
Metabolic Engineering of Saccharomyces cerevisiae for Production of Fragrant Terpenoids from Agarwood and Sandalwood
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strain, Media, and Transformation
2.2. RNA Isolation and Transcript Quantification
2.3. Quantification of Terpenoid Production in Engineered Strains
3. Results and Discussion
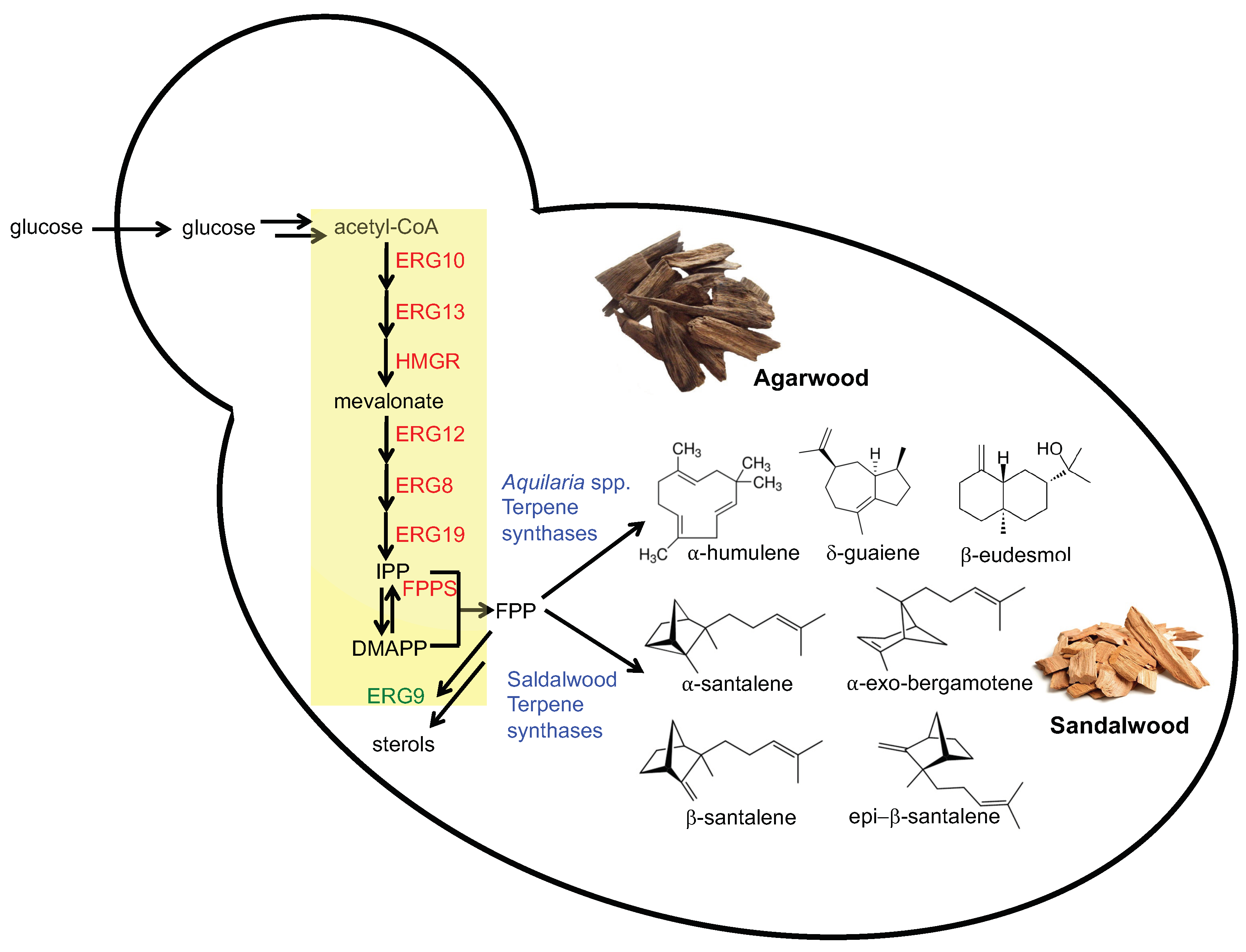
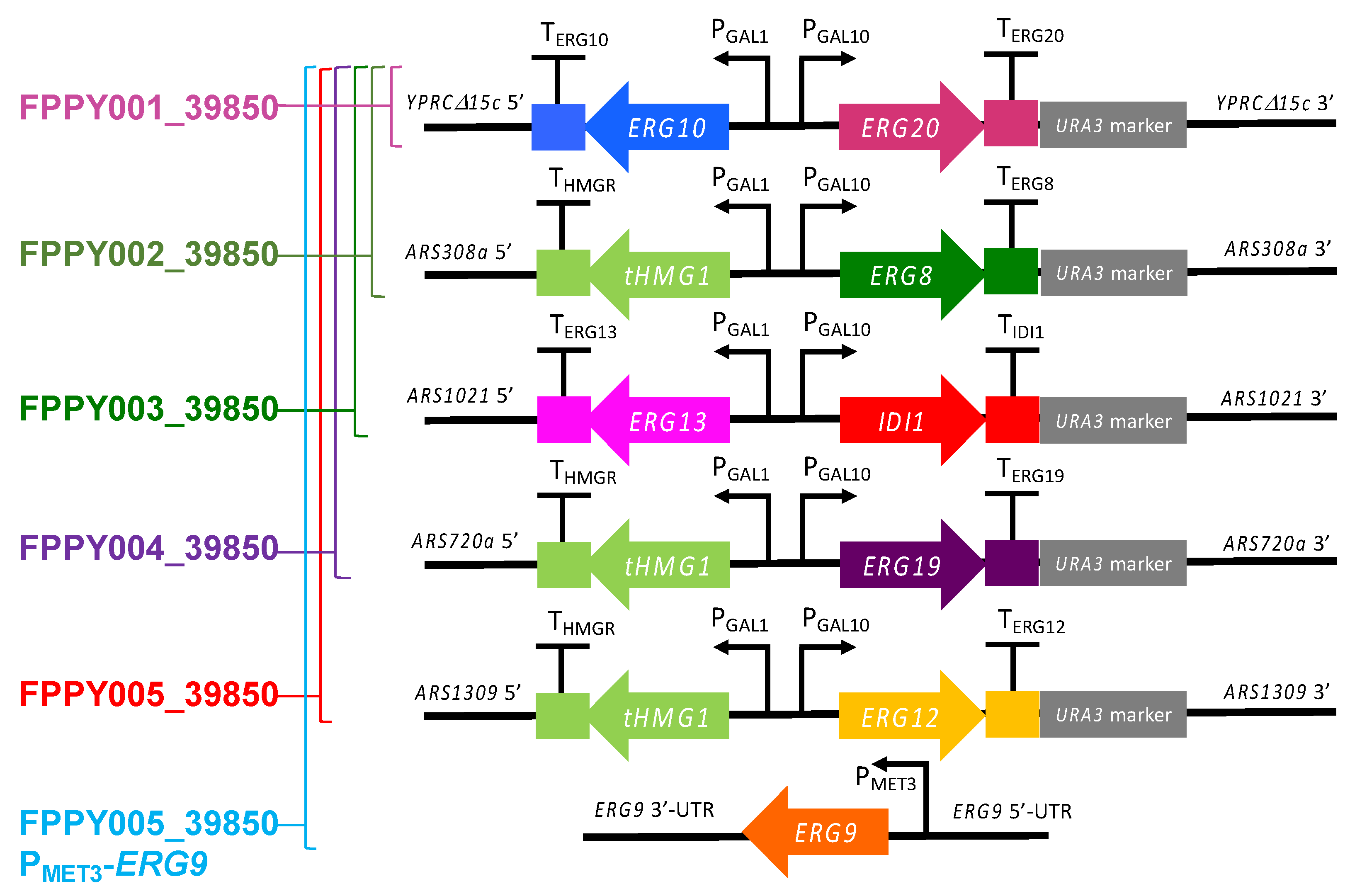
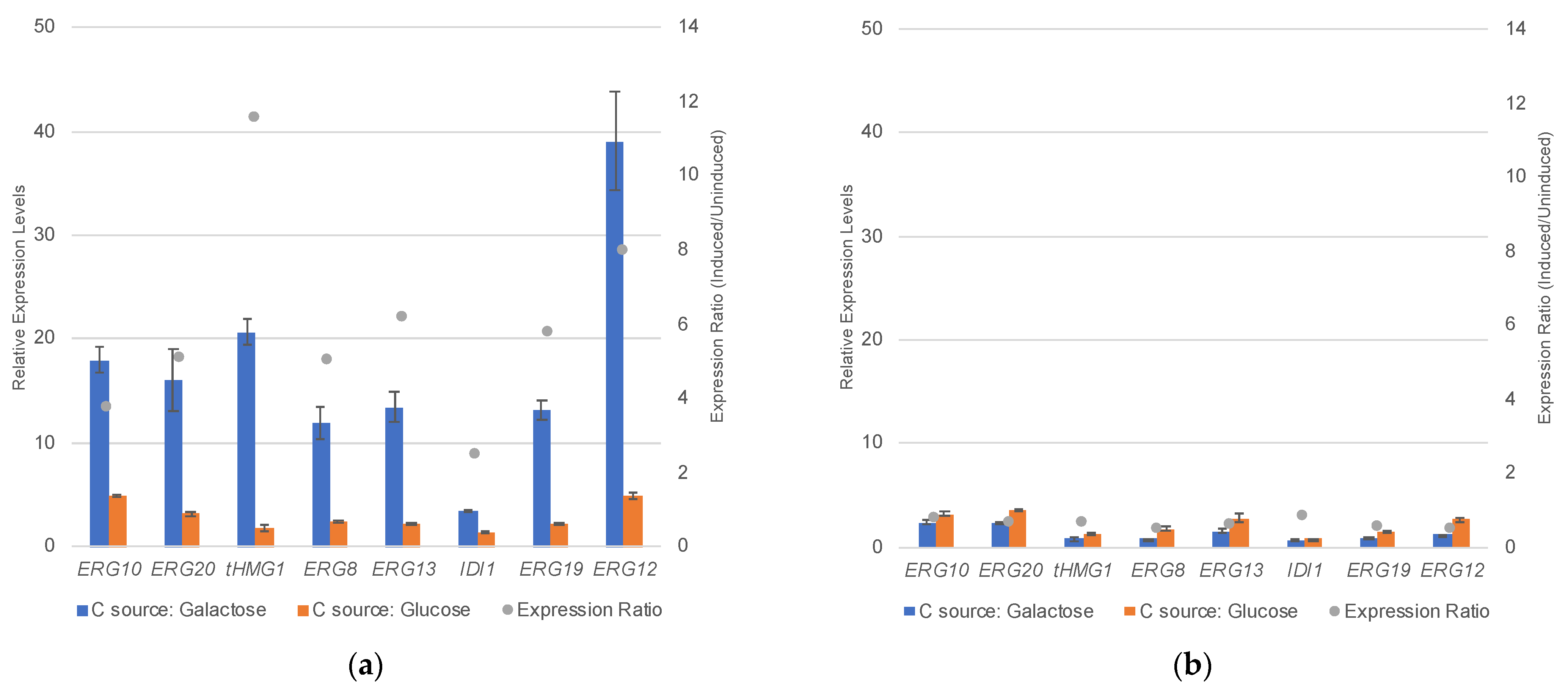
3.1. Overexpression of the Entire Mevalonate Pathway Genes in S. cerevisiae
3.2. Screening of Terpene Synthases in FPPY005_39850
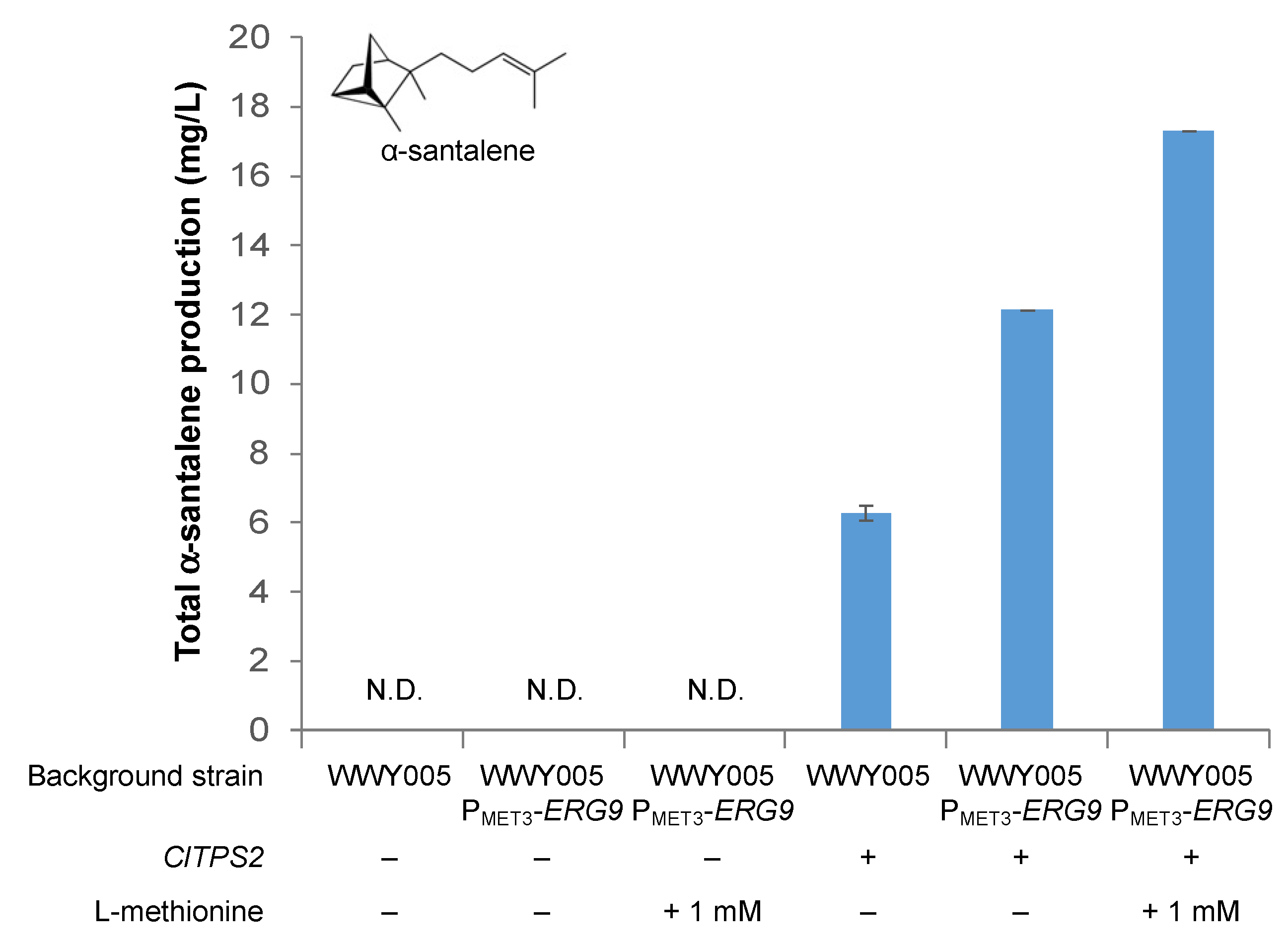
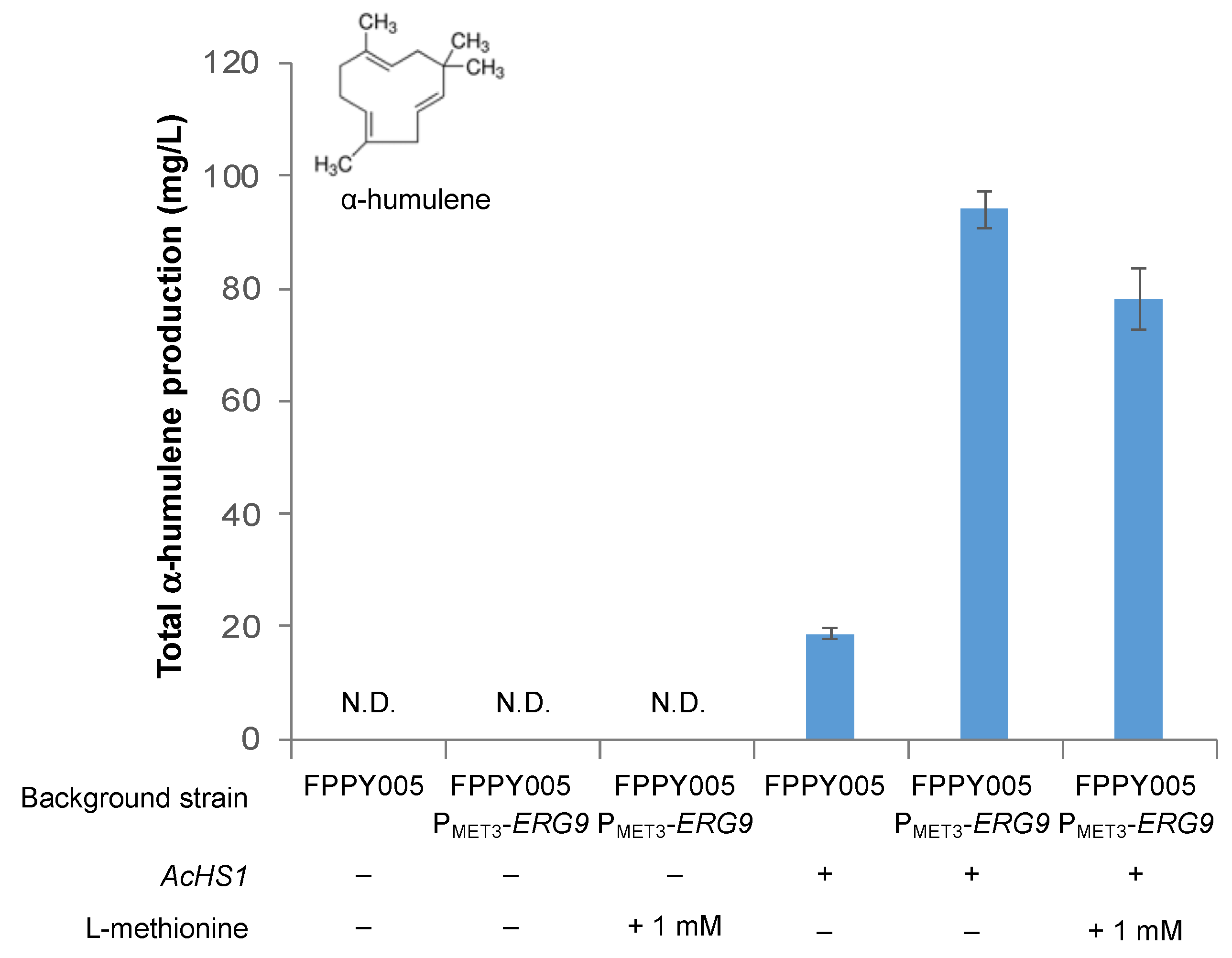
3.3. Improving Terpenoid Production via Downregulation of ERG9 Gene Expression
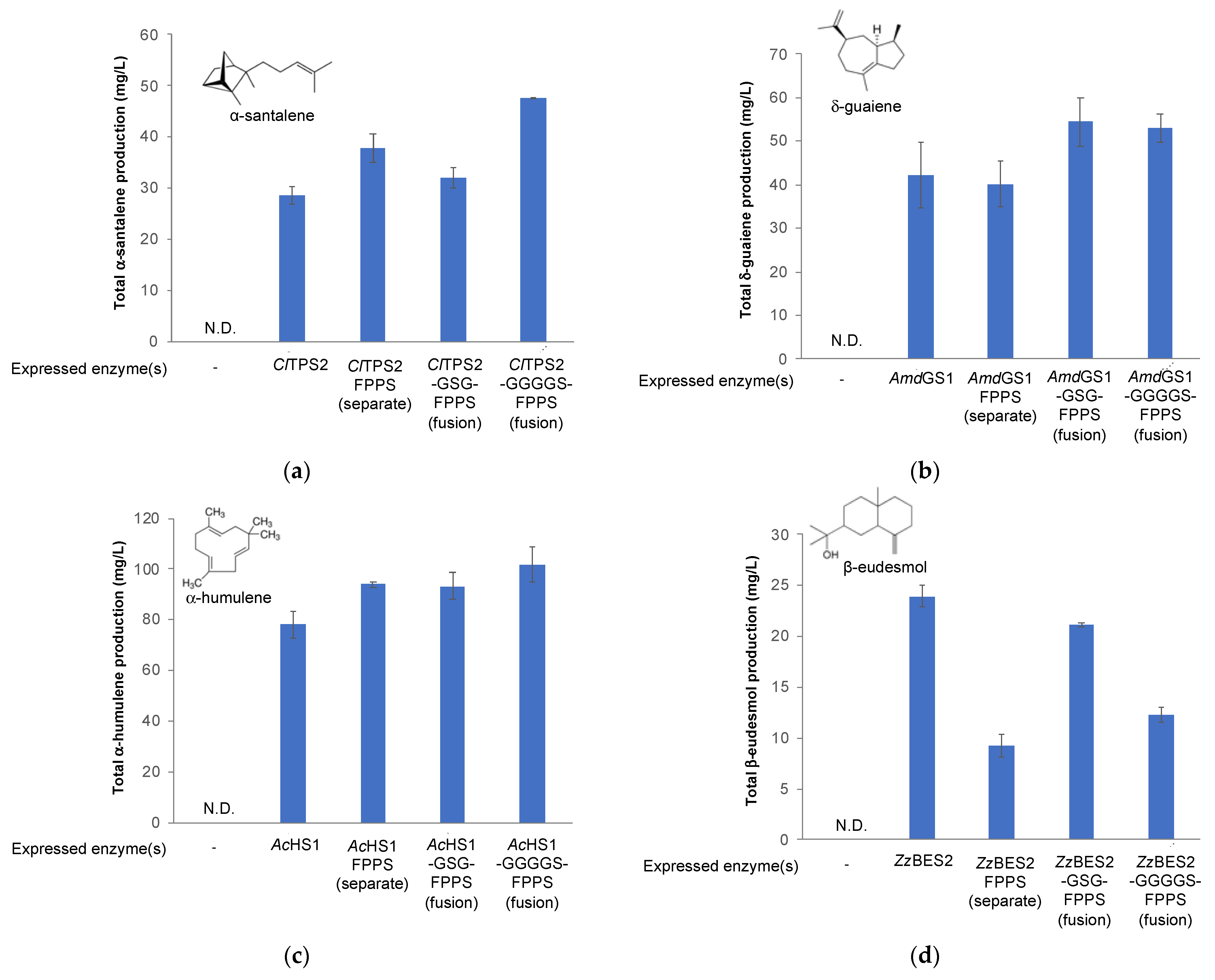
3.4. Improving Fragrant Terpenoid Production via Enzyme Fusion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, C.; Hong, K. Production of Terpenoids by Synthetic Biology Approaches. Front. Bioeng. Biotechnol. 2020, 8, 347. [Google Scholar] [CrossRef] [PubMed]
- Kung, Y.; Runguphan, W.; Keasling, J.D. From Fields to Fuels: Recent Advances in the Microbial Production of Biofuels. ACS Synth. Biol. 2012, 1, 498–513. [Google Scholar] [CrossRef] [PubMed]
- Belcher, M.S.; Mahinthakumar, J.; Keasling, J.D. New Frontiers: Harnessing Pivotal Advances in Microbial Engineering for the Biosynthesis of Plant-Derived Terpenoids. Curr. Opin. Biotechnol. 2020, 65, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.Y.; Keasling, J.D. Production of Isoprenoid Pharmaceuticals by Engineered Microbes. Nat. Chem. Biol. 2006, 2, 674–681. [Google Scholar] [CrossRef]
- Navale, G.R.; Dharne, M.S.; Shinde, S.S. Metabolic Engineering and Synthetic Biology for Isoprenoid Production in Escherichia Coli and Saccharomyces Cerevisiae. Appl. Microbiol. Biotechnol. 2021, 105, 457–475. [Google Scholar] [CrossRef]
- US20080083158A1-Fuel Compositions Comprising Farnesane and Farnesane Derivatives and Method of Making and Using Same-Google Patents. Available online: https://patents.google.com/patent/US20080083158A1/en (accessed on 5 August 2022).
- Westfall, P.J.; Pitera, D.J.; Lenihan, J.R.; Eng, D.; Woolard, F.X.; Regentin, R.; Horning, T.; Tsuruta, H.; Melis, D.J.; Owens, A.; et al. Production of Amorphadiene in Yeast, and Its Conversion to Dihydroartemisinic Acid, Precursor to the Antimalarial Agent Artemisinin. Proc. Natl. Acad. Sci. USA 2012, 109, E111–E118. [Google Scholar] [CrossRef]
- Paddon, C.J.; Keasling, J.D. Semi-Synthetic Artemisinin: A Model for the Use of Synthetic Biology in Pharmaceutical Development. Nat. Rev. Microbiol. 2014, 12, 355–367. [Google Scholar] [CrossRef]
- Anderson, M.S.; Yarger, J.G.; Burck, C.L.; Poulter, C.D. Farnesyl Diphosphate Synthetase. Molecular Cloning, Sequence, and Expression of an Essential Gene from Saccharomyces Cerevisiae. J. Biol. Chem. 1989, 264, 19176–19184. [Google Scholar] [CrossRef]
- Zhang, X.; Niu, M.; Teixeira Da Silva, J.A.; Zhang, Y.; Yuan, Y.; Jia, Y.; Xiao, Y.; Li, Y.; Fang, L.; Zeng, S.; et al. Identification and Functional Characterization of Three New Terpene Synthase Genes Involved in Chemical Defense and Abiotic Stresses in Santalum Album. BMC Plant. Biol. 2019, 19, 115. [Google Scholar] [CrossRef]
- Jones, C.G.; Moniodis, J.; Zulak, K.G.; Scaffidi, A.; Plummer, J.A.; Ghisalberti, E.L.; Barbour, E.L.; Bohlmann, J. Sandalwood Fragrance Biosynthesis Involves Sesquiterpene Synthases of Both the Terpene Synthase (TPS)-a and TPS-b Subfamilies, Including Santalene Synthases. J. Biol. Chem. 2011, 286, 17445–17454. [Google Scholar] [CrossRef] [Green Version]
- Hashim, Y.Z.H.Y.; Kerr, P.G.; Abbas, P.; Mohd Salleh, H. Aquilaria Spp. (Agarwood) as Source of Health Beneficial Compounds: A Review of Traditional Use, Phytochemistry and Pharmacology. J. Ethnopharmacol. 2016, 189, 331–360. [Google Scholar] [CrossRef] [PubMed]
- Samadi, M.; Zainal Abidin, Z.; Yoshida, H.; Yunus, R.; Awang Biak, D.R. Towards Higher Oil Yield and Quality of Essential Oil Extracted from Aquilaria Malaccensis Wood via the Subcritical Technique. Molecules 2020, 25, 3872. [Google Scholar] [CrossRef] [PubMed]
- Kumeta, Y.; Ito, M. Characterization of Delta-Guaiene Synthases from Cultured Cells of Aquilaria, Responsible for the Formation of the Sesquiterpenes in Agarwood. Plant. Physiol. 2010, 154, 1998–2007. [Google Scholar] [CrossRef]
- Watcharawipas, A.; Sansatchanon, K.; Phithakrotchanakoon, C.; Tanapongpipat, S.; Runguphan, W.; Kocharin, K. Novel Carotenogenic Gene Combinations from Red Yeasts Enhanced Lycopene and Beta-Carotene Production in Saccharomyces Cerevisiae from the Low-Cost Substrate Sucrose. FEMS Yeast Res. 2021, 21, foab062. [Google Scholar] [CrossRef] [PubMed]
- Farzadfard, F.; Perli, S.D.; Lu, T.K. Tunable and Multifunctional Eukaryotic Transcription Factors Based on CRISPR/Cas. ACS Synth. Biol. 2013, 2, 604–613. [Google Scholar] [CrossRef]
- Chee, M.K.; Haase, S.B. New and Redesigned PRS Plasmid Shuttle Vectors for Genetic Manipulation of Saccharomyces Cerevisiae. G3 Genes Genomes Genet. 2012, 2, 515–526. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. Frozen Competent Yeast Cells That Can Be Transformed with High Efficiency Using the LiAc/SS Carrier DNA/PEG Method. Nat. Protoc. 2007, 2, 1–4. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. High-Efficiency Yeast Transformation Using the LiAc/SS Carrier DNA/PEG Method. Nat. Protoc. 2007, 2, 31–34. [Google Scholar] [CrossRef]
- Zha, W.; An, T.; Li, T.; Zhu, J.; Gao, K.; Sun, Z.; Xu, W.; Lin, P.; Zi, J. Reconstruction of the Biosynthetic Pathway of Santalols under Control of the GAL Regulatory System in Yeast. ACS Synth. Biol. 2020, 9, 449–456. [Google Scholar] [CrossRef]
- Ro, D.-K.; Paradise, E.M.; Ouellet, M.; Fisher, K.J.; Newman, K.L.; Ndungu, J.M.; Ho, K.A.; Eachus, R.A.; Ham, T.S.; Kirby, J.; et al. Production of the Antimalarial Drug Precursor Artemisinic Acid in Engineered Yeast. Nature 2006, 440, 940–943. [Google Scholar] [CrossRef]
- Gueldener, U.; Heinisch, J.; Koehler, G.J.; Voss, D.; Hegemann, J.H. A Second Set of LoxP Marker Cassettes for Cre-Mediated Multiple Gene Knockouts in Budding Yeast. Nucleic Acids Res. 2002, 30, e23. [Google Scholar] [CrossRef] [PubMed]
- Reider Apel, A.; d’Espaux, L.; Wehrs, M.; Sachs, D.; Li, R.A.; Tong, G.J.; Garber, M.; Nnadi, O.; Zhuang, W.; Hillson, N.J.; et al. A Cas9-Based Toolkit to Program Gene Expression in Saccharomyces Cerevisiae. Nucleic Acids Res. 2016, 45, 496–508. [Google Scholar] [CrossRef]
- Hong, J.; Park, S.-H.; Kim, S.; Kim, S.-W.; Hahn, J.-S. Efficient Production of Lycopene in Saccharomyces Cerevisiae by Enzyme Engineering and Increasing Membrane Flexibility and NAPDH Production. Appl. Microbiol. Biotechnol. 2018, 103, 211–223. [Google Scholar] [CrossRef]
- Callari, R.; Meier, Y.; Ravasio, D.; Heider, H. Dynamic Control of ERG20 and ERG9 Expression for Improved Casbene Production in Saccharomyces Cerevisiae. Front. Bioeng. Biotechnol. 2018, 6, 160. [Google Scholar] [CrossRef] [PubMed]
- Conrado, R.J.; Varner, J.D.; DeLisa, M.P. Engineering the Spatial Organization of Metabolic Enzymes: Mimicking Nature’s Synergy. Curr. Opin. Biotechnol. 2008, 19, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Albertsen, L.; Chen, Y.; Bach, L.S.; Rattleff, S.; Maury, J.; Brix, S.; Nielsen, J.; Mortensen, U.H. Diversion of Flux toward Sesquiterpene Production in Saccharomyces Cerevisiae by Fusion of Host and Heterologous Enzymes. Appl. Environ. Microbiol. 2011, 77, 1033–1040. [Google Scholar] [CrossRef]
- Aalbers, F.S.; Fraaije, M.W. Enzyme Fusions in Biocatalysis: Coupling Reactions by Pairing Enzymes. Chembiochem 2019, 20, 20–28. [Google Scholar] [CrossRef]
- Chen, X.; Bai, Y.; Zaro, J.L.; Shen, W.C. Design of an in Vivo Cleavable Disulfide Linker in Recombinant Fusion Proteins. Biotechniques 2010, 49, 513–518. [Google Scholar] [CrossRef]
- Amet, N.; Lee, H.F.; Shen, W.C. Insertion of the Designed Helical Linker Led to Increased Expression of Tf-Based Fusion Proteins. Pharm. Res. 2009, 26, 523–528. [Google Scholar] [CrossRef]
- Reddy, S.T.; Ge, X.; Miklos, A.E.; Hughes, R.A.; Kang, S.H.; Hoi, K.H.; Chrysostomou, C.; Hunicke-Smith, S.P.; Iverson, B.L.; Tucker, P.W.; et al. Monoclonal Antibodies Isolated without Screening by Analyzing the Variable-Gene Repertoire of Plasma Cells. Nat. Biotechnol. 2010, 28, 965–969. [Google Scholar] [CrossRef]
- Bergeron, L.M.; Gomez, L.; Whitehead, T.A.; Clark, D.S. Self-Renaturing Enzymes: Design of an Enzyme-Chaperone Chimera as a New Approach to Enzyme Stabilization. Biotechnol. Bioeng. 2009, 102, 1316–1322. [Google Scholar] [CrossRef] [PubMed]
- Werner, S.; Marillonnet, S.; Hause, G.; Klimyuk, V.; Gleba, Y. Immunoabsorbent Nanoparticles Based on a Tobamovirus Displaying Protein A. Proc. Natl. Acad. Sci. USA 2006, 103, 17678–17683. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Feng, M.G. Bifunctional Enhancement of a Beta-Glucanase-Xylanase Fusion Enzyme by Optimization of Peptide Linkers. Appl. Microbiol. Biotechnol. 2008, 79, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.L.; Xue, C.; Du, J.L.; Ren, M.; Xia, S.; Liu, Z.M. Balancing the Pharmacokinetics and Pharmacodynamics of Interferon-A2b and Human Serum Albumin Fusion Protein by Proteolytic or Reductive Cleavage Increases Its in Vivo Therapeutic Efficacy. Mol. Pharm. 2012, 9, 664–670. [Google Scholar] [CrossRef]
- Guo, H.; Yang, Y.; Xue, F.; Zhang, H.; Huang, T.; Liu, W.; Liu, H.; Zhang, F.; Yang, M.; Liu, C.; et al. Effect of Flexible Linker Length on the Activity of Fusion Protein 4-Coumaroyl-CoA Ligase::Stilbene Synthase. Mol. Biosyst. 2017, 13, 598–606. [Google Scholar] [CrossRef]
- Weimer, T.; Wormsbächer, W.; Kronthaler, U.; Lang, W.; Liebing, U.; Schulte, S. Prolonged In-Vivo Half-Life of Factor VIIa by Fusion to Albumin. Thromb. Haemost. 2008, 99, 659–667. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, S.Z.; Li, J.; Pan, X.; Cahoon, R.E.; Jaworski, J.G.; Wang, X.; Jez, J.M.; Chen, F.; Yu, O. Using Unnatural Protein Fusions to Engineer Resveratrol Biosynthesis in Yeast and Mammalian Cells. J. Am. Chem. Soc. 2006, 128, 13030–13031. [Google Scholar] [CrossRef]
- Klein, J.S.; Jiang, S.; Galimidi, R.P.; Keeffe, J.R.; Bjorkman, P.J.; Regan, L. Design and Characterization of Structured Protein Linkers with Differing Flexibilities. Protein Eng. Des. Sel. 2014, 27, 325–330. [Google Scholar] [CrossRef]
- Subramaniam, S.; Kleywegt, G.J. A Paradigm Shift in Structural Biology. Nat. Methods 2022, 19, 20–23. [Google Scholar] [CrossRef]
- Noé, F.; De Fabritiis, G.; Clementi, C. Machine Learning for Protein Folding and Dynamics. Curr. Opin. Struct. Biol. 2020, 60, 77–84. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively Expanding the Structural Coverage of Protein-Sequence Space with High-Accuracy Models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhou, Y.J.; Bao, J.; Huang, L.; Nielsen, J.; Krivoruchko, A. Metabolic Engineering of Saccharomyces Cerevisiae for Production of Germacrene A, a Precursor of Beta-Elemene. J. Ind. Microbiol. Biotechnol. 2017, 44, 1065–1072. [Google Scholar] [CrossRef]
- Wang, Y.; Gong, X.; Li, F.; Zuo, S.; Li, M.; Zhao, J.; Han, X.; Wen, M. Optimized Biosynthesis of Santalenes and Santalols in Saccharomyces Cerevisiae. Appl. Microbiol. Biotechnol. 2021, 105, 8795–8804. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, M.; Zhao, G.R.; Lu, W. Harnessing Yeast Peroxisomes and Cytosol Acetyl-CoA for Sesquiterpene α-Humulene Production. J. Agric. Food Chem. 2020, 68, 1382–1389. [Google Scholar] [CrossRef]
- Kurosaki, F.; Kato, T.; Misawa, N.; Taura, F.; Kurosaki, F.; Kato, T.; Misawa, N.; Taura, F. Efficient Production of δ-Guaiene, an Aroma Sesquiterpene Compound Accumulated in Agarwood, by Mevalonate Pathway-Engineered Escherichia Coli Cells. Adv. Biosci. Biotechnol. 2016, 7, 435–445. [Google Scholar] [CrossRef]
- Yu, F.; Harada, H.; Yamasaki, K.; Okamoto, S.; Hirase, S.; Tanaka, Y.; Misawa, N.; Utsumi, R. Isolation and Functional Characterization of a Beta-Eudesmol Synthase, a New Sesquiterpene Synthase from Zingiber Zerumbet Smith. FEBS Lett. 2008, 582, 565–572. [Google Scholar] [CrossRef]
- An, T.; Li, L.; Lin, Y.; Zeng, F.; Lin, P.; Zi, J. Characterization of Guaiene Synthases from Stellera Chamaejasme L. Flowers and Their Application in de Novo Production of (-)-Rotundone in Yeast. J. Agric. Food Chem. 2020, 68, 3214–3219. [Google Scholar] [CrossRef]
- Dusséaux, S.; Wajn, W.T.; Liu, Y.; Ignea, C.; Kampranis, S.C. Transforming Yeast Peroxisomes into Microfactories for the Efficient Production of High-Value Isoprenoids. Proc. Natl. Acad. Sci. USA 2020, 117, 31789–31799. [Google Scholar] [CrossRef]
- Hoepfner, D.; Schildknegt, D.; Braakman, I.; Philippsen, P.; Tabak, H.F. Contribution of the Endoplasmic Reticulum to Peroxisome Formation. Cell 2005, 122, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Nielsen, J.; Liu, Z. Engineering Yeast Metabolism for Production of Terpenoids for Use as Perfume Ingredients, Pharmaceuticals and Biofuels. FEMS Yeast Res. 2017, 17, fox080. [Google Scholar] [CrossRef] [PubMed]
- Shiba, Y.; Paradise, E.M.; Kirby, J.; Ro, D.K.; Keasling, J.D. Engineering of the Pyruvate Dehydrogenase Bypass in Saccharomyces Cerevisiae for High-Level Production of Isoprenoids. Metab. Eng. 2007, 9, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Meadows, A.L.; Hawkins, K.M.; Tsegaye, Y.; Antipov, E.; Kim, Y.; Raetz, L.; Dahl, R.H.; Tai, A.; Mahatdejkul-Meadows, T.; Xu, L.; et al. Rewriting Yeast Central Carbon Metabolism for Industrial Isoprenoid Production. Nature 2016, 537, 694–697. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name | Genotype | Description |
|---|---|---|
| BCC39850hlu | Matα; his3Δ; leu2Δ; ura3Δ | BCC39850 with triple deletions (his3Δ, leu2Δ and ura3Δ) |
| FPPY001_39850 | BCC39850hlu YPRCΔ15c::PGAL1/10-ERG10-ERG20 | BCC39850hlu overexpressing ERG10 and ERG20 from S. cerevisiae |
| FPPY002_39850 | FPPY001_39850 ARS308a::PGAL1/10-tHMG1-ERG8 | FPPY001_39850 overexpressing tHMG1 and ERG8 from S. cerevisiae |
| FPPY003_39850 | FPPY002_39850 ARS1021::PGAL1/10-ERG13-IDI1 | FPPY002_39850 overexpressing ERG13 and IDI1 from S. cerevisiae |
| FPPY004_39850 | FPPY003_39850 ARS720a::PGAL1/10-tHMG1-ERG19 | FPPY003_39850 overexpressing tHMG1 and ERG19 from S. cerevisiae |
| FPPY005_39850 | FPPY004_39850 ARS1309::PGAL1/10-tHMG1-ERG12 | FPPY004_39850 overexpressing tHMG1 and ERG12 from S. cerevisiae |
| FPPY005_39850 PMET3-ERG9 | FPPY005_39850 ERG9::PMET3-ERG9 | FPPY005_39850 with the endogenous promoter of ERG9 replaced with the L-methionine repressible promoter (PMET3) |
| WWY005 PMET3-ERG9 | WWY005 ERG9::PMET3-ERG9 | WWY005 with the endogenous promoter of ERG9 replaced with the L-methionine repressible promoter (PMET3) |
| Plasmid Name | Overexpressed Gene(s) 1 | Promoter Used for Overexpression | Origin of Replication | Selectable Marker |
|---|---|---|---|---|
| pRSII416-ERG10-ERG20 | ERG10 and ERG20 from Saccharomyces cerevisiae | GAL1/GAL10 | CEN6/ARS4 | Ura3 |
| pRSII416-tHMG1-ERG12 | tHMG1 and ERG12 from S. cerevisiae | GAL1/GAL10 | CEN6/ARS4 | Ura3 |
| pRSII416-tHMG1-ERG8 | tHMG1 and ERG8 from S. cerevisiae | GAL1/GAL10 | CEN6/ARS4 | Ura3 |
| pRSII416-tHMG1-ERG19 | tHMG1 and ERG19 from S. cerevisiae | GAL1/GAL10 | CEN6/ARS4 | Ura3 |
| pRSII416-ERG13-IDI1 | ERG13 and IDI1 from S. cerevisiae | GAL1/GAL10 | CEN6/ARS4 | Ura3 |
| pRSII426-Gal1/10 | None | GAL1/GAL10 | 2 micron | Ura3 |
| pRSII426-Gal1/10-AsSesTPS | AsSesTPS from Aquilaria sinensis | GAL1 | 2 micron | Ura3 |
| pRSII426-Gal1/10-AsSesTPS1 | AsSesTPS1 from A. sinensis | GAL1 | 2 micron | Ura3 |
| pRSII426-Gal1/10-AsASS1 | AsASS1 from A. sinensis | GAL1 | 2 micron | Ura3 |
| pRSII426-Gal1/10-AcHS1 | AcHS1 from A. crassna | GAL1 | 2 micron | Ura3 |
| pRSII426-Gal1/10-AmdGS1 | AmdGS1 from A. microcarpa | GAL1 | 2 micron | Ura3 |
| pRSII426-Gal1/10-ZzBES2 | ZzBES2 from Zingiber zerumbet | GAL1 | 2 micron | Ura3 |
| pRSII426-Gal1/10-ClTPS2 | ClTPS2 (or ClSanSyn) from Clausena lansium | GAL1 | 2 micron | Ura3 |
| pRSII426-Gal1/10-FPPS-GSG-AcHS1 | ERG20 fused with AcHS1 from A. crassna (N→C; GSG linker) | GAL1 | 2 micron | Ura3 |
| pRSII426-Gal1/10-FPPS-GSG-AmdGS1 | ERG20 fused with AmdGS1 from A. microcarpa (N→C; GSG linker) | GAL1 | 2 micron | Ura3 |
| pRSII426-Gal1/10-FPPS-GSG-ZzBES2 | ERG20 fused with ZzBES2 from Z. zerumbet (N→C; GSG linker) | GAL1 | 2 micron | Ura3 |
| pRSII426-Gal1/10-FPPS-GSG-ClTPS2 | ERG20 fused with ClTPS2 from C. lansium (N→C; GSG linker) | GAL1 | 2 micron | Ura3 |
| pRSII426-Gal1/10-FPPS-GGGGS-AcHS1 | ERG20 fused with AcHS1 from A. crassna (N→C; GGGGS linker) | GAL1 | 2 micron | Ura3 |
| pRSII426-Gal1/10-FPPS-GGGGS-AmdGS1 | ERG20 fused with AmdGS1 from A. microcarpa (N→C; GGGGS linker) | GAL1 | 2 micron | Ura3 |
| pRSII426-Gal1/10-FPPS-GGGGS-ZzBES2 | ERG20 fused with ZzBES2 from Z. zerumbet (N→C; GGGGS linker) | GAL1 | 2 micron | Ura3 |
| pRSII426-Gal1/10-FPPS-GGGGS-ClTPS2 | ERG20 fused with ClTPS2 from C. lansium (N→C; GGGGS linker) | GAL1 | 2 micron | Ura3 |
| pRPR1-gRNA-ERG9p | Single-guide RNA for ERG9 promoter | RPR1 | 2 micron | Leu2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Promdonkoy, P.; Sornlek, W.; Preechakul, T.; Tanapongpipat, S.; Runguphan, W. Metabolic Engineering of Saccharomyces cerevisiae for Production of Fragrant Terpenoids from Agarwood and Sandalwood. Fermentation 2022, 8, 429. https://doi.org/10.3390/fermentation8090429
Promdonkoy P, Sornlek W, Preechakul T, Tanapongpipat S, Runguphan W. Metabolic Engineering of Saccharomyces cerevisiae for Production of Fragrant Terpenoids from Agarwood and Sandalwood. Fermentation. 2022; 8(9):429. https://doi.org/10.3390/fermentation8090429
Chicago/Turabian StylePromdonkoy, Peerada, Warasirin Sornlek, Thanchanok Preechakul, Sutipa Tanapongpipat, and Weerawat Runguphan. 2022. "Metabolic Engineering of Saccharomyces cerevisiae for Production of Fragrant Terpenoids from Agarwood and Sandalwood" Fermentation 8, no. 9: 429. https://doi.org/10.3390/fermentation8090429