
Colaconema formosanum, Sarcodia suae, and Nostoc commune as Fermentation Substrates for Bioactive Substance Production

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Algae and Bacterial Strain
2.2. Fermentation of the Three Types of Algae
2.3. Two-Stage Fermentation
2.4. Determination of Algae Moisture Content
2.5. Determination of Phycobiliproteins
total wet weight of algae]
2.6. Chlorophyll a Analysis
2.7. Estimation of the Amount of Reducing Sugar
2.8. Purification and Determination of Polysaccharide Composition
2.9. Determination of Saccharide Components
2.10. Statistical Analysis
3. Results
3.1. Determination of Moisture Content
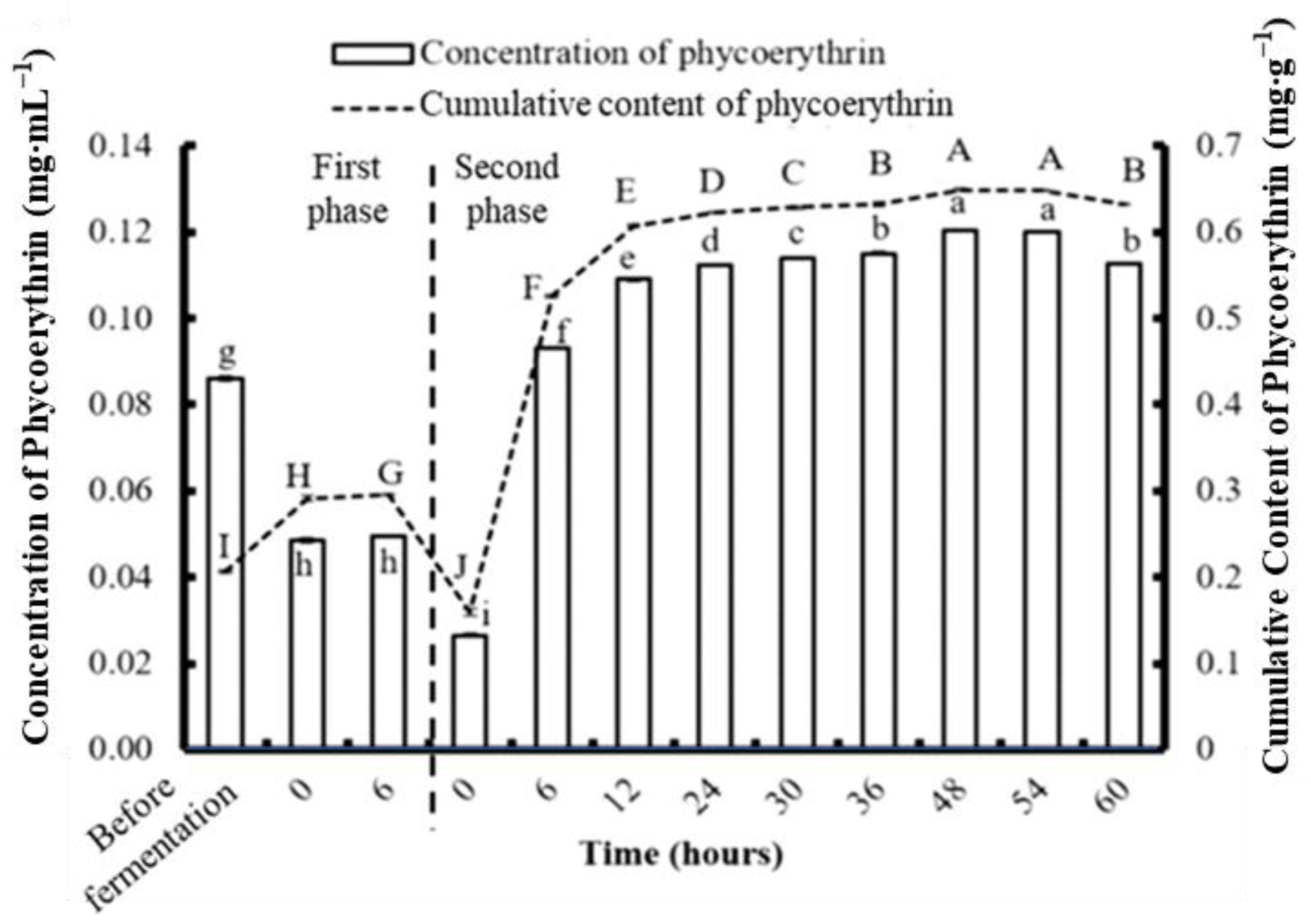
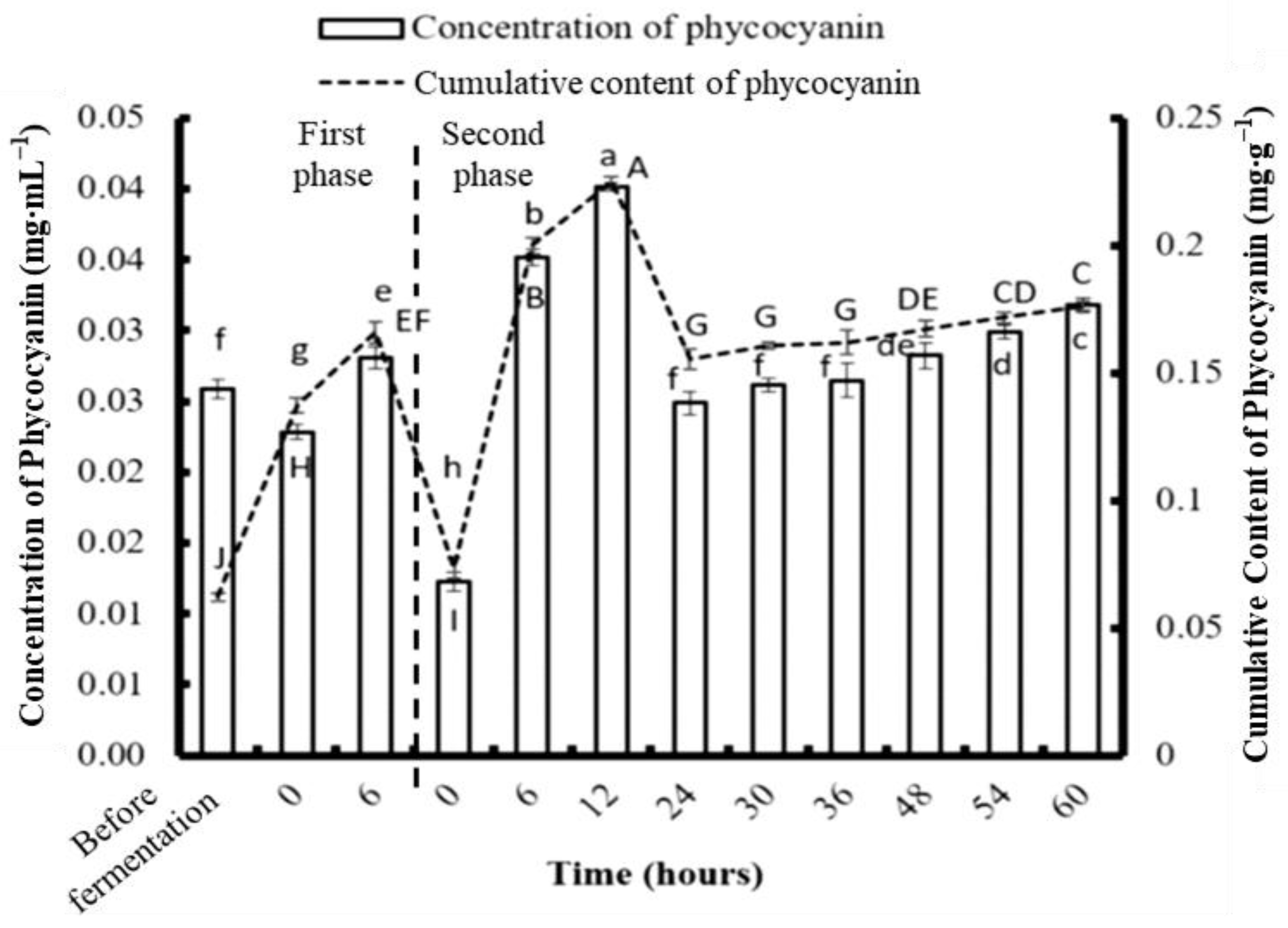
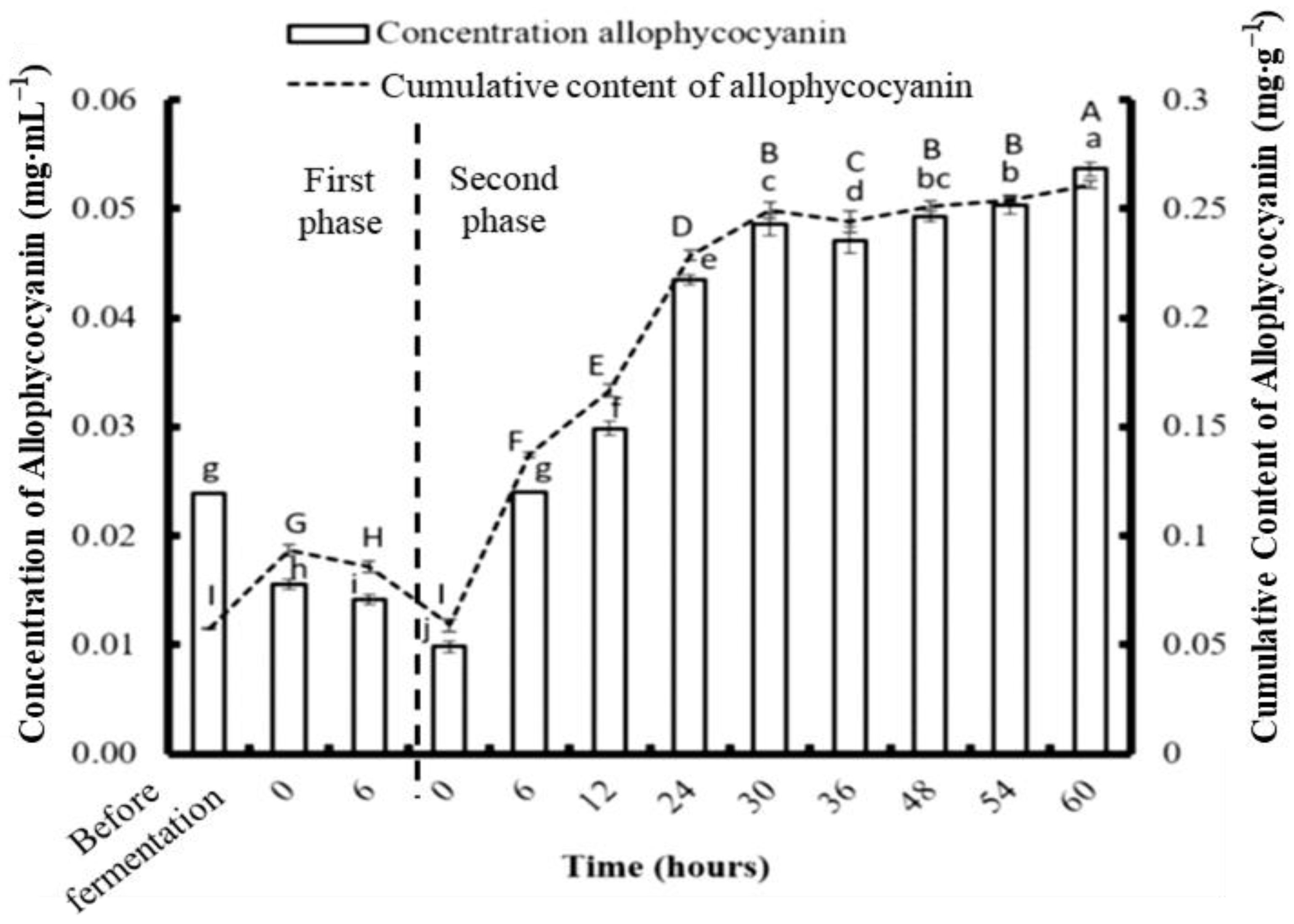
3.2. Concentrations of Phycobiliproteins and Chlorophyll a Obtained from the Three Types of Algae
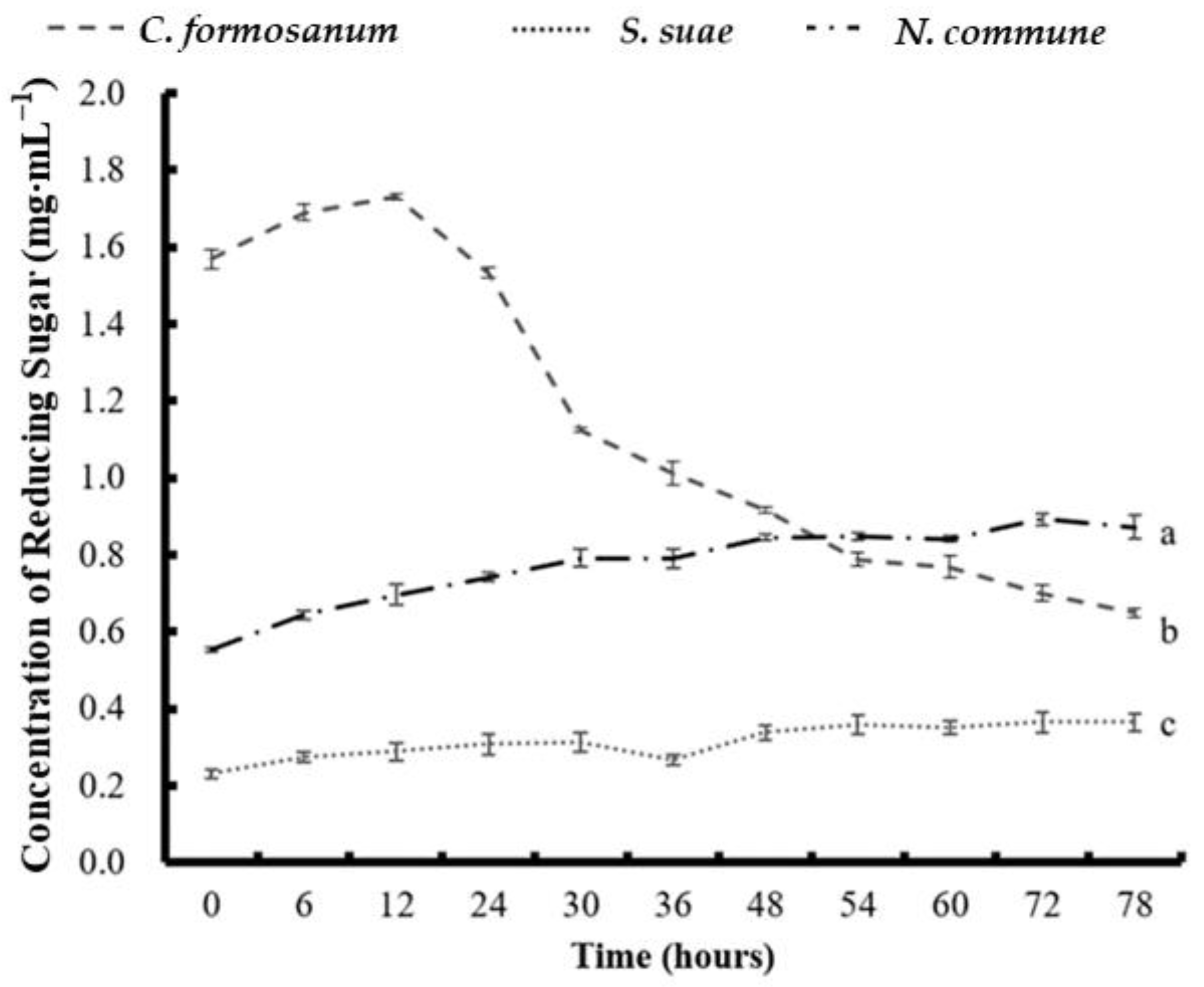
3.3. Determination of the Reducing Sugar Concentration and Monosaccharide Composition of the Three Types of Algae
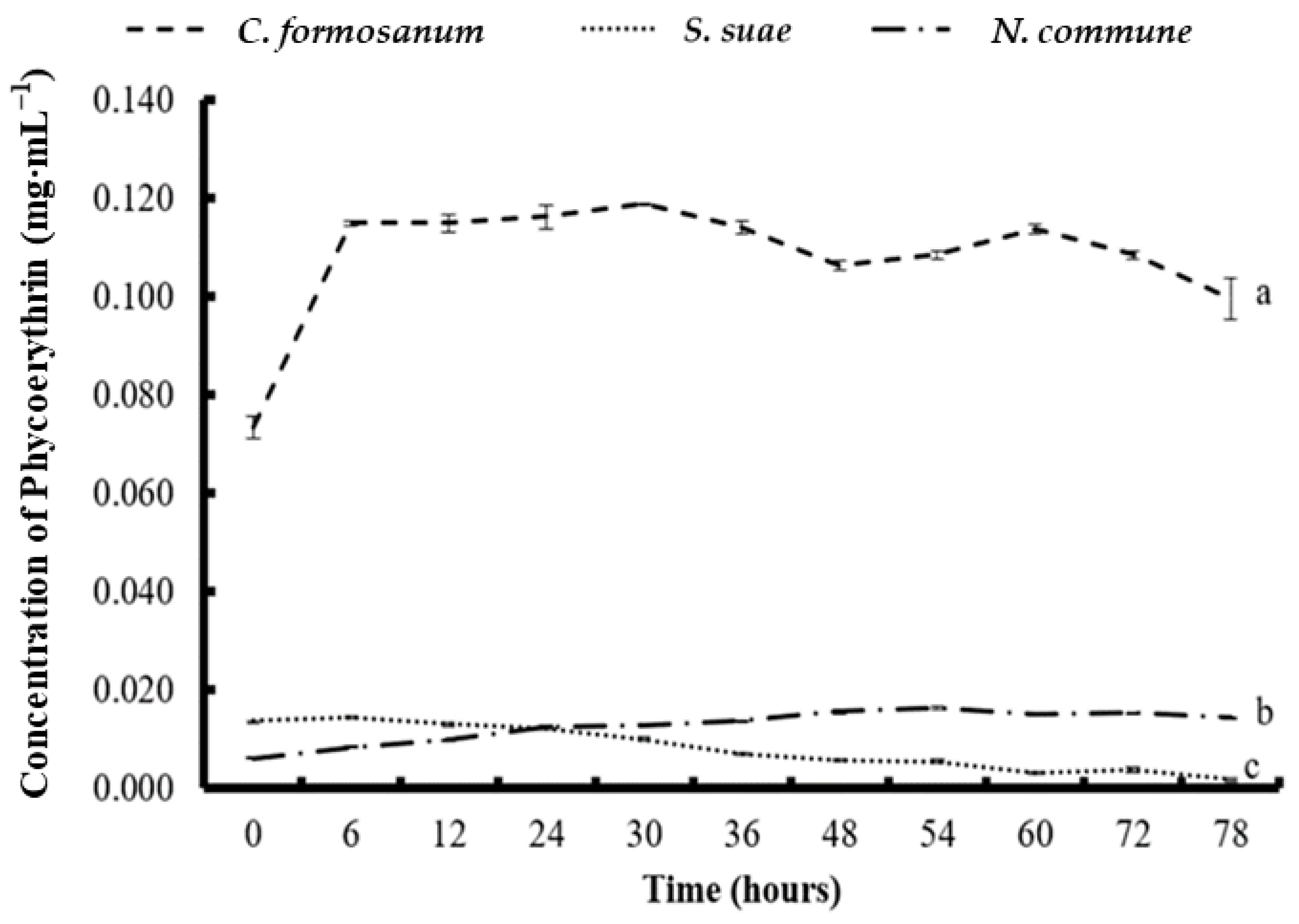
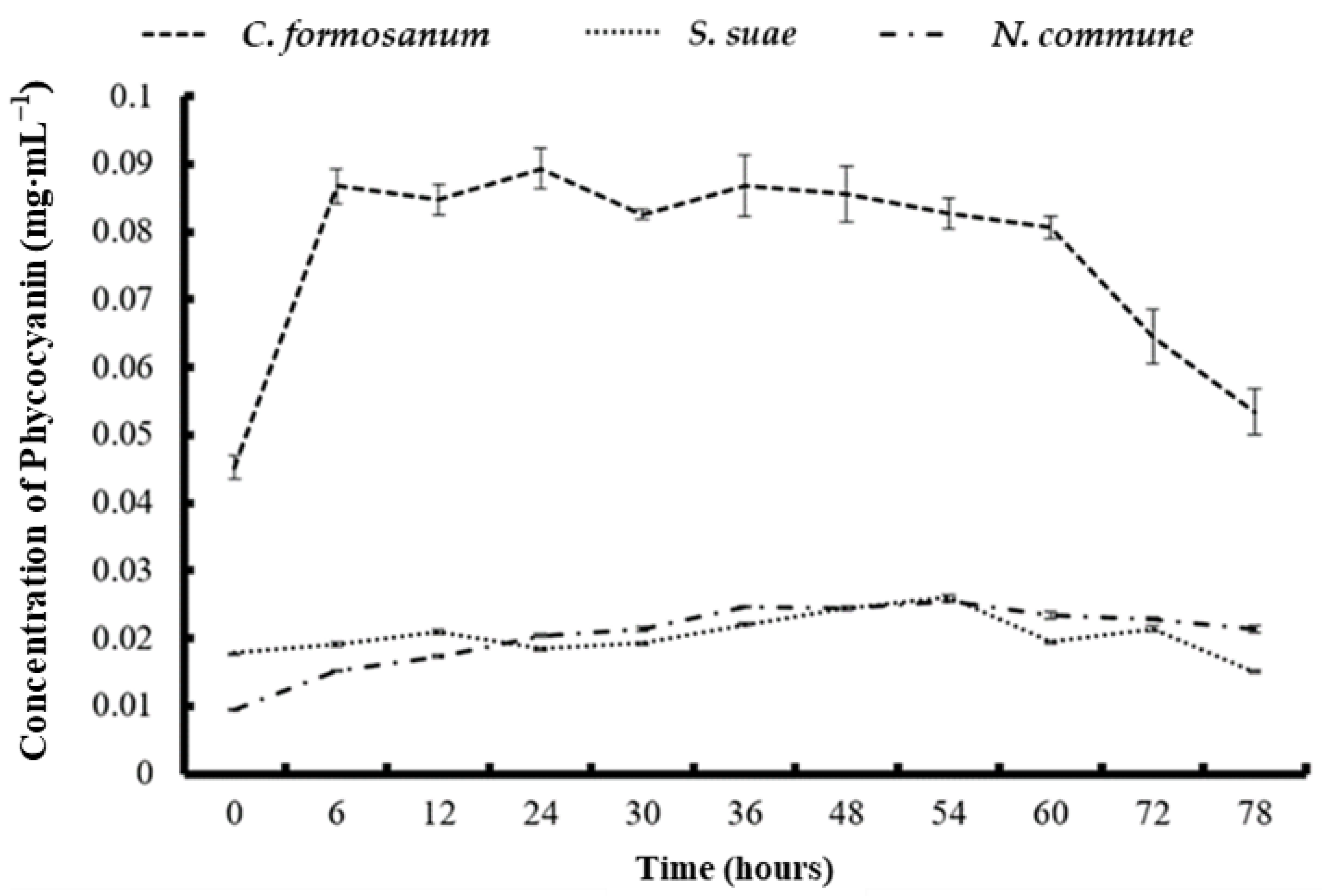
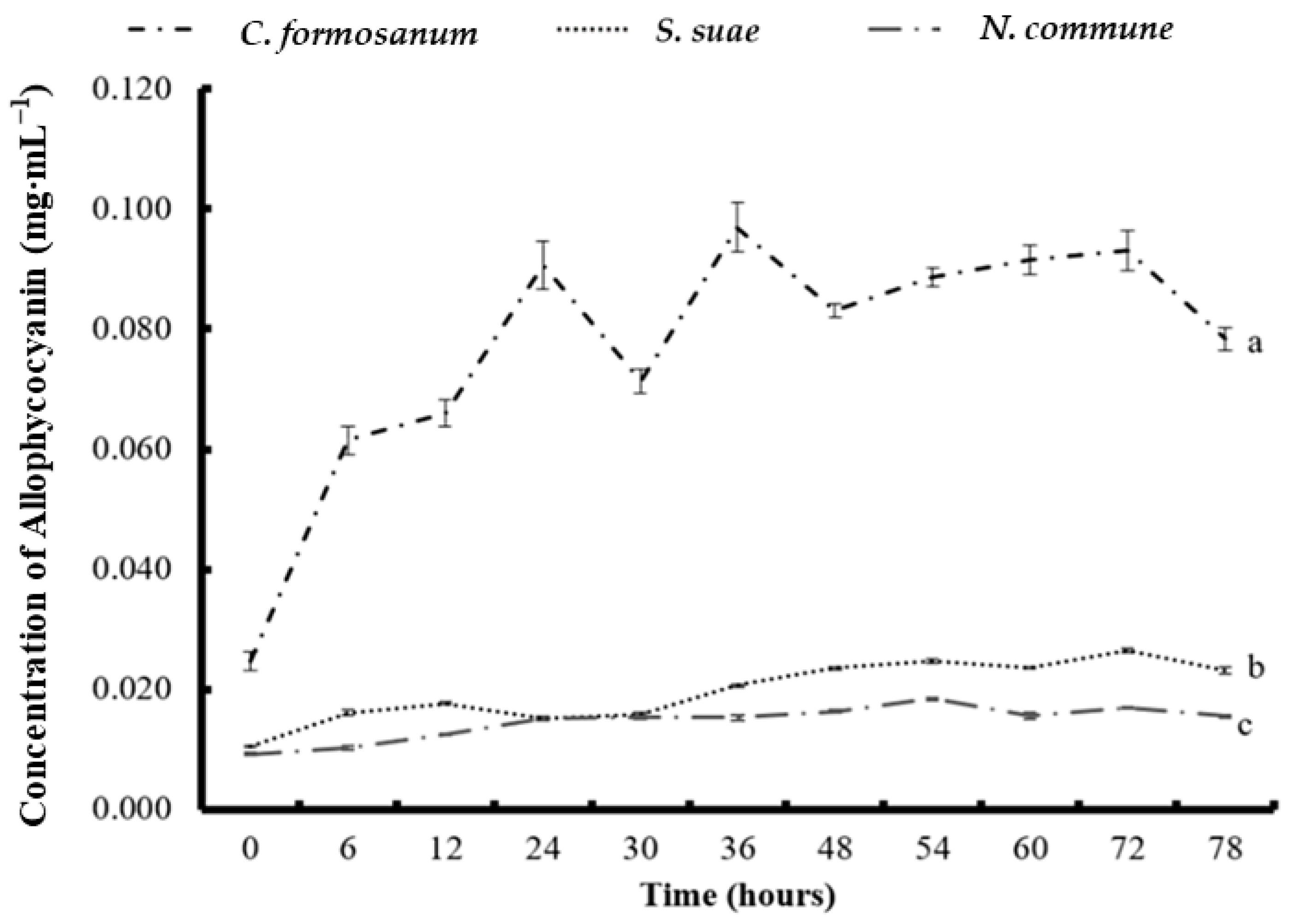
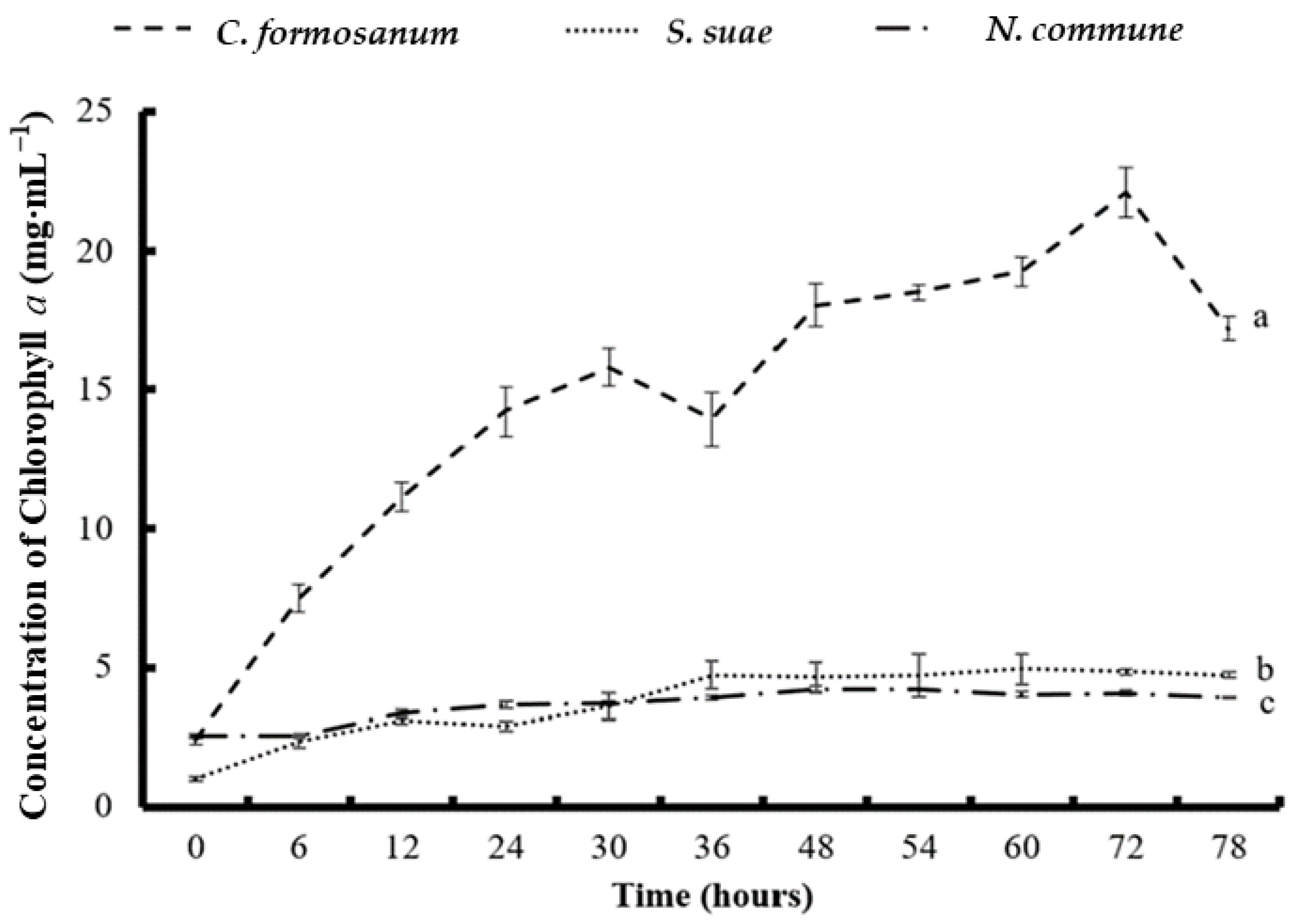
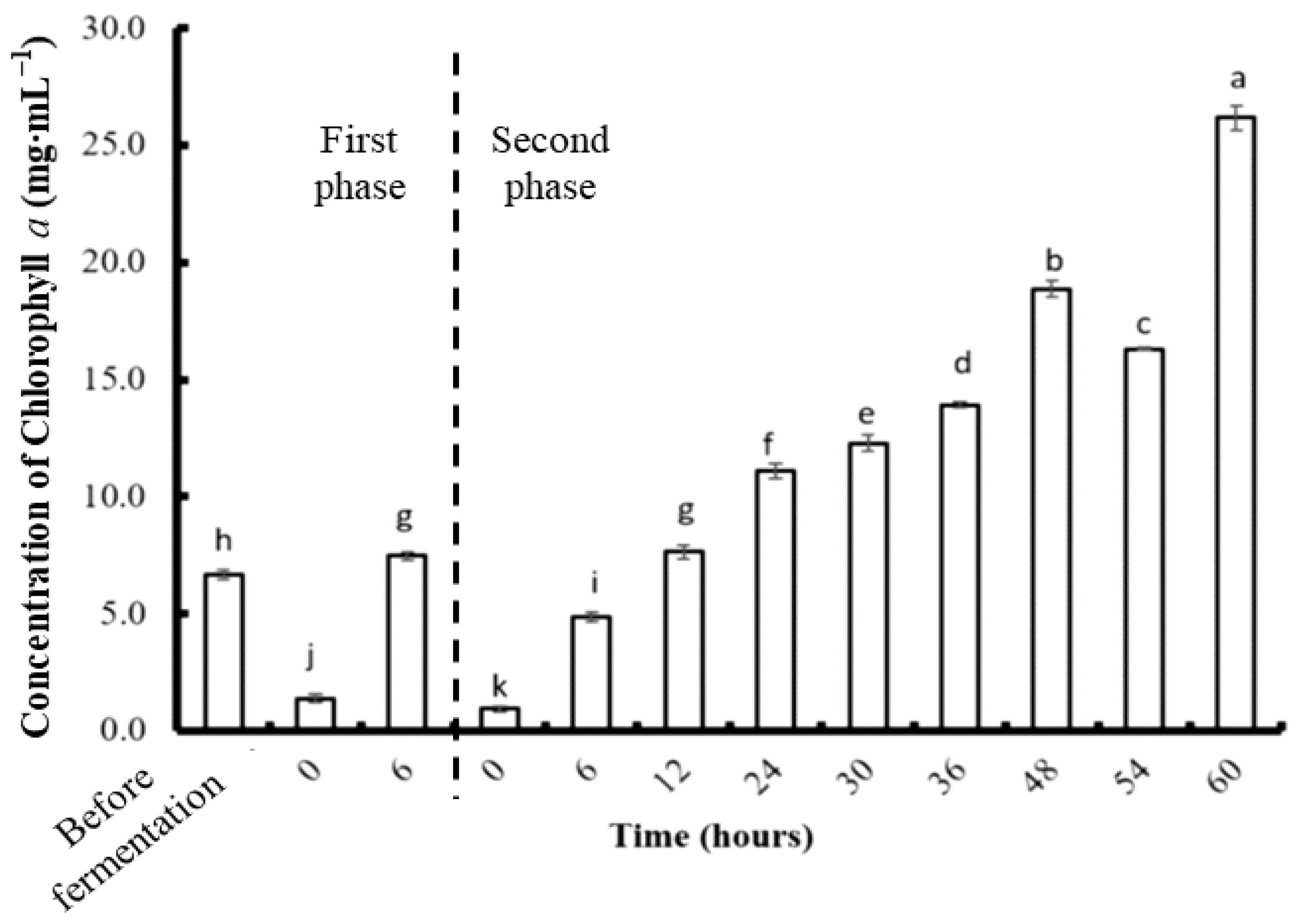
3.4. Phycobiliprotein and Chlorophyll a Concentration after Two-Stage Fermentation
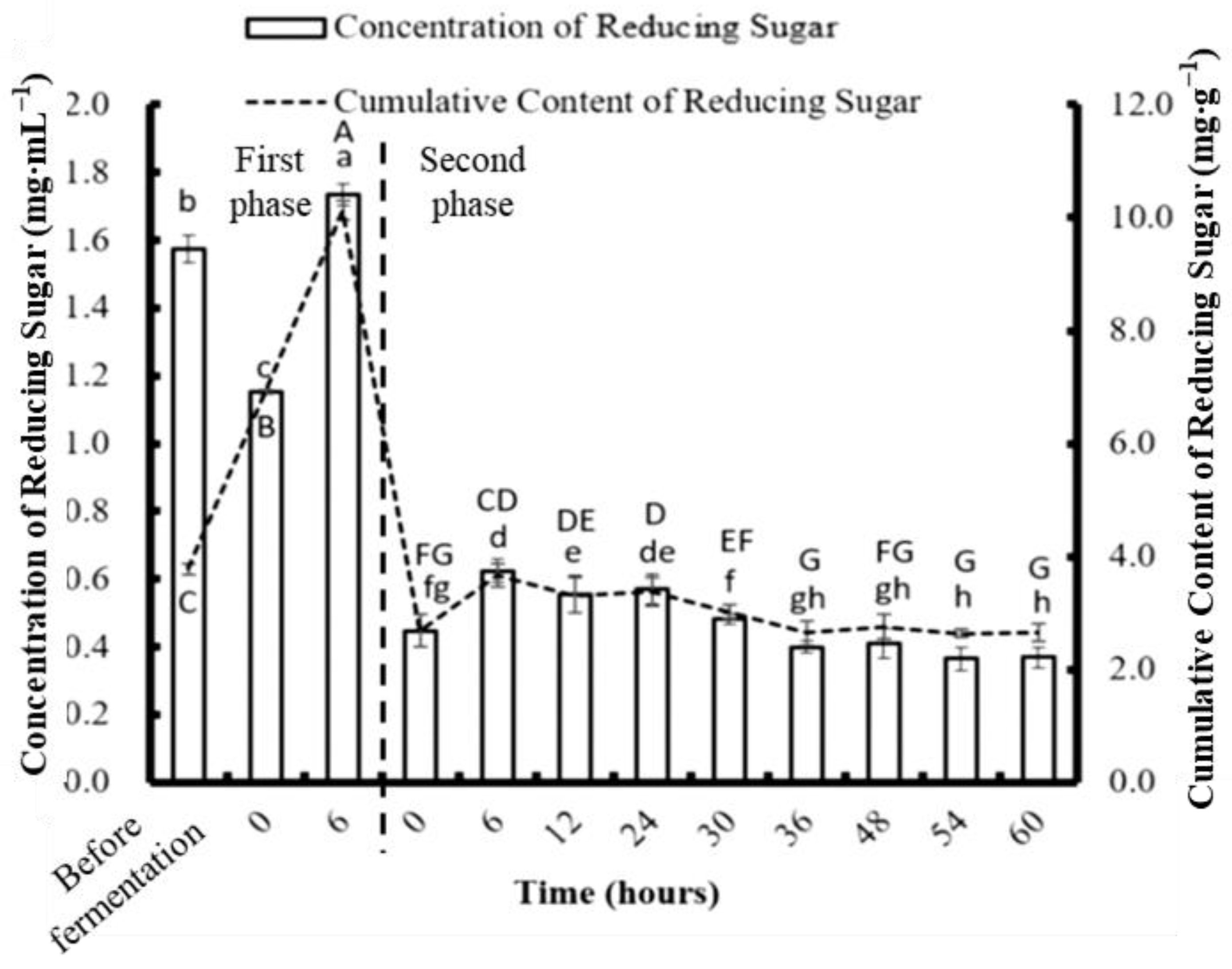
3.5. Determination of Reducing Sugar Concentration Obtained from Two-Stage Fermentation
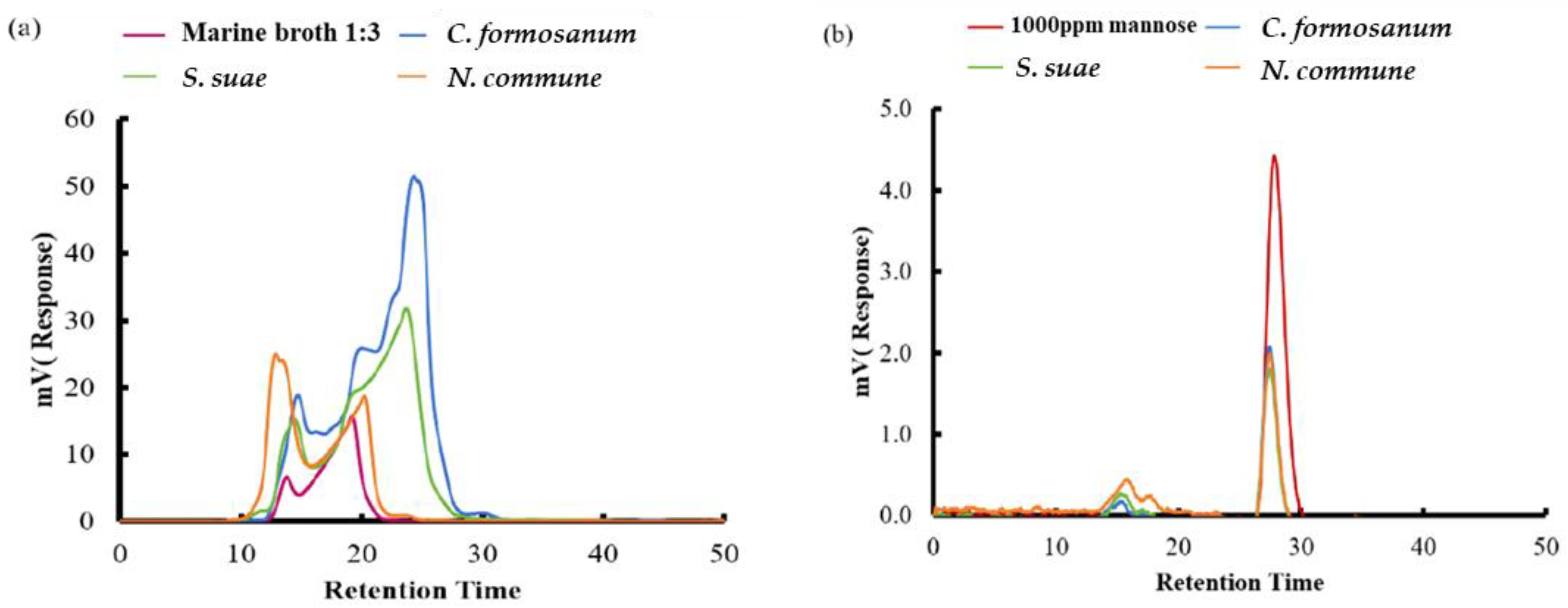
3.6. Determination of Saccharide Composition Obtained from Fermentation with P. haloplanktis ATCC 14393
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| S. suae | Sarcodia suae |
| N. commune | Nostoc commune |
| P. haloplanktis, ATCC 14393 | Pseudoalteromonas haloplanktis ATCC 14393 |
| PE | phycoerythrin |
| PC | phycocyanin |
| APC | allophycocyanin |
| Chl a | Chlorophyll a |
| NMR | nuclear magnetic resonance |
Appendix A
References
- Ganesan, A.R.; Tiwari, U.; Rajauria, G. Seaweed nutraceuticals and their therapeutic role in disease prevention. Food Sci. Hum. Wellness 2019, 8, 252–263. [Google Scholar] [CrossRef]
- Domozych, D.S.; Ciancia, M.; Fangel, J.U.; Mikkelsen, M.D.; Ulvskov, P.; Willats, W.G. The Cell Walls of Green Algae: A Journey through Evolution and Diversity. Front. Plant Sci. 2012, 3, 82. [Google Scholar] [CrossRef] [Green Version]
- Synytsya, A.; Čopíková, J.; Kim, W.-J.; Park, Y.I. Cell Wall Polysaccharides of Marine Algae. In Springer Handbook of Marine Biotechnology; Kim, S.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar] [CrossRef]
- Deniaud-Bouët, E.; Hardouin, K.; Potin, P.; Kloareg, B.; Hervé, C. A review about brown algal cell walls and fucose-containing sulfated polysaccharides: Cell wall context, biomedical properties and key research challenges. Carbohydr. Polym. 2017, 175, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Terauchi, M.; Nagasato, C.; Inoue, A.; Ito, T.; Motomura, T. Distribution of alginate and cellulose and regulatory role of calcium in the cell wall of the brown alga Ectocarpus siliculosus (Ectocarpales, Phaeophyceae). Planta 2016, 244, 361–377. [Google Scholar] [CrossRef] [PubMed]
- Ferrell, J. Sarisky-Reed, Valerie. National Algal Biofuels Technology Roadmap; EERE Publication and Product Library: Washington, DC, USA, 2010. [Google Scholar] [CrossRef] [Green Version]
- Sawant, S.; Salunke, B.; Tran, K.; Kim, B.S. Lignocellulosic and marine biomass as resource for production of polyhydroxyalkanoates. Korean J. Chem. Eng. 2016, 33, 1505–1513. [Google Scholar] [CrossRef]
- Phwan, C.K.; Ong, H.C.; Chen, W.-H.; Ling, T.C.; Ng, E.P.; Show, P.L. Overview: Comparison of pretreatment technologies and fermentation processes of bioethanol from microalgae. Energy Convers. Manag. 2018, 173, 81–94. [Google Scholar] [CrossRef]
- Wilmes, B.; Hartung, A.; Lalk, M.; Liebeke, M.; Schweder, T.; Neubauer, P. Fed-batch process for the psychrotolerant marine bacterium Pseudoalteromonas haloplanktis. Microb. Cell Fact. 2010, 9, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.D.; Kim, J.Y.; Park, J.K.; Lee, C.G. Selective control of the Prorocentrum minimum harmful algal blooms by a novel algal-lytic bacterium Pseudoalteromonas haloplanktis AFMB-008041. Mar. Biotechnol. 2009, 11, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Sekar, S.; Chandramohan, M. Phycobiliproteins as a commodity: Trends in applied research, patents and commercialization. J. Appl. Phycol. 2008, 20, 113–136. [Google Scholar] [CrossRef]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Limantara, L.; Dettling, M.; Indrawati, R.; Indriatmoko Brotosudarmo, T.H.P. Analysis on the Chlorophyll Content of Commercial Green Leafy Vegetables. Procedia Chem. 2015, 14, 225–231. [Google Scholar] [CrossRef] [Green Version]
- Ghose, T. Measurement of cellulase activities. Pure Appl. Chem. 1987, 59, 257–268. [Google Scholar] [CrossRef]
- Tseng, C.-C.; Yeh, H.-Y.; Liao, Z.-H.; Hung, S.-W.; Chen, B.; Lee, P.-T.; Nan, F.-H.; Shih, W.-L.; Chang, C.-C.; Lee, M.-C. An in vitro study shows the potential of Nostoc commune (Cyanobacteria) polysaccharides extract for wound-healing and anti-allergic use in the cosmetics industry. J. Funct. Foods 2021, 87, 104754. [Google Scholar] [CrossRef]
- Özkan, G.; Ersus Bilek, S. Enzyme-assisted extraction of stabilized chlorophyll from spinach. Food Chem. 2015, 176, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Holmström, C.; Kjelleberg, S. Marine Pseudoalteromonas species are associated with higher organisms and produce biologically active extracellular agents. FEMS Microbiol. Ecol. 1999, 30, 285–293. [Google Scholar] [CrossRef]
- Xu, W.; Fan, C.; Zhang, T.; Jiang, B.; Mu, W. Cloning, Expression, and Characterization of a Novel L-Arabinose Isomerase from the Psychrotolerant Bacterium Pseudoalteromonas haloplanktis. Mol. Biotechnol. 2016, 58, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Mittal, R.; Raghavarao, K.S.M.S. Extraction of R-Phycoerythrin from marine macro-algae, Gelidium pusillum, employing consortia of enzymes. Algal Res. 2018, 34, 1–11. [Google Scholar] [CrossRef]
- Le Guillard, C.; Bergé, J.-P.; Donnay-Moreno, C.; Bruzac, S.; Ragon, J.-Y.; Baron, R.; Fleurence, J.; Dumay, J. Soft liquefaction of the red seaweed Grateloupia turuturu Yamada by ultrasound-assisted enzymatic hydrolysis process. J. Appl. Phycol. 2016, 28, 2575–2585. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, Q.A.; Cho, E.; Trinh, L.T.P.; Jeong, J.S.; Bae, H.J. Development of an integrated process to produce d-mannose and bioethanol from coffee residue waste. Bioresour. Technol. 2017, 244, 1039–1048. [Google Scholar] [CrossRef] [PubMed]
- Tario, J.D.; Wallace, P.K. Reagents and Cell Staining for Immunophenotyping by Flow Cytometry. In Pathobiology of Human Diseas; McManus, L.M., Mitchell, R.N., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 3678–3701. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Sugar % (w·w−1) | Mannose % (w·w−1) | Glucose % (w·w−1) | Galactose % (w·w−1) | Xylose % (w·w−1) | Others % (w·w−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Before | After | Before | After | Before | After | Before | After | Before | After | Before | After | |
| C. formosanum | 70.7 | 47.2 | 11.7 | 18.8 | 24.5 | - | 35.6 | 17.1 | 28.2 | 64.1 | - | - |
| S. suae | 68.9 | 53.8 | - | 37.5 | - | - | 91 | 39.2 | 9.0 | 23.3 | - | - |
| N. commune | 42.2 | 81.9 | - | 6.5 | 73.2 | 64.2 | 20.9 | 17.5 | 3.9 | - | 2.0 | 11.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.-C.; Huang, C.-Y.; Lai, C.-L.; Yeh, H.-Y.; Huang, J.; Lung, W.Q.C.; Lee, P.-T.; Nan, F.-H. Colaconema formosanum, Sarcodia suae, and Nostoc commune as Fermentation Substrates for Bioactive Substance Production. Fermentation 2022, 8, 343. https://doi.org/10.3390/fermentation8070343
Lee M-C, Huang C-Y, Lai C-L, Yeh H-Y, Huang J, Lung WQC, Lee P-T, Nan F-H. Colaconema formosanum, Sarcodia suae, and Nostoc commune as Fermentation Substrates for Bioactive Substance Production. Fermentation. 2022; 8(7):343. https://doi.org/10.3390/fermentation8070343
Chicago/Turabian StyleLee, Meng-Chou, Chin-Yi Huang, Chin-Ling Lai, Han-Yang Yeh, Jing Huang, Wei Qing Chloe Lung, Po-Tsang Lee, and Fan-Hua Nan. 2022. "Colaconema formosanum, Sarcodia suae, and Nostoc commune as Fermentation Substrates for Bioactive Substance Production" Fermentation 8, no. 7: 343. https://doi.org/10.3390/fermentation8070343