
Influence of Long-Term Agar-Slant Preservation at 4 °C on the Recombinant Enzyme Activity of Engineered Yeast

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
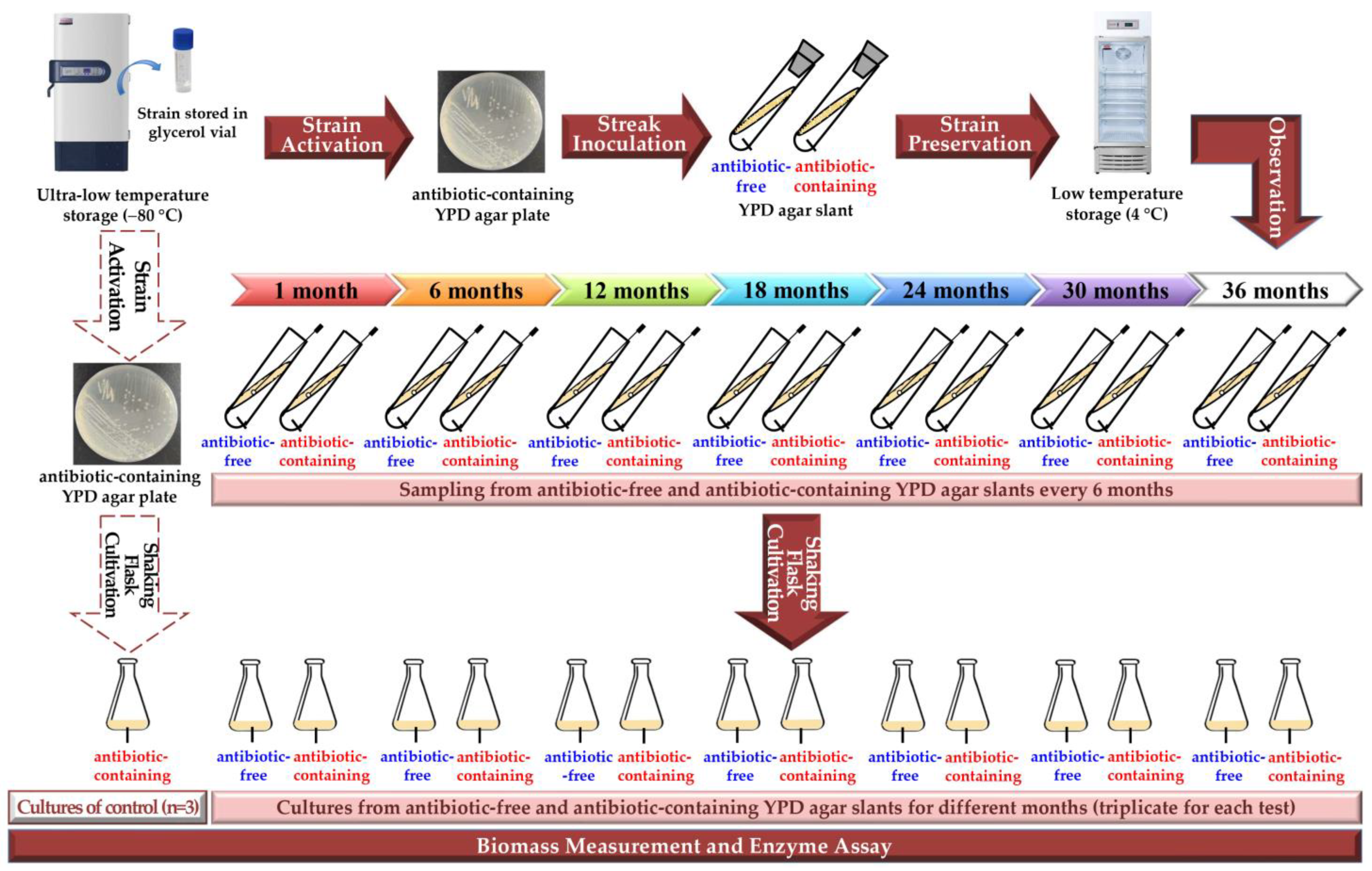
2.1. Strain and Media
2.2. Agar-Slant Preparation
2.3. Strain Preservation
2.4. Shaking Flask Cultivation
2.5. Biomass Measurement and Enzyme Assay
2.6. Statistical Analysis
3. Results
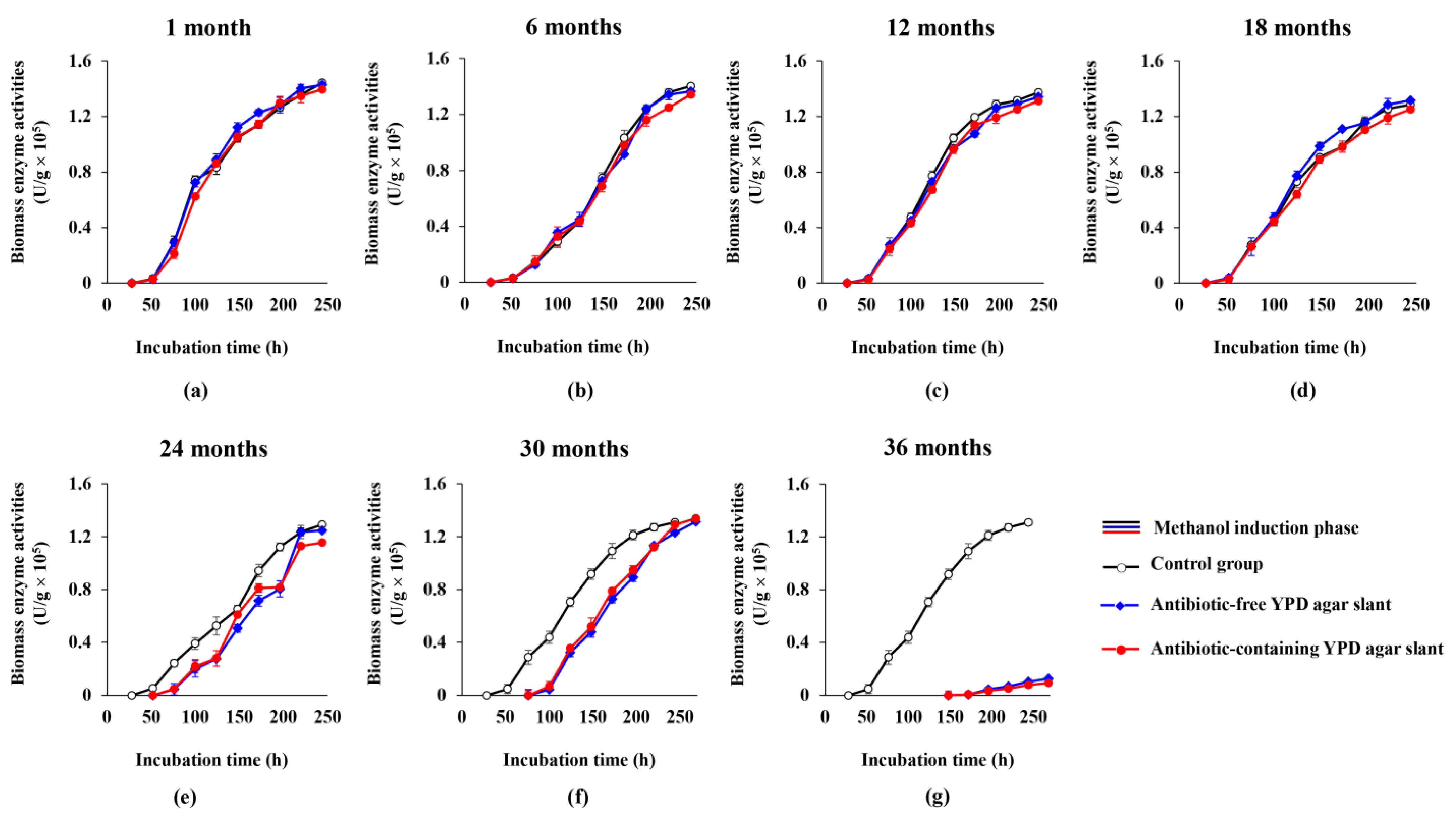
3.1. Cell Growth Ability of the Engineered Yeast GS115-3.5K-P1-2 in the Slants Preserved at 4 °C
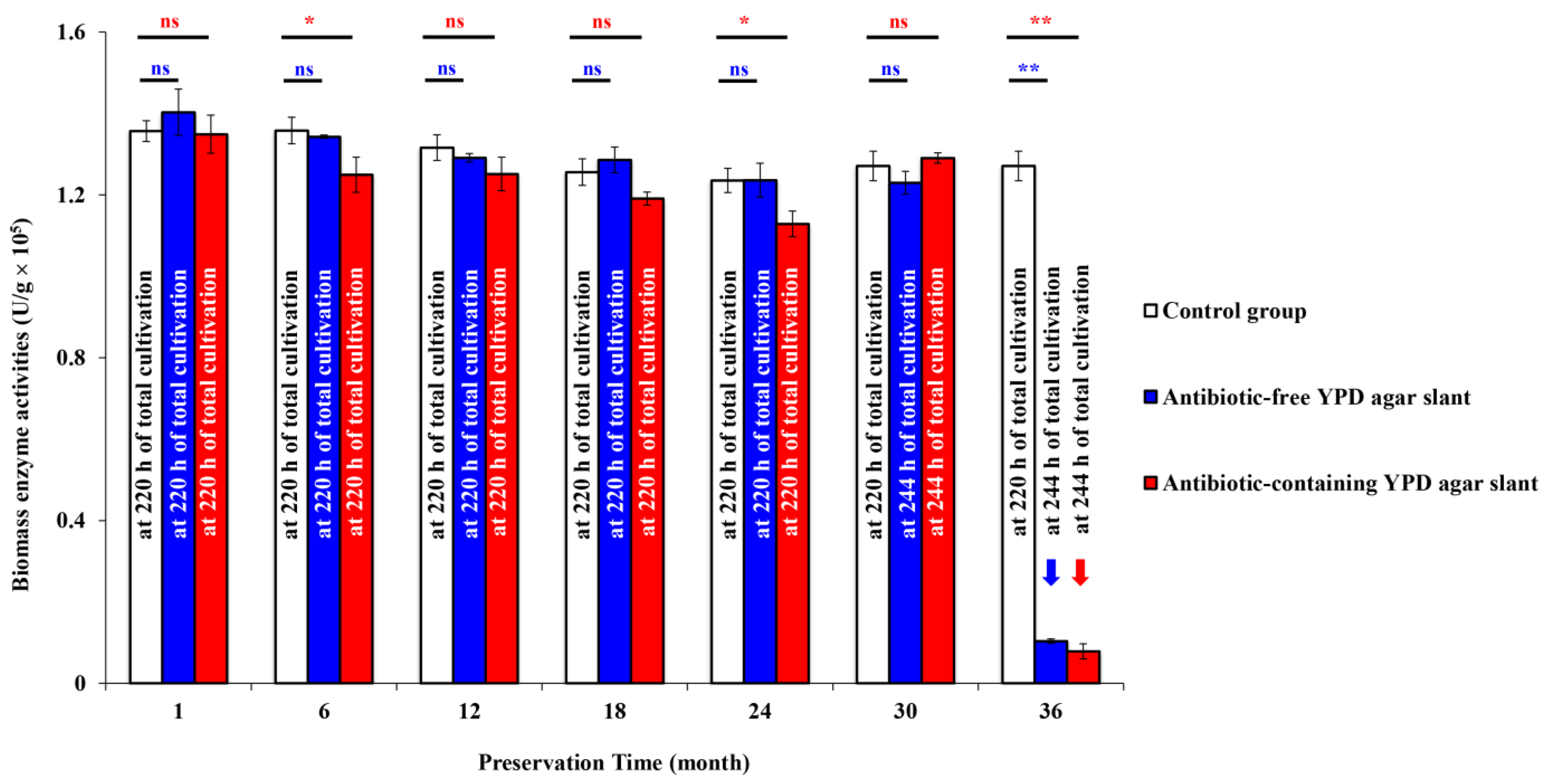
3.2. Biomass Enzyme Activity of the Engineered Yeast GS115-3.5K-P1-2 in the Slants Preserved at 4 °C
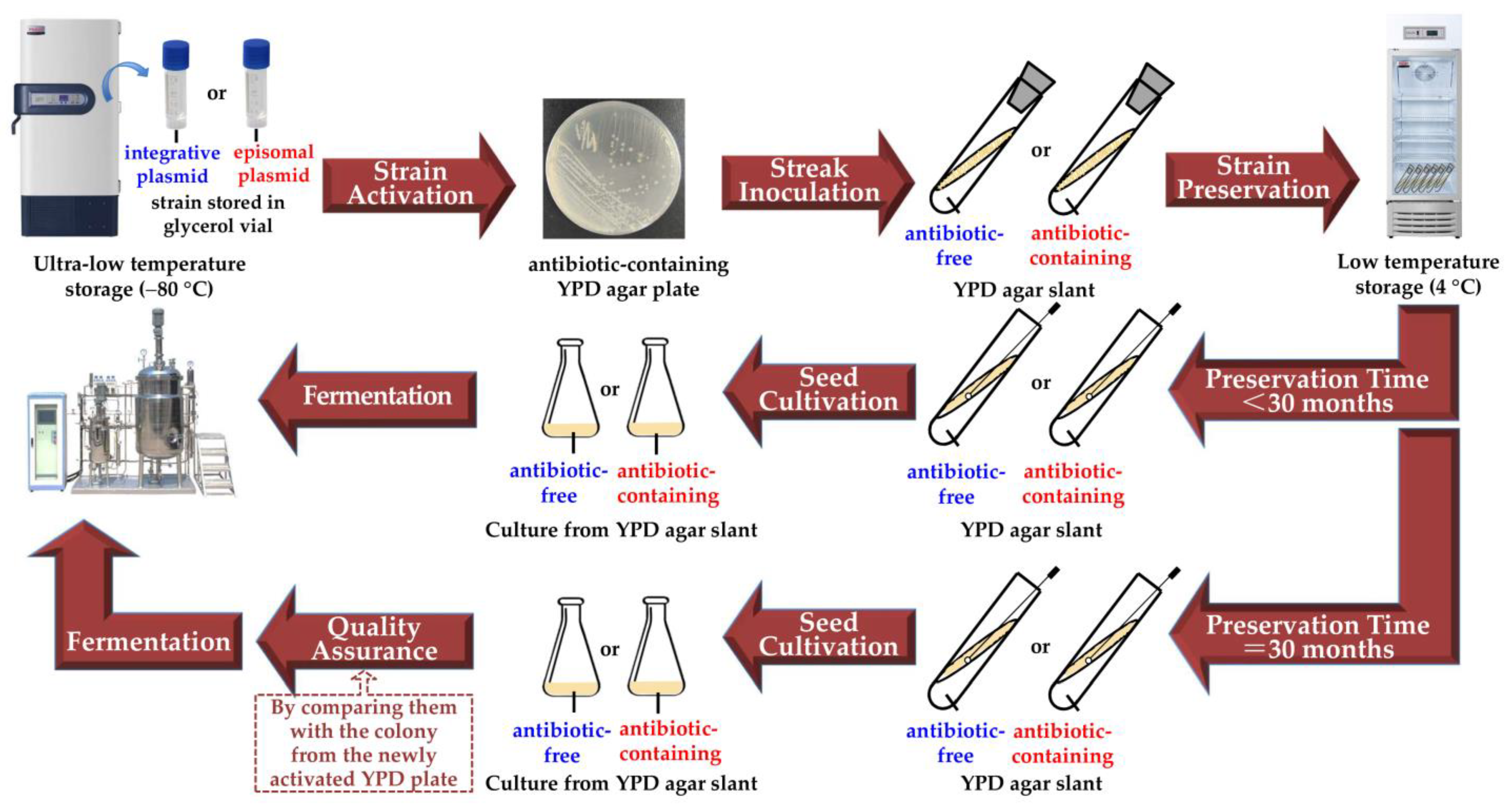
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cregg, J.M.; Vedvick, T.S.; Raschke, W.C. Recent advances in the expression of foreign genes in Pichia pastoris. Bio/Technology 1993, 11, 905–910. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.X.; Yi, P.; Liu, J.; Yan, Q.J.; Jiang, Z.Q. High-level expression of an engineered β-mannanase (mRmMan5A) in Pichia pastoris for manno-oligosaccharide production using steam explosion pretreated palm kernel cake. Bioresour. Technol. 2018, 256, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Werten, M.W.; Eggink, G.; Stuart, M.A.C.; Wolf, F.A. Production of protein-based polymers in Pichia pastoris. Biotechnol. Adv. 2019, 37, 642–666. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.M.; Ning, C.; Yuan, M.X.; Fu, X.D.; Yang, S.X.; Wei, X.Y.; Xiao, M.S.; Mou, H.J.; Zhu, C.L. High-efficiency expression of a superior β-mannanase engineered by cooperative substitution method in Pichia pastoris and its application in preparation of prebiotic mannooligosaccharides. Bioresour. Technol. 2020, 311, 123482. [Google Scholar] [CrossRef] [PubMed]
- Cai, P.; Duan, X.P.; Wu, X.Y.; Gao, L.H.; Ye, M.; Zhou, Y.J. Recombination machinery engineering facilitates metabolic engineering of the industrial yeast Pichia pastoris. Nucleic Acids Res. 2021, 49, 7791–7805. [Google Scholar] [CrossRef]
- Ding, J.J.; You, S.P.; Zhang, J.X.; Zhang, H.T.; Wang, H.; Zhang, W.; Qi, W.; Su, R.X.; He, Z.M. Rational design of 17β-hydroxysteroid dehydrogenase type3 for improving testosterone production with an engineered Pichia pastoris. Bioresour. Technol. 2021, 341, 125833. [Google Scholar] [CrossRef]
- Jin, X.R.; Zhang, W.J.; Wang, Y.; Sheng, J.Y.; Xu, R.R.; Li, J.H.; Du, G.C.; Kang, Z. Biosynthesis of non-animal chondroitin sulfate from methanol using genetically engineered Pichia pastoris. Green Chem. 2021, 23, 4365–4374. [Google Scholar] [CrossRef]
- Shao, Y.R.; Xue, C.L.; Liu, W.Q.; Zuo, S.Q.; Wei, P.L.; Huang, L.; Lian, J.Z.; Xu, Z.N. High-level secretory production of leghemoglobin in Pichia pastoris through enhanced globin expression and heme biosynthesis. Bioresour. Technol. 2022, 363, 127884. [Google Scholar] [CrossRef]
- Mohammadzadeh, R.; Karbalaei, M.; Soleimanpour, S.; Mosavat, A.; Rezaee, S.A.; Ghazvini, K.; Farsiani, H. Practical methods for expression of recombinant protein in the Pichia pastoris system. Curr. Protoc. 2021, 1, e155. [Google Scholar] [CrossRef]
- User Manual of Multi-Copy Pichia Expression Kit for the Isolation and Expression of Recombinant Proteins from Pichia Pastoris Strains Containing Multiple Copies of a Particular Gene; Cat.no.K1750-01., Manual part no.25-0170; Invitrogen TM: Waltham, MA, USA, 2010.
- Athmaram, T.N.; Singh, A.K.; Saraswat, S.; Srivastava, S.; Misra, P.; Kameswara Rao, M.; Gopalan, N.; Rao, P.V.L. A simple Pichia pastoris fermentation and downstream processing strategy for making recombinant pandemic Swine Origin Influenza a virus Hemagglutinin protein. J. Ind. Microbiol. Biotechnol. 2013, 40, 245–255. [Google Scholar] [CrossRef]
- Onions, A.H.S. Chapter IV preservation of fungi. Methods Microbiol. 1971, 4, 113–151. [Google Scholar]
- Lei, Y.P. Preserving bacteria at low temperature involves inoculating purified bacteria into suitable solid slant culture medium, and performing culturing process, sealing cultured product using sealing film and refrigerating product. Chinese Patent CN106609244-A, 2017. [Google Scholar]
- Lei, Y.P. Preservation of low temperature-bacterial slant culture involves storing pure bacterial inoculum in solid slant medium, culturing under preset conditions, sealing with parafilm, and storing at preset temperature. Chinese Patent CN105695332-A, 2016. [Google Scholar]
- Schneider, F.; Pan, L.; Ottenbruch, M.; List, T.; Gaich, T. The chemistry of nonclassical taxane diterpene. Acc. Chem. Res. 2021, 54, 2347–2360. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, L.Y.; Zhuang, W.B.; Zhang, F.J.; Shu, X.C.; Wang, N.; Wang, Z. Recent research progress in taxol biosynthetic pathway and acylation reactions mediated by Taxus acyltransferases. Molecules 2021, 26, 2855. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, S.K.; Sharma, R.P.; Kumar, S.; Madhusudanan, K.P. A process for the production of Taxol. EP Patent 090 5130 B1, 2002. [Google Scholar]
- Li, B.J.; Wang, H.; Gong, T.; Chen, J.J.; Chen, T.J.; Yang, J.L.; Zhu, P. Improving 10-deacetylbaccatin III-10-β-O-acetyltransferase catalytic fitness for Taxol production. Nat. Commun. 2017, 8, 15544. [Google Scholar] [CrossRef] [Green Version]
- Rao, K.V. Semi-synthesis of paclitaxel from naturally occurring glycosidic precursors. J. Heterocycl. Chem. 1997, 34, 675–680. [Google Scholar] [CrossRef]
- Cheng, H.L.; Zhao, R.Y.; Chen, T.J.; Yu, W.B.; Wang, F.; Cheng, K.D.; Zhu, P. Cloning and characterization of the glycoside hydrolases that remove xylosyl groups from 7-β-xylosyl-10-deacetyltaxol and its analogues. Mol. Cell. Proteomics. 2013, 12, 2236–2248. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.B.; Liang, X.; Zhu, P. High-cell-density fermentation and pilot-scale biocatalytic studies of an engineered yeast expressing the heterologous glycoside hydrolase of 7-β-xylosyltaxanes. J. Ind. Microbiol. Biotechnol. 2013, 40, 133–140. [Google Scholar] [CrossRef]
- Liu, W.C.; Zhu, P. Pilot studies on scale-up biocatalysis of 7-β-xylosyl-10-deacetyltaxol and its analogues by an engineered yeast. J. Ind. Microbiol. Biotechnol. 2015, 42, 867–876. [Google Scholar] [CrossRef]
- Liu, W.C.; Gong, T.; Wang, Q.H.; Liang, X.; Chen, J.J.; Zhu, P. Scaling-up fermentation of Pichia pastoris to demonstration-scale using new methanol-feeding strategy and increased air pressure instead of pure oxygen supplement. Sci. Rep. 2016, 6, 18439. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.J.; Liang, X.; Li, H.X.; Chen, T.J.; Zhu, P. Improving the catalytic property of the glycoside hydrolase LXYL-P1-2 by directed evolution. Molecules 2017, 22, 2133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.J.; Liang, X.; Wang, F.; Wen, Y.H.; Chen, T.J.; Liu, W.C.; Gong, T.; Yang, J.L.; Zhu, P. Combination mutation on the β-glycosidase specific to 7-β-xylosyltaxanes and increasing the mutated enzyme production by engineering the recombinant yeast. Acta Pharm. Sin. B 2019, 9, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Liang, X.; Chen, T.J.; Yang, J.L.; Zhu, P. Site-directed mutagenesis of a β-glycoside hydrolase from Lentinula edodes. Molecules 2019, 24, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tedeschi, R.; Paoli, P.D. Collection and preservation of frozen microorganisms. In Methods in biobanking; Humana Press: Totowa, NJ, USA, 2011; pp. 313–326. [Google Scholar]
- Cregg, J.M.; Higgins, D.R. Pichia Protocols; Humana Press: Totowa, NJ, USA, 2007. [Google Scholar]
- Blanquet, S.; Garrait, G.; Beyssac, E.; Perrier, C.; Denis, S.; H´ebrard, G.; Alric, M. Effects of cryoprotectants on the viability and activity of freeze dried recombinant yeasts as novel oral drug delivery systems assessed by an artificial digestive system. Eur. J. Pharm. Biopharm. 2005, 61, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Koštál, V.; Zahradníčková, H.; Šimek, P. Hyperprolinemic larvae of the drosophilid fly, Chymomyza costata, survive cryopreservation in liquid nitrogen. Proc. Natl. Acad. Sci. USA 2011, 108, 13041–13046. [Google Scholar] [CrossRef] [Green Version]
- Isachenko, E.; Isachenko, V.; Rahimi, G.; Nawroth, F. Cryopreservation of human ovarian tissue by direct plunging into liquid nitrogen. Eur. J. Obstet. Gynecol. Reprod. Biol. 2003, 108, 186–193. [Google Scholar] [CrossRef]
- Elliott, M.L. Survival, growth and pathogenicity of Gaeumannomyces graminis var. graminis with different methods of long-term storage. Mycologia 2005, 97, 901–907. [Google Scholar]
- Bond, C. Cryopreservation of yeast cultures. In Cryopreservation and Freeze-Drying Protocols; Humana Press: Totowa, NJ, USA, 2007; pp. 109–117. [Google Scholar]
- Alvarado-Fernández, A.M.; Rodríguez-López, E.A.; Espejo-Mojica, A.J.; Mosquera-Arévalo, A.R.; Alméciga-Díaz, C.J.; Trespalacios-Rangel, A.A. Effect of two preservation methods on the viability and enzyme production of a recombinant Komagataella phaffii (Pichia pastoris) strain. Cryobiology 2022, 105, 32–40. [Google Scholar] [CrossRef]
- Bryukhanov, A.L.; Netrusov, A.I. Long-term storage of obligate anaerobic microorganisms in glycerol. Appl. Biochem. Microbiol. 2006, 42, 177–180. [Google Scholar] [CrossRef]
- Siberry, G.; Brahmadathan, K.N.; Pandian, R.; Lalitha, M.K.; Steinhoff, M.C.; John, T.J. Comparison of different culture media and storage temperatures for the long-term preservation of Streptococcus pneumoniae in the tropics. Bull. World Health Organ. 2001, 79, 43–47. [Google Scholar]
- Day, J.; Stacey, G. Cryopreservation and Freeze-Drying Protocols; Humana Press: Totowa, NJ, USA, 2007; Volume 2. [Google Scholar]
- Miyamoto-Shinohara, Y.; Imaizumi, T.; Sukenobe, J.; Murakami, Y.; Kawamura, S.; Komatsu, Y. Survival rate of microbes after freeze-drying and long-term storage. Cryobiology 2000, 41, 251–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homolka, L.; Lisá, L. Long-term maintenance of fungal cultures on perlite in cryovials—an alternative for agar slants. Folia Microbiol. 2008, 53, 534–536. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, X.; Gong, T.; Chen, J.-J.; Chen, T.-J.; Yang, J.-L.; Zhu, P. Influence of Long-Term Agar-Slant Preservation at 4 °C on the Recombinant Enzyme Activity of Engineered Yeast. Fermentation 2023, 9, 104. https://doi.org/10.3390/fermentation9020104
Liang X, Gong T, Chen J-J, Chen T-J, Yang J-L, Zhu P. Influence of Long-Term Agar-Slant Preservation at 4 °C on the Recombinant Enzyme Activity of Engineered Yeast. Fermentation. 2023; 9(2):104. https://doi.org/10.3390/fermentation9020104
Chicago/Turabian StyleLiang, Xiao, Ting Gong, Jing-Jing Chen, Tian-Jiao Chen, Jin-Ling Yang, and Ping Zhu. 2023. "Influence of Long-Term Agar-Slant Preservation at 4 °C on the Recombinant Enzyme Activity of Engineered Yeast" Fermentation 9, no. 2: 104. https://doi.org/10.3390/fermentation9020104