculated fermentation. Front. Microbiol. 2018, 9, 1287. [Google Scholar] [CrossRef]
- Anagnostopoulos, D.A.; Kamilari, E.; Tsaltas, D. Evolution of Bacterial Communities, Physicochemical Changes and Sensorial Attributes of Natural Whole and Cracked Picual Table Olives during Spontaneous and Inoculated Fermentation. Front. Microbiol. 2020, 11, 1128. [Google Scholar] [CrossRef]
- Ramírez, E.; Brenes, M.; García, P.; Medina, E.; Romero, C. Oleuropein hydrolysis in natural green olives: Importance of the endogenous enzymes. Food Chem. 2016, 206, 204–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Antuono, I.; Bruno, A.; Linsalata, V.; Minervini, F.; Garbetta, A.; Tufariello, M.; Mita, G.; Logrieco, A.F.; Bleve, G.; Cardinali, A. Fermented Apulian table olives: Effect of selected microbial starters on polyphenols composition, antioxidant activities and bioaccessibility. Food Chem. 2018, 248, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; He, X.W.; Jiang, J.G.; Xu, X.L. Hydroxytyrosol and its potential therapeutic effects. J. Agric. Food Chem. 2014, 62, 1449–1455. [Google Scholar] [CrossRef] [PubMed]
- Gallazzi, M.; Festa, M.; Corradino, P.; Sansone, C.; Albini, A.; Noonan, D.M. An extract of olive mill wastewater downregulates growth, adhesion and invasion pathways in lung cancer cells: Involvement of CXCR4. Nutrients 2020, 12, 903. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sugars and Alcohols | ppm |
|---|---|
| Sucrose | 703.93 ± 125.43 |
| Glucose | 9477.74 ± 203.12 |
| Fructose | 615.80 ± 26.43 |
| Mannitol | 3619.97 ± 570.04 |
| Sorbitol | 92.41 ± 66.39 |
| Phenolic Compounds | ppm |
| Oleuropein | 682.09 ± 230.58 |
| Hydroxytyrosol | 1632.23 ± 675.70 |
| Tyrosol | 1816.50 ± 545.35 |
| Day | 23 | 40 | 105 | 140 | 188 | |
|---|---|---|---|---|---|---|
| Treatments | Sucrose (ppm) | |||||
| SP | 305.49 ± 4.85 e | 52.76 ± 3.88 b | 2.16 ± 0.25 a | 80.71 ± 1.73 a | 0.00 ± 0.00 d | |
| LP | 426.14 ± 6.52 ab | 124.19 ± 7.67 a | 0.00 ± 0.00 b | 103.55 ± 6.64 a | 0.00 ± 0.00 d | |
| WA | 410.69 ± 14.47 abc | 141.17 ± 17.57 a | 0.00 ± 0.00 b | 94.39 ± 3.51 a | 11.78 ± 8.98 cd | |
| MIX1 | 386.85 ± 19.64 c | 0.00 ± 0.00 c | 0.00 ± 0.00 b | 93.32 ± 3.03 a | 133.98 ± 12.17 a | |
| LY1 | 397.47 ± 5.29 bc | 0.00 ± 0.00 c | 0.00 ± 0.00 b | 31.69 ± 2.33 c | 40.14 ± 2.31 b | |
| MIX2 | 340.79 ± 7.95 d | 0.00 ± 0.00 c | 0.00 ± 0.00 b | 72.33 ± 6.96 ab | 23.25 ± 8.05 bc | |
| LY2 | 439.57 ± 11.71 a | 0.00 ± 0.00 c | 0.00 ± 0.00 b | 34.10 ± 9.14 bc | 0.00 ± 0.00 d | |
| Glucose (ppm) | ||||||
| SP | 1336.53 ± 30.22 f | 264.96 ± 5.08 c | 5.14 ± 4.06 ab | 225.35 ± 64.80 a | 0.00 ± 0.00 b | |
| LP | 2678.28 ± 89.35 ab | 589.65 ± 138.75 b | 0.73 ± 0.73 b | 20.60 ± 1.27 b | 0.00 ± 0.00 b | |
| WA | 2370.15 ± 74.86 cd | 886.36 ± 209.44 a | 0.00 ± 0.00 b | 18.90 ± 3.25 b | 17.17 ± 7.52 a | |
| MIX1 | 2091.05 ± 104.84 e | 231.74 ± 17.91 c | 0.00 ± 0.00 b | 12.64 ± 0.94 b | 9.27 ± 2.12 ab | |
| LY1 | 2730.10 ± 38.10 a | 172.82 ± 7.96 c | 0.00 ± 0.00 b | 4.93 ± 0.72 b | 3.80 ± 0.55 b | |
| MIX2 | 2472. 06 ± 34.42 bc | 217.64 ± 18.48 c | 1.38 ± 0.20 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | |
| LY2 | 2204.15 ± 108.27 de | 212.25 ± 3.30 c | 10.17 ± 4.36 a | 2.54 ± 0.37 b | 0.00 ± 0.00 b | |
| Fructose (ppm) | ||||||
| SP | 425.31 ± 11.99 ab | 234.09 ± 3.06 a | 1.05 ± 0.87 b | 37.69 ± 14.64 a | 0.00 ± 0.00 c | |
| LP | 366. 63 ± 33.94 ab | 377.97 ± 52.56 a | 5.62 ± 0.87 b | 1.92 ± 0.56 b | 0.00 ± 0.00 c | |
| WA | 379.57 ± 13.07 ab | 248.65 ± 114.49 a | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 17.52 ± 2.73 b | |
| MIX1 | 468.63 ± 59.45 a | 60.56 ± 7.77 b | 0.00 ± 0.00 b | 3.29 ± 0.73 b | 44.27 ± 3.28 a | |
| LY1 | 374.99 ± 7.21 ab | 83.75 ± 9.93 b | 0.00 ± 0.00 b | 2.91 ± 0.69 b | 6.24 ± 2.15 c | |
| MIX2 | 359.85 ± 12.16 ab | 42.41 ± 20.16 b | 21.93 ± 6.53 b | 12.01 ± 4.14 b | 0.00 ± 0.00 c | |
| LY2 | 287.43 ± 118.51 b | 53.84 ± 8.28 b | 63.09 ± 19.98 a | 20.09 ± 10.27 ab | 0.00 ± 0.00 c | |
| Day | 23 | 40 | 105 | 140 | 188 | |
|---|---|---|---|---|---|---|
| Treatments | Mannitol (ppm) | |||||
| SP | 2841.82 ± 184.23 bc | 330. 47 ± 15.47 b | 1.03 ± 0.52 a | 107.09 ± 13.69 b | 66.47 ± 20.02 b | |
| LP | 2721.22 ± 443.95 c | 881.01 ± 406.13 a | 0.00 ± 0.00 b | 0.00 ± 0.00 d | 666.53 ± 20.87 a | |
| WA | 4248.15 ± 117.43 a | 533.65 ± 12.23 ab | 0.00 ± 0.00 b | 0.00 ± 0.00 d | 608.07 ± 111.80 a | |
| MIX1 | 4205.16 ± 192.19 a | 613.97 ± 89.47 ab | 0.00 ± 0.00 b | 99.60 ± 10.39 b | 123.15 ± 16.39 b | |
| LY1 | 3505.10 ± 184.46 ab | 460.93 ± 31.92 ab | 0.00 ± 0.00 b | 66.89 ± 5.74 c | 103.76 ± 19.62 b | |
| MIX2 | 3146.29 ± 29.75 bc | 445.93 ± 51.91 ab | 0.00 ± 0.00 b | 166.02 ± 6.45 a | 0.00 ± 0.00 b | |
| LY2 | 2539.49 ± 361.20 c | 466.11 ± 72.69 ab | 0.00 ± 0.00 b | 183.03 ± 9.68 a | 43.10 ± 7.92 b | |
| Sorbitol (ppm) | ||||||
| SP | 37.29 ± 13.06 ab | 125.09 ± 3.96 bcd | 0.13 ± 0.08 a | 808.75 ± 39.18 a | 90.20 ± 9.68 c | |
| LP | 27.99 ± 5.01 ab | 97.59 ± 0.44 cd | 0.00 ± 0.00 b | 321.37 ± 41.25 bc | 46.20 ± 5.41 d | |
| WA | 36.42 ± 5.99 ab | 59.67 ± 27.49 d | 0.00 ± 0.00 b | 402.61 ± 54.24 bc | 113.06 ± 0.20 b | |
| MIX1 | 66.90 ± 16.76 a | 211.87 ± 39.55 b | 0.00 ± 0.00 b | 235.62 ± 75.17 c | 198.39 ± 8.32 a | |
| LY1 | 18.58 ± 7.61 b | 201.85 ± 10.38 bc | 0.00 ± 0.00 b | 432.52 ± 46.01 b | 0.00 ± 0.00 e | |
| MIX2 | 16.25 ± 2.69 b | 337.44 ± 84.51 a | 0.00 ± 0.00 b | 290.22 ± 88.32 bc | 16.42 ± 4.77 e | |
| LY2 | 41.03 ± 26.42 ab | 177.08 ± 8.37 bc | 0.00 ± 0.00 b | 479.53 ± 86.09 b | 10.22 ± 4.86 e | |
| Ethanol (ppm) | ||||||
| SP | 0.00 ± 0.00 | 0.00 ± 0.00 | 467.75 ± 132.03 d | 0.00 ± 0.00 | 0.00 ± 0.00 b | |
| LP | 0.00 ± 0.00 | 0.00 ± 0.00 | 822.96 ± 32.33 ab | 0.00 ± 0.00 | 0.00 ± 0.00 b | |
| WA | 0.00 ± 0.00 | 0.00 ± 0.00 | 851.71 ± 78.11 a | 0.00 ± 0.00 | 0.00 ± 0.00 b | |
| MIX1 | 0.00 ± 0.00 | 0.00 ± 0.00 | 598.26 ± 50.76 bcd | 0.00 ± 0.00 | 0.00 ± 0.00 b | |
| LY1 | 0.00 ± 0.00 | 0.00 ± 0.00 | 773.19 ± 40.86 abc | 0.00 ± 0.00 | 0.00 ± 0.00 b | |
| MIX2 | 0.00 ± 0.00 | 0.00 ± 0.00 | 576.26 ± 23.88 cd | 0.00 ± 0.00 | 41.49 ± 5.33 a | |
| LY2 | 0.00 ± 0.00 | 0.00 ± 0.00 | 535.13 ± 102.12 d | 0.00 ± 0.00 | 0.00 ± 0.00 b | |
| Glycerol (ppm) | ||||||
| SP | 0.00 ± 0.00 | 0.00 ± 0.00 c | 0.00 ± 0.00 | 1342.62 ± 287.81 a | 2362.26 ± 496.59 a | |
| LP | 0.00 ± 0.00 | 0.00 ± 0.00 c | 0.00 ± 0.00 | 1331.19 ± 52.87 a | 1487.78 ± 150.80 bc | |
| WA | 0.00 ± 0.00 | 470.96 ± 156.54 b | 0.00 ± 0.00 | 1353.50 ± 156.54 a | 950.54 ± 137.67 cd | |
| MIX1 | 0.00 ± 0.00 | 740.64 ± 228.03 b | 0.00 ± 0.00 | 1726.35 ± 49.04 a | 1792.01 ± 77.12 ab | |
| LY1 | 0.00 ± 0.00 | 880.76 ± 74.01 ab | 0.00 ± 0.00 | 1681.73 ± 207.55 a | 450.85 ± 32.96 de | |
| MIX2 | 0.00 ± 0.00 | 1221.38 ± 274.93 a | 0.00 ± 0.00 | 1345.29 ± 137.99 a | 0.00 ± 0.00 e | |
| LY2 | 0.00 ± 0.00 | 858.80 ± 73.42 ab | 0.00 ± 0.00 | 1368.59 ± 205.04 a | 0.00 ± 0.00 e | |
| Day | 23 | 40 | 105 | 140 | 188 | |
|---|---|---|---|---|---|---|
| Treatments | Oleuropein (ppm) | |||||
| SP | 2368.83 ± 1194.84 bc | 555.58 ± 502.88 a | 258.69 ± 26.71 a | 724.3 ± 680.23 a | 435.61 ± 63.41 a | |
| LP | 4709.37 ± 455.77 a | 342.6 ± 37.11 a | 228.39 ± 51.85 a | 313.01 ± 198.86 a | 456.20 ± 159.71 a | |
| WA | 4281.64 ± 647.67 a | 1887.59 ± 2613.48 a | 242.86 ± 53.21 a | 421.81 ± 144.07 a | 328.18 ± 53.63 a | |
| MIX1 | 3821.5 ± 159.55 ab | 398.43 ± 146.34 a | 379.31 ± 173.46 a | 410.80 ± 108.86 a | 545.42 ± 368.49 a | |
| LY1 | 3969.62 ± 1211.10 a | 374.93 ± 32.43 a | 283.02 ± 97.63 a | 415.54 ± 81.80 a | 337.83 ± 193.36 a | |
| MIX2 | 2470.98 ± 1182.53 bc | 358.59 ± 30.75 a | 270.93 ± 49.17 a | 450.19 ± 117.43 a | 318.80 ± 98.89 a | |
| LY2 | 1367.09 ± 394.85 c | 384.06 ± 61.36 a | 230.36 ± 63.59 a | 466.44 ± 77.55 a | 290.03 ± 21.01 a | |
| Hydroxytyrosol (ppm) | ||||||
| SP | 2009.22 ± 1752.86 a | 2130.88 ± 823.28 b | 2428.56 ± 174.15 a | 1940.845 ± 860.32 a | 1917.57 ± 879.92 ab | |
| LP | 2881.68 ± 78.045 a | 3567.21 ± 257.33 a | 2381.63 ± 399.71 a | 1675.35 ± 397.48 a | 1725.02 ± 865.91 ab | |
| WA | 3574.43 ± 251.95 a | 4034.87 ± 90.911 a | 2494.15 ± 357.76 a | 1506.91 ± 995.07 a | 1877.16 ± 184.35 ab | |
| MIX1 | 1991.57 ± 1059.90 a | 3878.65 ± 398.48 a | 2379.66 ± 932.88 a | 2143.85 ± 315.19 a | 1015.06 ± 105.44 b | |
| LY1 | 2292.77 ± 852.84 a | 3531.42 ± 544.91 a | 1768.14 ± 1134.18 ab | 1813.44 ± 582.48 a | 1258.71 ± 666.4 ab | |
| MIX2 | 3397.36 ± 479.51 a | 3741.07 ± 214.48 a | 1131.05 ± 1794.00 b | 2407.07 ± 1195.88 a | 1275.51 ± 239.39 ab | |
| LY2 | 2822.89 ± 828.89 a | 3033.28 ± 1070.70 ab | 1411.71 ± 897.04 b | 2276.69 ± 573.87 a | 2254.53 ± 439.46 a | |
| Tyrosol (ppm) | ||||||
| SP | 719.41 ± 221.50 b | 379.89 ± 260.59 a | 163.25 ± 6.44 ab | 413.38 ± 103.69 a | 277.7 ± 40.36 a | |
| LP | 767.18 ± 55.87 b | 192.77 ± 9.35 b | 150.4 ± 13.28 b | 357.33 ± 21.34 a | 304.11 ± 32.64 a | |
| WA | 1069.78 ± 170.11 ab | 219.16 ± 5.20 ab | 161.03 ± 29.77 ab | 365.74 ± 80.85 a | 255.81 ± 24.90 a | |
| MIX1 | 913.75 ± 65.43 ab | 211.62 ± 13.81 ab | 219.47 ± 56.96 a | 359.77 ± 36.99 a | 310.21 ± 75.51 a | |
| LY1 | 978.33 ± 345.91 ab | 205.96 ± 20.49 ab | 190.66 ± 58.56 ab | 398.12 ± 37.70 a | 244.64 ± 75.15 a | |
| MIX2 | 1187.25 ± 379.71 a | 216.53 ± 21.95 ab | 188.65 ± 25.85 ab | 391.93 ± 71.35 a | 284.12 ± 55.74 a | |
| LY2 | 759.83 ± 99.78 b | 200.88 ± 21.34 b | 192.52 ± 36.58 ab | 323.54 ± 84.63 a | 261.95 ± 35.40 a | |
| Treatments | Firmness (g) | Area (g × mm) | Rigidity Index (g/mm) |
|---|---|---|---|
| SP | 37.59 ± 1.01 a | 30.73 ± 1.24 a | 20.68 ± 0.56 ab |
| LP | 36.21 ± 0.91 ab | 28.46 ± 1.01 a | 21.01 ± 0.71 ab |
| WA | 36.05 ± 0.83 ab | 29.30 ± 1.03 a | 19.36 ± 0.36 ab |
| MIX1 | 36.44 ± 0.90 ab | 28.26 ± 0.89 a | 20.69 ± 0.55 ab |
| LY1 | 33.28 ± 0.96 b | 26.68 ± 1.16 a | 18.90 ± 0.48 b |
| MIX2 | 38.23 ± 1.27 a | 30.05 ± 1.27 a | 21.75 ± 0.63 a |
| LY2 | 36.81 ± 1.04 b | 28.04 ± 1.14 a | 22.00 ± 0.78 a |
| Treatments | Hardness (g) | Springiness (mm) | Cohesiveness | Gumminess (g) | Chewiness (g) |
|---|---|---|---|---|---|
| SP | 286.35 ± 11.47 ab | 0.66 ± 0.01 ab | 0.50 ± 0.01 a | 142.46 ± 5.45 abc | 96.79 ± 4.79 bc |
| LP | 315.20 ± 17.71 ab | 0.67 ± 0.01 ab | 0.50 ± 0.01 a | 155.40 ± 8.30 ab | 126.90 ± 6.43 a |
| WA | 334.29 ± 12.26 ab | 0.66 ± 0.01 ab | 0.48 ± 0.01 a | 158.83 ± 5.65 ab | 107.31 ± 4.31 abc |
| MIX1 | 337.11 ± 10.55 a | 0.69 ± 0.01 a | 0.48 ± 0.01 a | 162.80 ± 5.14 a | 116.23 ± 3.90 ab |
| LY1 | 321.51 ± 13.74 ab | 0.69 ± 0.01 a | 0.48 ± 0.01 a | 155.81 ± 6.29 abc | 115.08 ± 4.54 ab |
| MIX2 | 277.62 ± 11.61 b | 0.61 ± 0.01 c | 0.48 ± 0.01 a | 130.37 ± 5.39 c | 81.52 ± 4.04 c |
| LY2 | 314.41 ± 23.30 ab | 0.65 ± 0.01 cb | 0.47 ± 0.01 a | 134.42 ± 5.75 bc | 101.06 ± 6.45 abc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bencresciuto, G.F.; Mandalà, C.; Migliori, C.A.; Cortellino, G.; Vanoli, M.; Bardi, L. Assessment of Starters of Lactic Acid Bacteria and Killer Yeasts: Selected Strains in Lab-Scale Fermentations of Table Olives (Olea europaea L.) cv. Leccino. Fermentation 2023, 9, 182. https://doi.org/10.3390/fermentation9020182
Bencresciuto GF, Mandalà C, Migliori CA, Cortellino G, Vanoli M, Bardi L. Assessment of Starters of Lactic Acid Bacteria and Killer Yeasts: Selected Strains in Lab-Scale Fermentations of Table Olives (Olea europaea L.) cv. Leccino. Fermentation. 2023; 9(2):182. https://doi.org/10.3390/fermentation9020182
Chicago/Turabian StyleBencresciuto, Grazia Federica, Claudio Mandalà, Carmela Anna Migliori, Giovanna Cortellino, Maristella Vanoli, and Laura Bardi. 2023. "Assessment of Starters of Lactic Acid Bacteria and Killer Yeasts: Selected Strains in Lab-Scale Fermentations of Table Olives (Olea europaea L.) cv. Leccino" Fermentation 9, no. 2: 182. https://doi.org/10.3390/fermentation9020182