
Production of Pigments under Submerged Culture through Repeated Batch Fermentation of Immobilized Talaromyces atroroseus GH2
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism
2.2. Media
2.3. Inoculum
2.4. Support Selection for Immobilization
2.5. Improvement of Immobilization Conditions
2.6. Kinetics Evaluation under Improved Immobilization Conditions
2.7. Analytical Methods
2.8. Data Analysis
3. Results and Discussion
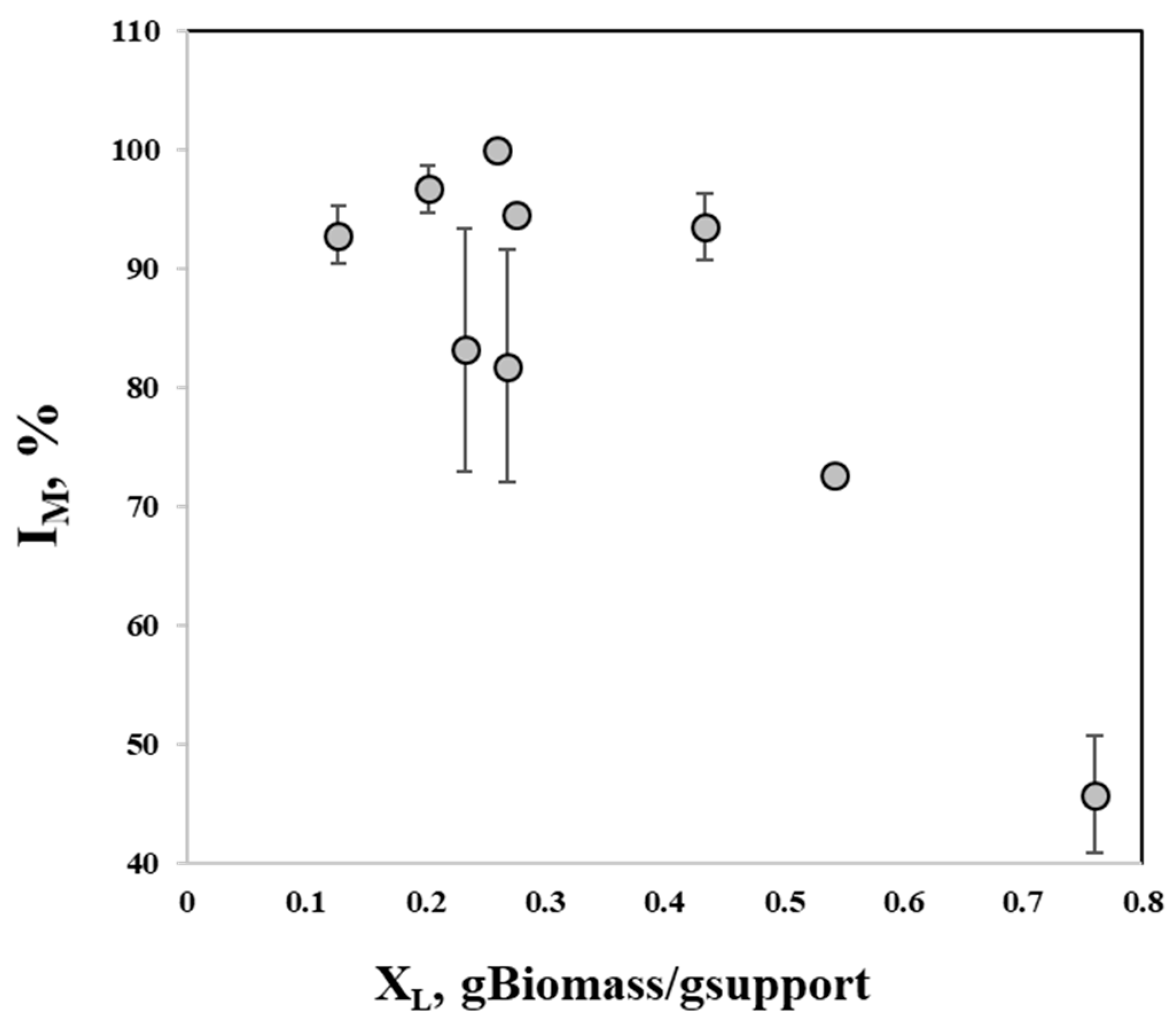
3.1. Support Selection for Immobilization
3.2. Improvement of Immobilization Conditions
3.2.1. Taguchi Experimental Design L9 Results
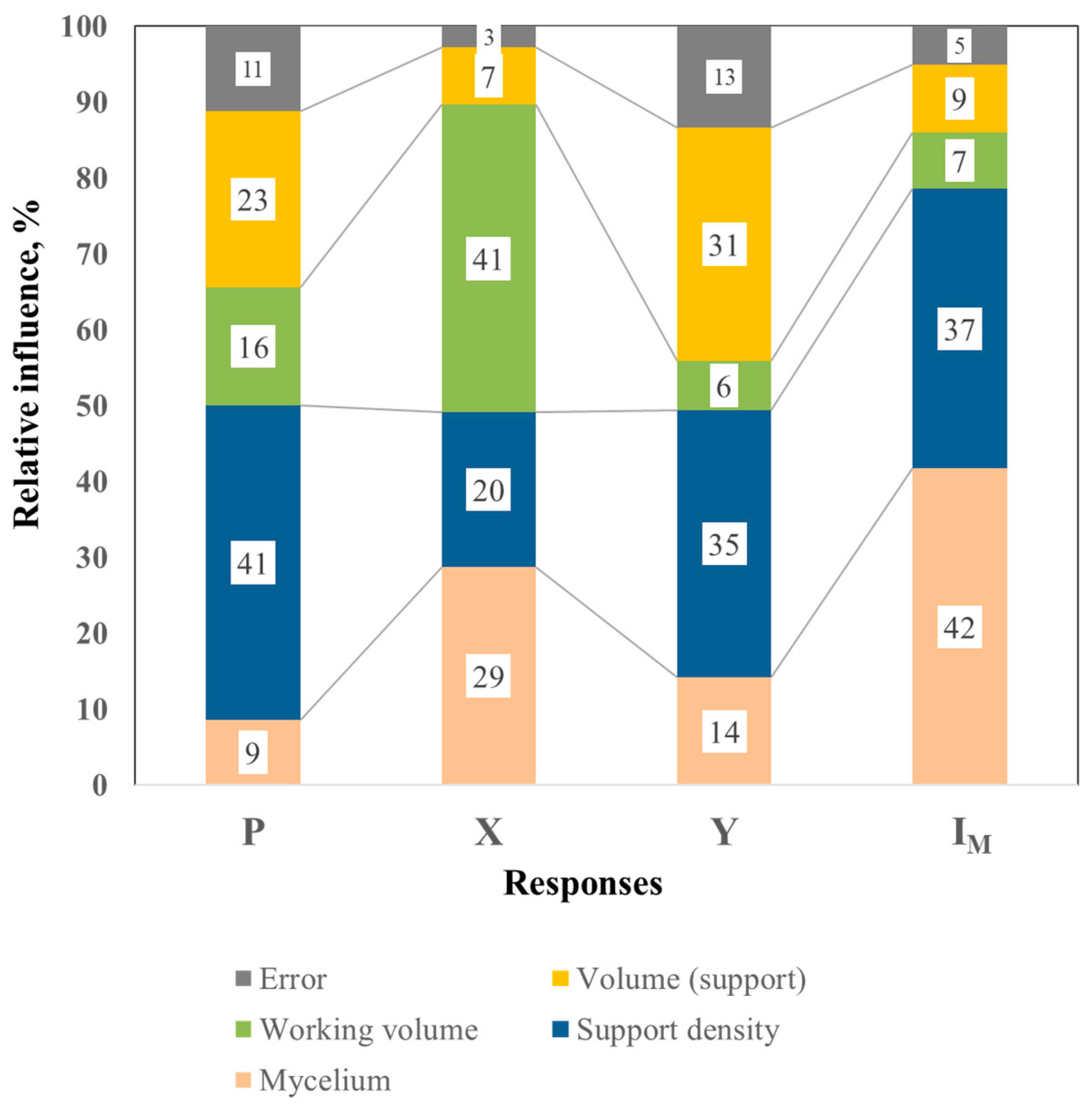
3.2.2. Analysis of the Relative Influence of Factors
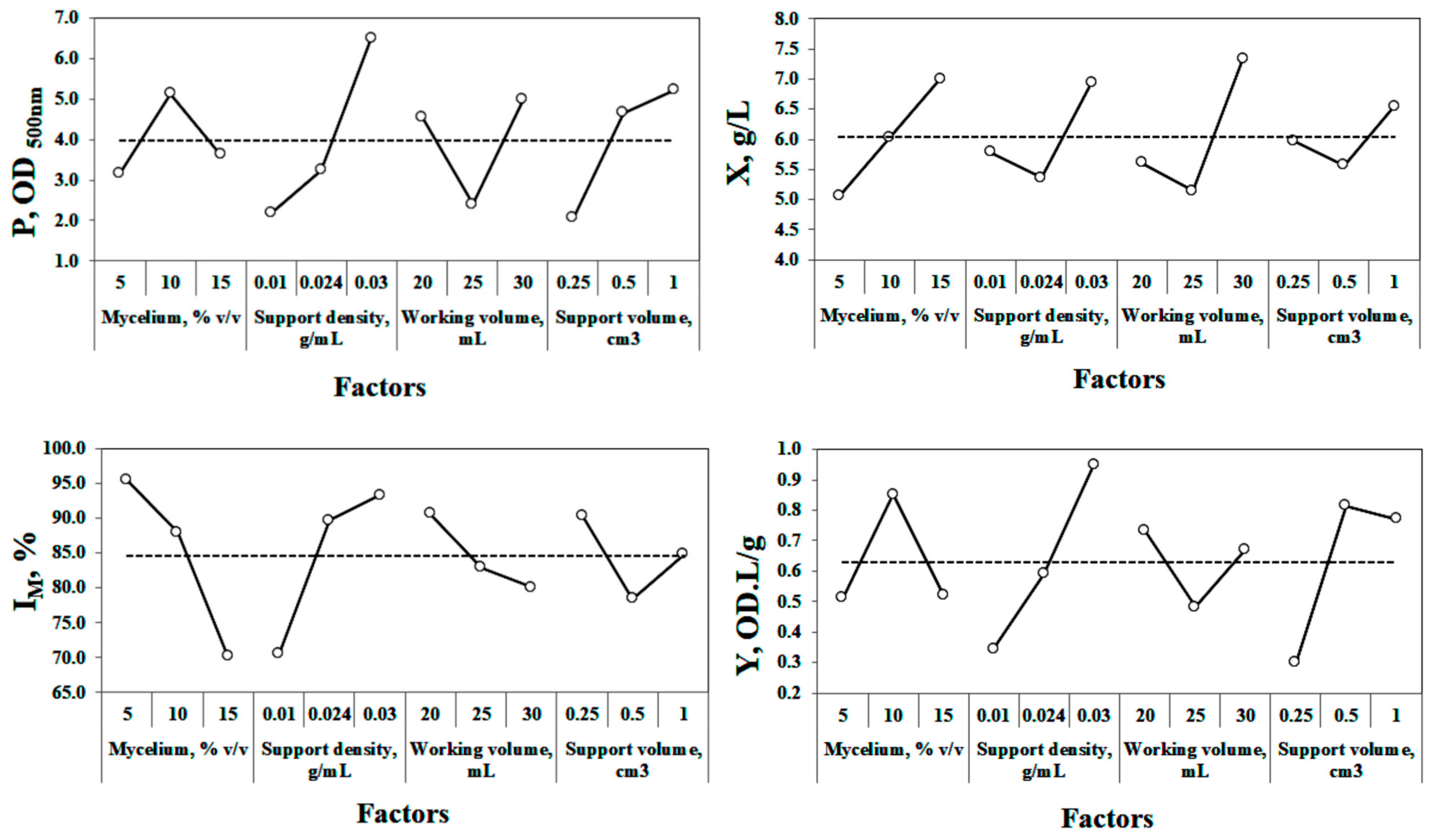
3.2.3. Analysis of Individual Factors
3.2.4. Validation Trials
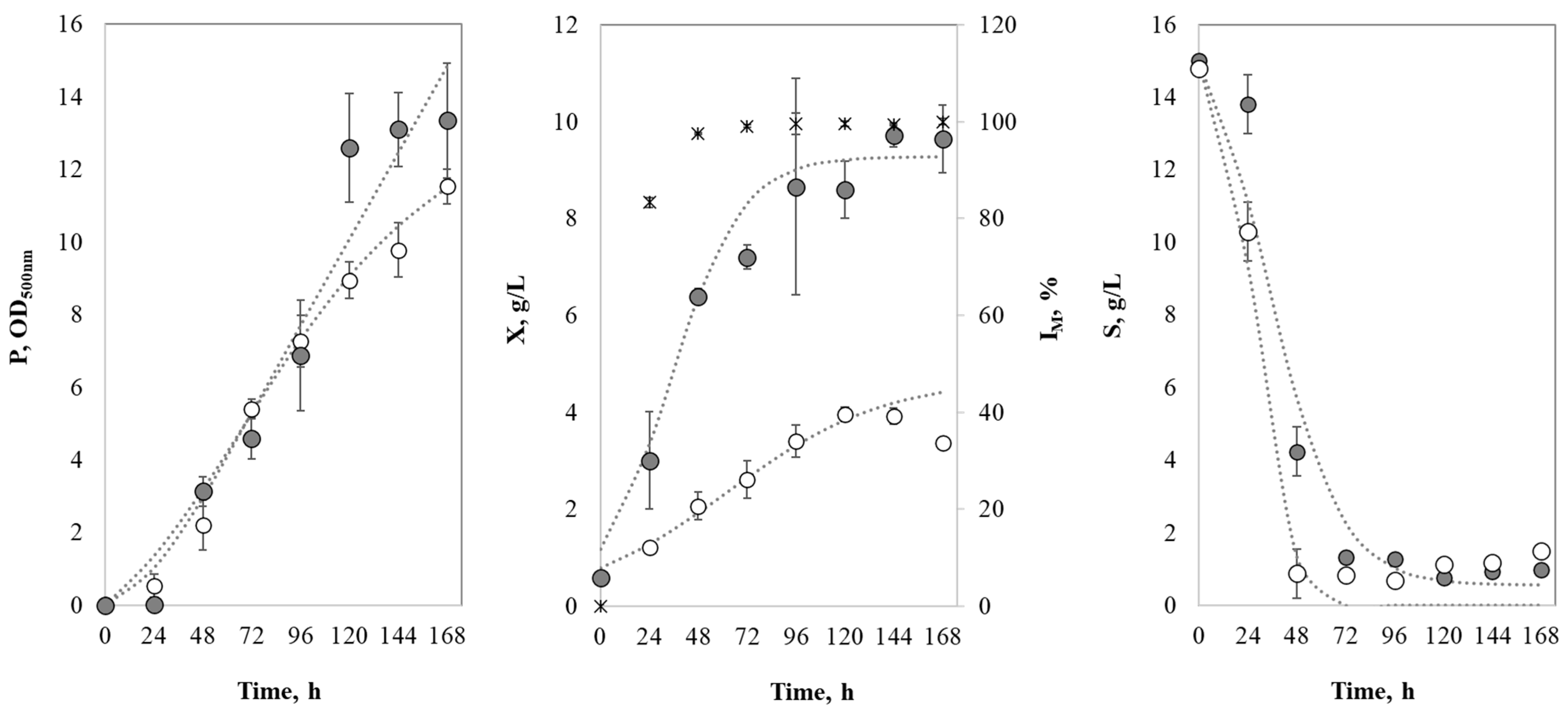
3.3. Process Kinetics Evaluation
3.3.1. Process Kinetics and Model Fitting Using Free and Immobilized Biomass
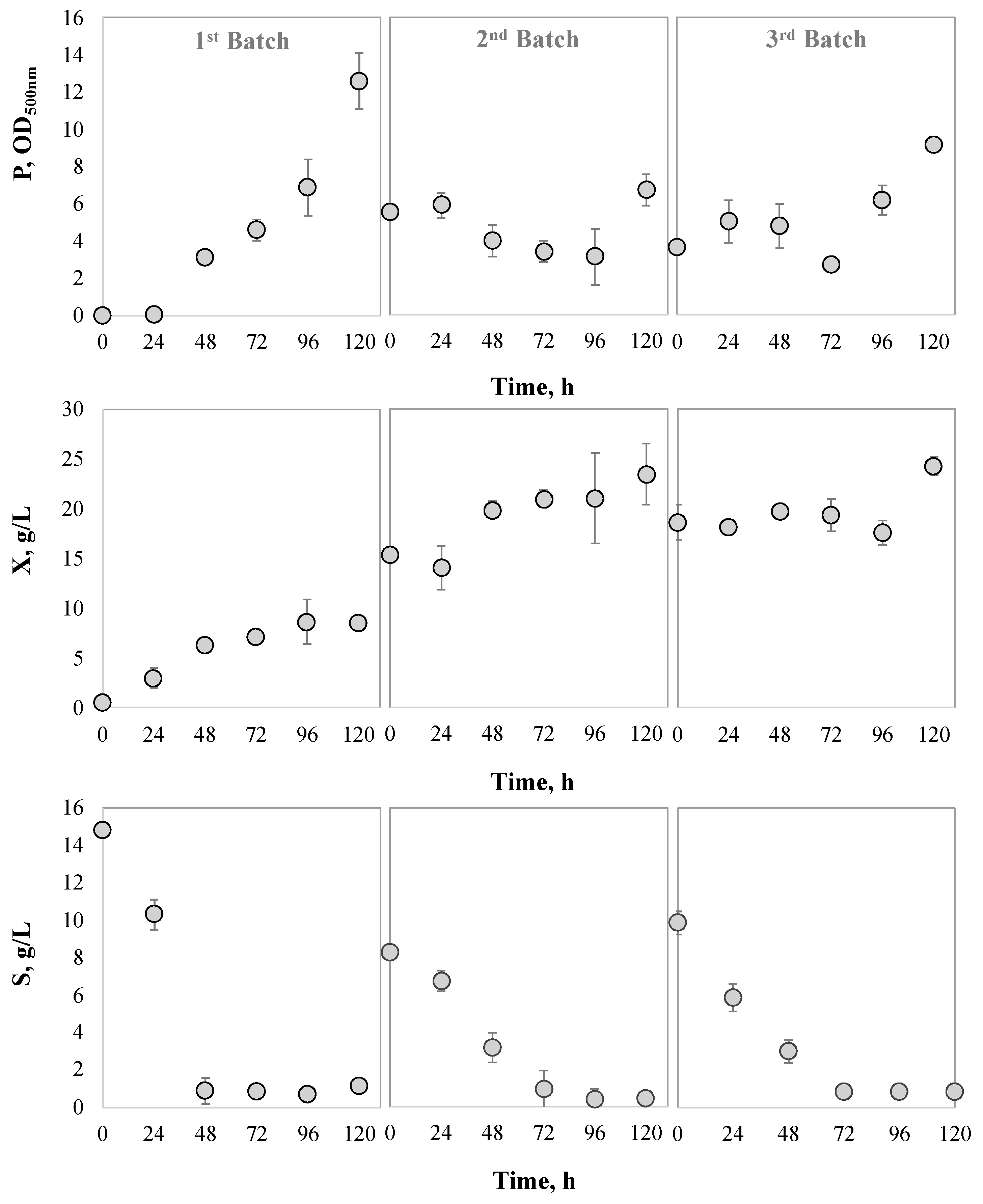
3.3.2. Cell Recycling Assessment during Consecutive Batches
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aman Mohammadi, M.; Ahangari, H.; Mousazadeh, S.; Hosseini, S.M.; Dufossé, L. Microbial Pigments as an Alternative to Synthetic Dyes and Food Additives: A Brief Review of Recent Studies. Bioprocess Biosyst. Eng. 2022, 45, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, M.; Zelena, M.; Cacciola, F.; Ceslova, L.; Girard-Valenciennes, E.; Clerc, P.; Dugo, P.; Mondello, L.; Fouillaud, M.; Rotondo, A.; et al. Partial Characterization of the Pigments Produced by the Marine-Derived Fungus Talaromyces Albobiverticillius 30548. Towards a New Fungal Red Colorant for the Food Industry. J. Food Compos. Anal. 2018, 67, 38–47. [Google Scholar] [CrossRef]
- Gomah, N.; Abdel-Raheam, H. Stability and Components Identification of Monascus Ruber’s Pigments. J. Food Dairy Sci. 2017, 8, 31–36. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, J.; Huang, Y.; Xin, Q.; Wang, Z. Diversifying of Chemical Structure of Native Monascus Pigments. Front. Microbiol. 2018, 9, 3143. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Feng, Y.; Molnár, I.; Chen, F. Nature and Nurture: Confluence of Pathway Determinism with Metabolic and Chemical Serendipity Diversifies: Monascus Azaphilone Pigments. Nat. Prod. Rep. 2019, 36, 561–572. [Google Scholar] [CrossRef]
- Tsiailanis, A.D.; Pateraki, C.; Kyriazou, M.; Chatzigiannis, C.M.; Chatziathanasiadou, M.; Parisis, N.; Mandala, I.; Tzakos, A.G.; Koutinas, A. Chemical Profiling, Bioactivity Evaluation and the Discovery of a Novel Biopigment Produced by Penicillium Purpurogenum Cbs 113139. Molecules 2021, 27, 69. [Google Scholar] [CrossRef] [PubMed]
- Peter, A.P.; Koyande, A.K.; Chew, K.W.; Ho, S.H.; Chen, W.H.; Chang, J.S.; Krishnamoorthy, R.; Banat, F.; Show, P.L. Continuous Cultivation of Microalgae in Photobioreactors as a Source of Renewable Energy: Current Status and Future Challenges. Renew. Sustain. Energy Rev. 2022, 154, 111852. [Google Scholar] [CrossRef]
- Villegas-Méndez, M.Á.; Aguilar-Machado, D.E.; Balagurusamy, N.; Montañez, J.; Morales-Oyervides, L. Agro-Industrial Wastes for the Synthesis of Carotenoids by Xanthophyllomyces dendrorhous: Mesquite Pods-Based Medium Design and Optimization. Biochem. Eng. J. 2019, 150, 107260. [Google Scholar] [CrossRef]
- Morales-Oyervides, L.; Ruiz-Sánchez, J.P.; Oliveira, J.C.; Sousa-Gallagher, M.J.; Méndez-Zavala, A.; Giuffrida, D.; Dufossé, L.; Montañez, J. Biotechnological Approaches for the Production of Natural Colorants by Talaromyces/Penicillium: A Review. Biotechnol. Adv. 2020, 43, 107601. [Google Scholar] [CrossRef]
- Maswanna, T.; Phunpruch, S.; Lindblad, P.; Maneeruttanarungroj, C. Enhanced Hydrogen Production by Optimization of Immobilized Cells of the Green Alga Tetraspora Sp. CU2551 Grown under Anaerobic Condition. Biomass Bioenergy 2018, 111, 88–95. [Google Scholar] [CrossRef]
- Rasheed, A.; Ghous, T.; Mumtaz, S.; Zafar, M.N.; Akhter, K.; Shabir, R.; Abdin, Z.U.; Shafqat, S.S. Immobilization of Pseudomonas aeruginosa Static Biomass on Eggshell Powder for On-Line Preconcentration and Determination of Cr (VI). Open Chem. 2020, 18, 303–313. [Google Scholar] [CrossRef]
- Wang, J.; Liang, J.; Sun, L.; Li, G.; Temmink, H.; Rijnaarts, H.H.M. Granule-Based Immobilization and Activity Enhancement of Anammox Biomass via PVA/CS and PVA/CS/Fe Gel Beads. Bioresour. Technol. 2020, 309, 123448. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Bisson, L.F.; García-Martínez, T.; Mauricio, J.C.; Moreno-García, J. New Insights on Yeast and Filamentous Fungus Adhesion in a Natural Co-Immobilization System: Proposed Advances and Applications in Wine Industry. Appl. Microbiol. Biotechnol. 2019, 103, 4723–4731. [Google Scholar] [CrossRef] [PubMed]
- Hua, X.; Du, G.; Xu, Y. Cost-Practical of Glycolic Acid Bioproduction by Immobilized Whole-Cell Catalysis Accompanied with Compressed Oxygen Supplied to Enhance Mass Transfer. Bioresour. Technol. 2019, 283, 326–331. [Google Scholar] [CrossRef]
- Liu, J.; Guo, T.; Luo, Y.; Chai, X.; Wu, J.; Zhao, W.; Jiao, P.; Luo, F.; Lin, Q. Enhancement of Monascus Pigment Productivity via a Simultaneous Fermentation Process and Separation System Using Immobilized-Cell Fermentation. Bioresour. Technol. 2019, 272, 552–560. [Google Scholar] [CrossRef]
- Arikan, E.B.; Isik, Z.; Bouras, H.D.; Dizge, N.; Kaleem, I.; Shen, H.; Lv, B.; Wei, B.; Rasool, A.; Li, C.; et al. Microbial Cell Immobilization in Biohydrogen Production: A Short Overview. Bioresour. Technol. 2019, 144, 37–42. [Google Scholar] [CrossRef]
- Oliveira, J.C.; Montañez, J.C.; Méndez-Zavala, A.; Morales-Oyervides, L.; Sousa-Gallagher, M.J. Selection of Best Conditions of Inoculum Preparation for Optimum Performance of the Pigment Production Process by Talaromyces Spp. Using the Taguchi Method. Biotechnol. Prog. 2017, 33, 621–632. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Yilmaz, N.; Thrane, U.; Rasmussen, K.B.; Houbraken, J.; Samson, R.A. Talaromyces atroroseus, a New Species Efficiently Producing Industrially Relevant Red Pigments. PLoS ONE 2013, 8, e84102. [Google Scholar] [CrossRef]
- Zavala, A.M. Identification of factors affecting the pigment production by Penicillium purpurogenum GH2 and chromatographic profiles obtention. Master’s Thesis, Universidad A utónoma de Coahuila, Saltillo, México, 2011. [Google Scholar]
- Castro, C.C.; Nobre, C.; Duprez, M.E.; De Weireld, G.; Hantson, A.L. Screening and Selection of Potential Carriers to Immobilize Aureobasidium pullulans Cells for Fructo-Oligosaccharides Production. Biochem. Eng. J. 2017, 118, 82–90. [Google Scholar] [CrossRef]
- Méndez, A.; Pérez, C.; Montañéz, J.C.; Martínez, G.; Aguilar, C.N. Red Pigment Production by Penicillium purpurogenum GH2 Is Influenced by PH and Temperature. J. Zhejiang Univ. Sci. B 2011, 12, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Gilles, K.; Hamilton, J. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 300–306. [Google Scholar] [CrossRef]
- Butler, C. A Primer on the Taguchi Method; Society of Manufacturing Engineers: Southfield, MI, USA, 2003; Volume 5, ISBN 0872638642. [Google Scholar]
- Weiss, R.M.; Ollis, D.F. Extracellular Microbial Polysaccharides. I. Substrate, Biomass, and Product Kinetic Equations for Batch Xanthan Gum Fermentation. Biotechnol. Bioeng. 1980, 22, 859–873. [Google Scholar] [CrossRef]
- Luedeking, R.; Piret, E.L. Kinetic Study of the Lactic Acid Fermentation. Batch Process at Controlled PH. Biotechnol. Bioeng. 2000, 67, 636–644. [Google Scholar] [CrossRef]
- Rajendran, A.; Thangavelu, V. Statistical Experimental Design for Evaluation of Medium Components for Lipase Production by Rhizopus arrhizus MTCC 2233. LWT-Food Sci. Technol. 2009, 42, 985–992. [Google Scholar] [CrossRef]
- Kurade, M.B.; Waghmode, T.R.; Xiong, J.Q.; Govindwar, S.P.; Jeon, B.H. Decolorization of Textile Industry Effluent Using Immobilized Consortium Cells in Upflow Fixed Bed Reactor. J. Clean. Prod. 2019, 213, 884–891. [Google Scholar] [CrossRef]
- Zhang, J.; Wei, J.; Massey, I.Y.; Peng, T.; Yang, F. Immobilization of Microbes for Biodegradation of Microcystins: A Mini Review. Toxins 2022, 14, 573. [Google Scholar] [CrossRef]
- Nirmal, L.A.; Vishal, R.; Bhakthochidan, S.A.; Roshini, V.B.; Jacob, S. Cell Leakage Minimization by Immobilization Modulation of Chlorella sorokiniana NCIM 5561 and Phosphate Removal from Wastewater. Int. J. Environ. Sci. Technol. 2022, 19, 3015–3030. [Google Scholar] [CrossRef]
- de Oliveira, C.F.D.; da Costa, J.P.V.; Vendruscolo, F. Maltose Syrup Residue as the Substrate for Monascus pigments Production. Biocatal. Agric. Biotechnol. 2019, 18, 101101. [Google Scholar] [CrossRef]
- Ha, N.T.H.; Toan, N.C.; Kajitvichyanukul, P. Enhanced Paraquat Removal from Contaminated Water Using Cell-Immobilized Biochar. Clean Technol. Environ. Policy 2022, 24, 1073–1085. [Google Scholar] [CrossRef]
- Santaolalla, A.; Gutierrez, J.; Gallastegui, G.; Barona, A.; Rojo, N. Immobilization of Acidithiobacillus perrooxidans in Bacterial Cellulose for a More Sustainable Bioleaching Process. J. Environ. Chem. Eng. 2021, 9, 105283. [Google Scholar] [CrossRef]
- Bahry, H.; Abdalla, R.; Pons, A.; Taha, S.; Vial, C. Optimization of Lactic Acid Production Using Immobilized Lactobacillus rhamnosus and Carob Pod Waste from the Lebanese Food Industry. J. Biotechnol. 2019, 306, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, T.; Dev, S.; Banerjee, A.; Chatterjee, A.; Singh, R.; Aggarwal, S. Use of Immobilized Bacteria for Environmental Bioremediation: A Review. J. Environ. Chem. Eng. 2021, 9, 105920. [Google Scholar] [CrossRef]
- Naramittanakul, A.; Buttranon, S.; Petchsuk, A.; Chaiyen, P.; Weeranoppanant, N. Development of a Continuous-Flow System with Immobilized Biocatalysts towards Sustainable Bioprocessing. React. Chem. Eng. 2021, 6, 1771–1790. [Google Scholar] [CrossRef]
- Gao, H.; Lu, J.; Jiang, Y.; Fang, Y.; Tang, Y.; Yu, Z.; Zhang, W.; Xin, F.; Jiang, M. Material-Mediated Cell Immobilization Technology in the Biological Fermentation Proces. Biofuels Bioprod. Biorefining 2021, 15, 1160–1173. [Google Scholar] [CrossRef]
- Molasses, S. Capability of Immobilized Clostridium beijerinckii TISTR 1461. Processes 2021, 9, 573. [Google Scholar]
- Ugwu, C.T.; Ogbonna, C.N.; Ogbonna, J.C.; Aoyagi, H. Production and Stability of Pigments by Talaromyces purpurogenus LC128689 in an Alternating Air Phase–liquid Phase Cultivation System. Biotechnol. Appl. Biochem. 2022, 69, 1317–1326. [Google Scholar] [CrossRef]
- Maslova, O.; Stepanov, N.; Senko, O.; Efremenko, E. Production of Various Organic Acids from Different Renewable Sources by Immobilized Cells in the Regimes of Separate Hydrolysis and Fermentation (SHF) and Simultaneous Saccharification and Fermentation (SFF). Bioresour. Technol. 2019, 272, 1–9. [Google Scholar] [CrossRef]
- Widayanti, V.T.; Estiasih, T.; Zubaidah, E.; Taher, M. The Role of Carbon and Nitrogen Sources in the Production of Bioactive Compounds in Monascus Fermentation Products: A Mini Review. IOP Conf. Ser. Earth Environ. Sci. 2021, 924, 012083. [Google Scholar] [CrossRef]
- Basto, B.; da Silva, N.R.; Teixeira, J.A.; Silvério, S.C. Production of Natural Pigments by Penicillium brevicompactum Using Agro-Industrial Byproducts. Fermentation 2022, 8, 536. [Google Scholar] [CrossRef]
- Huang, Z.-R.; Zhou, W.-B.; Yang, X.-L.; Tong, A.-J.; Hong, J.-L.; Guo, W.-L.; Li, T.-T.; Jia, R.-B.; Pan, Y.-Y.; Lin, J.; et al. The Regulation Mechanisms of Soluble Starch and Glycerol for Production of Azaphilone Pigments in Monascus purpureus FAFU618 as Revealed by Comparative Proteomic and Transcriptional Analyses. Food Res. Int. 2018, 106, 626–635. [Google Scholar] [CrossRef]
- Morales-Oyervides, L.; Ruiz-Sánchez, J.P.; Oliveira, J.C.; Sousa-Gallagher, M.J.; Morales-Martínez, T.K.; Albergamo, A.; Salvo, A.; Giuffrida, D.; Dufossé, L.; Montañez, J. Medium Design from Corncob Hydrolyzate for Pigment Production by Talaromyces atroroseus GH2: Kinetics Modeling and Pigments Characterization. Biochem. Eng. J. 2020, 161, 107698. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Batch | Supports | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | ||||||||||
| P, OD500nm | XL, g/L | X, g/L | IM, % | P, OD500nm | XL, g/L | X, g/L | IM, % | P, OD500nm | XL, g/L | X, g/L | IM, % | |
| 1 | b 3.00 ± 1.12 | a 4.97 ± 1.97 | - | - | b 2.88 ± 3.43 | b 0.48 ± 0.25 | - | - | a 8.63 ± 0.69 | b 1.35 ± 0.24 | - | - |
| 2 | a 9.54 ± 4.99 | a 3.55 ± 1.18 | - | - | a 2.41 ± 1.51 | a 1.12 ± 0.66 | - | - | a 15.86 ± 9.98 | a 2.91 ± 1.07 | - | - |
| 3 | a 2.68 ± 0.71 | a 1.85 ± 0.48 | a 13.01 ± 2.02 | a 85.90 ± 1.37 | a 2.73 ± 2.40 | a 2.00 ± 1.42 | a 13.68 ± 1.88 | a 85.35 ± 10.03 | a 4.56 ± 2.36 | a 2.96 ± 1.01 | a 14.76 ± 1.50 | a 79.73 ± 7.17 |
| Actual | Experimental Results | |||||||
|---|---|---|---|---|---|---|---|---|
| Run | Mycelium, % v/v | Support Density, g/mL | Working Volume, mL | Support Volume, cm3 | P, OD500nm | IM, % | X, g/L | Y, OD.L/g |
| 1 | 5 | 0.01 | 20 | 0.25 | 0.02 ± 0.03 | 93.50 ± 2.82 | 4.34 ± 0.14 | 0.00 ± 0.01 |
| 2 | 5 | 0.024 | 25 | 0.5 | 1.55 ± 0.35 | 92.83 ± 2.39 | 3.04 ± 0.17 | 0.51 ± 0.14 |
| 3 | 5 | 0.03 | 30 | 1 | 7.94 ± 0.03 | 100.00 ± 0.00 | 7.80 ± 0.28 | 1.02 ± 0.03 |
| 4 | 10 | 0.01 | 25 | 1 | 3.00 ± 1.81 | 72.68 ± 0.52 | 5.42 ± 0.31 | 0.56 ± 0.37 |
| 5 | 10 | 0.024 | 30 | 0.25 | 3.50 ± 1.41 | 94.56 ± 0.12 | 6.62 ± 0.14 | 0.53 ± 0.20 |
| 6 | 10 | 0.03 | 20 | 0.5 | 8.89 ± 2.67 | 96.72 ± 2.01 | 4.26 ± 2.40 | 2.69 ± 2.15 |
| 7 | 15 | 0.01 | 30 | 0.5 | 3.53 ± 0.69 | 45.73 ± 4.93 | 7.60 ± 0.17 | 0.47 ± 0.10 |
| 8 | 15 | 0.024 | 20 | 1 | 4.70 ± 0.48 | 81.80 ± 9.82 | 6.44 ± 0.57 | 0.73 ± 0.01 |
| 9 | 15 | 0.03 | 25 | 0.25 | 2.66 ± 1.75 | 83.15 ± 10.22 | 6.98 ± 0.71 | 0.37 ± 0.21 |
| Factor | Responses | ||||
|---|---|---|---|---|---|
| P, OD500nm | X, g/L | Y, OD.L/g | IM, % | ||
| Contribution | Mycelium, % v/v | 1.15 | 0.00 | 0.22 | 3.43 |
| Support density, g/mL | 2.52 | 0.91 | 0.32 | 8.74 | |
| Working volume, mL | 1.01 | 1.31 | 0.04 | −4.45 | |
| Support volume, cm3 | 1.24 | 0.52 | 0.14 | 0.27 | |
| Expected response | Minimum | 8.15 | 8.32 | 1.08 | 85.81 |
| Maximum | 11.65 | 9.23 | 1.64 | 99.28 | |
| Experimental | 13.17 ± 1.52 | 9.68 ± 0.47 | 1.36 ± 0.15 | 99.01 ± 0.37 | |
| Parameter | Control | Immobilized |
|---|---|---|
| , g/L | 0.79 | 0.81 |
| , g/L | 3.75 | 9.32 |
| , g/L | 15.00 | 15.00 |
| , OD500nm | 0.00 | 0.00 |
| , h−1 | 0.03 | 0.06 |
| 2.53 | 0.22 | |
| 0.00 | 0.01 | |
| , gBiomass/gSubstrate | 0.26 | 0.65 |
| , OD/gSubstrate | 0.89 | 0.93 |
| 0.09 | 0.00 | |
| 0.75 | 0.91 | |
| 0.95 | 0.96 | |
| 0.99 | 0.93 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Sánchez, J.P.; Morales-Oyervides, L.; Giuffrida, D.; Dufossé, L.; Montañez, J.C. Production of Pigments under Submerged Culture through Repeated Batch Fermentation of Immobilized Talaromyces atroroseus GH2. Fermentation 2023, 9, 171. https://doi.org/10.3390/fermentation9020171
Ruiz-Sánchez JP, Morales-Oyervides L, Giuffrida D, Dufossé L, Montañez JC. Production of Pigments under Submerged Culture through Repeated Batch Fermentation of Immobilized Talaromyces atroroseus GH2. Fermentation. 2023; 9(2):171. https://doi.org/10.3390/fermentation9020171
Chicago/Turabian StyleRuiz-Sánchez, Juan Pablo, Lourdes Morales-Oyervides, Daniele Giuffrida, Laurent Dufossé, and Julio César Montañez. 2023. "Production of Pigments under Submerged Culture through Repeated Batch Fermentation of Immobilized Talaromyces atroroseus GH2" Fermentation 9, no. 2: 171. https://doi.org/10.3390/fermentation9020171