
Genomic and Transcriptional Characteristics of Strain Rum-meliibacillus sp. TYF-LIM-RU47 with an Aptitude of Directly Producing Acetoin from Lignocellulose

Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Strains
2.2. Metabolite Analysis
2.3. Genome Sequencing, Assembly and Annotation
2.4. Total RNA Extraction, Reverse Transcription and Real-Time PCR
3. Results and Discussion
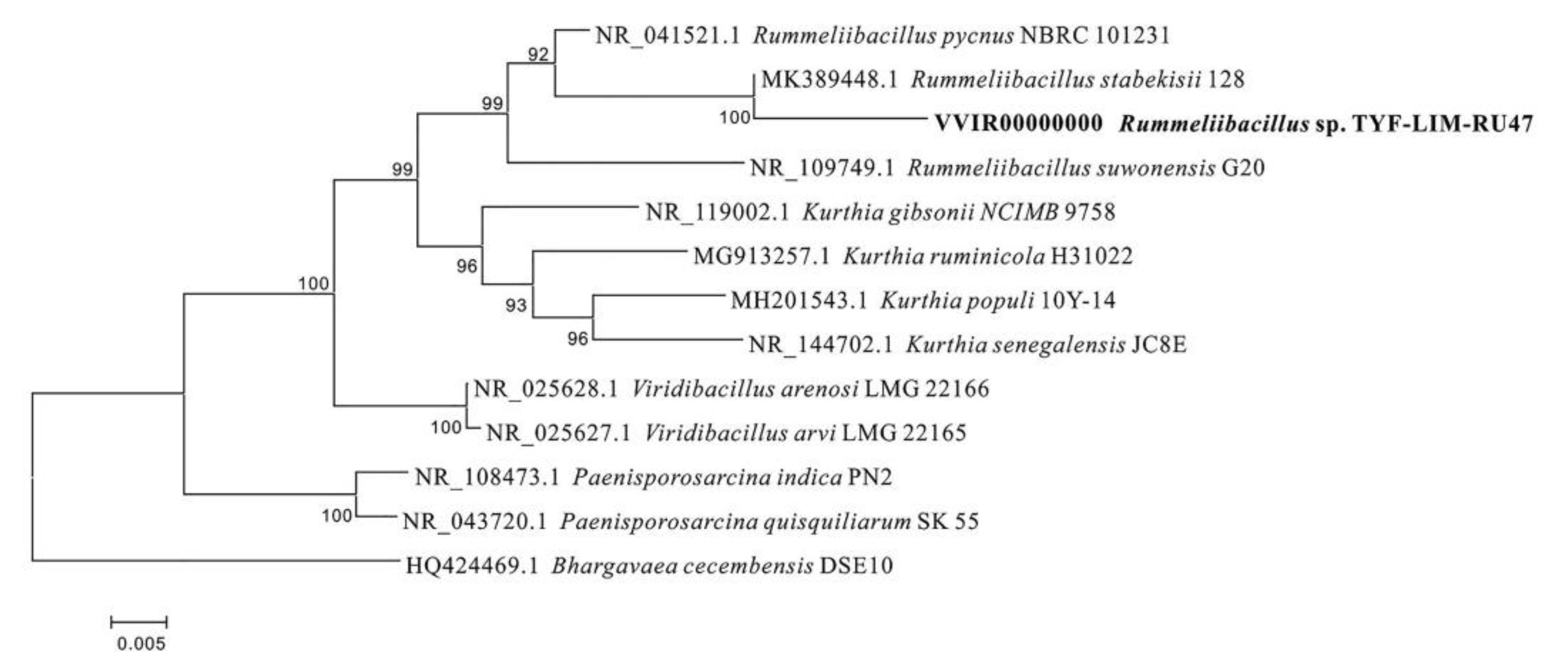
3.1. Identification of Strains
3.2. Metabolites Analysis
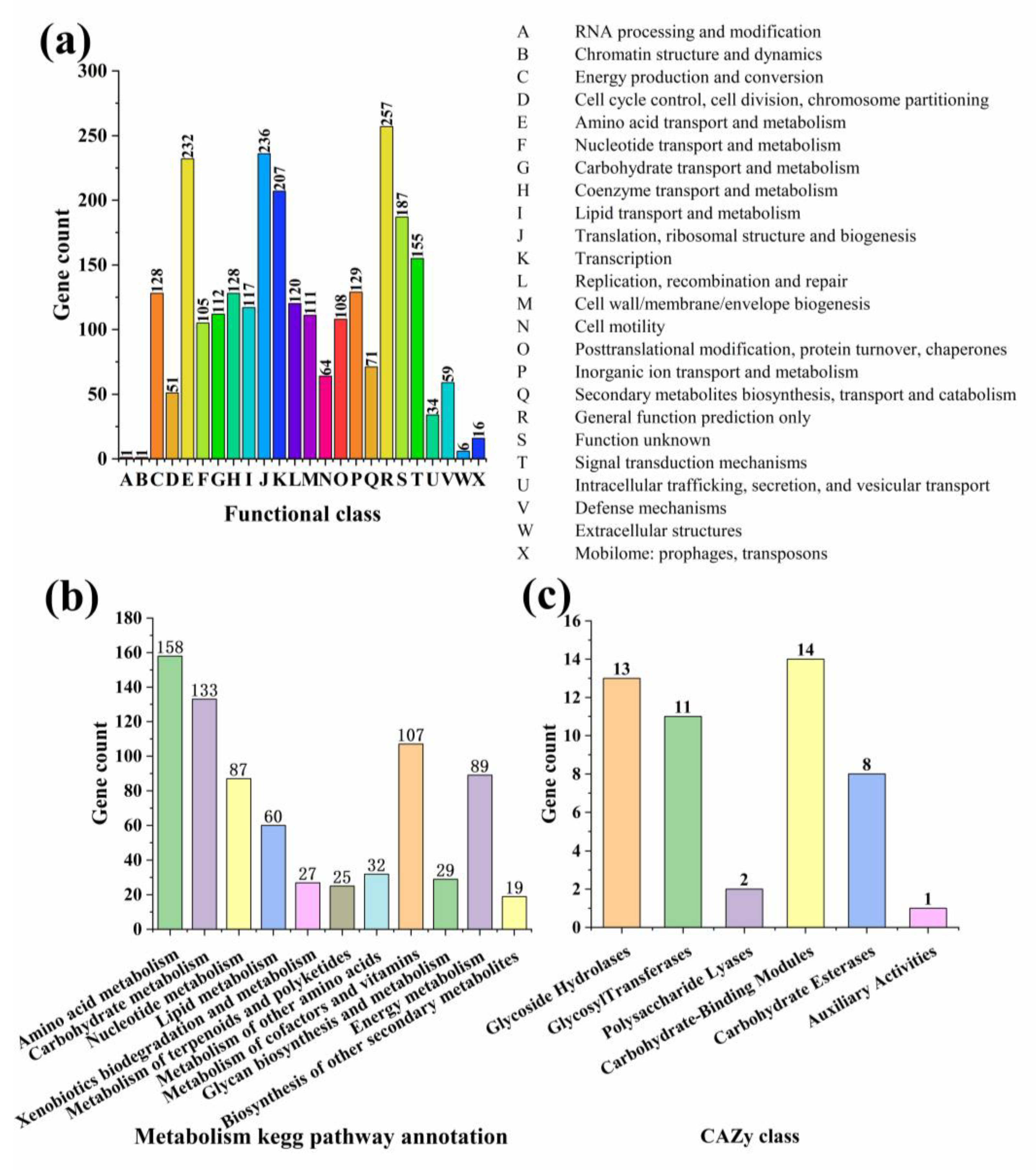
3.3. Genome Assembly and Annotation
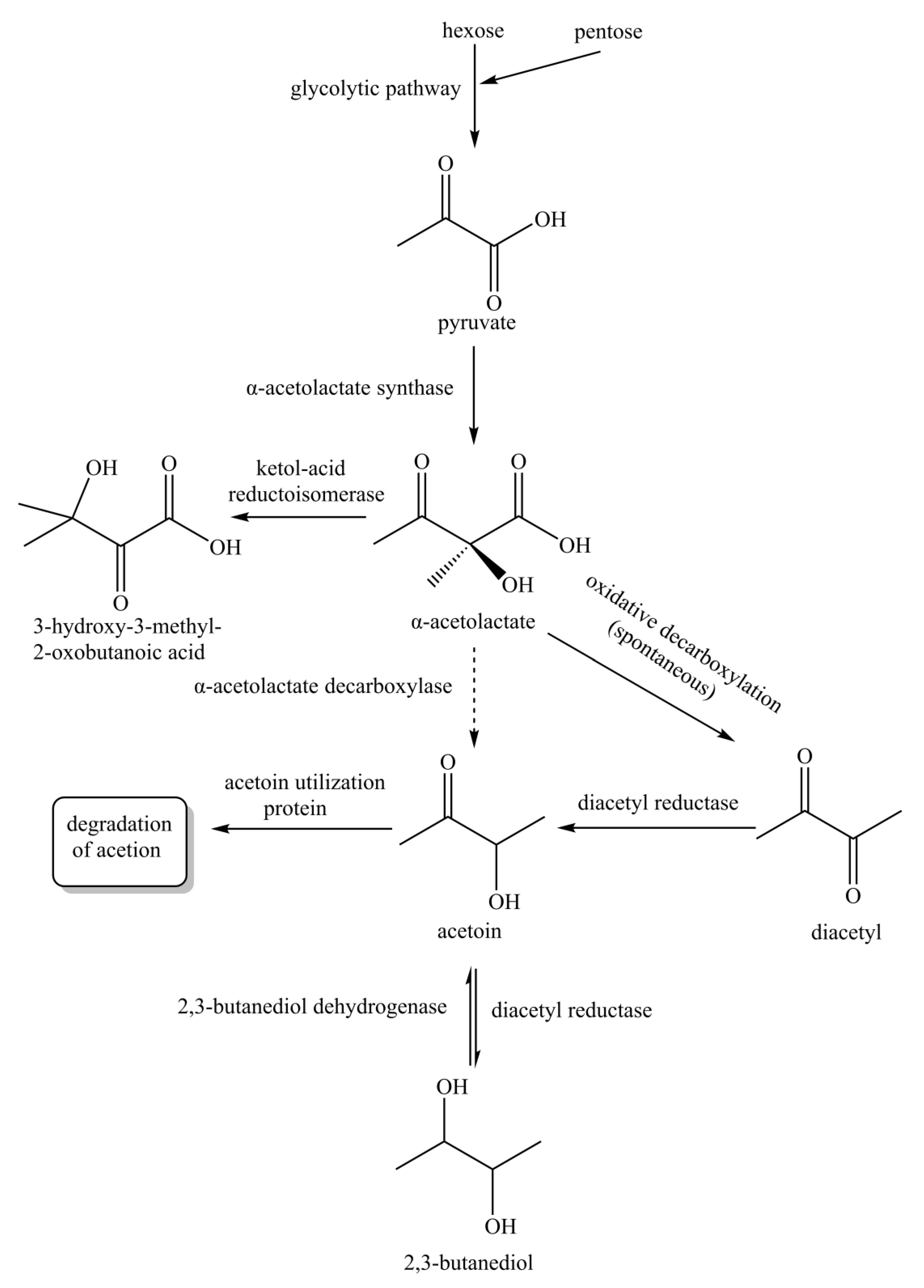
3.4. Acetoin Bioproduction Pathway of TYF-LIM-RU47
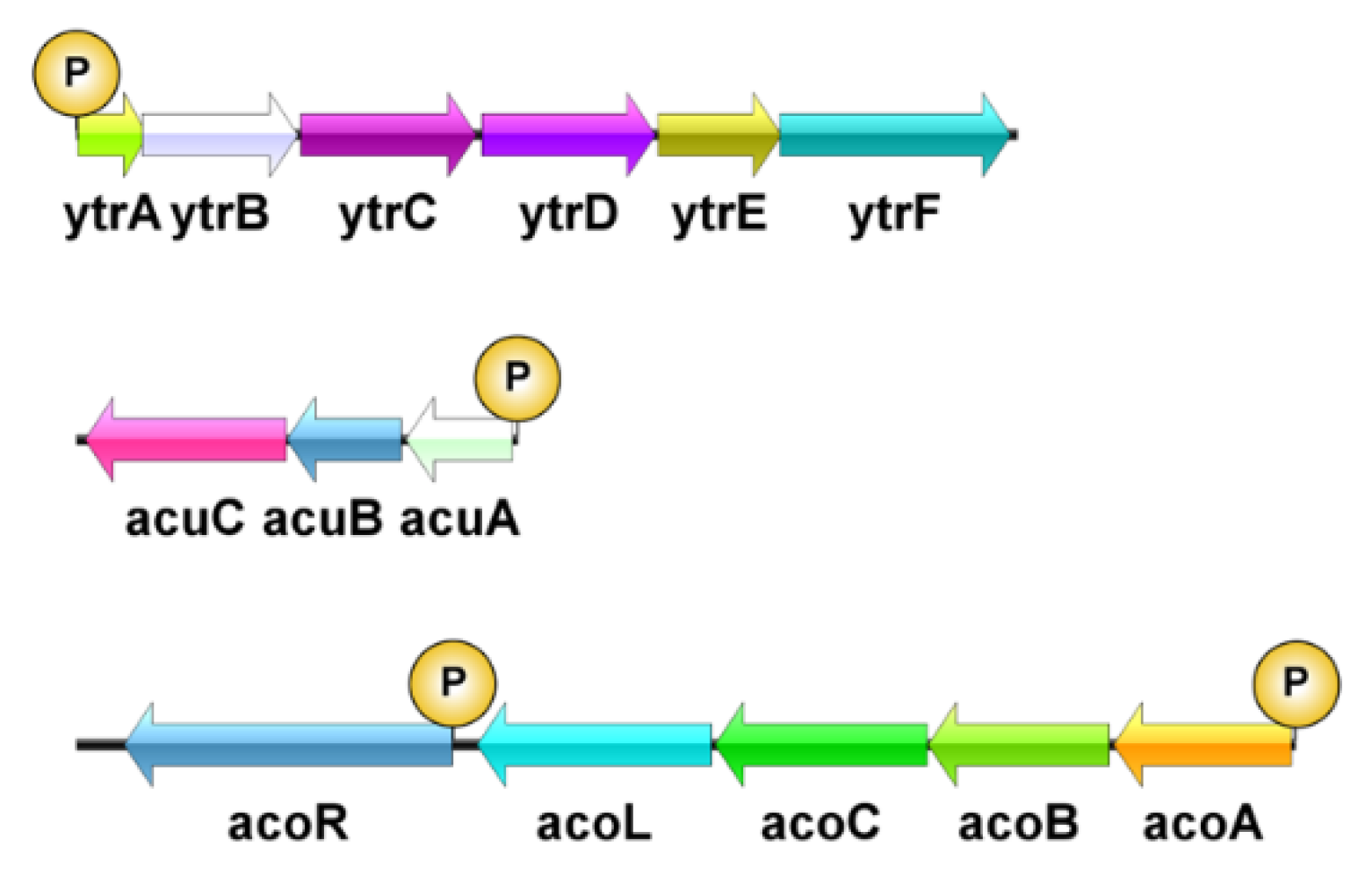
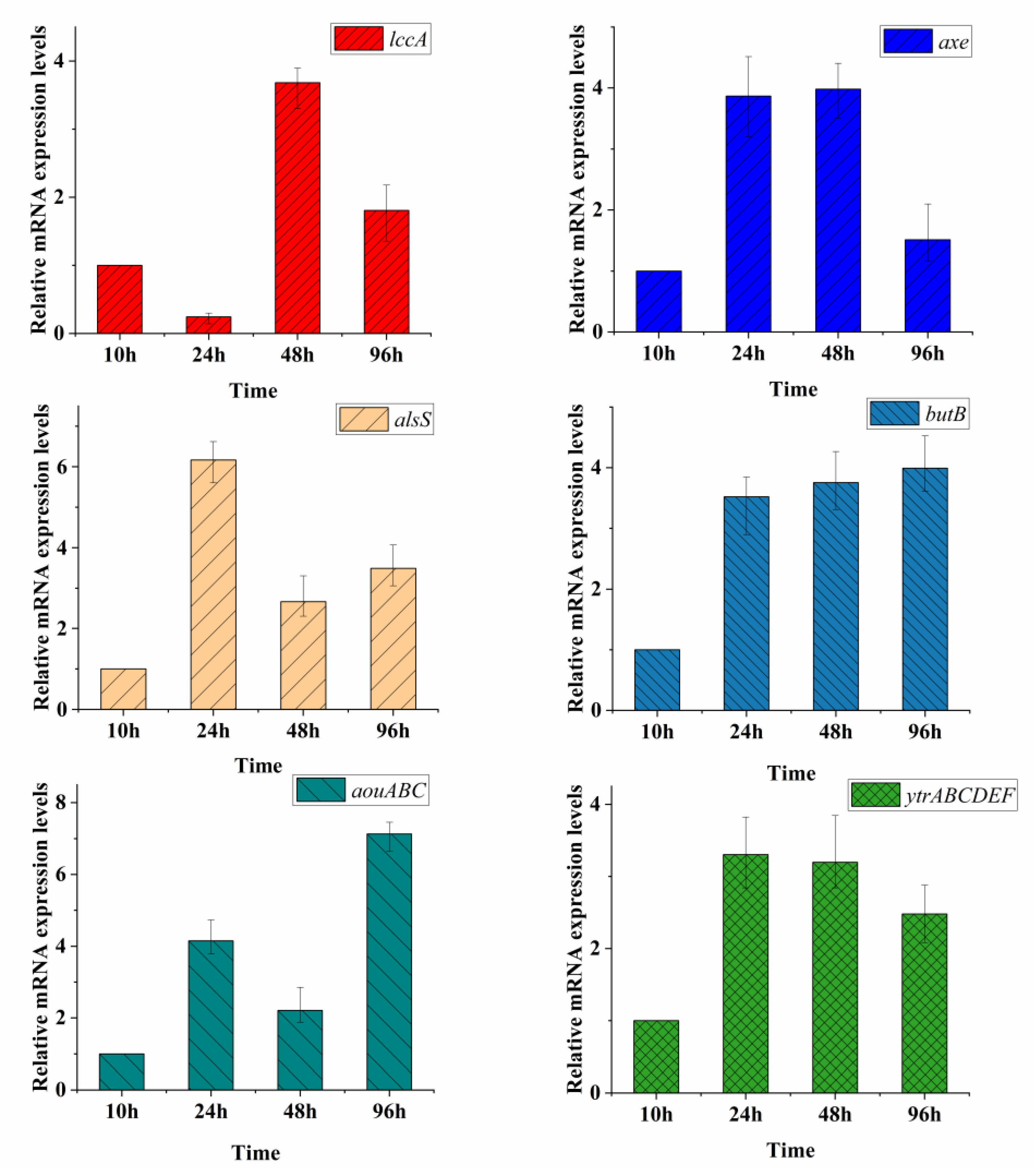
3.5. Transcriptional Level of Genes Related to Acetoin Metabolite
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tu, W.C.; Hallett, J.P. Recent advances in the pretreatment of lignocellulosic biomass. Curr. Opin. Green Sustain. Chem. 2019, 20, 11–17. [Google Scholar] [CrossRef]
- Kucharska, K.; Hołowacz, I.; Konopacka-Łyskawa, D.; Rybarczyk, P.; Kamiński, M. Key issues in modeling and optimization of lignocellulosic biomass fermentative conversion to gaseous biofuels. Renew. Energy 2018, 129, 384–408. [Google Scholar] [CrossRef]
- Shi, F.; Yu, H.; Zhang, N.; Wang, S.; Li, P.; Yu, Q.; Liu, J.; Pei, Z. Microbial succession of lignocellulose degrading bacteria during composting of corn stalk. Bioengineered 2021, 12, 12372–12382. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.Q.; Shen, Y.Q.; Liu, Y. State of the art of straw treatment technology: Challenges and solutions forward. Bioresour. Technol. 2020, 313, 123656. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wu, K.K.; Chen, C.; Ren, H.Y.; Wang, Z.H.; Nan, J.; Ren, N.Q. Role of residue cornstalk derived biochar for the enhanced bio-hydrogen production via simultaneous saccharification and fermentation of cornstalk. Bioresour. Technol. 2021, 330, 125006. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, Z.; Ren, H.Y.; Chen, C.; Nan, J.; Cao, G.L.; Yang, S.S.; Ren, N.Q. Residue cornstalk derived biochar promotes direct bio-hydrogen production from anaerobic fermentation of cornstalk. Bioresour. Technol. 2021, 320, 124338. [Google Scholar] [CrossRef]
- Xiao, Z.; Lu, J.R. Strategies for enhancing fermentative production of acetoin: A review. Biotechnol. Adv. 2014, 32, 492–503. [Google Scholar] [CrossRef]
- Zhang, B.; Li, X.L.; Fu, J.; Li, N.; Wang, Z.; Tang, Y.-J.; Chen, T. Production of acetoin through simultaneous utilization of glucose, xylose, and arabinose by engineered Bacillus subtilis. PLoS ONE 2016, 11, e0159298. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, R.Z.; Bao, T.; Yang, T.W.; Xu, M.J.; Li, H.Z.; Xu, Z.H.; Rao, Z.M. Moderate expression of the transcriptional regulator ALsR enhances acetoin production by Bacillus subtilis. J. Ind. Microbiol. Biotechnol. 2013, 40, 1067–1076. [Google Scholar] [CrossRef]
- Qiu, M.; Chen, C.F.; Zhang, A.Z.; Zhou, X.J. Partial Reduction of α-Diketones by Sodium Hydrogen Selenide. Synth. Commun. 1992, 22, 1529–1534. [Google Scholar] [CrossRef]
- Roncal, T.; Caballero, S.; Guereñu, M.D.; Rincón, I.; Prieto-Fernández, S.; Ochoa-Gómez, J.R. Efficient production of acetoin by fermentation using the newly isolated mutant strain Lactococcus lactis subsp. lactis CML B4. Process Biochem. 2017, 58, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, R.Z.; Bao, T.; Rao, Z.M.; Yang, T.W.; Xu, M.J.; Xu, Z.H.; Li, H.Z.; Yang, S.T. The rebalanced pathway significantly enhances acetoin production by disruption of acetoin reductase gene and moderate-expression of a new water-forming NADH oxidase in Bacillus subtilis. Metab. Eng. 2014, 23, 34–41. [Google Scholar] [CrossRef]
- Passerini, D.; Laroute, V.; Coddeville, M.; Le Bourgeois, P.; Loubière, P.; Ritzenthaler, P.; Cocaign-Bousquet, M.; Daveran-Mingot, M.L. New insights into Lactococcus lactis diacetyl-and acetoin-producing strains isolated from diverse origins. Int. J. Food Microbiol. 2013, 160, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Geckil, H.; Barak, Z.E.; Chipman, D.M.; Erenler, S.O.; Webster, D.A.; Stark, B.C. Enhanced production of acetoin and butanediol in recombinant Enterobacter aerogenes carrying Vitreoscilla hemoglobin gene. Bioprocess Biosyst. Eng. 2004, 26, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.N.; Zhang, L.Y.; Rao, B.; Han, Y.B.; Chu, J.; Zhu, J.W.; Shen, Y.L.; Wei, D.Z. Enhanced acetoin production by Serratia marcescens H32 using statistical optimization and a two-stage agitation speed control strategy. Biotechnol. Bioprocess Eng. 2012, 17, 598–605. [Google Scholar] [CrossRef]
- Luo, Q.L.; Wu, J.; Wu, M.C. Enhanced acetoin production by Bacillus amyloliquefaciens through improved acetoin tolerance. Process Biochem. 2014, 49, 1223–1230. [Google Scholar] [CrossRef]
- Jung, M.Y.; Jung, H.M.; Lee, J.; Oh, M.K. Alleviation of carbon catabolite repression in Enterobacter aerogenes for efficient utilization of sugarcane molasses for 2,3-butanediol production. Biotechnol. Biofuels 2015, 8, 106. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Jia, S.R.; Liu, J.J. Production of acetoin by Bacillus subtilis TH-49. In Proceedings of the 2011 International Conference on Consumer Electronics, Communications and Networks (CECNet), Xianning, China, 16–18 April 2011; pp. 1524–1527. [Google Scholar] [CrossRef]
- Breuil, C.; Saddler, J.N. Comparison of the 3, 5-dinitrosalicylic acid and Nelson-Somogyi methods of assaying for reducing sugars and determining cellulase activity. Enzym. Microb. Technol. 1985, 7, 327–332. [Google Scholar] [CrossRef]
- Lim, H.J.; Lee, E.H.; Yoon, Y.; Chua, B.; Son, A. Portable lysis apparatus for rapid single-step DNA extraction of Bacillus subtilis. J. Appl. Microbiol. 2016, 120, 379–387. [Google Scholar] [CrossRef] [Green Version]
- Li, R.Q.; Li, Y.R.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Zhu, H.; Ruan, J.; Qian, W.; Fang, X.; Shi, Z.; Li, Y.; Li, S.; Shan, G.; Kristiansen, K.; et al. De novo assembly of human genomes with massively parallel short read sequencing. Genome Res. 2010, 20, 265–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, P.P.; Daub, J.; Tate, J.G.; Nawrocki, E.P.; Kolbe, D.L.; Lindgreen, S.; Wilkinson, A.C.; Finn, R.D.; Griffiths-Jones, S.; Eddy, S.R.; et al. Rfam: Updates to the RNA families database. Nucleic Acids Res. 2009, 37, D136–D140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, W.; Wan, I.; Jones, S.J.; Brinkman, F.S. IslandPath: Aiding detection of genomic islands in prokaryotes. Bioinformatics 2003, 19, 418–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawrocki, E.P.; Kolbe, D.L.; Eddy, S.R. Infernal 1.0: Inference of RNA alignments. Bioinformatics 2009, 25, 1335–1337. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Liang, Y.; Lynch, K.H.; Dennis, J.J.; Wishart, D.S. PHAST: A fast phage search tool. Nucleic Acids Res. 2011, 39, W347–W352. [Google Scholar] [CrossRef]
- Grissa, I.; Vergnaud, G.; Pourcel, C. CRISPRFinder: A web tool to identify clustered regularly interspaced short palindromic repeats. Nucleic Acids Res. 2007, 35, W52–W57. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Andlar, M.; Rezić, T.; Marđetko, N.; Kracher, D.; Ludwig, R.; Šantek, B. Lignocellulose degradation: An overview of fungi and fungal enzymes involved in lignocellulose degradation. Eng. Life Sci. 2018, 18, 768–778. [Google Scholar] [CrossRef]
- Grundy, F.J.; Waters, D.A.; Takova, T.Y.; Henkin, T.M. Identification of genes involved in utilization of acetate and acetoin in Bacillus subtilis. Mol. Microbiol. 1993, 10, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.I.; Fujita, Y.; Ehrlich, S.D. An operon for a putative ATP-binding cassette transport system involved in acetoin utilization of Bacillus subtilis. J. Bacteriol. 2000, 182, 5454–5461. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Description | Primer Sequence (5′-3′) Forward/Reverse | Product (bp) |
|---|---|---|---|
| 16S rRNA | 16S rRNA | AGAGTTTGATCCTGGCTCAG GGTTACCTTGTTACGACTT | 1500 |
| GAPDH | glyceraldehyde-3 phosphate dehydrogenase | GTCGTATCGGCCGTATGGTT CAAAGCTGCTTTGCTACGCT | 293 |
| lccA | laccase | GGCTGGCAAGGGACGATTAA TGGTAATGTGAGCGCTTGGT | 280 |
| axe | acetylxylan esterase | ACCAACGCTATTCTCGCTCC TGGCCATCTGCGTACACTTT | 206 |
| alsS | α-acetolactate synthase | ACAGGTGCTGCTAATGCCAT GCTGTTCTCTTGGCAGGAGT | 299 |
| butB | diacetyl reductase | GGCAGGCATTTGTGGAAGTG TCCACCGTCTCCATGCAATC | 271 |
| acuABC | acetoin utilization protein acuABC | TGCTTCAGGTGTCGATGCTT TCCGTTGCAGCAACCTCTAG | 166 |
| ytrABCDEF | operon related to acetoin transport ytrABCDEF | TAGGGGTCTTAGGGACGGTG TAGTTGCCTTTCGAGCTGGG | 225 |
| Substrate | Keep Time (min) | Product Name | Relative Content % |
|---|---|---|---|
| Corn straw | 6.815 | Ethanol | 4.41 |
| 7.538 | Acetone | 10.88 | |
| 10.146 | 2,3-butanedione | 18.12 | |
| 10.45 | Butanone | 2.93 | |
| 12.341 | Valeraldehyde | 3.38 | |
| 12.627 | Ethyl allyl ether | 1.71 | |
| 15.524 | Acetoin | 53.88 | |
| 16.077 | Propane | 1.23 | |
| 22.071 | Propionaldehyde | 0.92 | |
| 25.19 | Benzaldehyde | 1.51 |
| Attribute | Value |
|---|---|
| GC content (%) | 38.37 |
| Scaffolds (>500 bp) | 14 |
| Scaffold N50 (bp) | 2,231,473 |
| Scaffold N90 (bp) | 188,997 |
| Total sequence length (bp) | 3,298,449 |
| Gene length (bp) | 2,834,418 |
| Total number of genes | 3418 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, G.; Fan, X.; Liang, Y.; Li, C.; Xing, J.; He, Y. Genomic and Transcriptional Characteristics of Strain Rum-meliibacillus sp. TYF-LIM-RU47 with an Aptitude of Directly Producing Acetoin from Lignocellulose. Fermentation 2022, 8, 414. https://doi.org/10.3390/fermentation8080414
Feng G, Fan X, Liang Y, Li C, Xing J, He Y. Genomic and Transcriptional Characteristics of Strain Rum-meliibacillus sp. TYF-LIM-RU47 with an Aptitude of Directly Producing Acetoin from Lignocellulose. Fermentation. 2022; 8(8):414. https://doi.org/10.3390/fermentation8080414
Chicago/Turabian StyleFeng, Guoyang, Xiaojun Fan, Yanna Liang, Chen Li, Junde Xing, and Yongji He. 2022. "Genomic and Transcriptional Characteristics of Strain Rum-meliibacillus sp. TYF-LIM-RU47 with an Aptitude of Directly Producing Acetoin from Lignocellulose" Fermentation 8, no. 8: 414. https://doi.org/10.3390/fermentation8080414