
Growth Kinetics of Kazachstania unispora and Its Interaction with Lactic Acid Bacteria during Qymyz Production
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Growth Media Composition
2.3. Growth Experiments
2.3.1. Effect of Temperature
2.3.2. Effect of Carbohydrate Source and pH
2.3.3. LAB–K. unispora Coculture Experiments
2.3.4. Plate-Counting Methods for the Calibration Curve
2.4. Chemical Analyses
2.4.1. HPLC Analysis
2.4.2. Volatile Organic Compound Analysis
2.4.3. Odor Activity Value
2.5. Mathematical Modeling
2.5.1. Microorganism Growth
2.5.2. Limiting Substrate
2.5.3. Metabolite Production Modeling
2.5.4. Model Fitting and Statistical Analyses
3. Results and Discussion
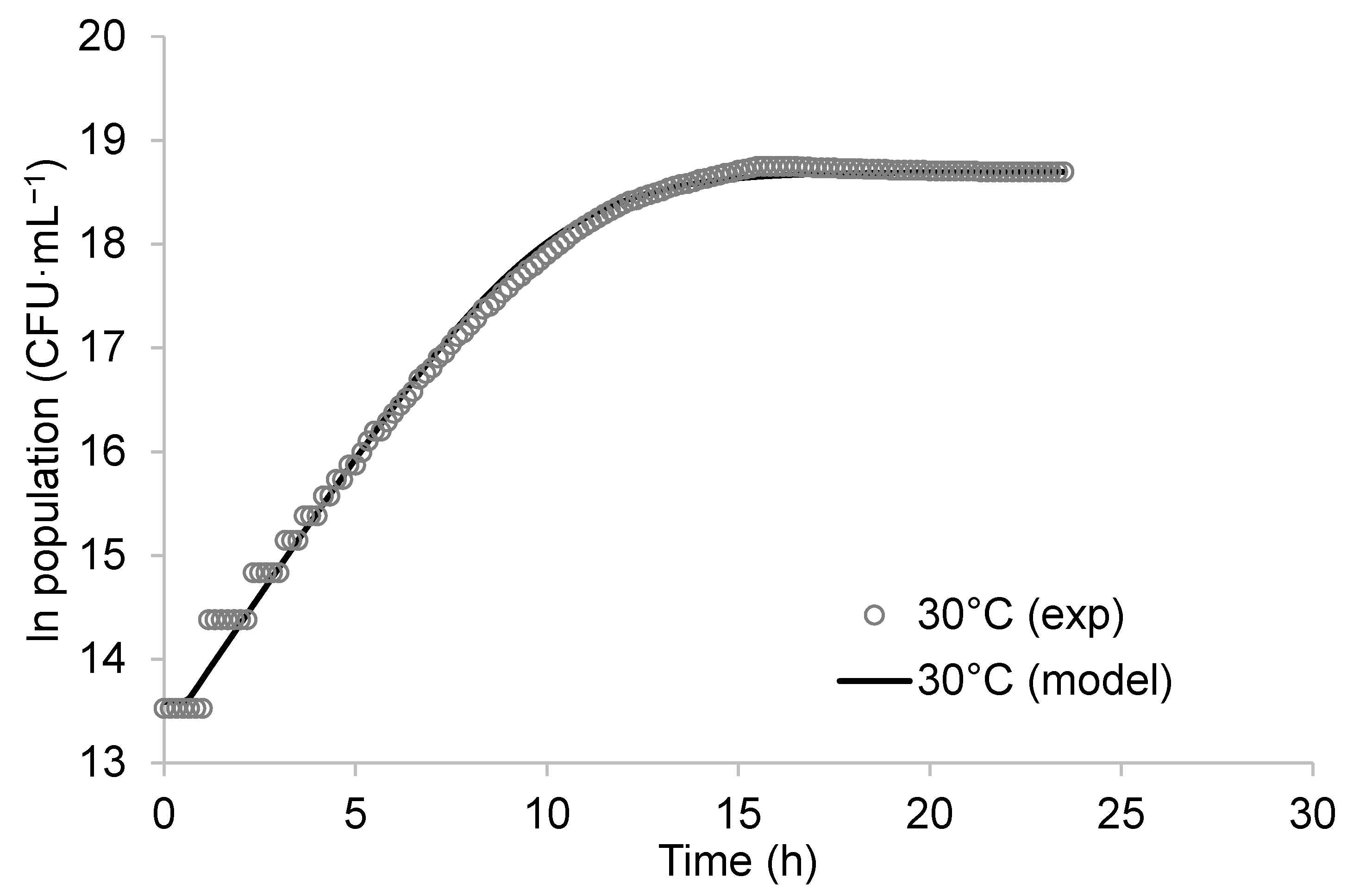
3.1. Growth Characteristic of K. unispora
3.1.1. Microorganism Growth Modeling
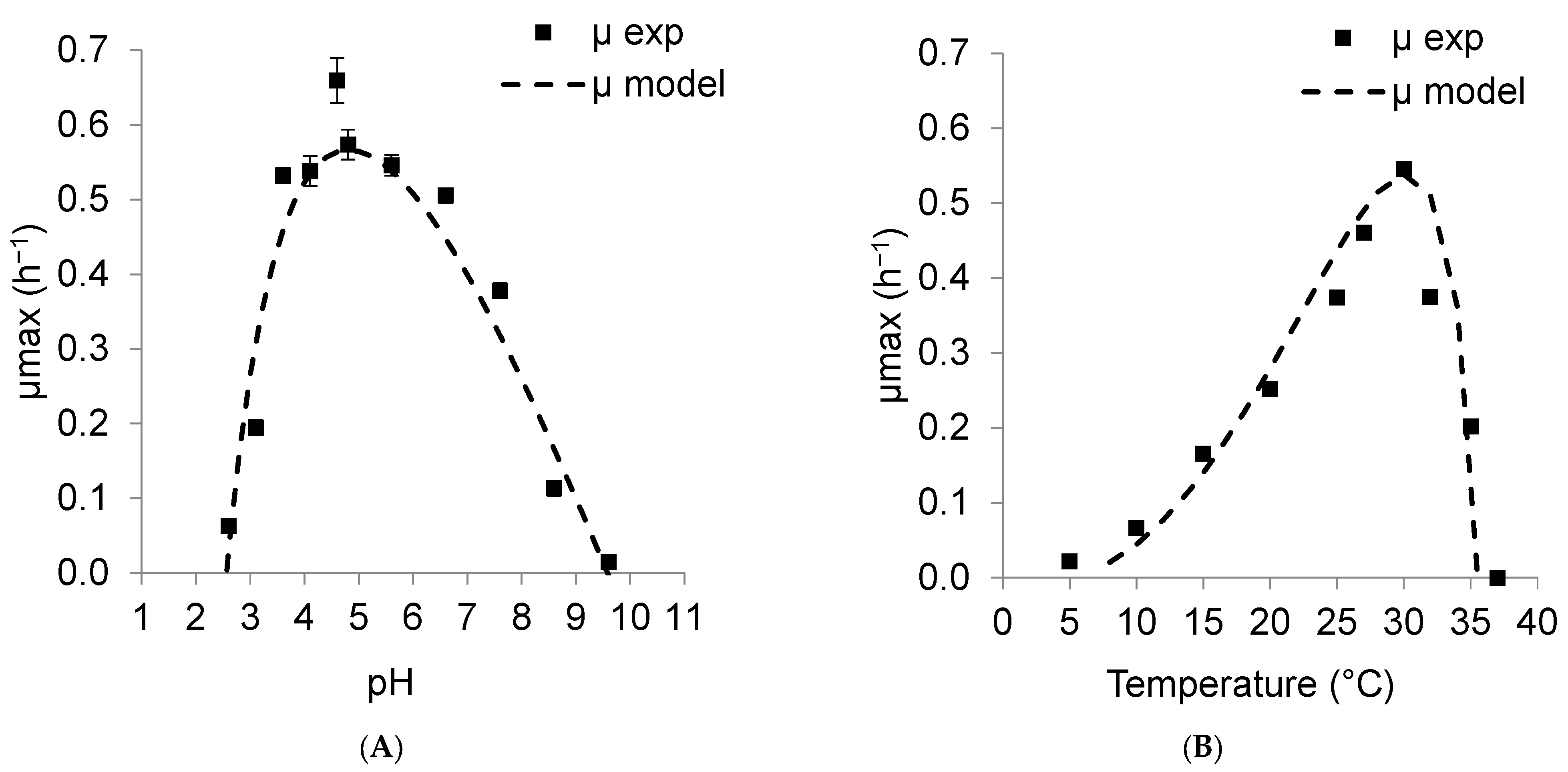
3.1.2. Modeling Effect of pH and Temperature for K. unispora
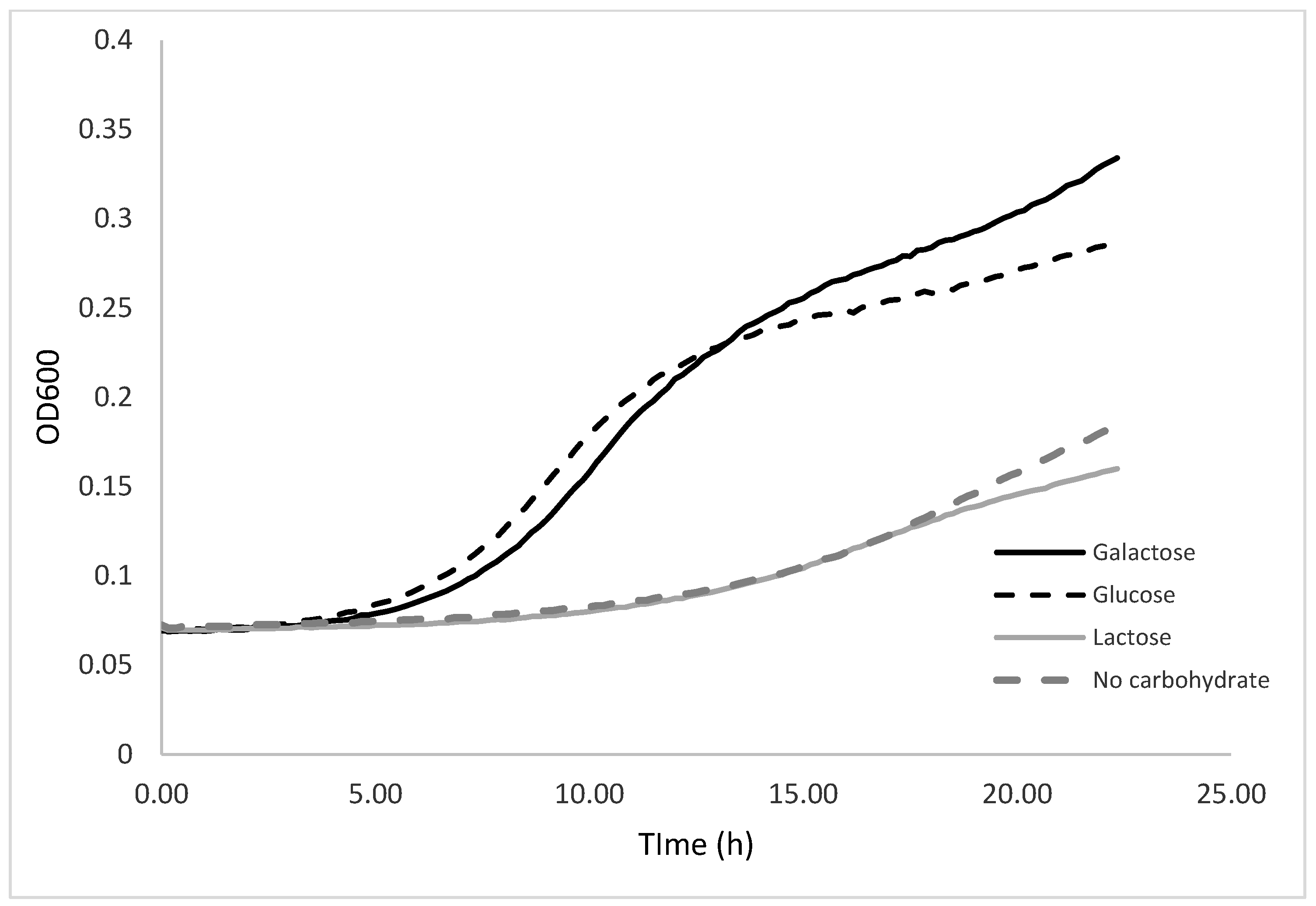
3.1.3. Effect of the Carbohydrate Source on K. unispora Growth
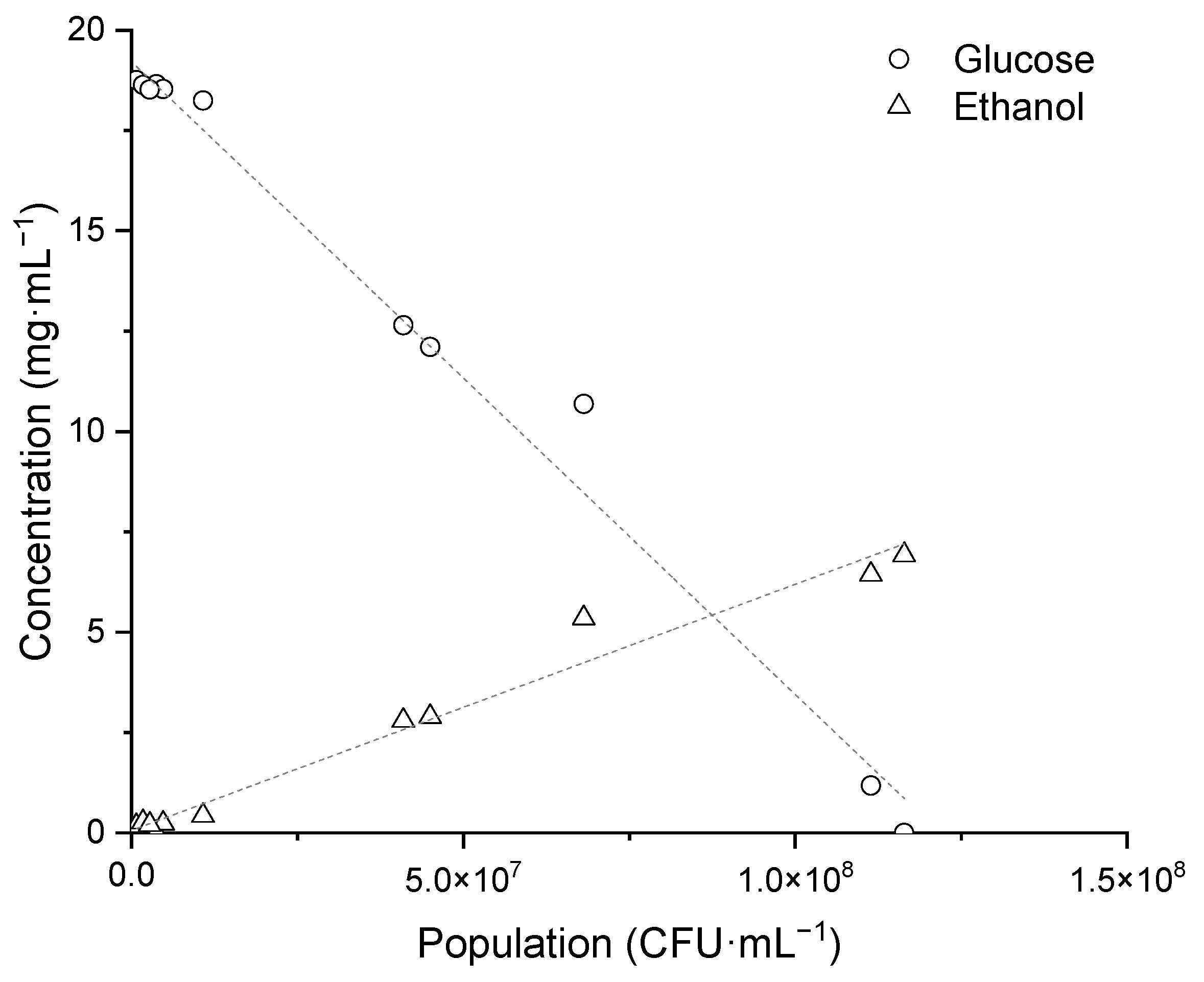
3.2. Ethanol Production from K. unispora Growth
3.3. Resulting Parameters of Models
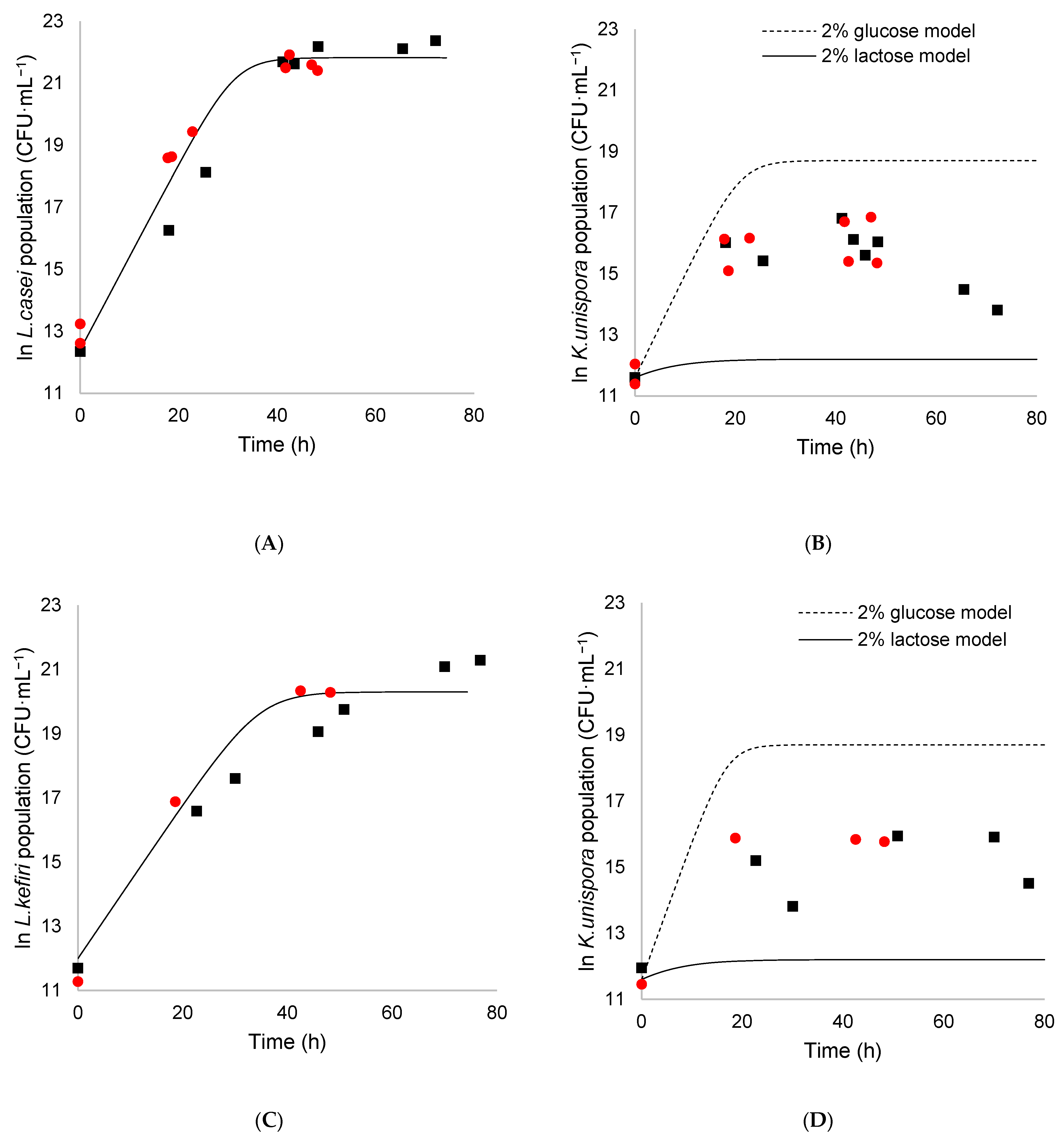
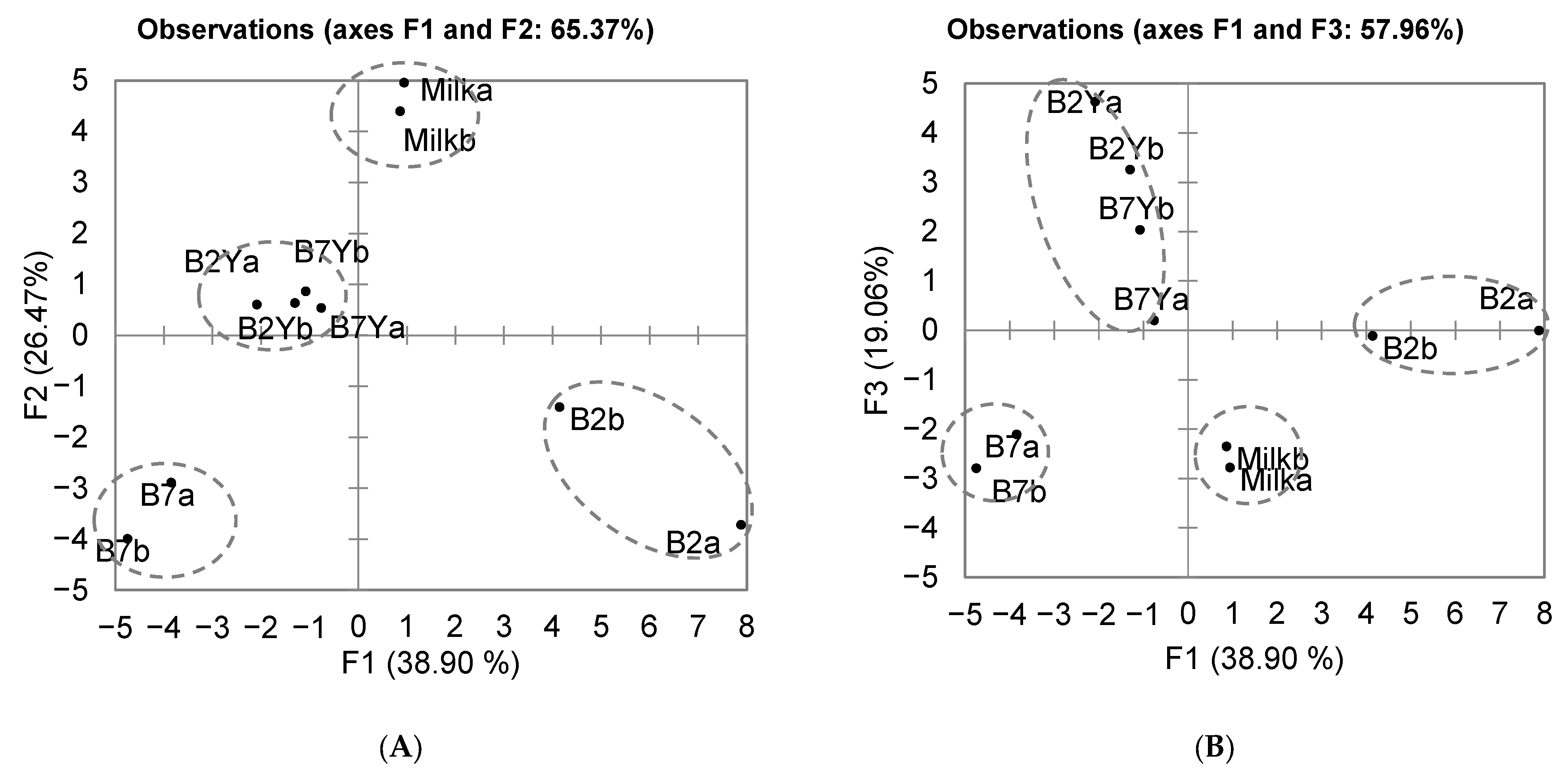
3.4. Coculture of K. unispora with Lactic Acid Bacteria
3.4.1. Evolution of the Population
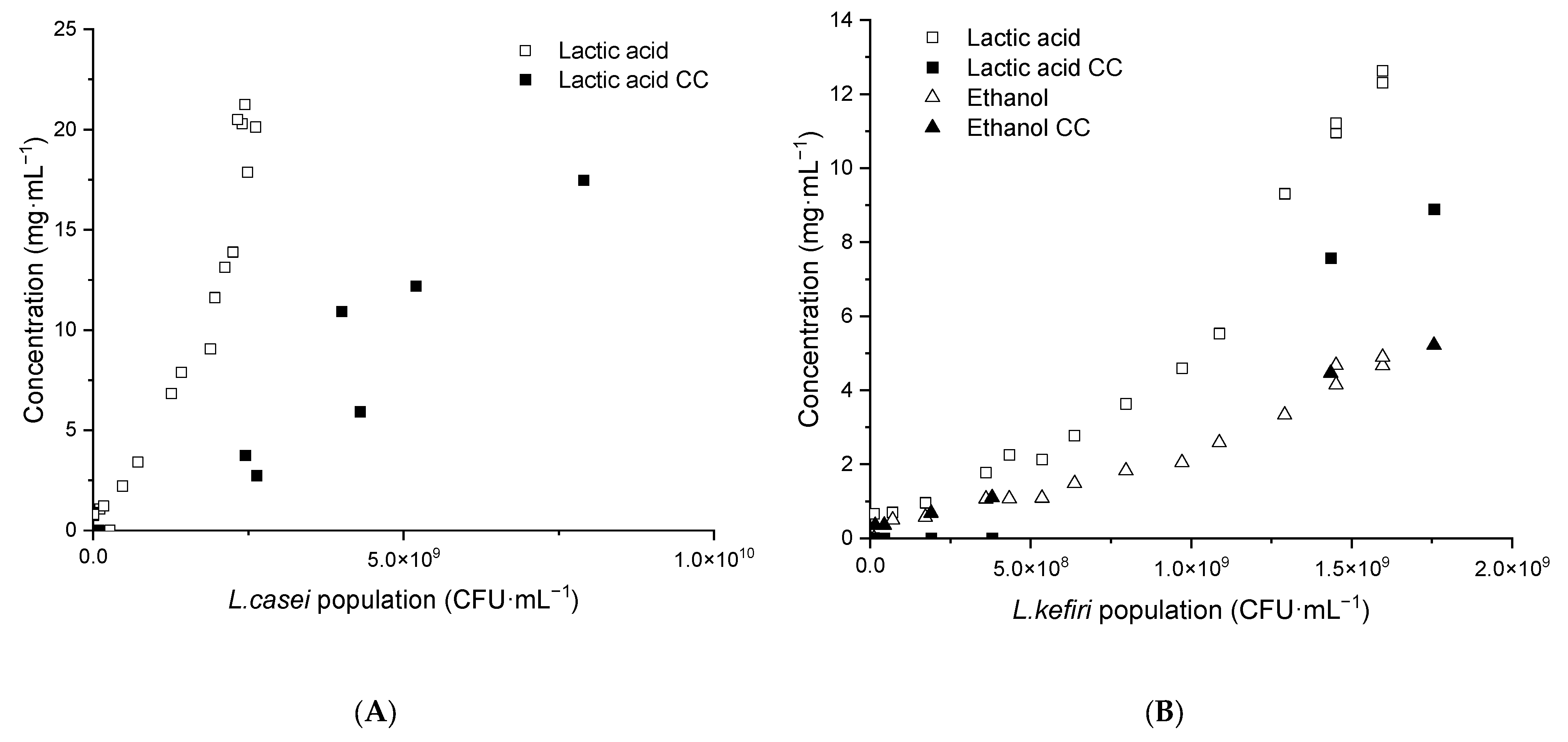
3.4.2. Lactic Acid and Ethanol Production
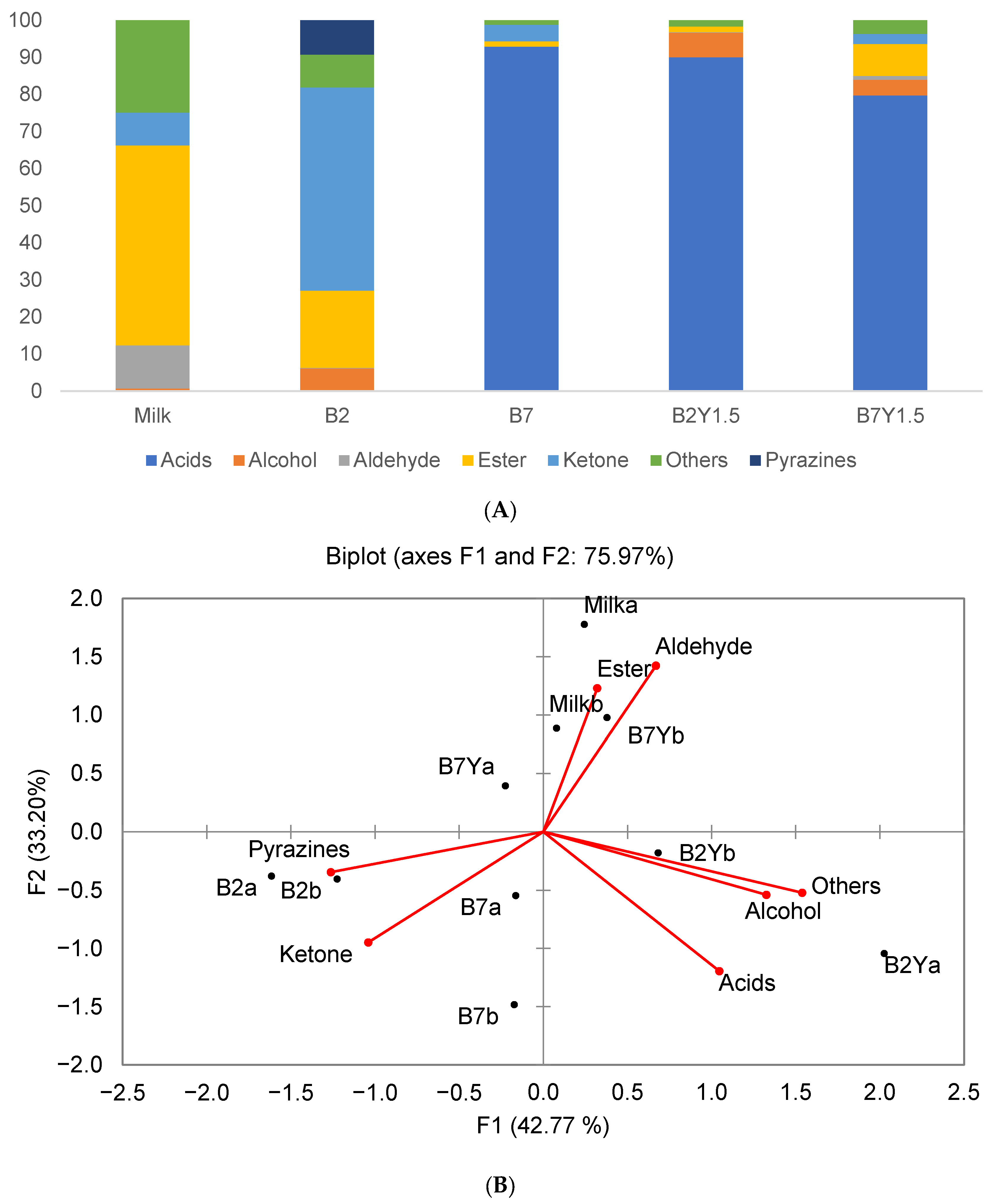
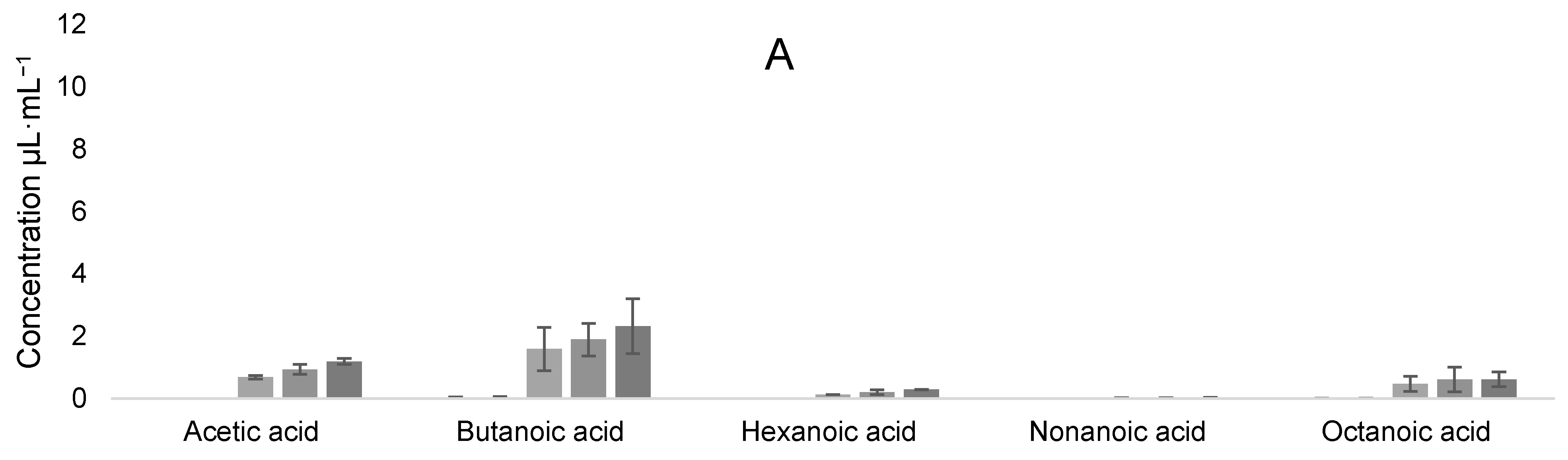
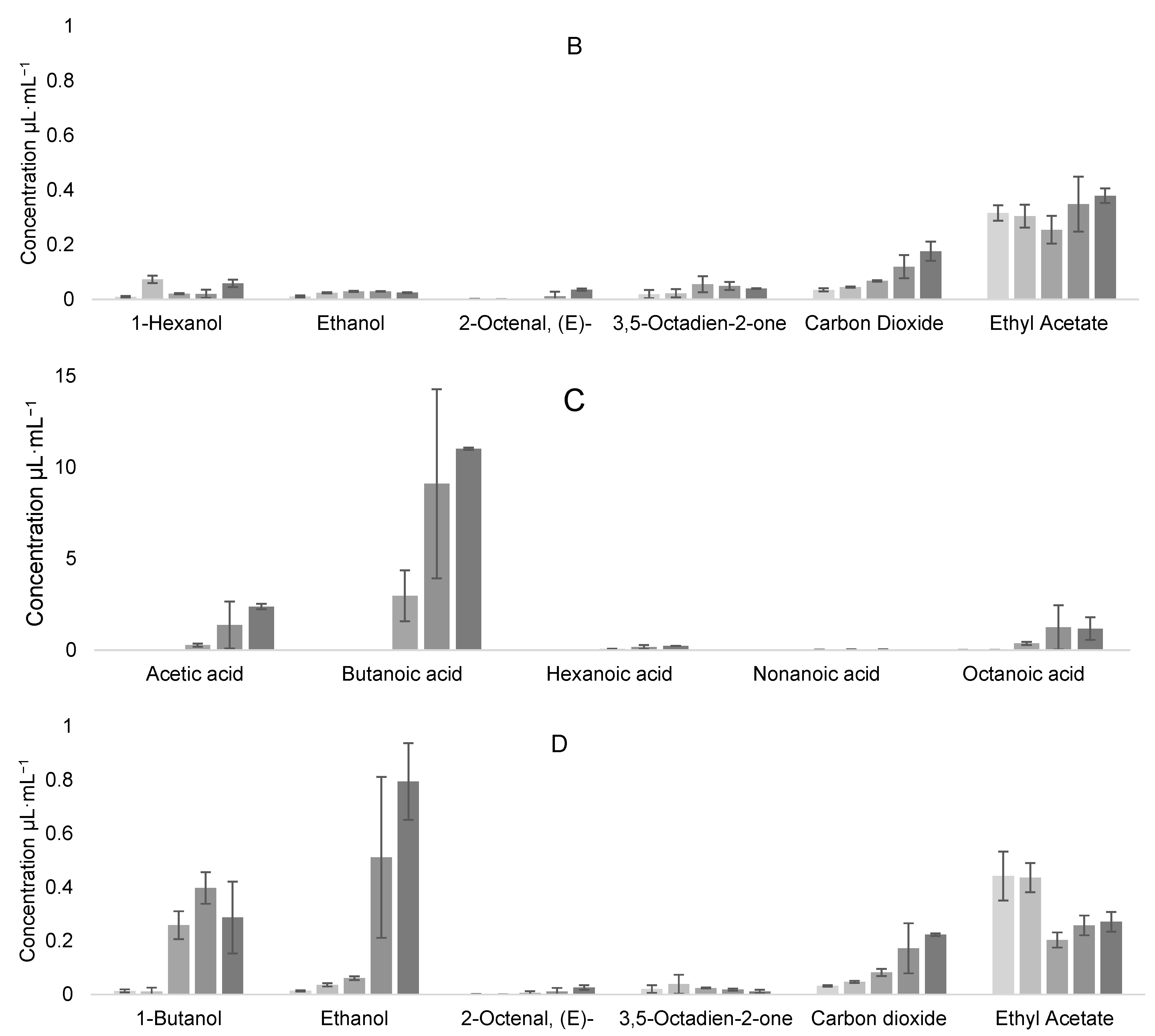
3.5. Volatile Organic Compound Production
Evolution of VOCs during Coculture Fermentation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tang, H.; Ma, H.; Hou, Q.; Li, W.; Xu, H.; Liu, W.; Sun, Z.; Haobisi, H.; Menghe, B. Profiling of koumiss microbiota and organic acids and their effects on koumiss taste. BMC Microbiol. 2020, 20, 85. [Google Scholar] [CrossRef] [PubMed]
- ST RK 1004-98. Natural Koumiss. Committee for Standardization, Metrology and Certification of the Republic of Kazakhstan: Astana City, Kazakhstan, 1999.
- Kondybayev, A.; Loiseau, G.; Achir, N.; Mestres, C.; Konuspayeva, G. Fermented mare milk product (Qymyz, Koumiss). Int. Dairy J. 2021, 119, 105065. [Google Scholar] [CrossRef]
- McKellar, R.C.; Lu, X. Modeling Microbial Responses in Food; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Rosso, L.; Lobry, J.R.; Bajard, S.; Flandrois, J.P. Convenient Model To Describe the Combined Effects of Temperature and pH on Microbial Growth. Appl. Environ. Microbiol. 1995, 61, 610–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munanga, B.D.J.C.; Loiseau, G.; Grabulos, J.; Mestres, C. Modeling Lactic Fermentation of Gowé Using Lactobacillus Starter Culture. Microorganisms 2016, 4, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornu, M.; Billoir, E.; Bergis, H.; Beaufort, A.; Zuliani, V. Modeling microbial competition in food: Application to the behavior of Listeria monocytogenes and lactic acid flora in pork meat products. Food Microbiol. 2011, 28, 639–647. [Google Scholar] [CrossRef]
- Montanari, G.; Zambonelli, C.; Grazia, L.; Kamesheva, G.K.; Shigaeva, M.K. Saccharomyces unisporus as the principal alcoholic fermentation microorganism of traditional koumiss. J. Dairy Res. 1996, 63, 327–331. [Google Scholar] [CrossRef]
- Wood, B.J.; Holzapfel, W. The Genera of Lactic Acid Bacteria; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1992; Volume 2. [Google Scholar]
- Hickey, M.W.; Hillier, A.J.; Jago, G.R. Transport and Metabolism of Lactose, Glucose, and Galactose in Homofermentative Lactobacilli. Appl. Environ. Microbiol. 1986, 51, 825–831. [Google Scholar] [CrossRef] [Green Version]
- Hutkins, R.W.; Ponne, C. Lactose uptake driven by galactose efflux in Streptococcus thermophilus: Evidence for a galactose-lactose antiporter. Appl. Environ. Microbiol. 1991, 57, 941–944. [Google Scholar] [CrossRef] [Green Version]
- Gadaga, T.H.; Mutukumira, A.N.; Narvhus, J.A. The growth and interaction of yeasts and lactic acid bacteria isolated from Zimbabwean naturally fermented milk in UHT milk. Int. J. Food Microbiol. 2001, 68, 21–32. [Google Scholar] [CrossRef]
- Mu, Z.; Yang, X.; Yuan, H. Detection and identification of wild yeast in Koumiss. Food Microbiol. 2012, 31, 301–308. [Google Scholar] [CrossRef]
- Meng, Y.; Chen, X.; Sun, Z.; Li, Y.; Chen, D.; Fang, S.; Chen, J. Exploring core microbiota responsible for the production of volatile flavor compounds during the traditional fermentation of Koumiss. LWT 2021, 135, 110049. [Google Scholar] [CrossRef]
- Kondybayev, A.; Konuspayeva, G.; Strub, C.; Loiseau, G.; Mestres, C.; Grabulos, J.; Manzano, M.; Akhmetsadykova, S.; Achir, N. Growth and Metabolism of Lacticaseibacillus casei and Lactobacillus kefiri Isolated from Qymyz, a Traditional Fermented Central Asian Beverage. Fermentation 2022, 8, 367. [Google Scholar] [CrossRef]
- Stiles, J.; Penkar, S.; Plocková, M.; Chumchalova, J.; Bullerman, L. Antifungal activity of sodium acetate and Lactobacillus rhamnosus. J. Food Prot. 2002, 65, 1188–1191. [Google Scholar] [CrossRef]
- Augustin, J.-C.; Rosso, L.; Carlier, V. Estimation of temperature dependent growth rate and lag time of Listeria monocytogenes by optical density measurements. J. Microbiol. Methods 1999, 38, 137–146. [Google Scholar] [CrossRef]
- Dan, T.; Chen, H.; Li, T.; Tian, J.; Ren, W.; Zhang, H.; Sun, T. Influence of Lactobacillus plantarum P-8 on fermented milk flavor and storage stability. Front. Microbiol. 2019, 9, 3133. [Google Scholar] [CrossRef]
- Delhalle, L.; Daube, G.; Adolphe, Y.; Crevecoeur, S.; Clinquart, A. Les modèles de croissance en microbiologie prévisionnelle pour la maitrise de la sécurité des aliments (synthèse bibliographique). Biotechnol. Agron. Soc. Environ. 2012, 16, 369–381. [Google Scholar]
- Zwietering, M.H.; Wijtzes, T.; Rombouts, F.M.; Riet, K. A decision support system for prediction of microbial spoilage in foods. J. Ind. Microbiol. 1993, 12, 324–329. [Google Scholar] [CrossRef]
- Rodrigues, C.I.S.; Wahl, A.; Gombert, A.K. Aerobic growth physiology of Saccharomyces cerevisiae on sucrose is strain-dependent. FEMS Yeast Res. 2021, 21, foab021. [Google Scholar] [CrossRef]
- Korcari, D.; Ricci, G.; Capusoni, C.; Fortina, M.G. Physiological performance of Kazachstania unispora in sourdough environments. World J. Microbiol. Biotechnol. 2021, 37, 88. [Google Scholar] [CrossRef]
- Bruder, S.; Reifenrath, M.; Thomik, T.; Boles, E.; Herzog, K. Parallelised online biomass monitoring in shake flasks enables efficient strain and carbon source dependent growth characterisation of Saccharomyces cerevisiae. Microb. Cell Factories 2016, 15, 127. [Google Scholar] [CrossRef] [Green Version]
- Canonico, L.; Zannini, E.; Ciani, M.; Comitini, F. Assessment of non-conventional yeasts with potential probiotic for protein-fortified craft beer production. LWT 2021, 145, 111361. [Google Scholar] [CrossRef]
- Korcari, D.; Secchiero, R.; Laureati, M.; Marti, A.; Cardone, G.; Rabitti, N.S.; Ricci, G.; Fortina, M.G. Technological properties, shelf life and consumer preference of spelt-based sourdough bread using novel, selected starter cultures. LWT 2021, 151, 112097. [Google Scholar] [CrossRef]
- Shigaeva, M.K.; Ospanova, M.S. Microflora of National Fermented Milk Drinks; Nauka: Alma-Ata, Kazakhstan, 1983. [Google Scholar]
- Dan, T.; Wang, D.; Wu, S.; Jin, R.; Ren, W.; Sun, T. Profiles of Volatile Flavor Compounds in Milk Fermented with Different Proportional Combinations of Lactobacillus delbrueckii subsp. bulgaricus and Streptococcus thermophilus. Molecules 2017, 22, 1633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kourkoutas, Y.; Bosnea, L.; Taboukos, S.; Baras, C.; Lambrou, D.; Kanellaki, M. Probiotic Cheese Production Using Lactobacillus casei Cells Immobilized on Fruit Pieces. J. Dairy Sci. 2006, 89, 1439–1451. [Google Scholar] [CrossRef]
- Tabit, F.T. Prevalence and growth characteristics of Bacillus sporothermodurans in UHT milk. Br. Food J. 2018, 120, 2250–2260. [Google Scholar] [CrossRef]
- Dickschat, J.S.; Wickel, S.; Bolten, C.J.; Nawrath, T.; Schulz, S.; Wittmann, C. Pyrazine Biosynthesis in Corynebacterium glutamicum. Eur. J. Org. Chem. 2010, 2010, 2687–2695. [Google Scholar] [CrossRef]
- Boonbumrung, S.; Tamura, H.; Mookdasanit, J.; Nakamoto, H.; Ishihara, M.; Yoshizawa, T.; Varanyanond, W. Characteristic aroma components of the volatile oil of yellow keaw mango fruits determined by limited odor unit method. Food Sci. Technol. Res. 2001, 7, 200–206. [Google Scholar] [CrossRef] [Green Version]
- Qian, M.; Reineccius, G. Quantification of aroma compounds in Parmigiano Reggiano cheese by a dynamic headspace gas chromatography-mass spectrometry technique and calculation of odor activity value. J. Dairy Sci. 2003, 86, 770–776. [Google Scholar] [CrossRef] [Green Version]
- Buttery, R.G.; Turnbaugh, J.G.; Ling, L.C. Contribution of volatiles to rice aroma. J. Agric. Food Chem. 1988, 36, 1006–1009. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
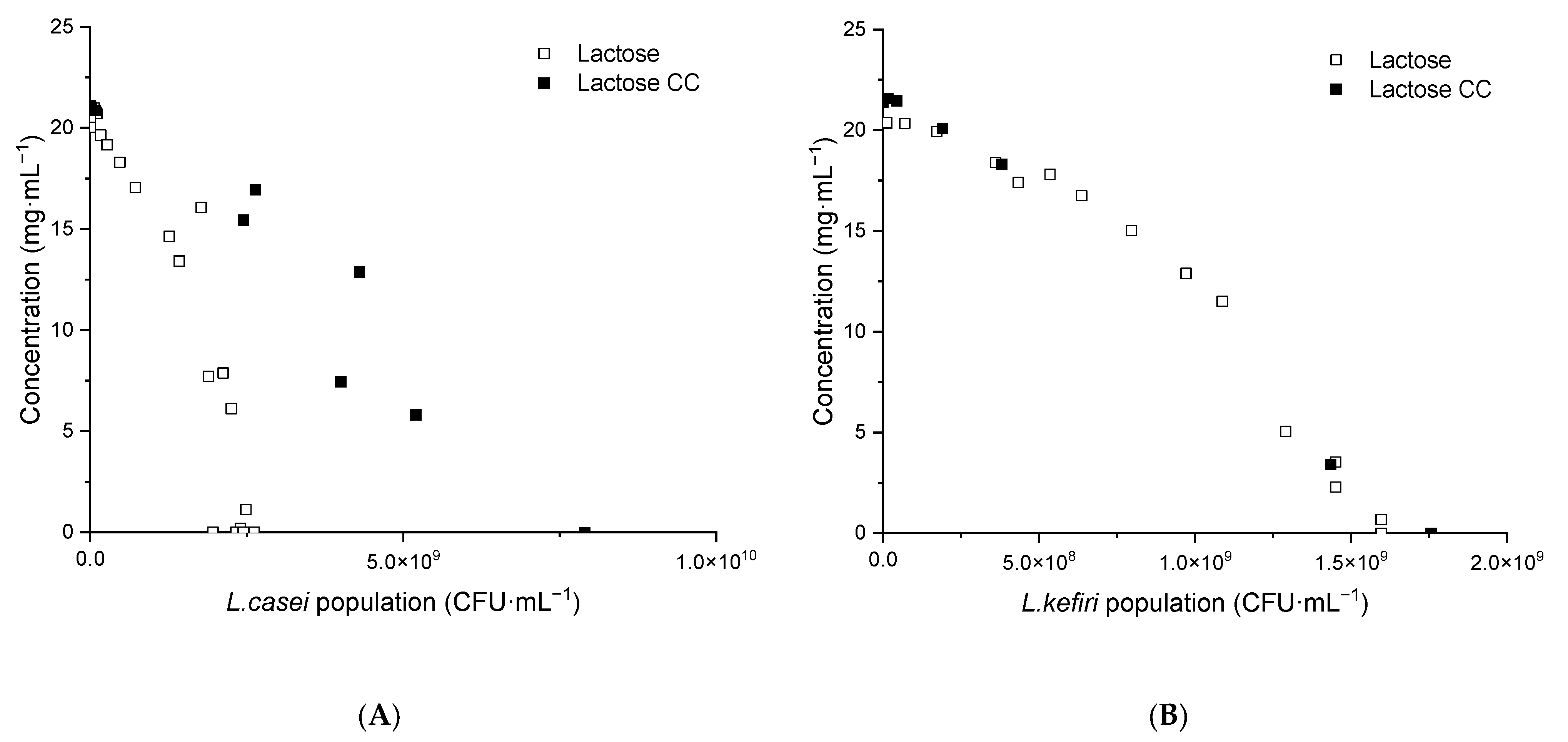{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Kazachstania unispora (Value ± SD) |
|---|---|
| pHmin | 2.56 ± 0.09 |
| pHmax | 9.58 ± 030 |
| pHopt | 4.81 ± 0.22 |
| Tmin (°C) | 3.81 ± 3.33 |
| Tmax (°C) | 35.70 ± 0.20 |
| Topt (°C) | 30.16 ± 0.53 |
| µopt (h−1) | 0.57 ± 0.02 |
| Glucose µopt (h−1) | 0.45 ± 0.01 |
| Glucose Ks (g·L−1) | 0.24 ± 0.03 |
| Galactose µopt (h−1) | 0.45 ± 0.01 |
| Galactose Ks (g·L−1) | 0.13 ± 0.04 |
| Ethanol Production (10−8 mg·CFU−1) | 6.13 ± 0.27 |
| Compound | OT ppm | L. kefiri–K. unispora | L. casei–K. unispora | References | ||
|---|---|---|---|---|---|---|
| OAV 0 h | OAV 138 h | OAV 0 h | OAV 138 h | |||
| 1-Butanol | 4.3 | 0.0032 | 0.0668 | 0.0022 | 0.0138 | [31] |
| Ethanol | 4.5 | 0.0031 | 0.1766 | 0.0025 | 0.0056 | [31] |
| 2-Octenal, (E)- | 0.003 | 0.6559 | 8.7322 | 0.7572 | 12.0579 | [18] |
| 3,5-Octadien-2-one | - | |||||
| Ethyl Acetate | 3.2 | 0.1382 | 0.0848 | 0.0990 | 0.1186 | [31] |
| Acetic acid | 32 | 0.0000 | 0.0739 | 0.0000 | 0.0367 | [32] |
| Butanoic acid | 2.7 | 0.0017 | 4.0827 | 0.0137 | 0.8597 | [32] |
| Hexanoic acid | 27 | 0.0000 | 0.0087 | 0.0000 | 0.0104 | [32] |
| Nonanoic acid | 3 | 0.0014 | 0.0136 | 0.0006 | 0.0104 | [33] |
| Octanoic acid | 11 | 0.0004 | 0.1048 | 0.0006 | 0.0547 | [32] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondybayev, A.; Achir, N.; Mestres, C.; Collombel, I.; Strub, C.; Grabulos, J.; Akhmetsadykov, N.; Aubakirova, A.; Kamidinkyzy, U.; Ghanmi, W.; et al. Growth Kinetics of Kazachstania unispora and Its Interaction with Lactic Acid Bacteria during Qymyz Production. Fermentation 2023, 9, 101. https://doi.org/10.3390/fermentation9020101
Kondybayev A, Achir N, Mestres C, Collombel I, Strub C, Grabulos J, Akhmetsadykov N, Aubakirova A, Kamidinkyzy U, Ghanmi W, et al. Growth Kinetics of Kazachstania unispora and Its Interaction with Lactic Acid Bacteria during Qymyz Production. Fermentation. 2023; 9(2):101. https://doi.org/10.3390/fermentation9020101
Chicago/Turabian StyleKondybayev, Askar, Nawel Achir, Christian Mestres, Ingrid Collombel, Caroline Strub, Joel Grabulos, Nurlan Akhmetsadykov, Aidana Aubakirova, Ulzhan Kamidinkyzy, Wijden Ghanmi, and et al. 2023. "Growth Kinetics of Kazachstania unispora and Its Interaction with Lactic Acid Bacteria during Qymyz Production" Fermentation 9, no. 2: 101. https://doi.org/10.3390/fermentation9020101