
Biodegradation of Methylene Blue Using a Novel Lignin Peroxidase Enzyme Producing Bacteria, Named Bacillus sp. React3, as a Promising Candidate for Dye-Contaminated Wastewater Treatment



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation Process of Functional Bacteria
2.2. Screening of MB-Detoxifying Bacteria
2.3. Characterization and Identification of Isolated Bacteria
2.4. Growth Curve of the Bacillus sp. React3 Isolate
2.5. Optimization of Lignin Peroxidase Enzyme Activity
2.6. Removal Test
2.6.1. Effect of the Temperature and pH
2.6.2. Effect of the Methylene Blue Concentration
2.6.3. Effect of the Inoculum Concentration
2.6.4. Effect of Agitation
2.6.5. Effect of the Carbon and Nitrogen Sources
2.7. Enzymes Activity Assays
2.8. MB Decolorization Assay
3. Results
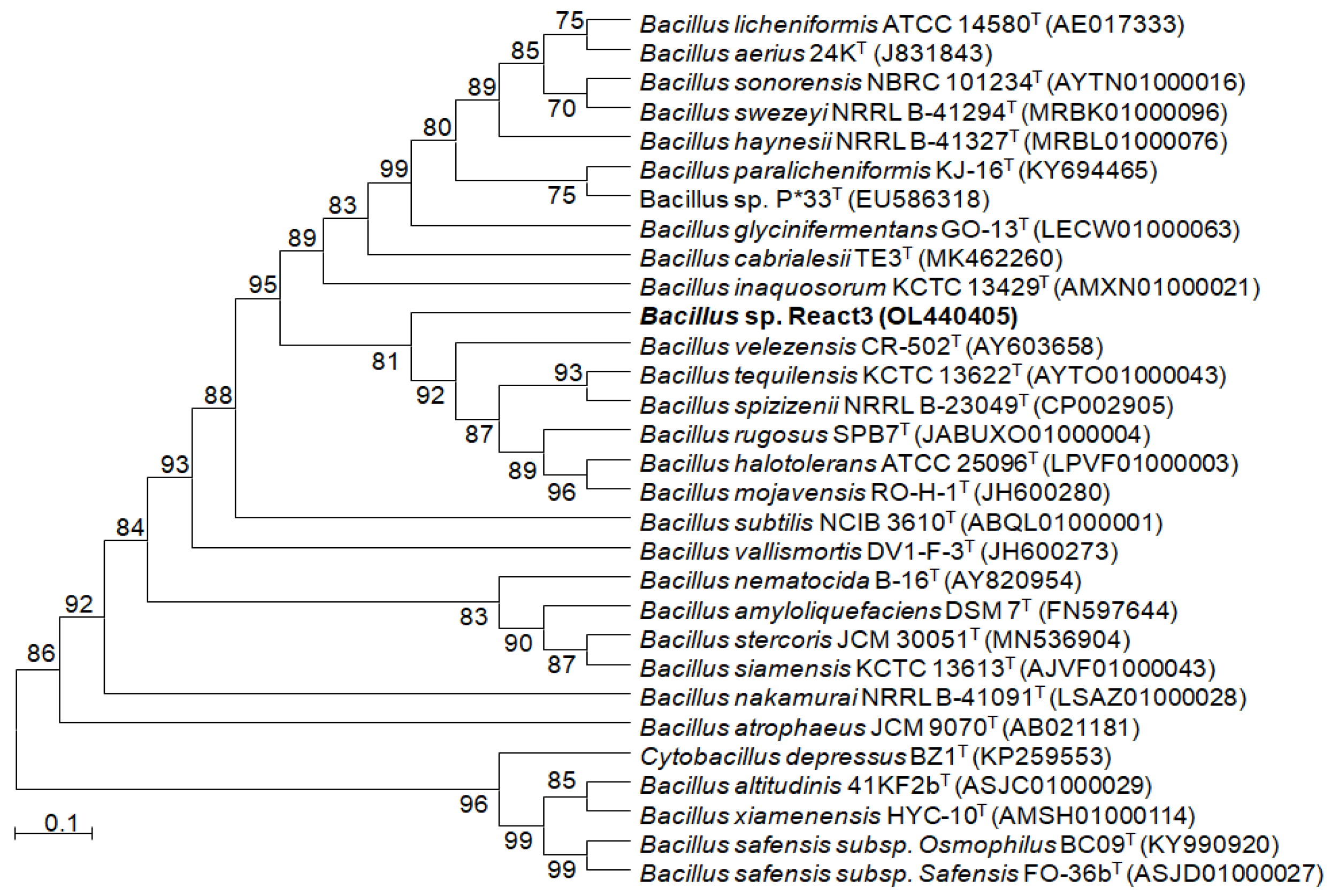
3.1. Characteristic and Phylogenetic Identification of Bacterium
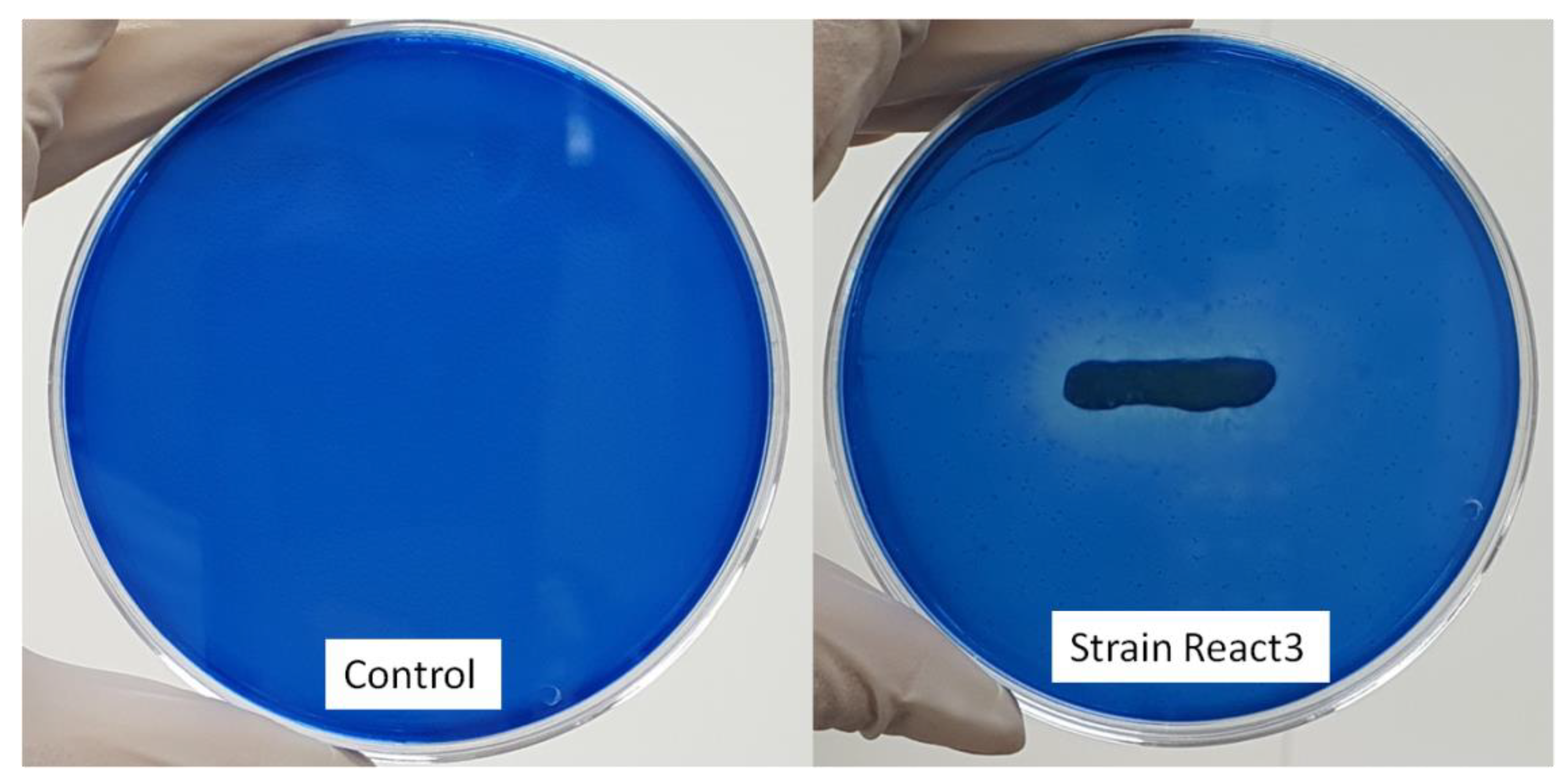
3.2. Potential LiP Enzyme Production
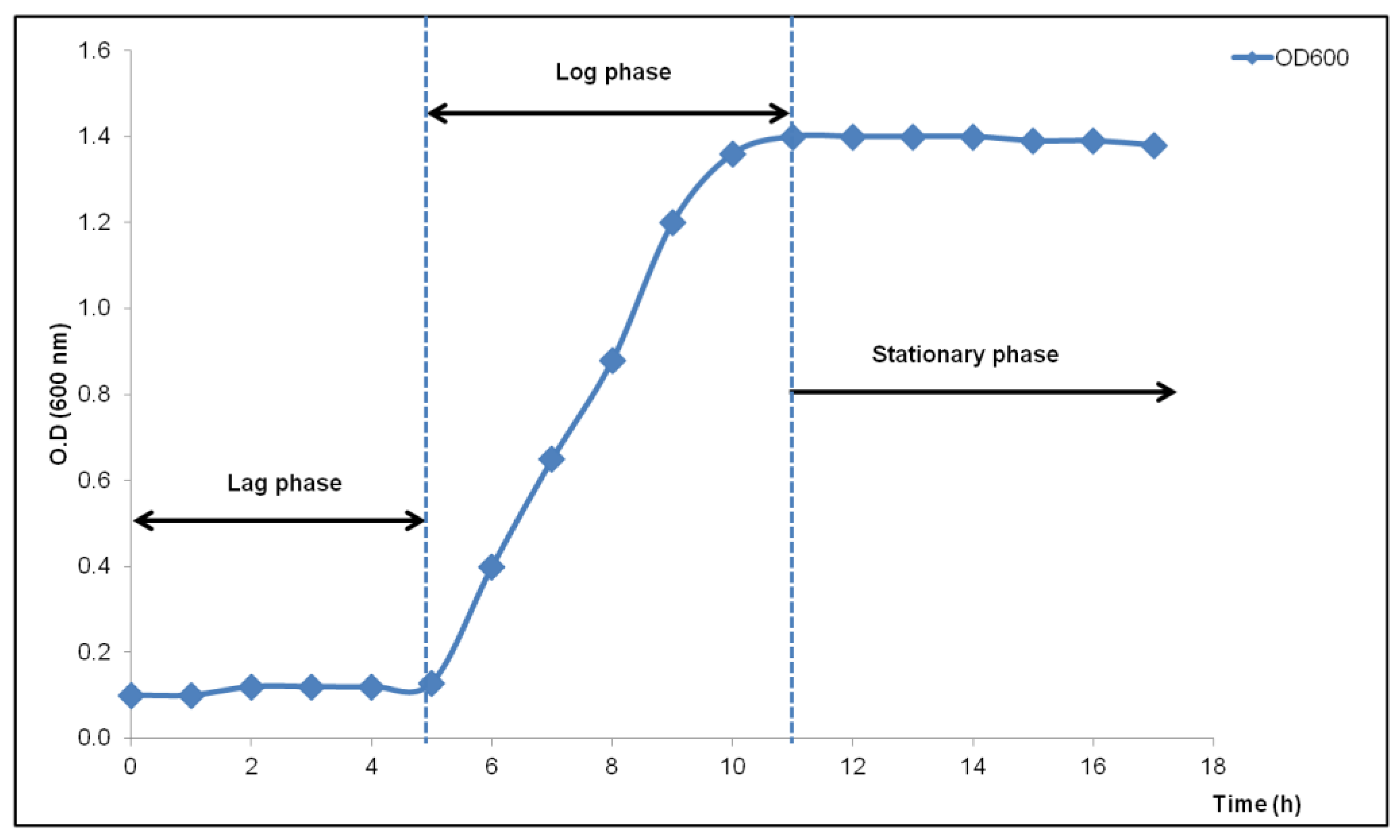
3.3. Optimal Conditions for Growth and Enzyme Production of B. React3
3.4. Effect of Different Factors on Decolorization of MB Dye
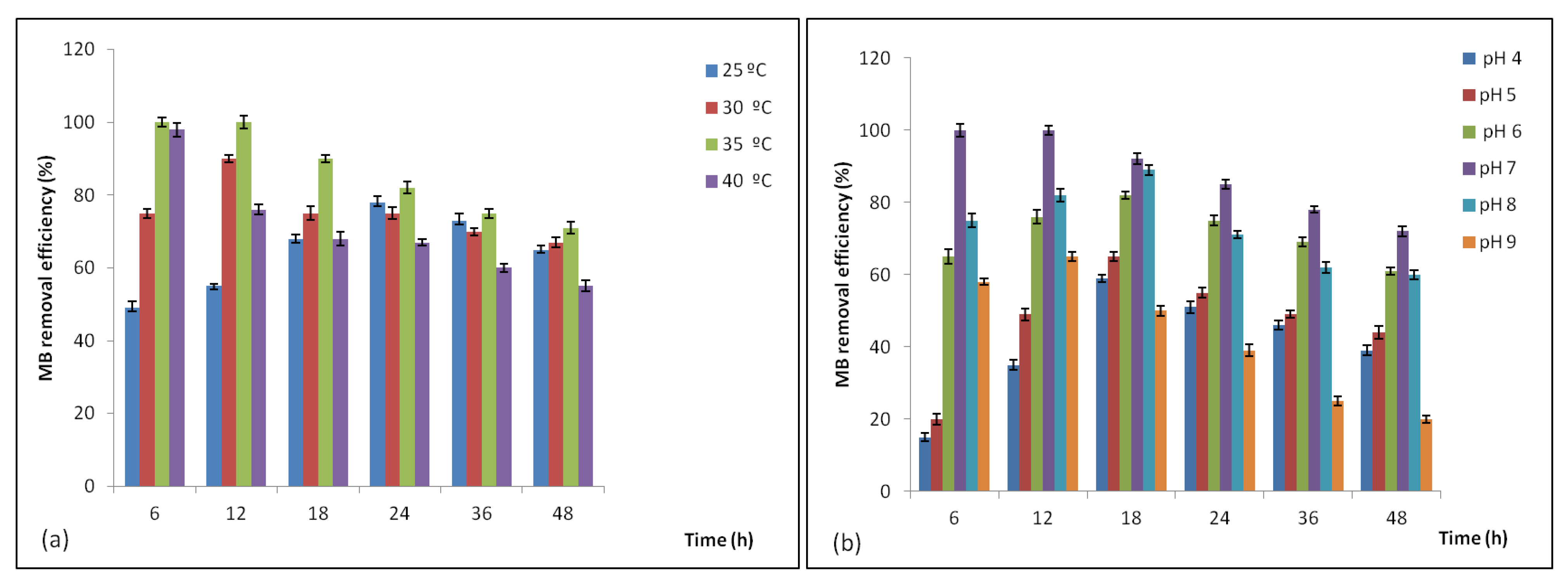
3.4.1. Effect of Temperature and pH
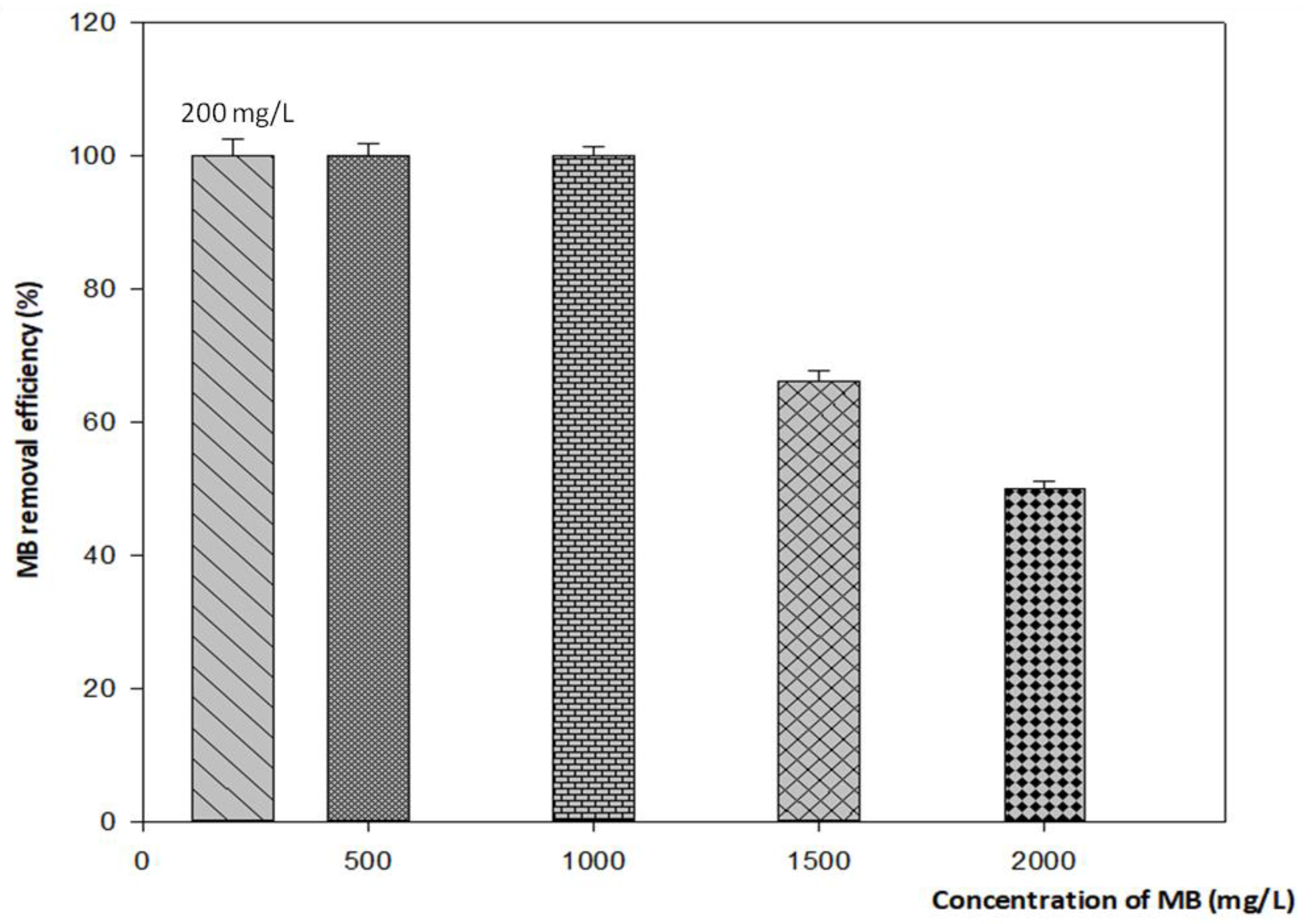
3.4.2. Effect of Initial MB Concentration
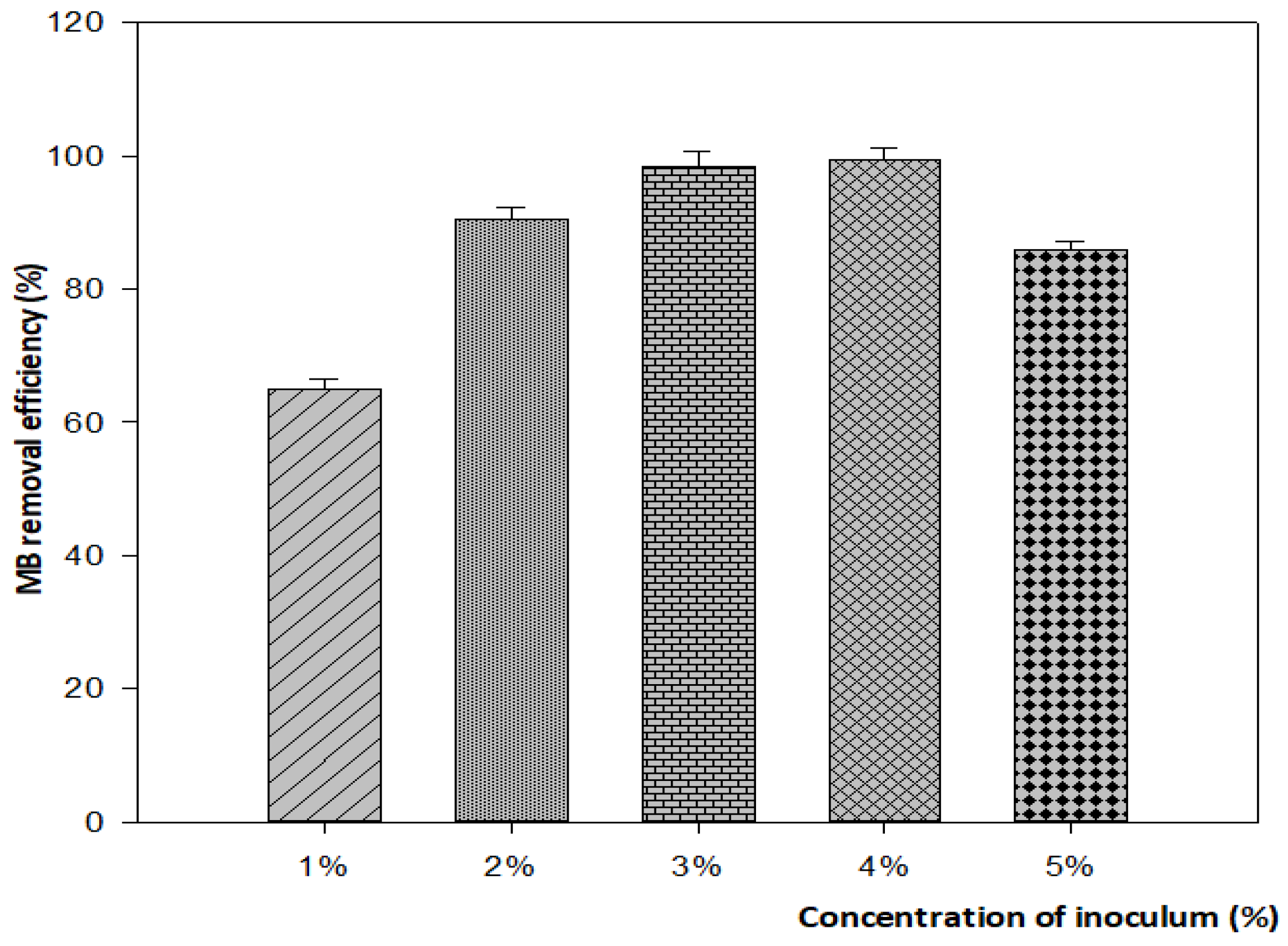
3.4.3. Effect of Inoculum Size
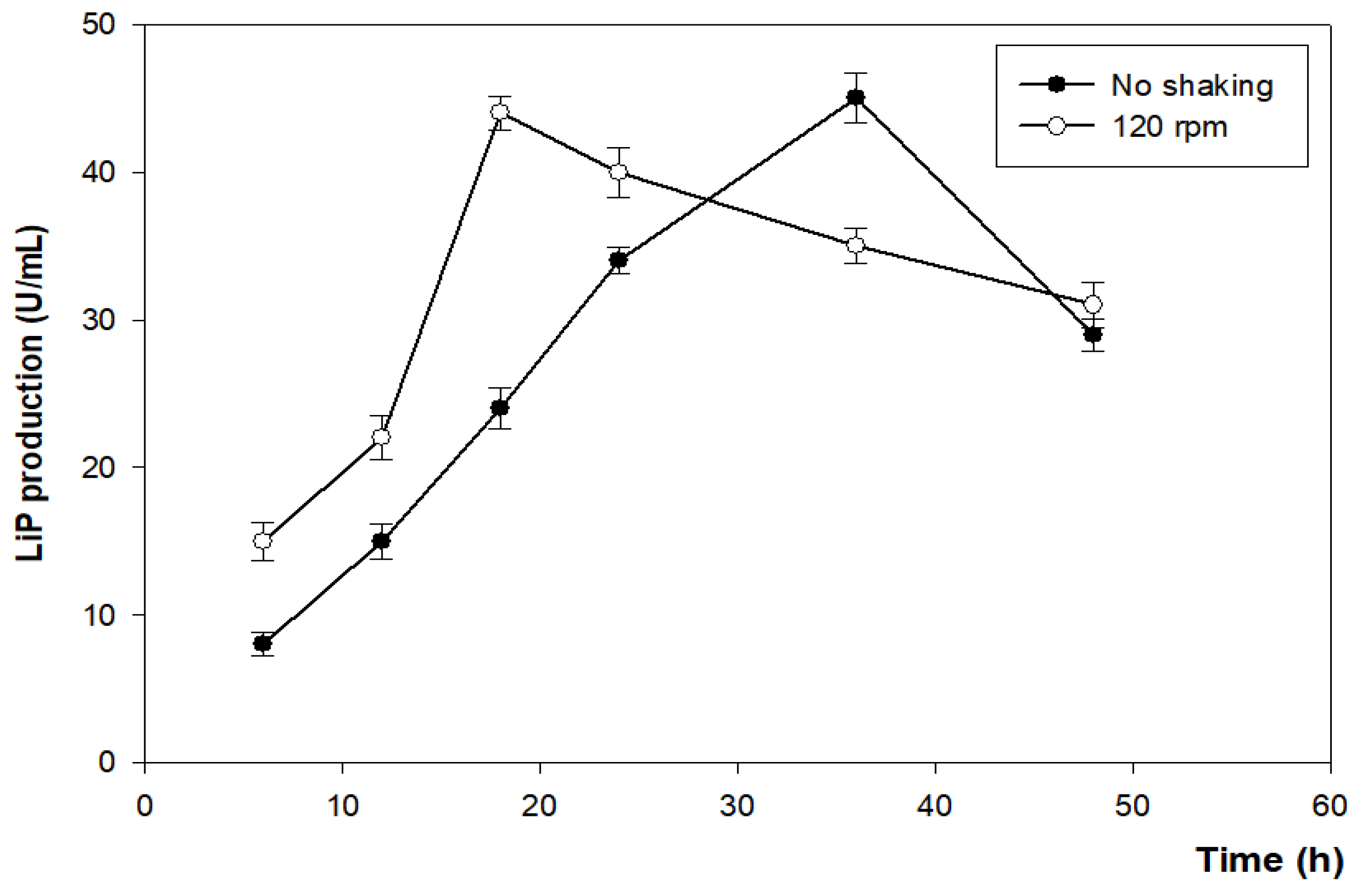
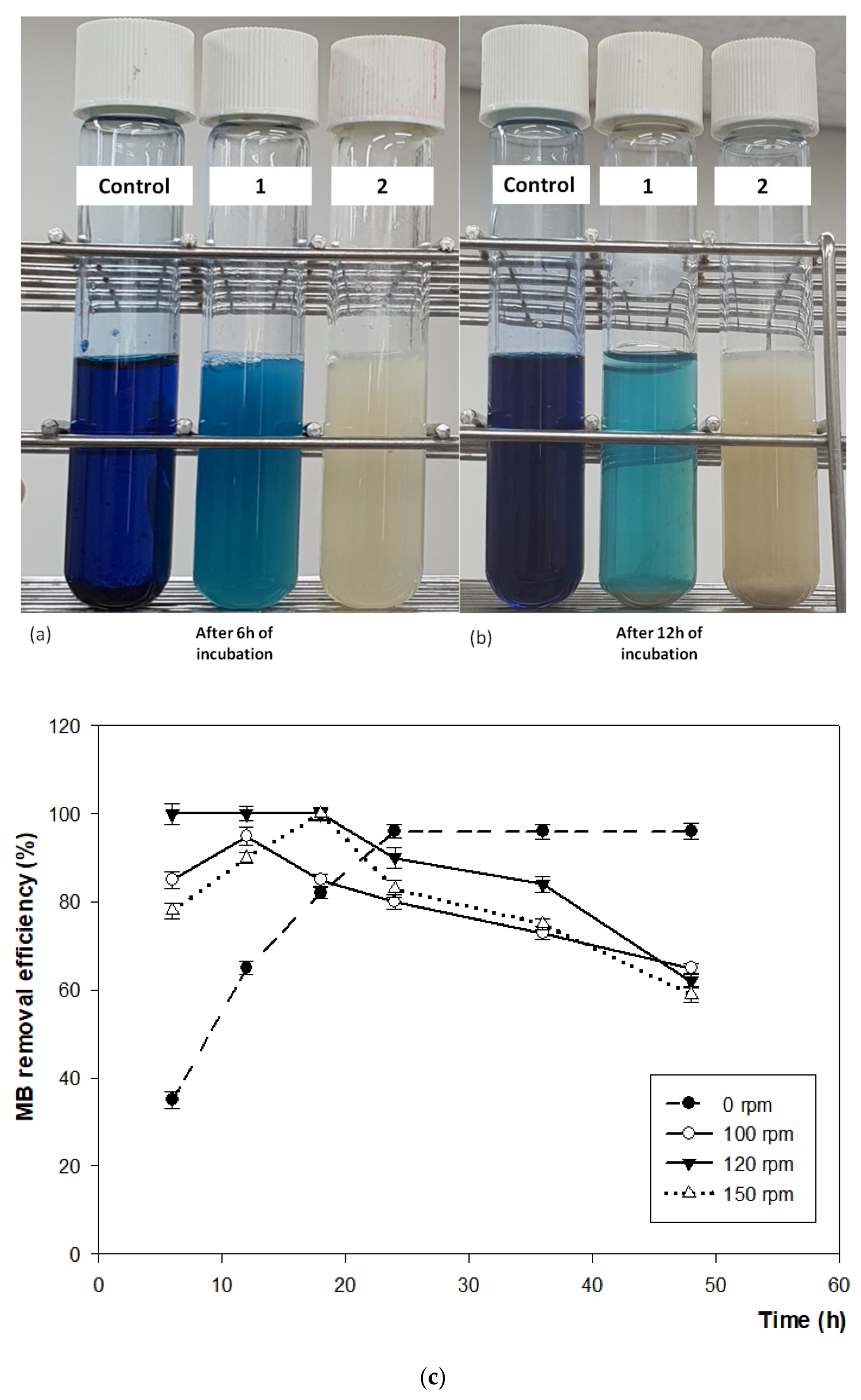
3.4.4. Effect of Agitation
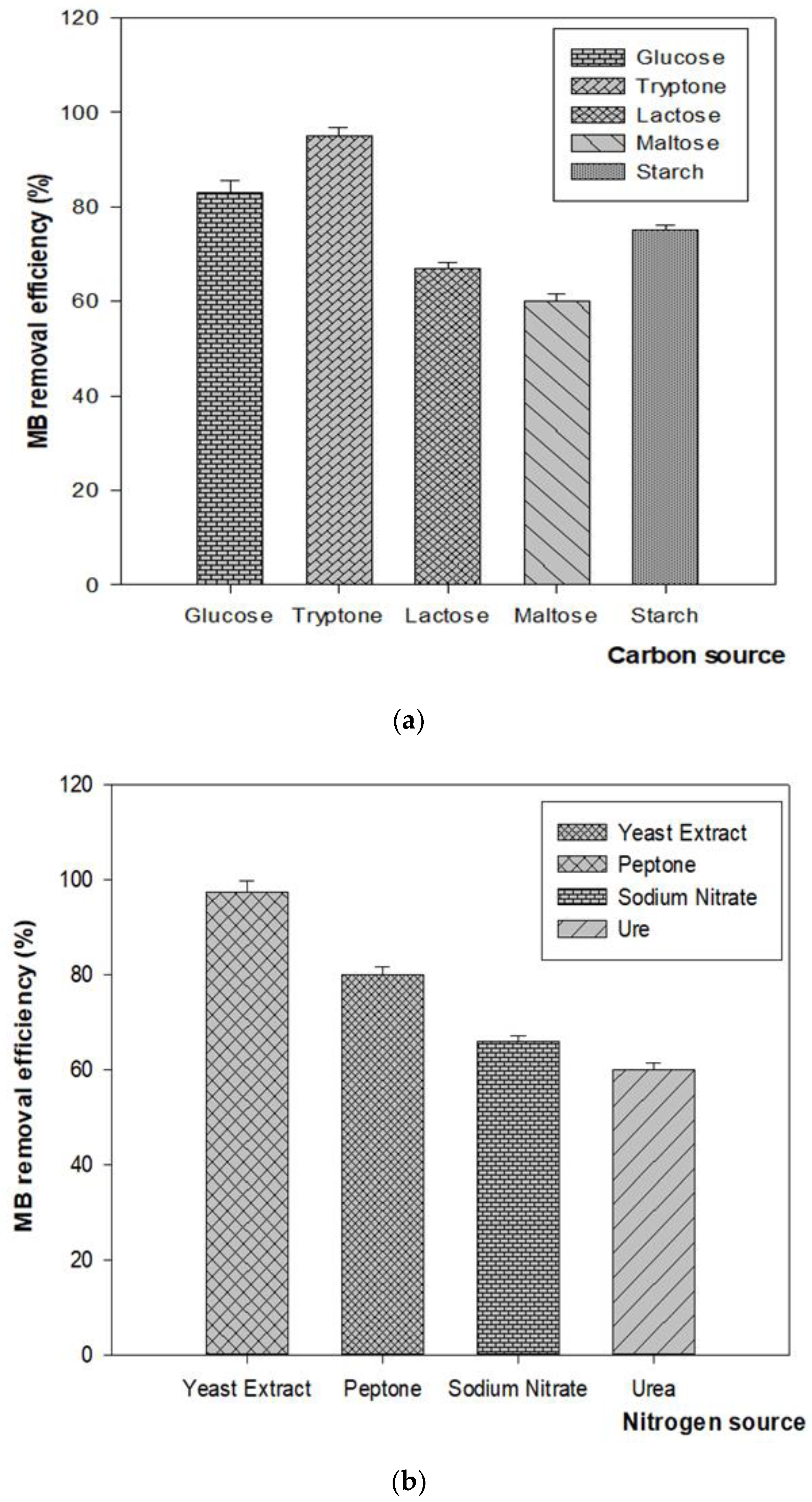
3.4.5. Effect of Different Carbon and Nitrogen Sources
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Benkhaya, B.; Harfi, S.E.; Harfi, A.E. Classifications, Properties and Applications of Textile Dyes: A Review. Appl. J. Environ. Eng. Sci. 2017, 3, 311–320. [Google Scholar]
- Benkhaya, S.; Mrabet, S.; El Harfi, A. A review on classifications, recent synthesis and applications of textile dyes. Inorg. Chem. Commun. 2020, 115, 107891. [Google Scholar] [CrossRef]
- Kishor, R.; Purchase, D.; Saratale, G.D.; Saratale, R.G.; Ferreira, L.F.R.; Bilal, M.; Chandra, R.; Bharagava, R.N. Ecotoxicological and health concerns of persistent coloring pollutants of textile industry wastewater and treatment approaches for environmental safety. J. Environ. Chem. Eng. 2021, 9, 105012. [Google Scholar] [CrossRef]
- Farhan Hanafi, M.; Sapawe, N. A review on the water problem associate with organic pollutants derived from phenol, methyl orange, and remazol brilliant blue dyes. Mater. Today Proc. 2020, 1, A141–A150. [Google Scholar] [CrossRef]
- Selvankumar, T.; Sudhakar, C.; Govarthanan, M. Microbial removal of dye stuffs. In Microbial Biodegradation of Xenobiotic Compounds; CRC Press: Boca Raton, FL, USA, 2019; Volume 95, pp. 95–110. [Google Scholar]
- Das, A.; Mishra, S. Removal of textile dye reactive green-19 using bacterial consortium: Process optimization using response surface methodology and kinetics study. J. Environ. Chem. Eng. 2017, 5, 612–627. [Google Scholar] [CrossRef]
- Bayomie, O.S.; Kandeel, H.; Shoeib, T.; Yang, H.; Youssef, N.; El-Sayed, M.M.H. Novel approach for effective removal of methylene blue dye from water using fava bean peel waste. Sci. Rep. 2020, 10, 7824. [Google Scholar] [CrossRef]
- Lade, H.; Kadam, A.; Paul, D.; Govindwar, S. Biodegradation and detoxification of textile azo dyes by bacterial consortium under sequential microaerophilic/aerobic processes. EXCLI J. 2015, 14, 158. [Google Scholar]
- Xi, Y.; Shen, Y.; Yang, F.; Yang, G.; Liu, C.; Zhang, Z.; Zhu, D. Removal of azo dye from aqueous solution by a new biosorbent prepared with Aspergillus nidulans cultured in tobacco wastewater. J. Taiwan Inst. Chem. Eng. 2013, 44, 815–820. [Google Scholar] [CrossRef]
- Kandisa, R.V.; KV, N.S.; Shaik, K.B.; Gopinath, R. Dye Removal by Adsorption: A Review. J. Bioremed. Biodegrad. 2016, 7, 371. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Chen, Y.; Li, X.; Cheng, Y.; Yang, C.; Zeng, G. Influence of salinity on microorganisms in activated sludge processes: A review. Int. Biodeterior. Biodegrad. 2017, 119, 520–527. [Google Scholar] [CrossRef]
- Pham, V.H.T.; Kim, J.; Chang, S.; Chung, W. Purification and characterization of strong simultaneous enzyme production of protease and amylase from an extremophile-Bacillus sp. FW2 and its possibility in food waste degradation. Fermentation 2022, 8, 12. [Google Scholar] [CrossRef]
- Pham, V.H.T.; Kim, J.; Shim, J.; Chang, S.; Chung, W. Coconut Mesocarp-Based Lignocellulosic Waste as a Substrate for Cellulase Production from High Promising Multienzyme-Producing Bacillus amyloliquefaciens FW2 without Pretreatments. Microorganisms 2022, 10, 327. [Google Scholar] [CrossRef]
- Karim, M.E.; Dhar, K.; Hossain, M.T. Decolorization of textile reactive dyes by bacterial monoculture and consortium screened from textile dyeing effluent. J. Genet. Eng. Biotechnol. 2018, 16, 375–380. [Google Scholar] [CrossRef]
- Bharti, V.; Vikrant, K.; Goswami, M.; Tiwari, H.; Sonwani, R.K.; Lee, J.; Tsang, D.C.W.; Kim, K.H.; Saeed, M.; Kumar, S.; et al. Biodegradation of methylene blue dye in a batch and continuous mode using biochar as packing media. Environ. Res. 2019, 1, 356–364. [Google Scholar] [CrossRef]
- Contreras, M.; Grande-Tovar, C.D.; Vallejo, W.; Chaves-López, C. Bio-removal of Methylene Blue from aqueous solution by Galactomyces geotrichum KL20A. Water 2019, 11, 282. [Google Scholar] [CrossRef] [Green Version]
- Eslami, H.; Khavidak, S.S.; Salehi, F.; Khosravi, R.; Ali Fallahzadeh, R.; Peirovi, R.; Sadeghi, S. Biodegrdation of methylene blue from aqueous solution by bacteria isolated from contaminated soil. J. Adv. Environ. Health Res. 2017, 5, 10–15. [Google Scholar]
- Ogunlaja, A.; Nwankwo, I.N.; Omaliko, M.E.; Olukanni, O.D. Biodegradation of Methylene Blue as an evidence of synthetic dyes mineralization during textile effluent biotreatment by Acinetobacter pittii. Environ. Process. 2020, 7, 931–947. [Google Scholar] [CrossRef]
- Haque, M.M.; Haque, M.A.; Mosharaf, M.K.; Marcus, P.K. Decolorization, degradation and detoxification of carcinogenic sulfonated azo dye methyl orange by newly developed biofilm consortia. Saudi. J. Biol. Sci. 2021, 28, 793–804. [Google Scholar] [CrossRef]
- Michelle, S.; RAN, S.A.; Lucy, J.; Pinontoan, R. Methylene blue decolorizing bacteria isolated from water sewage in Yogyakarta, Indonesia. Biodiversitas 2020, 21, 1136–1141. [Google Scholar] [CrossRef]
- Zhu, C.; Feng, Q.; Ma, H.; Wu, M.; Wang, D.; Wang, Z. Effect of methylene blue on the properties and microbial community of anaerobic granular sludge. BioResources 2018, 13, 6033–6046. [Google Scholar]
- Pham, V.H.T.; Kim, J. Cultivation of unculturable soil bacteria. Trends. Biotechnol. 2012, 30, 475–484. [Google Scholar] [CrossRef]
- Pham, V.H.T.; Kim, J.; Chang, S.; Chung, W. Investigation of lipolytic-secreting bacteria from an artificially polluted soil using a modified culture method and optimization of their lipase production. Microorganism 2021, 9, 2590. [Google Scholar] [CrossRef]
- Pham, V.H.T.; Kim, J.; Chang, S.; Chung, W. Improvement of Hydrogen Production during Anaerobic Fermentation of Food Waste Leachate by Enriched Bacterial Culture Using Biochar as an Additive. Microorganisms 2021, 9, 2438. [Google Scholar] [CrossRef]
- Pham, V.H.T.; Ahn, J.; Kim, J.; Lee, S.; Lee, I.; Kim, S.; Chang, S.; Chung, W. Volatile Fatty Acid Production from Food Waste Leachate Using Enriched Bacterial Culture and Soil Bacteria as Co-Digester. Sustainability 2021, 13, 9606. [Google Scholar] [CrossRef]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Felsenstein, J. Confidence limit on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Buntić, A.V.; Pavlović, M.D.; Antonović, D.G.; Šiler-Marinković, S.S.; Dimitrijević-Branković, S.I. A treatment of wastewater containing basic dyes by the use of new strain Streptomyces microflavus CKS6. J. Clean. Prod. 2017, 148, 347–354. [Google Scholar] [CrossRef]
- Blažeka, D.; Car, J.; Klobučar, N.; Jurov, A.; Zavašnik, J.; Jagodar, A.; Eva Kovačević, E.; Krstulović, N. Photodegradation of methylene blue and rhodamine B using laser-synthesized ZnO nanoparticles. Materials 2020, 13, 4357. [Google Scholar] [CrossRef]
- Ruiz-García, C.; Béjar, V.; Martínez-Checa, F.; Llamas, I.; Quesada, E. Bacillus velezensis sp. nov., a surfactant-producing bacterium isolated from the river Vélez in Málaga, southern Spain. Int. J. Syst. Evol. Microbiol. 2005, 55, 191–195. [Google Scholar] [CrossRef] [Green Version]
- Briggs, T.; Blunn, G.; Hislop, S.; Ramalhete, R.; Bagley, C.; McKenna, D.; Coathup, M. Antimicrobial photodynamic therapy-a promising treatment for prosthetic joint infections. Lasers Med. Sci. 2018, 33, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.F.; Santhanam, N.; Badri, D.V.; Hunter, W.J.; Manter, D.K.; Decker, S.R.; Vivanco, J.M.; Reardon, K.F. Isolation and characterization of lignin-degrading bacteria from rainforest soils. Biotechnol. Bioeng. 2013, 110, 1616–1626. [Google Scholar] [CrossRef]
- Bandounas, L.; Wierckx, N.J.; de Winde, J.H.; Ruijssenaars, H.J. Isolation and characterization of novel bacterial strains exhibiting ligninolytic potential. BMC Biotechnol. 2011, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Ngalimat, M.S.; Yahaya, R.S.R.; Baharudin, M.M.A.-a.; Yaminudin, S.M.; Karim, M.; Ahmad, S.A.; Sabri, S. A review on the biotechnological applications of the operational group Bacillus amyloliquefaciens. Microorganisms 2021, 9, 614. [Google Scholar] [CrossRef]
- Roy, D.C.; Kumar Biswas, S.; Kumar Saha, A.; Sikdar, B.; Rahman, M.; Kumar Roy, A.; Hossain Prodhan, Z.; Seong Tang, S. Biodegradation of Crystal Violet dye by bacteria isolated from textile industry effluents. PeerJ 2018, 6, e5015. [Google Scholar] [CrossRef]
- Pham, V.H.T.; Kim, J.; Chang, S.; Chung, W. Bacterial biosorbents, an efficient heavy metals green clean-up strategy: Prospects, Challenges, and Opportunities. Microorganisms 2022, 10, 610. [Google Scholar] [CrossRef]
- Lalnunhlimi, S.; Krishnaswamy, V. Decolorization of azo dyes (Direct Blue 151 and Direct Red 31) by moderately alkaliphilic bacterial consortium. Braz. J. Microbiol. 2016, 47, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Holey, B.A. Decolourization of Congo Red dye by bacteriaand consortium isolated from dye contaminated soil. Int. Res. J. Sci. Eng. 2015, 3, 107–112. [Google Scholar]
- Wanyonyi, W.C.; Onyari, J.M.; Shiundu, P.M.; Mulaa, F.J. Biodegradation and detoxification of malachite green dye using novel enzymes from Bacillus cereus strain KM201428: Kinetic and metabolite analysis. Energy Procedia 2017, 119, 38–51. [Google Scholar] [CrossRef]
- Kishor, R.; Saratale, G.D.; Saratale, R.G.; Romanholo Ferreira, L.F.; Bilal, M.; Iqbal, H.M.N.; Bharagava, R.N. Efficient degradation and detoxification of methylene blue dye by a newly isolated ligninolytic enzyme producing bacterium Bacillus albus MW407057. Colloids Surf. B Biointerfaces 2021, 206, 111947. [Google Scholar] [CrossRef]
- Bilińska, L.; Blus, K.; Gmurek, M.; Ledakowicz, S. Coupling of electrocoagulation and ozone treatment for textile wastewater reuse. Chem. Eng. J. 2019, 358, 992–1001. [Google Scholar] [CrossRef]
- Aksu, Z.; Tezer, S. Biosorption of reactive dyes on the green alga Chlorella vulgaris. Process Biochem. 2005, 40, 1347–1361. [Google Scholar] [CrossRef]
- Kumar, K.V.; Ramamurthi, V.; Sivanesan, S. Biosorption of malachite green, a cationic dye onto Pithophora sp., a fresh water algae. Dye. Pigment. 2006, 69, 102–107. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, L.; Shang, H.; Li, Q.; Zhou, W. Treatment of Azo Dye wastewater by the Self-Flocculating marine bacterium Aliiglaciecola Lipolytica. Environ. Technol. Innov. 2020, 19, 100810. [Google Scholar] [CrossRef]
- Bharagava, R.N.; Mani, S.; Mulla, S.I.; Saratale, G.D. Degradation and decolorization potential of an ligninolytic enzyme producing Aeromonas hydrophila for crystal violet dye and its phytotoxicity evaluation. Ecotoxicol. Environ. Saf. 2018, 156, 166–175. [Google Scholar] [CrossRef]
- Guadie, A.; Tizazu, S.; Melese, M.; Guo, W.; Ngo, H.H.; Xia, S. Biodecolourization of textile azo dye using Bacillus sp. strain CH12 isolated from alkaline lake. Biotechnol. Rep. 2017, 15, 92–100. [Google Scholar] [CrossRef]
- Bose, R.B.; Thillaichidambaram, M.; Paulraj, B.; Narayanan, K.; Ganesan, N.; Ramakritinan Chokalingam Muthiah, R.C.; Murugesan, R.K. Bio-decolourization of Reactive Blue EFAF using halotolerant. Exiguobacterium profundum strain CMR2 isolated from salt pan. Biocatal. Agric. Biotechnol. 2018, 16, 98–106. [Google Scholar]
- Mabrouk, M.E.M.; Yusef, H.H. Decolorization of fast red by Bacillus subtilis HM. J. Appl. Sci. Res. 2008, 4, 262–269. [Google Scholar]
- Haq, I.; Kumar, S.; Kumari, V.; Singh, S.K.; Raj, A. Evaluation of bioremediation potentiality of ligninolytic Serratia liquefaciens for detoxification of pulp and paper mill effluent. J. Hazard. Mater. 2016, 305, 190–199. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pham, V.H.T.; Kim, J.; Chang, S.; Chung, W. Biodegradation of Methylene Blue Using a Novel Lignin Peroxidase Enzyme Producing Bacteria, Named Bacillus sp. React3, as a Promising Candidate for Dye-Contaminated Wastewater Treatment. Fermentation 2022, 8, 190. https://doi.org/10.3390/fermentation8050190
Pham VHT, Kim J, Chang S, Chung W. Biodegradation of Methylene Blue Using a Novel Lignin Peroxidase Enzyme Producing Bacteria, Named Bacillus sp. React3, as a Promising Candidate for Dye-Contaminated Wastewater Treatment. Fermentation. 2022; 8(5):190. https://doi.org/10.3390/fermentation8050190
Chicago/Turabian StylePham, Van Hong Thi, Jaisoo Kim, Soonwoong Chang, and Woojin Chung. 2022. "Biodegradation of Methylene Blue Using a Novel Lignin Peroxidase Enzyme Producing Bacteria, Named Bacillus sp. React3, as a Promising Candidate for Dye-Contaminated Wastewater Treatment" Fermentation 8, no. 5: 190. https://doi.org/10.3390/fermentation8050190