
Effect of Homo-Fermentative Lactic Acid Bacteria Inoculants on Fermentation Characteristics and Bacterial and Fungal Communities in Alfalfa Silage

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ensiling
2.2. Analytical Procedures
2.3. Enumeration of Microorganisms on Agar Plates
2.4. Analysis of the Composition of the Microbial Communities by Next-Generation Sequencing
2.5. Statistical Analysis
3. Results
3.1. Targeted and Actual LAB Application Rate
3.2. Nutrient Composition and Numbers of Culturable Microorganisms in Fresh Forage
3.3. Nutrient Composition, Fermentation Profile and Numbers of Culturable Microorganisms in Silages
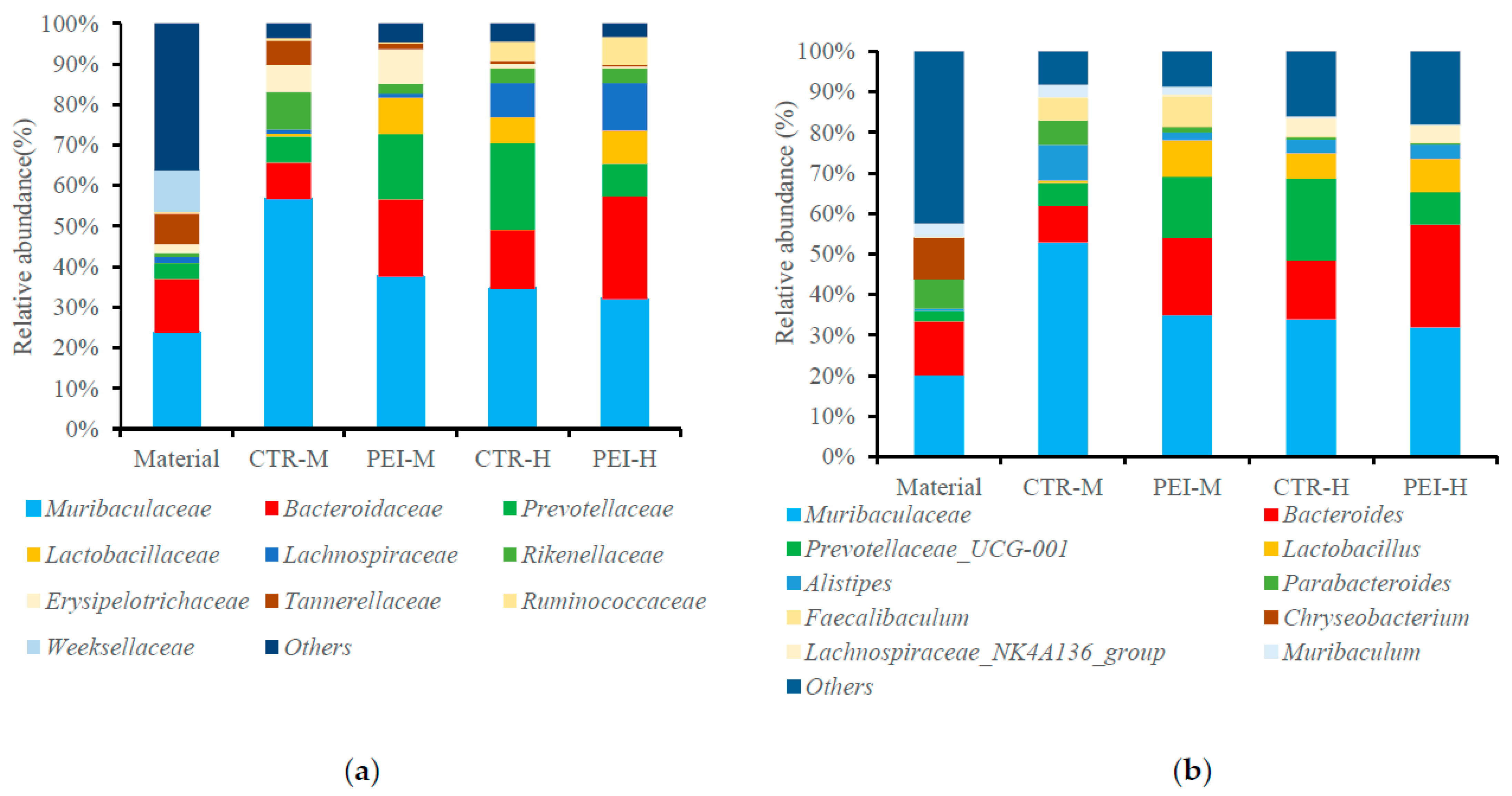
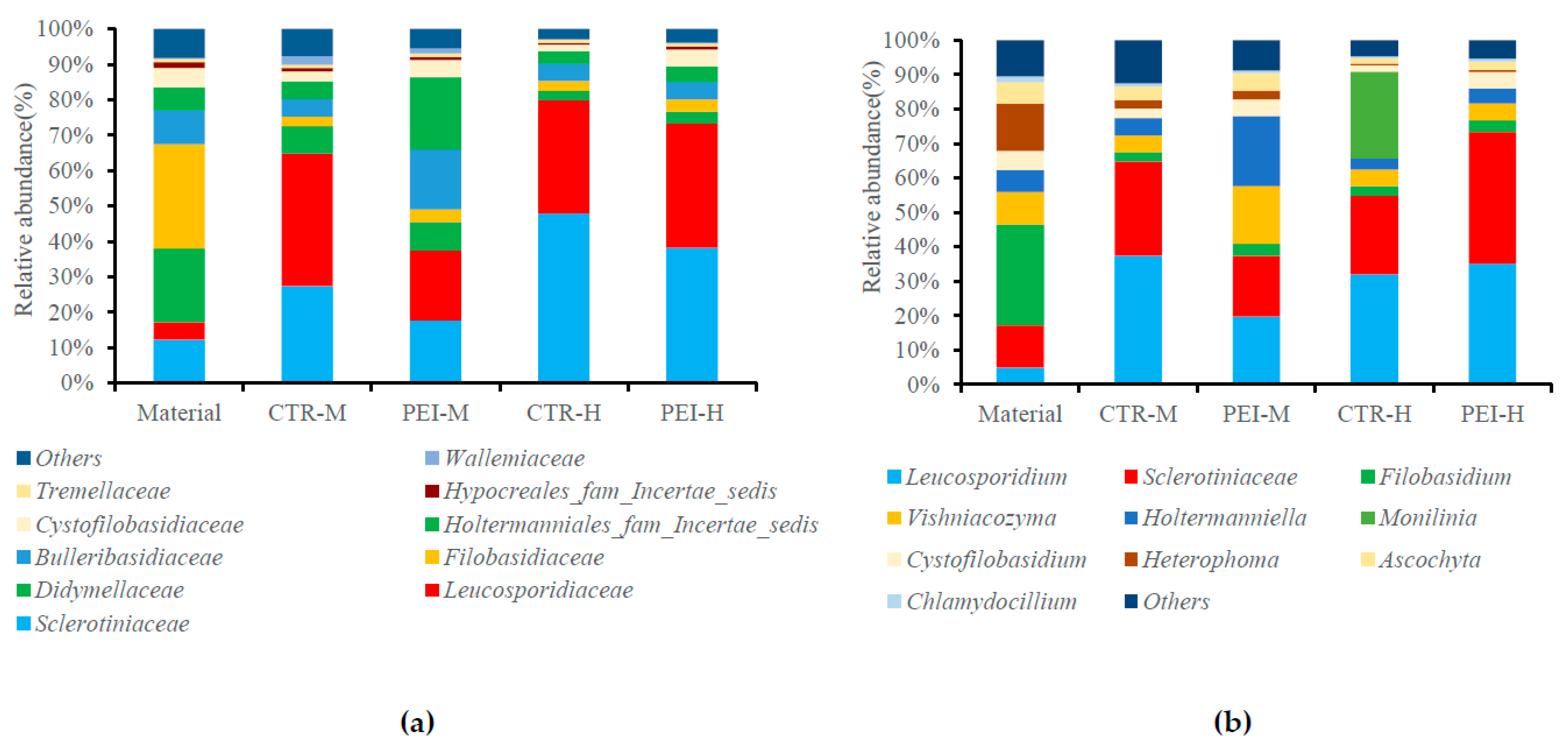
3.4. Analysis of the Composition of the Bacterial and Fungal Communities by Next-Generation Sequencing for Fresh Alfalfa and 90 d Silage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nkosi, B.D.; Meeske, R.; Langa, T.; Motiang, M.D.; Modiba, S.; Mkhize, N.R.; Groenewald, I.B. Effects of ensiling forage soybean (Glycine max (L.) Merr.) with or without bacterial inoculants on the fermentation characteristics, aerobic stability and nutrient digestion of the silage by Damara rams. Small Rumin. Res. 2016, 134, 90–96. [Google Scholar] [CrossRef]
- Przemieniecki, S.W.; Damszel, M.; Kurowski, T.P.; Mastalerz, J.; Kotlarz, K. Identification, ecological evaluation and phylogenetic analysis of non-symbiotic endophytic fungi colonizing timothy grass and perennial ryegrass grown in adjacent plots. Grass Forage Sci. 2019, 74, 42–52. [Google Scholar] [CrossRef] [Green Version]
- Weinberg, Z.G.; Muck, R.E. New trends in development and use of inoculants for silage. FEMS Microbiol. Rev. 1996, 19, 53–68. [Google Scholar] [CrossRef]
- Muck, R.E. Silage Additives and Management Issues. In Proceedings of the 2010 Idaho Alfalfa and Forage Conference Proceedings, Burley, ID, USA, 16–17 February 2010; pp. 49–55. [Google Scholar]
- Weinberg, Z.G.; Szakacs, G.; Ashbell, G.; Hen, Y. The effect of temperature on the ensiling process of corn and wheat. J. Appl. Microbiol. 2001, 90, 561–566. [Google Scholar] [CrossRef]
- Stallings, C.C.; Townes, R.; Jesse, B.W.; Thomas, J.W. Changes in alfalfa haylage during wilting and ensiling with and without additives. J. An. Sci. 1981, 53, 765–773. [Google Scholar] [CrossRef]
- McKersie, B.D. Effect of pH on proteolysis in ensiled legume forage. Agron. J. 1985, 77, 81–86. [Google Scholar] [CrossRef]
- Muck, R.E.; Nadeau, E.M.G.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.; Kung, L., Jr. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef]
- Sheperd, A.C.; Maslanka, M.; Quinn, D.; Kung, L., Jr. Additives containing bacteria and enzymes for alfalfa silage. J. Dairy Sci. 1995, 78, 565–572. [Google Scholar] [CrossRef]
- Whiter, A.G.; Kung, L., Jr. The effect of a dry or liquid application of Lactobacillus plantarum MTD1 on the fermentation of alfalfa silage. J. Dairy Sci. 2001, 84, 2195–2202. [Google Scholar] [CrossRef]
- Chen, Y.; Weinberg, Z.G. The effect of relocation of whole crop wheat and corn silages on their quality. J. Dairy Sci. 2014, 97, 406–410. [Google Scholar] [CrossRef]
- Lin, C.; Bolsen, K.K.; Brent, B.E.; Fung, D. Epiphytic lactic acid bacteria succession during the pre-ensiling and ensiling periods of alfalfa and maize. J. Appl. Bacteriol. 1992, 73, 375–387. [Google Scholar] [CrossRef]
- Weinberg, Z.G.; Ashbell, G.; Hen, Y.; Azrieli, A. The effect of applying lactic acid bacteria at ensiling on the aerobic stability of silages. J. Appl. Bacteriol. 1993, 75, 512–518. [Google Scholar] [CrossRef]
- Ranjit, N.K.; Kung, L. The effect of Lactobacillus buchneri, Lactobacillus plantarum, or a chemical preservative on the fermentation and aerobic stability of corn silage. J. Dairy Sci. 2000, 83, 526–535. [Google Scholar] [CrossRef]
- Li, F.; Ke, W.; Ding, Z.; Bai, J.; Zhang, Y.; Xu, D.; Li, Z.; Guo, X. Pretreatment of Pennisetum sinese silages with ferulic acid esterase-producing lactic acid bacteria and cellulase at two dry matter contents: Fermentation characteristics, carbohydrates composition and enzymatic saccharification. Bioresour. Technol. 2020, 295, 122261. [Google Scholar] [CrossRef] [PubMed]
- McAllister, T.A.; Dunière, L.; Drouin, P.; Xu, S.; Wang, Y.; Munns, K.; Zaheer, R. Silage review: Using molecular approaches to define the microbial ecology of silage. J. Dairy Sci. 2018, 101, 4060–4074. [Google Scholar] [CrossRef] [Green Version]
- Muck, R.E.; Dickerson, J.T. Storage temperature effects on proteolysis in alfalfa silage. Trans. ASAE 1988, 31, 1005–1009. [Google Scholar] [CrossRef]
- Armstrong, R.; Gibson, N.A.; Hosking, J.W.; Weatherburn, D.C. The preparation and properties of some quaternary arsonium compounds. Aust. J. Chem. 1967, 20, 2771–2776. [Google Scholar] [CrossRef]
- Nelson, N.A. Photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem. 1944, 153, 375–380. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2010. [Google Scholar]
- Krishnamoorthy, U.; Muscato, T.V.; Sniffen, C.J.; Van Soest, P.J. Nitrogen fractions in selected feedstuffs. J. Dairy Sci. 1982, 65, 217–225. [Google Scholar] [CrossRef]
- Van Soest, P.V.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Hall, M.B. Determination of starch, including maltooligosaccharides, in animal feeds: Comparison of methods and a method recommended for AOAC collaborative study. J. AOAC Int. 2008, 92, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Baker, G.C.; Smith, J.J.; Cowan, D.A. Review and re-analysis of domain specific 16S primers. J. Microbiol. Meth. 2003, 55, 541–555. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J.L. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, E.B.; Savage, R.M.; Biddle, A.S.; Polukis, S.A.; Smith, M.L.; Limin, K. Effects of a chemical additive on the fermentation, microbial communities, and aerobic stability of corn silage with or without air stress during storage. J. Anim. Sci. 2020, 98, skaa246. [Google Scholar] [CrossRef] [PubMed]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 7th ed.; The Iowa State University Press: Ames, IA, USA, 1980. [Google Scholar]
- Li, M.; Zhou, H.; Zi, X.; Cai, Y. Silage fermentation and ruminal degradation of stylo prepared with lactic acid bacteria and cellulase. Anim. Sci. J. 2017, 88, 1531–1537. [Google Scholar] [CrossRef]
- Li, Y.; da Silva, E.B.; Novinski, C.O.; Kung, L., Jr. Effect of microbial and chemical additives on the fermentation and aerobic stability of alfalfa silage ensiled at 2 dry matters and subjected to air stress during storage. J. Anim. Sci. 2021, 99, skab174. [Google Scholar] [CrossRef]
- Kung, L., Jr.; Stokes, M.R.; Lin, C.J. Silage additives. In Silage Science and Technology; Agronomy Series No. 42; Buxton, D.R., Muck, R.E., Harrison, J.H., Eds.; American Society of Agronomy: Madison, WI, USA, 2003; pp. 305–360. [Google Scholar]
- McDonald, P.; Henderson, N.; Heron, S.J.R. The Biochemistry of Silage; Chalcombe Publications: Marlow, UK, 1991. [Google Scholar]
- Kung, L., Jr.; Smith, M.L.; da Silva, E.B.; Windle, M.C.; da Silva, T.C.; Polukis, S.A. An evaluation of the effectiveness of a chemical additive based on sodium benzoate, potassium sorbate, and sodium nitrite on the fermentation and aerobic stability of corn silage. J. Dairy Sci. 2018, 101, 5949–5960. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, R.J.; Hu, W.; Mills, J.A.; Kung, L., Jr. The development of lactic acid bacteria and Lactobacillus buchneri and their effects on the fermentation of alfalfa silage. J. Dairy Sci. 2009, 92, 5005–5010. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Yao, Y.; Li, D.; Xu, H.; Wu, J.; Wen, A.; Xie, M.; Ni, Q.; Zhang, M.; Peng, G.; et al. Characterization of the Gut Microbiota in Six Geographical Populations of Chinese Rhesus Macaques (Macaca mulatta), Implying an Adaptation to High-Altitude Environment. Microb. Ecol. 2018, 76, 565–577. [Google Scholar] [CrossRef]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [Green Version]
- Bäckhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science. 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, R.K.; Anand, P.; Saran, S.; Isar, J.; Agarwal, L. Microbial production and applications of 1, 2-propanediol. Indian J. Microbiol. 2010, 50, 2–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fell, J.W.; Statzell, A.C.; Hunter, I.L.; Phaff, H.J. Leucosporidium gen. n., the heterbasidiomycetous stage of several yeasts of the genus Canidia. Antonie Van Leeuwenhoek 1969, 35, 433–462. [Google Scholar] [CrossRef] [PubMed]
- Bolton, M.D.; Thomma, B.P.H.J.; Nelson, B.D. Sclerotinia sclerotiorum (Lib.) de Bary: Biology and molecular traits of a cosmopolitan pathogen. Mol. Plant Pathol. 2006, 7, 1–16. [Google Scholar] [CrossRef]
- Gomes, N.C.M.; Fagbola, O.; Costa, R.; Rumjanek, N.G.; Buchner, A.; Mendona-Hagler, L.; Smalla, K. Dynamics of fungal communities in bulk and maize rhizosphere soil in the tropics. Appl. Environ. Microbiol. 2003, 69, 3758–3766. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Yang, J.; Qi, M.; Smiley, B.; Rutherford, W.; Wang, Y.; McAllister, T.A. Impact of Saccharomyces cerevisiae and Lactobacillus buchneri on microbial communities during ensiling and aerobic spoilage or corn silage. J. Anim. Sci. 2019, 97, 1273–1285. [Google Scholar] [CrossRef] [Green Version]
- Abdullabekova, D.A.; Magomedova, E.S.; Magomedov, G.G.; Aliverdieva, D.A.; Kachalkin, A.V. Yeast communities of chestnut soils under vineyards in Dagestan. Eurasian Soil Sci. 2017, 50, 1463–1467. [Google Scholar] [CrossRef]
- Vishniac, H.S. Cryptococcus tephrensis, sp. nov.; and Cryptococcus heimaeyensis, sp. nov.; a new anamorphic basidiomycetous yeast species from Iceland. Can. J. Microbiol. 2002, 48, 463–467. [Google Scholar] [CrossRef]
- Ball, D.M.; Collins, M.; Lacefield, G.D.; Martin, N.P.; Mertens, D.A.; Olson, K.E.; Putnam, D.H.; Undersand, D.J.; Wolf, M.W. Understanding Forage Quality; American Farm Bureau Federations Publication: Park Ridge, IL, USA, 2001. [Google Scholar]
- Pahlow, G.; Muck, R.E.; Driehuis, F.; Oude Elferink, S.J.W.H.; Spoelstra, S.F. Microbiology of ensiling. In Science and Technology Silage; Buxton, D.R., Muck, R.E., Harrison, J.H., Eds.; American Society of Agronmy: Madison, WI, USA, 2003; Chapter 2; pp. 31–93. [Google Scholar]



{kind=link}
{kind=link}
| Treatment | Targeted Final Lactic Acid Bacterial Targeted Application Rate—cfu/g of Fresh Forage | Actual Final Lactic Acid Bacterial Targeted Application Rate—cfu/g of Fresh Forage |
|---|---|---|
| Medium dry matter alfalfa | ||
| PEI 1 | 165,000 | 150,482 |
| High dry matter alfalfa | ||
| PEI | 165,000 | 106,533 |
| Item | Medium DM Alfalfa | High DM Alfalfa | SEM * |
|---|---|---|---|
| Dry matter | 37.82 | 46.05 | 1.46 |
| pH | 6.24 | 6.23 | 0.01 |
| Crude protein | 17.80 | 17.60 | 0.13 |
| Soluble crude protein, % of crude protein | 45.07 | 40.11 | 2.10 |
| Ammonia nitrogen | 0.03 | 0.03 | <0.01 |
| Acid detergent fiber | 35.58 | 35.04 | 0.19 |
| Neutral detergent fiber | 43.38 | 43.46 | 0.23 |
| Ash | 9.05 | 9.07 | 0.09 |
| Water soluble carbohydrates | 8.06 | 8.07 | 0.11 |
| Buffering capacity (gm lactic/100 g dry matter) | 5.56 | 5.65 | 0.04 |
| Lactic acid bacteria | 5.83 | 5.92 | 0.07 |
| Yeasts | 3.06 | 3.62 | 0.20 |
| Molds | 3.29 | 2.67 | 0.13 |
| Treatments | Dry Matter | Crude Protein | Soluble Crude Protein, % of Crude Protein | Ammonia Nitrogen | Acid Detergent Fiber | Neutral Detergent Fiber | Ash | Water Soluble Carbohydrates |
|---|---|---|---|---|---|---|---|---|
| Day 3 | ||||||||
| Medium dry matter alfalfa | ||||||||
| CTR * | 38.82 b | 17.76 b | 50.46 d | 0.11 d | nd | nd | nd | 6.85 c |
| PEI | 36.50 b | 18.24 a | 50.96 d | 0.12 d | nd | nd | nd | 7.01 bc |
| High dry matter alfalfa | ||||||||
| CTR | 45.51 a | 17.74 b | 49.74 d | 0.11 d | nd | nd | nd | 8.31 a |
| PEI | 45.51 a | 17.74 b | 48.56 d | 0.11 d | nd | nd | nd | 8.11 ab |
| Day 30 | ||||||||
| Medium DM alfalfa | ||||||||
| CTR | 37.06 b | 18.16 ab | 61.14 bc | 0.23 ab | nd | nd | nd | 3.50 efg |
| PEI | 36.54 b | 18.32 a | 59.18 c | 0.22 c | nd | nd | nd | 4.95 d |
| High dry matter alfalfa | ||||||||
| CTR | 46.21 a | 17.74 b | 59.38 c | 0.22 bc | nd | nd | nd | 4.12 def |
| PEI | 45.51 a | 18.06 ab | 60.66 bc | 0.22 bc | nd | nd | nd | 3.57 efg |
| Day 60 | ||||||||
| Medium dry matter alfalfa | ||||||||
| CTR | 36.81 b | 18.18 ab | 62.36 ab | 0.25 a | 36.00 ab | 41.32 a | 9.49 a | 4.44 de |
| PEI | 36.04 b | 18.48 a | 65.24 a | 0.21 bc | 35.24 b | 40.70 b | 9.67 a | 6.37 c |
| High dry matter alfalfa | ||||||||
| CTR | 44.29 a | 18.06 ab | 60.72 bc | 0.25 a | 36.64 a | 43.66 a | 9.86 a | 3.02 fg |
| PEI | 44.35 a | 18.30 a | 58.28 c | 0.23 ab | 36.24 ab | 43.54 a | 10.00 a | 2.71 g |
| SEM | 0.57 | 0.04 | 0.73 | 0.01 | 0.17 | 0.32 | 0.08 | 0.21 |
| Effects and interactions | p-value | |||||||
| INO | 0.05 | <0.01 | 0.67 | <0.01 | 0.04 | 0.14 | 0.25 | <0.01 |
| DRM | <0.01 | <0.01 | <0.01 | 0.55 | <0.01 | <0.01 | 0.02 | <0.01 |
| DAY | 0.02 | <0.01 | <0.01 | <0.01 | nd | nd | nd | <0.01 |
| INO × DRM | 0.17 | 0.27 | 0.08 | 0.11 | 0.50 | 0.31 | 0.88 | <0.01 |
| INO × DAY | 0.65 | 0.97 | 0.76 | <0.01 | nd | nd | nd | 0.06 |
| DRM × DAY | 0.30 | 0.40 | <0.01 | 0.16 | nd | nd | nd | <0.01 |
| DAY × DRM × INO | 0.36 | 0.08 | <0.01 | 0.01 | nd | nd | nd | 0.02 |
| Treatments | Dry Matter | Crude Protein | Acid Detergent Fiber | Neutral Detergent Fiber | Ash |
|---|---|---|---|---|---|
| INO | |||||
| CTR * | 41.45 | 17.94 b | 36.32 a | 42.49 | 9.67 |
| PEI | 40.74 | 18.19 a | 35.74 b | 42.12 | 9.83 |
| DRM | |||||
| Medium dry matter | 36.96 b | 18.25 a | 35.62 b | 41.01 b | 9.58 b |
| High dry matter | 45.20 a | 17.92 b | 36.44 a | 43.60 a | 9.93 a |
| Day | |||||
| 3 | 41.58 a | 17.87 c | nd | nd | nd |
| 30 | 41.33 ab | 18.07 b | nd | nd | nd |
| 60 | 40.37 b | 18.26 a | 36.03 | 42.30 | 9.76 |
| INO × DRM | |||||
| Medium dry matter alfalfa | |||||
| CTR | 37.57 b | 18.03 b | 36.00 ab | 41.32 b | 9.49 b |
| PEI | 36.36 c | 18.35 a | 35.24 b | 40.70 b | 9.67 ab |
| High dry matter alfalfa | |||||
| CTR | 45.33 a | 17.85 c | 36.64 a | 43.66 a | 9.86 ab |
| PEI | 45.12 a | 18.03 b | 36.24 b | 43.54 a | 10.00 a |
| Effects and interactions | p-value | ||||
| INO | 0.05 | <0.01 | 0.04 | 0.14 | 0.25 |
| DRM | <0.01 | <0.01 | <0.01 | <0.01 | 0.02 |
| DAY | 0.02 | <0.01 | nd | nd | nd |
| INO × DRM | 0.17 | 0.27 | 0.50 | 0.31 | 0.88 |
| Treatments | pH | Lactic Acid | Acetic Acid | 1,2- Propanediol | Propionic Acid | Ethanol | Lactic Acid Bacteria | Yeasts |
|---|---|---|---|---|---|---|---|---|
| Day 3 | ||||||||
| Medium dry matter alfalfa | ||||||||
| CTR * | 5.51 b | 1.61 ef | 0.71 d | 0.00 d | 0.12 bc | 1.12 | 9.12 abc | 3.87 a |
| PEI | 5.17 c | 2.28 e | 0.85 cd | 0.00 d | 0.13 bc | 0.97 | 9.26 ab | 3.77 a |
| High dry matter alfalfa | ||||||||
| CTR | 5.69 a | 1.26 f | 0.58 d | 0.00 d | 0.09 bc | 0.87 | 9.72 a | 1.90 cd |
| PEI | 5.73 a | 1.24 f | 0.60 b | 0.00 d | 0.10 bc | 0.92 | 9.22 abc | 1.69 d |
| Day 30 | ||||||||
| Medium dry matter alfalfa | ||||||||
| CTR | 4.35 f | 5.89 c | 2.40 ab | 0.18 c | 0.35 a | 1.12 | 7.12 cd | 3.02 abc |
| PEI | 4.08 h | 7.04 ab | 1.24 c | 0.00 d | 0.15 bc | 0.80 | 3.60 e | 3.00 abc |
| High dry matter alfalfa | ||||||||
| CTR | 4.70 d | 4.04 d | 2.37 ab | 0.39 b | 0.18 bc | 1.04 | 8.80 abc | 2.91 abc |
| PEI | 4.64 d | 3.75 d | 2.07 b | 0.36 b | 0.19 b | 0.90 | 8.94 abc | 2.59 bcd |
| Day 60 | ||||||||
| Medium dry matter alfalfa | ||||||||
| CTR | 4.29 g | 6.44 bc | 2.57 a | 0.28 bc | 0.05 c | 1.15 | 7.50 bcd | 2.75 abcd |
| PEI | 4.08 h | 7.32 a | 2.06 b | 0.17 c | 0.34 a | 0.83 | 5.58 de | 3.08 ab |
| High dry matter alfalfa | ||||||||
| CTR | 4.53 e | 4.13 d | 2.57 a | 0.66 a | 0.07 bc | 0.96 | 8.51 abc | 3.30 ab |
| PEI | 4.50 e | 4.17 d | 2.50 ab | 0.63 a | 0.18 b | 0.94 | 8.67 abc | 2.99 abc |
| SEM | 0.07 | 0.28 | 0.11 | 0.03 | 0.01 | 0.03 | 0.25 | 0.10 |
| Effects and interactions | p-value | |||||||
| INO | <0.01 | <0.01 | <0.01 | <0.01 | 0.01 | <0.01 | <0.01 | 0.54 |
| DRM | <0.01 | <0.01 | 0.01 | <0.01 | <0.01 | 0.21 | <0.01 | 0.39 |
| DAY | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | 0.99 | <0.01 | <0.01 |
| INO × DRM | <0.01 | <0.01 | <0.01 | 0.02 | 0.64 | 0.02 | 0.02 | 0.03 |
| INO × DAY | 0.08 | 0.85 | <0.01 | 0.03 | <0.01 | 0.31 | 0.06 | 0.14 |
| DRM × DAY | <0.01 | <0.01 | <0.01 | <0.01 | 0.50 | 0.37 | <0.01 | <0.01 |
| DAY × DRM × INO | <0.01 | 0.29 | <0.01 | 0.15 | <0.01 | 0.84 | <0.01 | <0.01 |
| Bacterial Genus | Fungal Genus | |||||||
|---|---|---|---|---|---|---|---|---|
| Item | Muribaculaceae | Bacteroides | Prevotellaceae-UCG-001 | Lactobacillus | Leucosporidium | Sclerotiniaceae | Filobasidium | Vishniacozyma |
| Material | 20.08 | 13.27 | 2.61 | 0.05 | 4.91 | 12.28 | 29.32 | 9.43 |
| Medium dry matter alfalfa | ||||||||
| CTR * | 52.86 a | 8.96 c | 5.66 b | 0.72 b | 37.40 a | 27.40 ab | 2.68 | 4.89 b |
| PEI | 34.94 b | 19.04 ab | 15.13 a | 8.95 a | 19.72 b | 17.72 b | 3.45 | 16.74 a |
| High dry matter alfalfa | ||||||||
| CTR | 33.88 b | 14.54 b | 20.15 a | 6.32 a | 32.00 a | 22.86 b | 2.75 | 4.81 b |
| PEI | 31.87 b | 25.38 a | 8.03 b | 8.18 a | 35.16 a | 38.19 a | 3.44 | 4.86 b |
| SEM | 4.20 | 2.64 | 2.14 | 1.52 | 3.26 | 3.61 | 2.46 | 1.07 |
| Effects and interactions | p-value | |||||||
| INO | 0.30 | 0.03 | 0.78 | 0.04 | 0.24 | 0.58 | 0.26 | <0.01 |
| DRM | 0.25 | 0.04 | 0.45 | 0.04 | 0.41 | 0.13 | 0.97 | <0.01 |
| INO × DRM | 0.04 | 0.04 | 0.04 | 0.03 | 0.04 | 0.03 | 0.95 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; da Silva, E.B.; Li, J.; Kung, L., Jr. Effect of Homo-Fermentative Lactic Acid Bacteria Inoculants on Fermentation Characteristics and Bacterial and Fungal Communities in Alfalfa Silage. Fermentation 2022, 8, 621. https://doi.org/10.3390/fermentation8110621
Li Y, da Silva EB, Li J, Kung L Jr. Effect of Homo-Fermentative Lactic Acid Bacteria Inoculants on Fermentation Characteristics and Bacterial and Fungal Communities in Alfalfa Silage. Fermentation. 2022; 8(11):621. https://doi.org/10.3390/fermentation8110621
Chicago/Turabian StyleLi, Yanbing, E. B. da Silva, Jingchun Li, and L. Kung, Jr. 2022. "Effect of Homo-Fermentative Lactic Acid Bacteria Inoculants on Fermentation Characteristics and Bacterial and Fungal Communities in Alfalfa Silage" Fermentation 8, no. 11: 621. https://doi.org/10.3390/fermentation8110621