
Screening and Molecular Identification of Lactic Acid Bacteria Producing β-Glucan in Boza and Cider

 ,
,  ,
,  , , ,
, , ,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Collection and Preparation of Samples
2.3. Bacterial Isolation, Purification, and Growth Conditions
2.4. Genomic DNA Extraction
2.5. Amplification of 16S rRNA and gtf Genes
2.6. Sequencing of Amplicon (16S rRNA Product) and Alignment
2.7. Blast N and Phylogenetic Tree Analysis
2.8. Gene Expression
2.8.1. RNA Extraction
2.8.2. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) for the gtf Gene Using LAB 16S rRNA as a Housekeeping Gene
2.9. The Screening Four LAB Isolates Using L. rhamnosus NRRL 1937 (LGG) as a Control to Produce β-Glucan
2.10. Total Carbohydrate and Protein Analysis
2.11. Determination of β-Glucans Extracted from LAB Isolates by HPLC
2.12. Statistical Analysis
3. Results and Discussions
3.1. Isolation and Purification of Lactic Acid Bacteria (LAB)
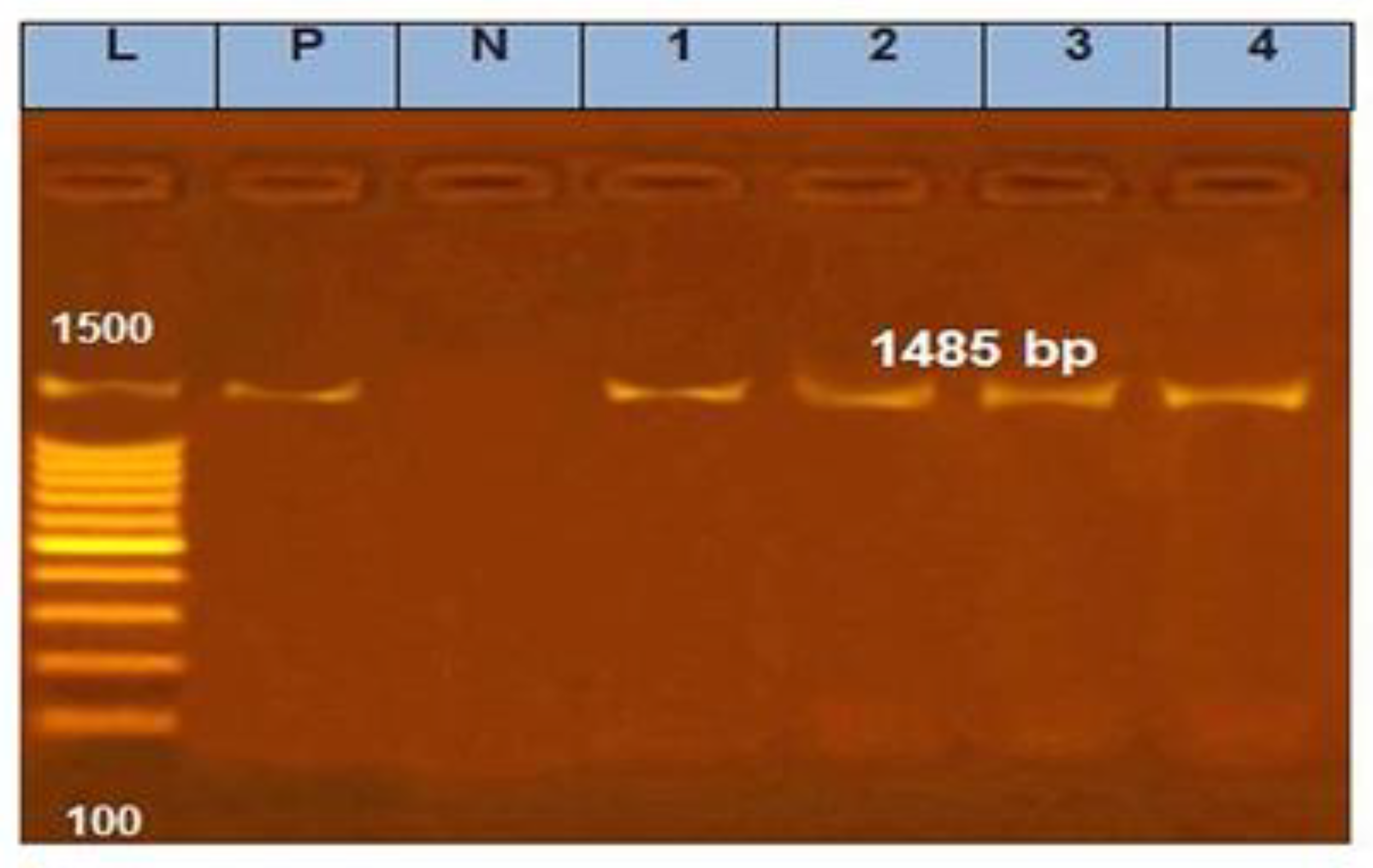
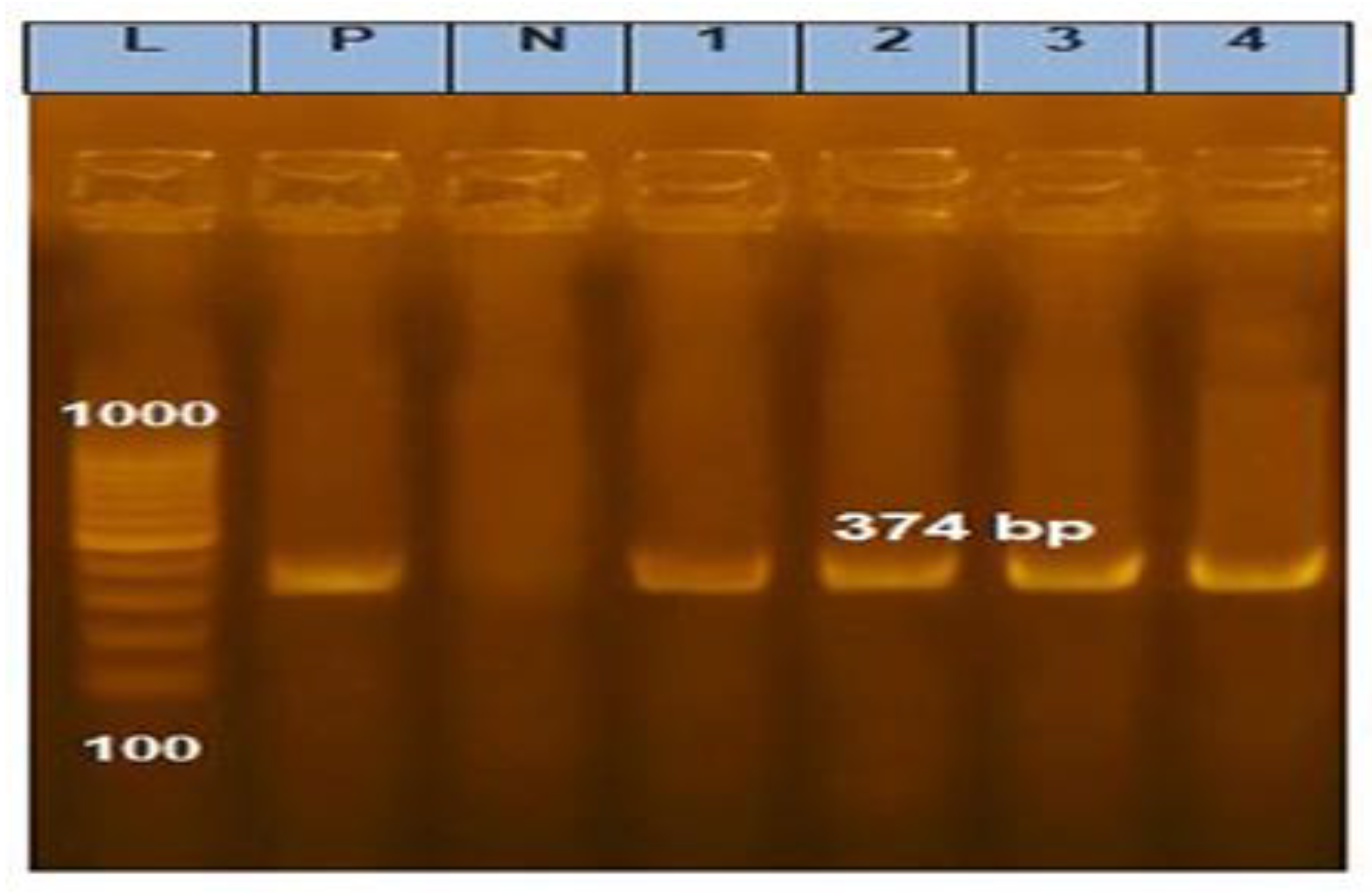
3.2. Amplification of 16S rRNA and gtf Gene for Selected LAB Isolates
3.3. Sequencing of Amplicon (16S rRNA Product) and Alignment for Selected LAB Isolates
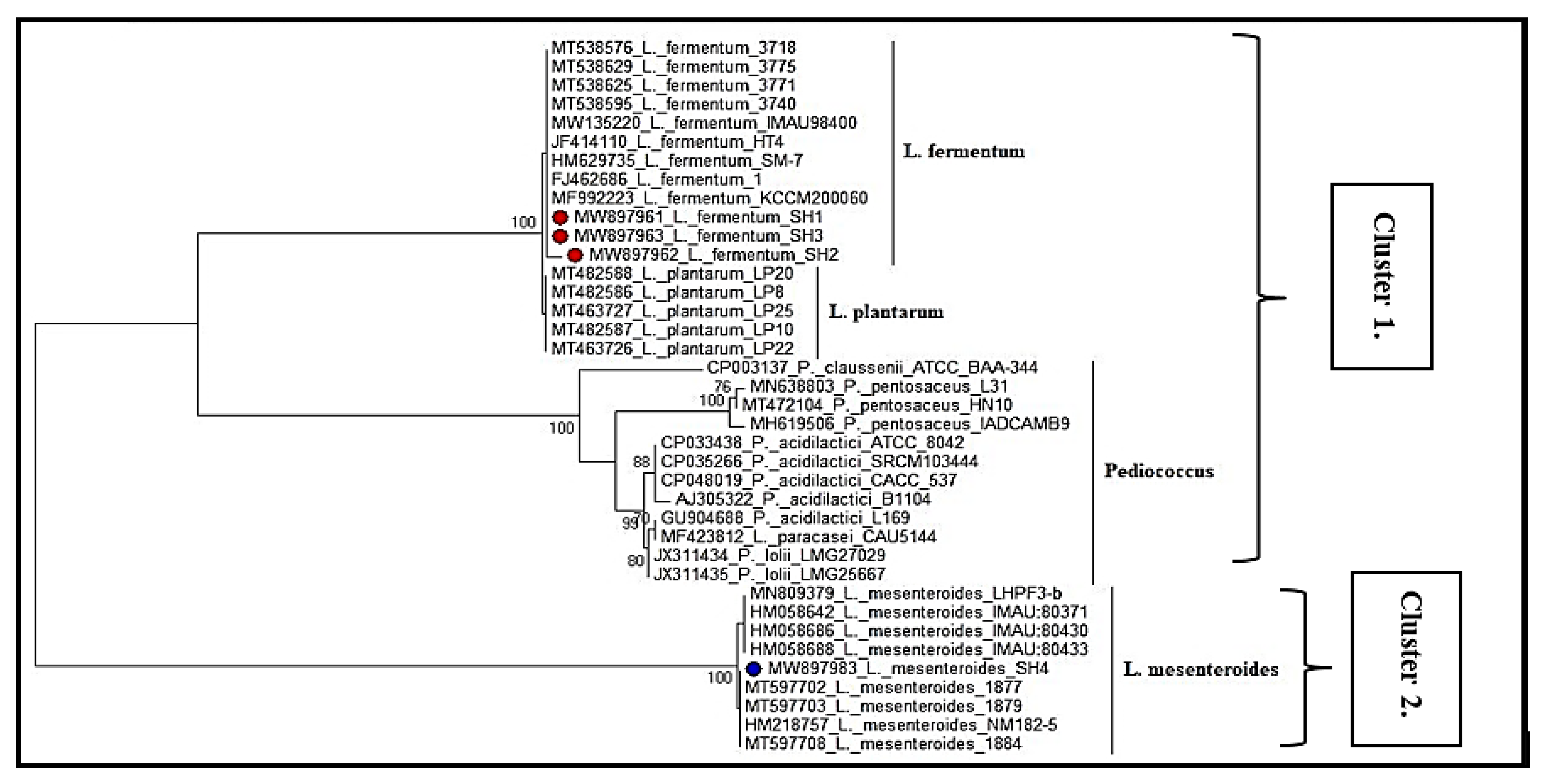
3.4. Blast N and Phylogenetic Tree Analysis of Selected LAB Isolates
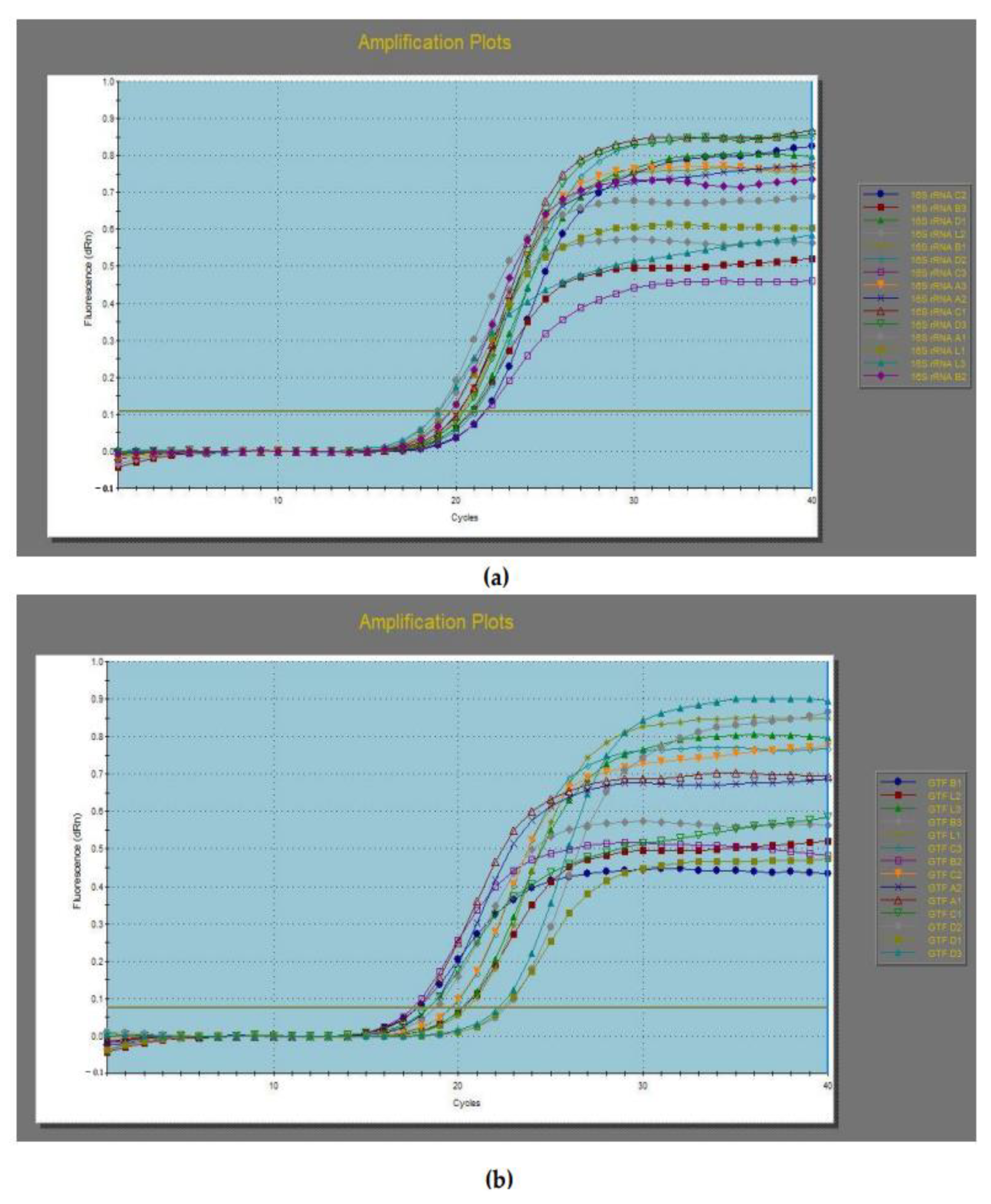
3.5. Gene Expression of Selected LAB Isolates by Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
3.6. The Screening of Four LAB Isolates Using L. rhamnosus NRRL 1937 (LGG) as a Control to Produce β-Glucan
3.6.1. Production and Calculation of EPS, Cell Mass, and pH Levels for Selected LAB Isolates
3.6.2. Determination of Total Carbohydrate and Total Protein in EPS Produced by Selected LAB Isolates
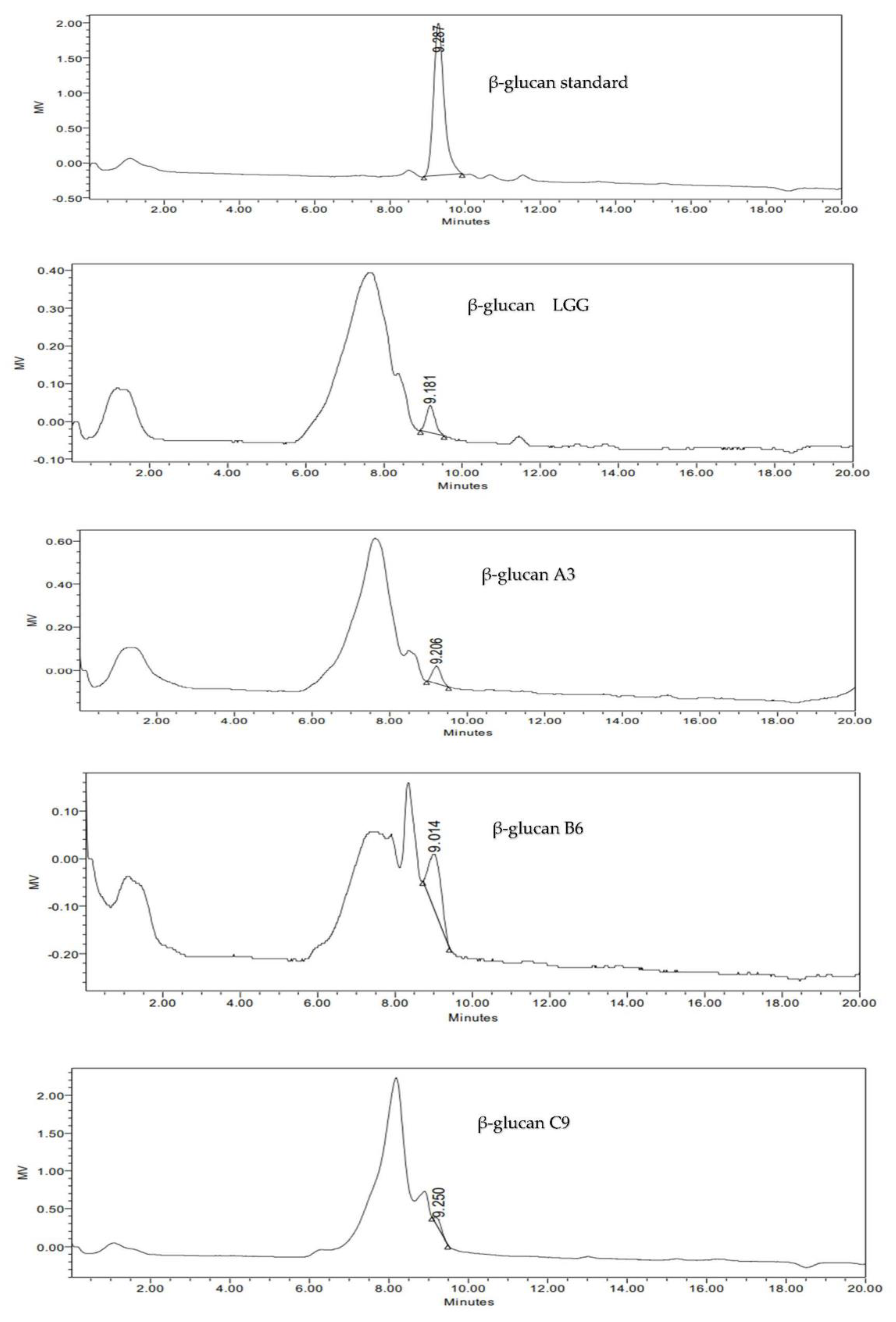
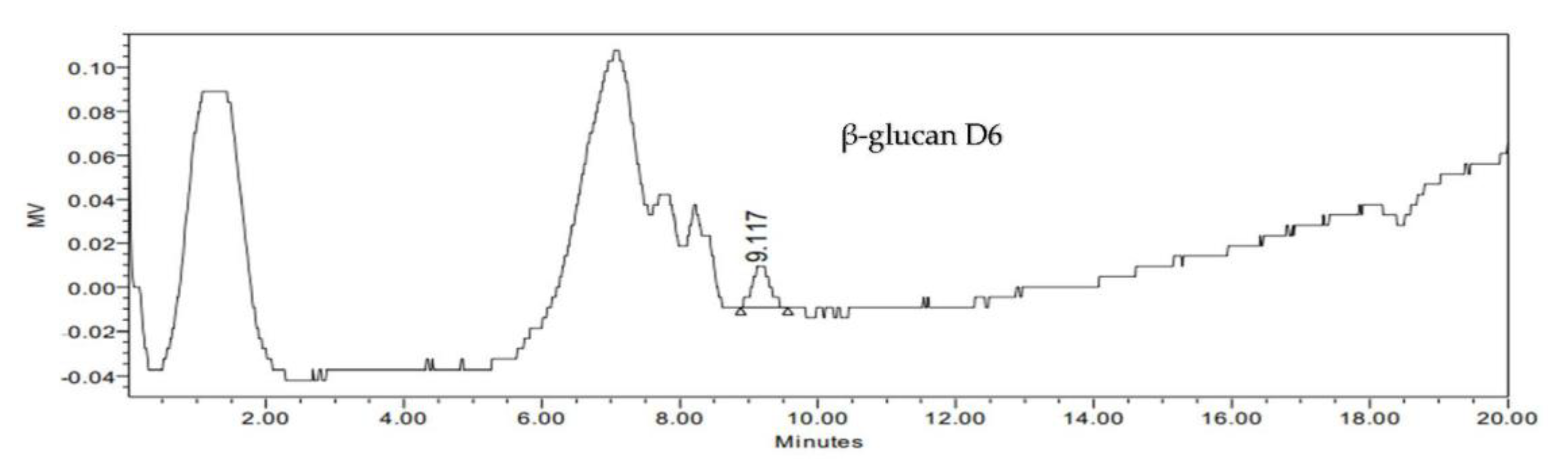
3.6.3. Detection and Identification of β-Glucan Produced by Selected LAB Isolates Using HPLC Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Henrion, M.; Francey, C.; Lê, K.-A.; Lamothe, L. Cereal _β-glucans: The Impact of Processing and How It affects Physiological Responses. Nutrients 2019, 11, 1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamothe, L.M.; Lê, K.-A.; Samra, R.A.; Roger, O.; Green, H.; Macé, K. The scientific basis for healthful carbohydrate profile. Crit. Rev. Food Sci. Nutr. 2019, 59, 1058–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abd El Ghany, K.; Hamouda, R.A.; Mahrous, H.A.; Sharaf, S.M.; Hamza, H.A. Molecular characterization using gtf gene detection and medium optimization of β -d-glucan production from Pediococcus parvulus F1030 isolated from local Egyptian boza. J. Pharm. Res. Int. 2017, 1–8. [Google Scholar]
- Donot, F.; Fontana, A.; Baccou, J.C.; Schorr-Galindo, S. Microbial Exopolysaccharides: Main examples of synthesis, excretion, genetics and extraction. Carbohydr. Polym. 2012, 87, 951–962. [Google Scholar] [CrossRef]
- Flemming, H.-C. EPS—Then and Now. Microorganisms 2016, 4, 41. [Google Scholar] [CrossRef] [Green Version]
- Verma, D.K.; Niamah, A.K.; Patel, A.R.; Thakur, M.; Sandhu, K.S.; Chávez-González, M.L.; Shah, N.; Aguilar, C.N. Chemistry and microbial sources of curdlan with potential application and safety regulations as prebiotic in food and health. Food Res. Int. 2020, 133, 109–136. [Google Scholar] [CrossRef]
- Zhu, F.; Du, B.; Xu, B. A critical review on production and industrial applications of β -glucans. Food Hydrocoll. 2016, 52, 275–288. [Google Scholar] [CrossRef]
- Schlörmann, W.; Bockwoldt, J.A.; Hübner, S.M.; Wittwer, E.; Reiners, S.; Lorkowski, S.; Dawczynski, C.; Ehrmann, M.A.; Glei, M. Use of the β-Glucan-Producing Lactic Acid Bacteria Strains Levilactobacillus brevis and Pediococcus claussenii for Sourdough Fermentation—Chemical Characterization and Chemopreventive Potential of In Situ-Enriched Wheat and Rye Sourdoughs and Breads. Nutrients 2022, 14, 1510. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration. GRAS Notice Inventory. Available online: http://www.fda.gov/Food/FoodIngredientsPackaging/GenerallyRecognizedasSafeGRAS/GRASListings/default.htm (accessed on 11 September 2009).
- Sutherland, I.W. Novel and established applications of microbial polysaccharides. Trends Biotechnol. 1998, 16, 41–46. [Google Scholar] [CrossRef]
- Miao, M.; Bai, A.; Jiang, B.; Song, Y.; Cui, S.W.; Zhang, T. Characterisation of a novel water-soluble polysaccharide from Leuconostoc citreum SK24. 002. Food Hydrocoll. 2014, 36, 265–272. [Google Scholar] [CrossRef]
- Verce, M.; De Vuyst, L.; Weckx, S. Comparative genomics of Lactobillus fermentum suggests a free-living lifestyle of this lactic acid bacterial species. Food microbiol. 2020, 89, 103448. [Google Scholar] [CrossRef] [PubMed]
- Ilango, S.; Antony, U. Probiotic microorganisms from non-dairy traditional fermented foods. Trends Food Sci. Tech. 2021, 118, 617–638. [Google Scholar] [CrossRef]
- Duenas, M.; Irastorza, A.; Fernandez, K.; Bilbao, A.; Huerta, A. Microbial populations and malolactic fermentation of apple cider using traditional and modified methods. J. Food Sci. 1994, 59, 1060–1064. [Google Scholar] [CrossRef]
- Marshall, E.; Mejia, D. Fermentation and sustainable livelihoods. In Traditional Fermented Food and Beverages for Improved Livelihoods, 1st ed.; Marshall, E., Ed.; FAO: Rome, Italy, 2011; Volume 18. [Google Scholar]
- Abd El Ghany, K.; Hamouda, R.; Mahrous, H.; Abd Elhafez, E.; Ahmed, F.A.H.; Hamza, H.A. Description of isolated LAB producing β -glucan from Egyptian sources and evaluation of its therapeutic effect. Int. J. Pharm. 2016, 12, 801–811. [Google Scholar] [CrossRef] [Green Version]
- Nel, S.; Davis, S.B.; Endo, A.; Dicks, L.M. Phylogenetic analysis of Leuconostoc and Lactobacillus species isolated from sugarcane processing streams. MicrobiologyOpen. 2020, 9, e1065. [Google Scholar] [CrossRef] [PubMed]
- Kurek, M.A.; Wyrwisz, J.; Brzeska, M.; Moczkowska, M.; Karp, S.; Wierzbicka, A. Effect of different β -glucan preparation pretreatments on fortified bread quality. Food Sci.Technol. 2018, 38, 606–611. [Google Scholar] [CrossRef] [Green Version]
- Hugenholtz, P.; Goebel, B.M.; Pace, N.R. Impact of culture independent studies on the emerging phylogenetic view of bacterial diversity. J. Bacteriol. 1998, 180, 4765–4774. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, D.J. Methods for analysis of the intestinal microflora. Curr. Issues Intest. Microbiol. 2000, 1, 39–50. [Google Scholar]
- Martinez-Murcia, A.J.; Collins, M.D. A phylogenetic analysis of the genus Leuconostoc based on reverse transcriptase sequencing of 16S rRNA. FEMS Microbiol. Lett. 1990, 70, 73–83. [Google Scholar] [CrossRef]
- Sanalibaba, P.; Çakmak, G.A. Exopolysaccharides production by lactic acid bacteria. Appl. Microbiol. 2016, 2, 1000115. [Google Scholar] [CrossRef]
- Werning, M.L.; Ibarburu, I.; Duenas, M.T.; Irastorza, A.N.A.; Navas, J.; Lopez, P. Pediococcus parvulus gtf gene encoding the GTF glycosyltransferase and its application for specific PCR detection of β-D-glucan–producing bacteria in foods and beverages. J. Food Prot. 2006, 69, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Garai-Ibabe, G.; Dueñas, M.T.; Irastorza, A.; Sierra-Filardi, E.; Werning, M.L.; López, P.; De Palencia, P.F. Naturally occurring 2-substituted (1, 3)-β-D-glucan producing Lactobacillus suebicus and Pediococcus parvulus strains with potential utility in the production of functional foods. Bioresource Technol. 2010, 101, 9254–9263. [Google Scholar] [CrossRef] [Green Version]
- Kralj, S.; van Geel-Schutten, G.H.; Dondorff, M.M.G.; Kirsanovs, S.; Van Der Maarel, M.J.E.C.; Dijkhuizen, L. Glucan sy thesis in the genus Lactobacillus: Isolation and characterization of glucansucrase genes, enzymes and glucan products from six different strains. Microbiology 2004, 150, 3681–3690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valles, B.S.; Bedriñana, R.P.; Tascón, N.F.; Simón, A.Q.; Madrera, R.R. Yeast species associated with the spontaneous fermentation of cider. Food Microbiol. 2007, 24, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Lagacé, L.; Pitre, M.; Jacques, M.; Roy, D. Identification of the bacterial community of maple sap by using amplified ribosomal DNA (rDNA) restriction analysis and rDNA sequencing. Appl. Environ. Microbiol. 2004, 70, 2052–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turpin, W.; Humblot, C.; Guyot, J.P. Genetic screening of functional properties of lactic acid bacteria in a fermented pearl millet slurry and in the metagenome of fermented starchy foods. Appl. Environ. Microbiol. 2011, 77, 8722–8734. [Google Scholar] [CrossRef] [Green Version]
- Green, M.R.; Hughes, H.; Sambrook, J.; MacCallum, P. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2012; p. 1890. [Google Scholar]
- Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi#alnHdr_2024212275 (accessed on 18 July 2022).
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. Clustal W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Chon, J.W.; Kim, H.; Kim, H.S.; Choi, D.; Hwang, D.G.; Seo, K.H. Detection and enumeration of lactic acid bacteria, acetic acid bacteria and yeast in kefir grain and milk using Quantitative Real Time PCR. J. Food Saf. 2015, 35, 102–107. [Google Scholar] [CrossRef]
- Yuan, J.S.; Reed, A.; Chen, F.; Stewart, C.N. Statistical analysis of real-time PCR data. BMC Bioinform. 2006, 7, 85. [Google Scholar] [CrossRef] [Green Version]
- Ale, E.C.; Batistela, V.A.; Correa Olivar, G.; Ferrado, J.B.; Sadiq, S.; Ahmed, H.I.; Reinheimer, J.A.; Vera-Candioti, L.; Laws, A.P.; Binetti, A.G. Statistical optimisation of the exopolysaccharide production by Lactobacillus fermentum Lf2 and analysis of its chemical composition. Int. J. Dairy Technol. 2020, 73, 76–87. [Google Scholar] [CrossRef]
- Othman, N.Z.; Din, A.R.J.M.; Azam, Z.M.; Rosli, M.A.; Sarmidi, M.R. Statistical optimization of medium compositions for high cell mass and exopolysaccharide production by Lactobacillus plantarum ATCC 8014. Appl. Food Biotechnol. 2018, 5, 87–96. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Classics Lowry, O.; Rosebrough, N.; Farr, A.; Randall, R. Protein measurement with the Folin phenol reagent. J. Biol Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Allaith, S.A. Genotyping and hemolytic characterization of pathogenic bacteria from some raw and cooked foods. Iraqi J. Agric. Sci. 2018, 49, 5. [Google Scholar]
- Balcázar, J.L.; De Blas, I.; Ruiz-Zarzuela, I.; Vendrell, D.; Girones, O.; Muzquiz, J.L. Sequencing of variable regions of the 16S rRNA gene for identification of lactic acid bacteria isolated from the intestinal microbiota of healthy salmonids. Comp. Immunol. Microbiol. Infect. Dis. 2007, 30, 111–118. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Fowler, V.J.; Woese, C.R. A phylogenetic analysis of lactobacilli, Pediococcus pentosaceus and Leuconostoc mesenteroides. Syst. Appl. Microbiol. 1983, 4, 326–337. [Google Scholar] [CrossRef]
- Goh, Y.J.; Klaenhammer, T.R. Functional roles of aggregation promoting like factor in stress tolerance and adherence of Lactobacillus acidophilus NCFM. Appl. Environ. Microbiol. 2010, 76, 5005–5012. [Google Scholar] [CrossRef] [Green Version]
- Llamas-Arriba, M.G.; Pérez-Ramos, A.; Puertas, A.I.; López, P.; Dueñas, M.T.; Prieto, A. Characterization of Pediococcus ethanolidurans CUPV141: A β-D-glucan-and heteropolysaccharide-producing bacterium. Front. Microbiol. 2018, 9, 2041. [Google Scholar] [CrossRef]
- Tajabadi, N.; Mardan, M.; Saari, N.; Mustafa, S.; Bahreini, R.; Manap, M.Y.A. Identification of Lactobacillus plantarum, Lactobacillus pentosus and Lactobillus fermentumfrom honey stomach of honeybee. Braz. J. Microbiol. 2013, 44, 717–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humblot, C.; Guyot, J.P. Pyrosequencing of tagged 16S rRNA gene amplicons for rapid deciphering of the microbiomes of fermented foods such as pearl millet slurries. Appl. Environ. Microbiol. 2009, 75, 4354–4361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, X. Bioinformatics and the Cell: Modern Computational Approaches in Genomics, Proteomics and Transcriptomics, 2nd ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; pp. 1–489. [Google Scholar] [CrossRef]
- Wasfi, R.; Ola, A.A.; Mai, M.Z.; Hossam, M.A. Probiotic Lactobacillus sp. inhibit growth, biofilm formation and gene expression of caries inducing Streptococcus mutans. J. Cell. Mol. Med. 2018, 22, 1972–1983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.M.; Lee, N.K.; Paik, H.D. Antibacterial and anticavity activity of probiotic Lactobacillus plantarum 200661 isolated from fermented foods against Streptococcus mutans. LWT 2020, 118, 108840. [Google Scholar] [CrossRef]
- Ispirli, H.; Dertli, E. Isolation and characterisation of lactic acid bacteria from traditional koumiss and kurut. Int. J. Food Prop. 2017, 20, S2441–S2449. [Google Scholar] [CrossRef] [Green Version]
- Bancalari, E.; D’Incecco, P.; Sardaro, M.L.S.; Neviani, E.; Pellegrino, L.; Gatti, M. Impedance microbiology to speed up the screening of lactic acid bacteria exopolysaccharide production. Int. J. Food Microbiol. 2019, 306, 108268. [Google Scholar] [CrossRef]
- Zeidan, A.A.; Poulsen, V.K.; Janzen, T.; Buldo, P.; Derkx, P.M.; Øregaard, G.; Neves, A.R. Polysaccharide production by lactic acid bacteria: From genes to industrial applications. Fems. Microbiol. Rev. 2017, 41, 168–200. [Google Scholar] [CrossRef] [Green Version]
- Leo, F.; Hashida, S.; Kumagai, D.; Uchida, K.; Motoshima, H.; Arai, I.; Asakuma, S.; Fukuda, K.; Urashima, T. Studies on a neutral exopolysaccharide of Lactobacillus fermentum TDS030603. J. Appl. Glycosci. 2007, 54, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, K.; Shi, T.; Nagami, K.; Leo, F.; Nakamura, T.; Yasuda, K.; Senda, A.; Motoshima, H.; Urashima, T. Effects of carbohydrate source on physicochemical properties of the exopolysaccharide produced by Lactobacillus fermentum TDS030603 in a chemically defined medium. Carbohydr. Polym. 2010, 79, 1040–1045. [Google Scholar] [CrossRef]
- De Albuquerque, T.M.R.; Garcia, E.F.; de Oliveira Araújo, A.; Magnani, M.; Saarela, M.; de Souza, E.L. In vitro characterization of Lactobacillus strains isolated from fruit processing by-products as potential probiotics. Probiotics Antimicrob. Proteins. 2018, 10, 704–716. [Google Scholar] [CrossRef]
- Notararigo, S.; Nácher-Vázquez, M.; Ibarburu, I.; Werning, M.L.; de Palencia, P.F.; Dueñas, M.T.; Aznar, R.; López, P.; Prieto, A. Comparative analysis of production and purification of homo-and hetero-polysaccharides produced by lactic acid bacteria. Carbohydr. Polym. 2013, 93, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Luyen, T.T.A.; Thuy, D.B.T.; Thi, T.T.V.; Huyen, P.T.T. Study on the biosynthesis and structure characterization of exopolysaccharide from Lactobacillus fermentum MC3. Hue Uni. J. Sci. Nat. Sci. 2018, 127, 13. [Google Scholar] [CrossRef] [Green Version]
- Ale, E.C.; Rojas, M.F.; Reinheimer, J.A.; Binetti, A.G. Lactobacillus fermentum: Could EPS production ability be responsible for functional properties? Food Microbiol. 2020, 90, 103465. [Google Scholar] [CrossRef]
- Polak-Berecka, M.; Waśko, A.D.A.M.; Kubik-Komar, A. Optimization of culture conditions for exopolysaccharide production by a probiotic strain of Lactobacillus rhamnosus E/N. Pol. J. Microbiol. 2014, 63, 253. [Google Scholar] [CrossRef]
- Oleksy-Sobczak, M.; Klewicka, E. Optimization of media composition to maximize the yield of exopolysaccharides production by Lactobacillus rhamnosus strains. Probiotics Antimicrob. Proteins 2020, 12, 774–783. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, Y.; Cao, C.; Zhu, X.; Wang, C.; Wu, R.; Wu, J. Extraction and biological activity of exopolysaccharide produced by Leuconostoc mesenteroides SN-8. Int. J. Biol. Macromol. 2020, 157, 36–44. [Google Scholar] [CrossRef]
- Van der Meulen, R.; Grosu-Tudor, S.; Mozzi, F.; Vaningelgem, F.; Zamfir, M.; de Valdez, G.F.; De Vuyst, L. Screening of lactic acid bacteria isolates from dairy and cereal products for exopolysaccharide production and genes involved. Int. J. Food Microbiol. 2007, 118, 250–258. [Google Scholar] [CrossRef]
- Leroy, F.; De Vuyst, L. Advances in production and simplified methods for recovery and quantification of exopolysaccharides for applications in food and health. J. Dairy Sci. 2016, 99, 3229–3238. [Google Scholar] [CrossRef] [Green Version]
- Padmanabhan, A.; Tong, Y.; Wu, Q.; Zhang, J.; Shah, N.P. Transcriptomic insights into the growth phase- and sugarassociated changes in the exopolysaccharide production of a high eps-producing Streptococcus thermophilus ASCC 1275. Front. Microbiol. 2018, 9, 1919. [Google Scholar] [CrossRef]
- Tukenmez, U.; Aktas, B.; Aslim, B.; Yavuz, S. The relationship between the structural characteristics of lactobacilli-eps and its ability to induce apoptosis in colon cancer cells in vitro. Sci. Rep. 2019, 9, 8268. [Google Scholar] [CrossRef] [Green Version]
- Nachtigall, C.; Surber, G.; Herbi, F.; Wefers, D.; Jaros, D.; Rohm, H. Production and Molecular Structure of Heteropolysaccharides from Two Lactic Acid Bacteria. Carbohydr. Polym. 2020, 236, 116019. [Google Scholar] [CrossRef]
- Imran, M.Y.M.; Reehana, N.; Jayaraj, K.A.; Ahamed, A.A.P.; Dhanasekaran, D.; Thajuddin, N.; Alharbi, N.S.; Muralitharan, G. Statistical optimization of exopolysaccharide production by Lactobacillus plantarum NTMI05 and NTMI20. Int. J. Biol. Macromol. 2016, 93, 731–745. [Google Scholar] [CrossRef]
- Vitlic, A.; Sadiq, S.; Ahmed, H.I.; Ale, E.C.; Binetti, A.G.; Collett, A.; Humpreys, P.N.; Laws, A.P. Isolation and characterization of a high molecular mass β-glucan from Lactobacillus fermentum Lf2 and evaluation of its immunomodulatory activity. Carbohydr. Res. 2019, 476, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Ruas-Madiedo, P.; De Los Reyes-Gavilán, C.G. Invited Review: Methods for the Screening, Isolation, and Characterization of Exopolysaccharides Produced by Lactic Acid Bacteria. J. Dairy Sci. 2005, 88, 843–856. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID Sequence | Accession Number | Putative Strain | TOP Hit Blast N | Food Source | Similarity (%) | e-Value | Range |
|---|---|---|---|---|---|---|---|
| SH1 | MW897961 | L. fermentum | L. fermentum 222 | Boza (A3) | 100% | 0.0 | 1–1458 |
| SH2 | MW897962 | L. fermentum | L. fermentum 222 | Fermented cider, fridge (B6) | 99.8% | 0.0 | 1–1458 |
| SH3 | MW897963 | L. fermentum | L. fermentum 222 | Fermented cider, ambient (C9) | 100% | 0.0 | 1–1458 |
| SH4 | MW897983 | L. mesenteroides | L. mesenteroides A16–17 | Unfermented cider (D6) | 100% | 0.0 | 1–1447 |
| Species | Gp ID | Replicate No. | LAB 16S rRNA Gene | gtf Gene | Fold Change in Expression Levels of Genes | |
|---|---|---|---|---|---|---|
| Ct | Ct | Fold Change | ||||
| L. rhamnosus LGG | Control | 1 | 19.46 | 20.51 | 1.000 | 1 ± 0.000 d |
| 2 | 19.40 | 20.49 | ||||
| 3 | 19.27 | 20.38 | ||||
| Mean | 19.38 | 20.46 | ||||
| L. fermentum SH1 | A3 | 1 | 19.17 | 17.22 | 8.1681 | 8.6139 ± 0.320 c |
| 2 | 20.28 | 18.24 | 8.6939 | |||
| 3 | 20.39 | 18.30 | 9.0005 | |||
| Mean | 19.95 | 17.92 | 8.6139 | |||
| L. fermentum SH2 | B6 | 1 | 20.72 | 17.91 | 14.8254 | 14.7230 ± 0.070 a |
| 2 | 19.92 | 17.09 | 15.0324 | |||
| 3 | 21.13 | 18.37 | 14.3204 | |||
| Mean | 20.59 | 17.79 | 14.7230 | |||
| L. fermentum SH3 | C9 | 1 | 20.22 | 18.00 | 9.8492 | 10.1730 ± 0.231 b |
| 2 | 21.66 | 19.37 | 10.3388 | |||
| 3 | 21.90 | 19.61 | 10.3388 | |||
| Mean | 21.26 | 18.99 | 10.1730 | |||
| L. mesenteroides SH4 | D6 | 1 | 21.05 | 22.26 | 0.9138 | 0.8566 ± 0.040 d |
| 2 | 21.31 | 22.63 | 0.8467 | |||
| 3 | 20.74 | 22.12 | 0.8123 | |||
| Mean | 21.03 | 22.34 | 0.8566 | |||
| Type of LAB | EPS mg/100 mL | Cell Mass mg/100 mL | pH | Total Carbohydrate % | Total Protein % | β-Glucans % |
|---|---|---|---|---|---|---|
| LGG | 199.7 ± 3.51 c | 33.33 ± 1.53 c | 4.1 ± 0.6 b | 9.61 ± 0.05 e | 180.92 ± 0.03 b | 0.31 ± 0.01 c |
| A3 | 199.5 ± 1.80 c | 225 ± 5.0 a | 5.0 ± 0.1 a | 99.17 ± 0.11 c | 98.31 ± 0.02 d | 0.12 ± 0.008 e |
| B6 | 481 ± 1.00 a | 116.33 ± 1.53 b | 4.2 ± 0.3 b | 301.48 ± 0.20 a | 150.60 ± 0.02 c | 5.56 ± 0.01 a |
| C9 | 440 ± 2.00 b | 41.7 ± 1.53 c | 5.0 ± 0.5 a | 74.77 ± 0.03 d | 328.00 ± 2.00 a | 4.46 ± 0.01 b |
| D6 | 161 ± 2.65 d | 12.33 ± 1.53 d | 5.1 ± 0.2a | 103.72 ± 0.28 b | 50.24 ± 0.04 e | 0.25 ± 0.008 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Allaith, S.A.; Abdel-aziz, M.E.; Thabit, Z.A.; Altemimi, A.B.; Abd El-Ghany, K.; Giuffrè, A.M.; Al-Manhel, A.J.A.; Ebrahim, H.S.; Mohamed, R.M.; Abedelmaksoud, T.G. Screening and Molecular Identification of Lactic Acid Bacteria Producing β-Glucan in Boza and Cider. Fermentation 2022, 8, 350. https://doi.org/10.3390/fermentation8080350
Allaith SA, Abdel-aziz ME, Thabit ZA, Altemimi AB, Abd El-Ghany K, Giuffrè AM, Al-Manhel AJA, Ebrahim HS, Mohamed RM, Abedelmaksoud TG. Screening and Molecular Identification of Lactic Acid Bacteria Producing β-Glucan in Boza and Cider. Fermentation. 2022; 8(8):350. https://doi.org/10.3390/fermentation8080350
Chicago/Turabian StyleAllaith, Shatha A., Mohamed E. Abdel-aziz, Zaid Akram Thabit, Ammar B. Altemimi, Khalid Abd El-Ghany, Angelo Maria Giuffrè, Alaa Jabbar Abd Al-Manhel, Hanan S. Ebrahim, Reda Mahgoub Mohamed, and Tarek Gamal Abedelmaksoud. 2022. "Screening and Molecular Identification of Lactic Acid Bacteria Producing β-Glucan in Boza and Cider" Fermentation 8, no. 8: 350. https://doi.org/10.3390/fermentation8080350