
Recent Advances in Multiple Strategies for the Synthesis of Terpenes by Engineered Yeast

Abstract
:1. Introduction
2. The Adaptive Screening and Expression of Terpene Synthases in Yeast
2.1. The Screening of Terpene Synthases from Different Origins
2.2. The Enhancement of Terpene Synthases’ Expression in Yeast
2.3. The Artificial Design of Amino Acid Sequences Based on the Computer-Aided Analysis
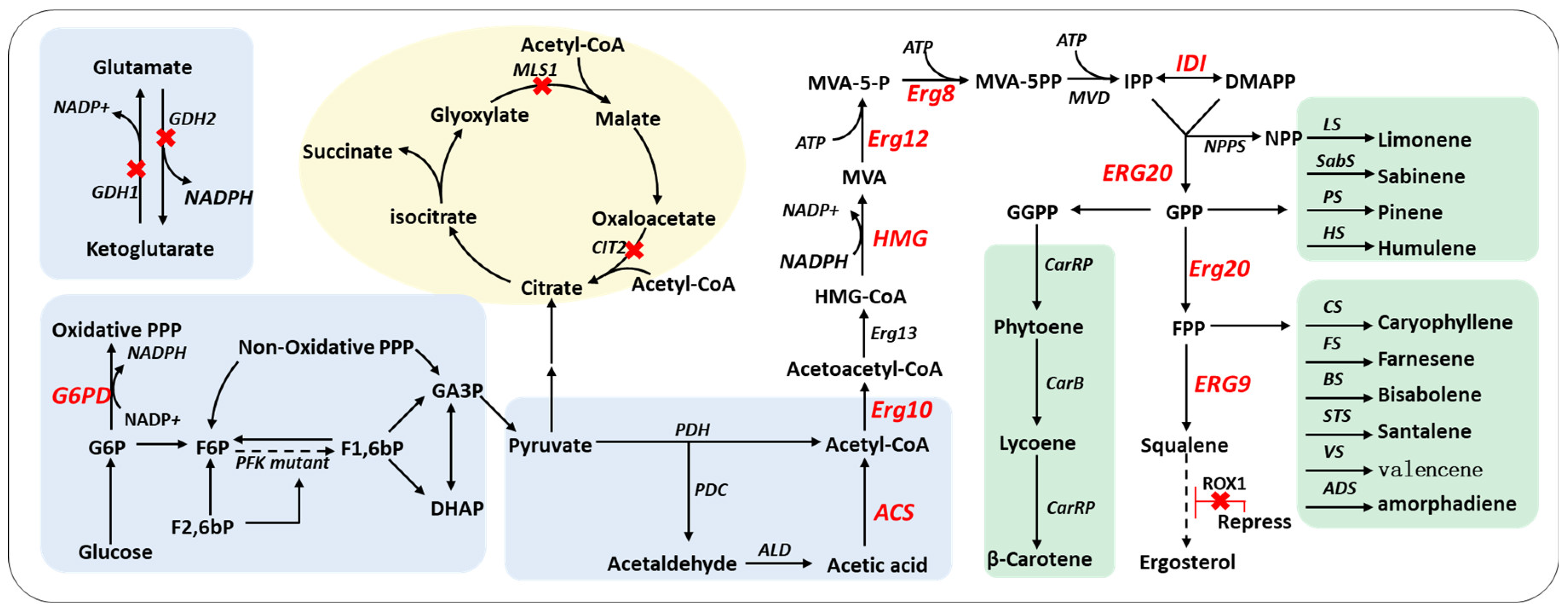
3. The Strategies for the Regulation of the Synthesis Pathway
3.1. The Regulation of the Rate-Limiting Steps in the Pathway
3.2. The Regulation of Cofactor NADPH Supply
3.3. The Enhancement of the Central Carbon Flux
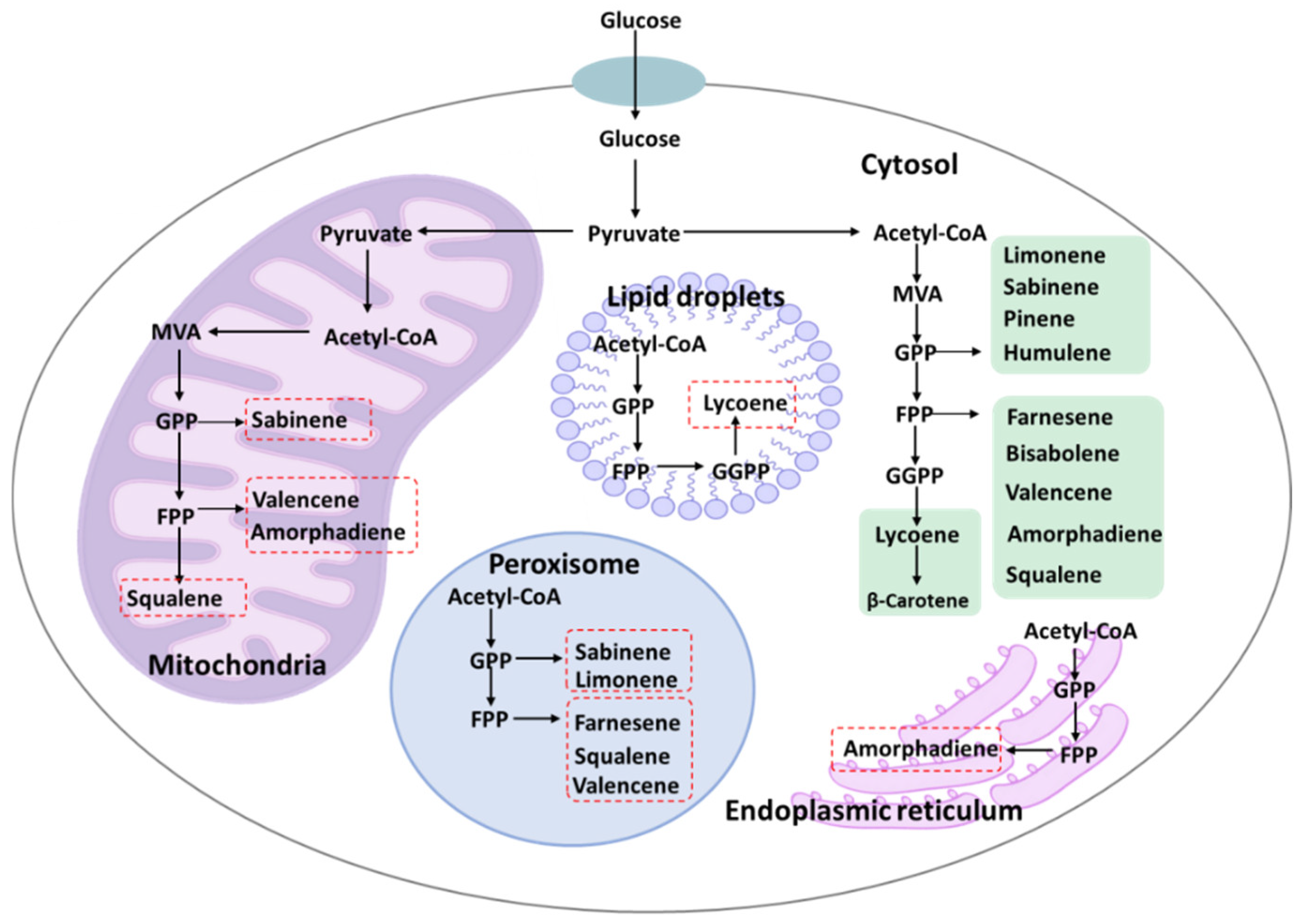
4. The Intracellular Compartmentalized Expression of Metabolic Pathways
4.1. The Promotion of Terpene Biosynthesis via Mitochondria Engineering
4.2. The Promotion of Terpene Biosynthesis via Peroxisome Engineering
4.3. The Development of Other Organelles for Terpene Biosynthesis
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tholl, D. Terpene synthases and the regulation, diversity and biological roles of terpene metabolism. Curr. Opin. Plant Biol. 2006, 9, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Chandi, G.K.; Gill, B.S. Production and characterization of microbial carotenoids as an alternative to synthetic colors: A review. Int. J. Food Prop. 2011, 14, 503–513. [Google Scholar] [CrossRef]
- Smith, T.J. Squalene: Potential chemopreventive agent. Expert Opin. Investig. Drug 2000, 9, 1841–1848. [Google Scholar] [CrossRef] [PubMed]
- Harvey, B.G.; Meylemans, H.A.; Gough, R.V.; Quintana, R.L.; Garrison, M.D. High-density biosynthetic fuels: The intersection of heterogeneous catalysis and metabolic engineering. Phys. Chem. 2014, 16, 9448–9457. [Google Scholar] [CrossRef]
- Harvey, B.G.; Wright, M.E.; Quintana, R.L. High-density renewable fuels based on the selective dimerization of pinenes. Energy Fuel 2010, 24, 267–273. [Google Scholar] [CrossRef]
- Naziri, E.; Mantzouridou, F.; Tsimidou, M.Z. Squalene resources and uses point to the potential of biotechnology. Lipid Technol. 2011, 23, 270–273. [Google Scholar] [CrossRef]
- Rosales-García, T.; Jimenez-Martinez, C.; Davila-Ortiz, G. Squalene Extraction: Biological Sources and Extraction Methods. IJEAB 2017, 2, 238838. [Google Scholar] [CrossRef]
- Global Market Insights, Squalene Market Size by Source, by Application, Industry Analysis Report, Regional Outlook, Application Potential, Price Trends, Competitive Market Share & Forecast 2015–2022. 2016. Available online: https://www.gminsights.com/industry-analysis/squalene-market (accessed on 16 September 2022).
- Zhu, Z.-T.; Du, M.-M.; Gao, B.; Tao, X.Y.; Zhao, M.; Ren, Y.-H.; Wang, F.-Q.; Wei, D.-Z. Metabolic compartmentalization in yeast mitochondria: Burden and solution for squalene overproduction. Metab. Eng. 2021, 68, 232–245. [Google Scholar] [CrossRef]
- Ciriminna, R.; Lomeli-Rodriguez, M.; Demma Carà, P.; Lopez-Sanchez, J.A.; Pagliaro, M. Limonene: A versatile chemical of the bioeconomy. Chem. Commun. 2014, 50, 15288–15296. [Google Scholar] [CrossRef]
- Felipe, L.d.O.; Oliveira, A.M.d.; Bicas, J.L. Bioaromas—Perspectives for sustainable development. Trends Food Sci. Technol. 2017, 62, 141–153. [Google Scholar] [CrossRef]
- Johnson, T.J.; Jahandideh, A.; Johnson, M.D.; Fields, K.H.; Richardson, J.W.; Muthukumarappan, K.; Cao, Y.; Gu, Z.; Halfmann, C.; Zhou, R.; et al. Producing next-generation biofuels from filamentous cyanobacteria: An economic feasibility analysis. Algal Res. 2016, 20, 218–228. [Google Scholar] [CrossRef] [Green Version]
- Mukhtar, Y.M.; Adu-Frimpong, M.; Xu, X.; Yu, J. Biochemical significance of limonene and its metabolites: Future prospects for designing and developing highly potent anticancer drugs. Biosci. Rep. 2018, 38, BSR20181253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Global Market Insights, Dipentene (Limonene) Market Size By Grade (Food Grade, Technical Grade), By End-user (Personal Care, Food & Beverages, Pharmaceutical, Electronics, Chemicals, Paints & Coatings, Rubber, Agriculture), Industry Analysis Report, Regional Outlook, Application Growth Potential, Price Trends, Competitive Market Share & Forecast 2018–2024. 2018. Available online: https://www.gminsights.com/industry-analysis/dipentene-market (accessed on 16 September 2022).
- Zhang, X.; Liu, X.; Meng, Y.; Zhang, L.; Qiao, J.; Zhao, G.-R. Combinatorial engineering of Saccharomyces cerevisiae for improving limonene production. Biochem. Eng. J. 2021, 176, 108155. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, X.; Zhan, Y.; Wang, K.; Francis, F.; Liu, Y. New slow release mixture of (E)-β-farnesene with methyl salicylate to enhance aphid biocontrol efficacy in wheat ecosystem. Pest Manag. Sci. 2021, 77, 3341–3348. [Google Scholar] [CrossRef] [PubMed]
- Peralta-Yahya, P.P.; Zhang, F.; del Cardayre, S.B.; Keasling, J.D. Microbial engineering for the production of advanced biofuels. Nature 2012, 488, 320–328. [Google Scholar] [CrossRef]
- Beller, H.R.; Lee, T.S.; Katz, L. Natural products as biofuels and bio-based chemicals: Fatty acids and isoprenoids. Nat. Prod. Rep. 2015, 32, 1508–1526. [Google Scholar] [CrossRef]
- Global Market Insights, Farnesene Market Size By Application (Cosmetics & Personal Care, Performance Material, Flavors & Fragrances, Fuels & Lubes), Industry Analysis Report, Regional Outlook, Application Potential, Price Trends, Competitive Market Share & Forecast 2021–2027. 2021. Available online: https://www.gminsights.com/industry-analysis/farnesene-market (accessed on 16 September 2022).
- Meadows, A.L.; Hawkins, K.M.; Tsegaye, Y.; Antipov, E.; Kim, Y.; Raetz, L.; Dahl, R.H.; Tai, A.; Mahatdejkul-Meadows, T.; Xu, L.; et al. Rewriting yeast central carbon metabolism for industrial isoprenoid production. Nature 2016, 537, 694–697. [Google Scholar] [CrossRef]
- Mezzomo, N.; Ferreira, S.R.S. Carotenoids Functionality, Sources, and Processing by Supercritical Technology: A Review. J. Chem. 2016, 2016, 3164312. [Google Scholar] [CrossRef] [Green Version]
- Global Market Insights, Carotenoids Market Size, Share and Industry Analysis Report by Source (Natural, Synthetic), Product (Beta-carotene, Lutein, Astaxanthin, Canthaxanthin, Lycopene) and Application (Food & Beverages, Pharmaceutical, Cosmetics, Animal Feed, Dietary supplements), Regional Outlook, Growth Potential, Competitive Market Share & Forecast 2016–2024. 2016. Available online: https://www.gminsights.com/industry-analysis/carotenoids-market (accessed on 16 September 2022).
- Liu, M.; Zhang, J.; Liu, X.; Hou, J.; Qi, Q. Rapid Gene Target Tracking for Enhancing β-Carotene Production Using Flow Cytometry-Based High-Throughput Screening in Yarrowia lipolytica. Appl. Environ. Microbiol. 2022, 88, e01149-22. [Google Scholar] [CrossRef]
- Chen, Y.; Xiao, W.; Wang, Y.; Liu, H.; Li, X.; Yuan, Y. Lycopene overproduction in Saccharomyces cerevisiae through combining pathway engineering with host engineering. Microb. Cell Factories 2016, 15, 113. [Google Scholar] [CrossRef]
- Novoveská, L.; Ross, M.E.; Stanley, M.S.; Pradelles, R.; Wasiolek, V.; Sassi, J.-F. Microalgal Carotenoids: A Review of Production, Current Markets, Regulations, and Future Direction. Mar. Drugs 2019, 17, 640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Global Market Insights, Astaxanthin Market Size By Source (Synthetic, Natural), By Application (Dietary Supplement, Personal Care, Pharmaceuticals, Food & Beverages, Animal Feed {Aquaculture, Livestock, Pets}), Industry Outlook Report, Regional Analysis, Application Potential, Covid-19 Impact Analysis, Price Trends, Competitive Market Share & Forecast 2021–2027. 2021. Available online: https://www.gminsights.com/industry-analysis/astaxanthin-market (accessed on 16 September 2022).
- Gong, Z.; Wang, H.; Tang, J.; Bi, C.; Li, Q.; Zhang, X. Coordinated Expression of Astaxanthin Biosynthesis Genes for Improved Astaxanthin Production in Escherichia coli. J. Agric. Food Chem. 2020, 68, 14917–14927. [Google Scholar] [CrossRef] [PubMed]
- Agledal, L.; Niere, M.; Ziegler, M. The phosphate makes a difference: Cellular functions of NADP. Redox Rep. 2010, 15, 2–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chemler, J.A.; Koffas, M.A.G. Metabolic engineering for plant natural product biosynthesis in microbes. Curr. Opin. Biotech. 2008, 19, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Marienhagen, J.; Bott, M. Metabolic engineering of microorganisms for the synthesis of plant natural products. J. Biotechnol. 2013, 163, 166–178. [Google Scholar] [CrossRef]
- Wang, C.; Pfleger, B.F.; Kim, S.-W. Reassessing Escherichia coli as a cell factory for biofuel production. Curr. Opin. Biotech. 2017, 45, 92–103. [Google Scholar] [CrossRef] [Green Version]
- Martin, V.J.J.; Pitera, D.J.; Withers, S.T.; Newman, J.D.; Keasling, J.D. Engineering a mevalonate pathway in Escherichia coli for production of terpenoids. Nat. Biotechnol. 2003, 21, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.S.; Thodey, K.; Trenchard, I.; Smolke, C.D. Advancing secondary metabolite biosynthesis in yeast with synthetic biology tools. FEMS Yeast Res. 2012, 12, 144–170. [Google Scholar] [CrossRef]
- Shi, B.; Ma, T.; Ye, Z.; Li, X.; Huang, Y.; Zhou, Z.; Ding, Y.; Deng, Z.; Liu, T. Systematic Metabolic Engineering of Saccharomyces cerevisiae for Lycopene Overproduction. J. Agric. Food Chem. 2019, 67, 11148–11157. [Google Scholar] [CrossRef]
- Wei, L.-J.; Zhong, Y.-T.; Nie, M.-Y.; Liu, S.-C. Biosynthesis of α-Pinene by Genetically Engineered Yarrowia lipolytica from Low-Cost Renewable Feedstocks. J. Agric. Food Chem. 2021, 69, 275–285. [Google Scholar]
- Zhao, Y.; Hua, Q. High-efficiency production of bisabolene from waste cooking oil by metabolically engineered Yarrowia lipolytica. Microb. Biotechnol. 2021, 14, 2497–2513. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chen, S.-L.; Xu, J.-Z.; Zhang, W.-G. Dual Regulation of Cytoplasm and Peroxisomes for Improved A-Farnesene Production in Recombinant Pichia pastoris. ACS Synth. Biol. 2021, 10, 1563–1573. [Google Scholar] [CrossRef] [PubMed]
- Bohlmann, J.; Meyer-Gauen, G.; Croteau, R. Plant terpenoid synthases: Molecular biology and phylogenetic analysis. Proc. Natl. Acad. Sci. USA 1998, 95, 4126–4133. [Google Scholar] [CrossRef] [Green Version]
- Peralta-Yahya, P.P.; Ouellet, M.; Chan, R.; Mukhopadhyay, A.; Keasling, J.D.; Lee, T.S. Identification and microbial production of a terpene-based advanced biofuel. Nature 2011, 2, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Katabami, A.; Li, L.; Iwasaki, M.; Furubayashi, M.; Saito, K.; Umeno, D. Production of squalene by squalene synthases and their truncated mutants in Escherichia coli. J. Biosci. Bioeng. 2015, 119, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, X.; Chappell, J. Building terpene production platforms in yeast. Biotechnol. Bioeng. 2015, 112, 1854–1864. [Google Scholar] [CrossRef]
- Foster, A.J.; Hall, D.E.; Mortimer, L.; Abercromby, S.; Gries, R.; Gries, G.; Bohlmann, J.; Russell, J.; Mattsson, J. Identification of genes in Thuja plicata foliar terpenoid defenses. Plant Physiol. 2013, 161, 1993–2004. [Google Scholar] [CrossRef] [Green Version]
- Roach, C.R.; Hall, D.E.; Zerbe, P.; Bohlmann, J. Plasticity and evolution of (+)-3-carene synthase and (−)-sabinene synthase functions of a Sitka spruce monoterpene synthase gene family associated with weevil resistance. J. Biol. Chem. 2014, 289, 23859–23869. [Google Scholar] [CrossRef] [Green Version]
- Hyatt, D.C.; Youn, B.; Zhao, Y.; Santhamma, B.; Coates, R.M.; Croteau, R.B.; Kang, C. Structure of limonene synthase, a simple model for terpenoid cyclase catalysis. Proc. Natl. Acad. Sci. USA 2007, 104, 5360–5365. [Google Scholar] [CrossRef] [Green Version]
- Schilmiller, A.L.; Schauvinhold, I.; Larson, M.; Xu, R.; Charbonneau, A.L.; Schmidt, A.; Wilkerson, C.; Last, R.L.; Pichersky, E. Monoterpenes in the glandular trichomes of tomato are synthesized from a neryl diphosphate precursor rather than geranyl diphosphate. Proc. Natl. Acad. Sci. India B 2009, 106, 10865–10870. [Google Scholar] [CrossRef] [Green Version]
- Cheng, B.-Q.; Wei, L.-J.; Lv, Y.-B.; Hua, Q. Elevating Limonene Production in Oleaginous Yeast Yarrowia lipolytica via Genetic Engineering of Limonene Biosynthesis Pathway and Optimization of Medium Composition. Biotechnol. Bioproc. E 2019, 24, 500–506. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, J.; Ye, J.; Qi, Q.; Hou, J. Morphological and Metabolic Engineering of Yarrowia lipolytica to Increase β-Carotene Production. ACS Synth. Biol. 2021, 10, 3551–3560. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.Z.; Rouvière, P.E.; LaRossa, R.A.; Suh, W. Chromosomal promoter replacement of the isoprenoid pathway for enhancing carotenoid production in E. coli. Metab. Eng. 2006, 8, 79–90. [Google Scholar] [CrossRef]
- Dulermo, R.; Brunel, F.; Dulermo, T.; Ledesma-Amaro, R.; Vion, J.; Trassaert, M.; Thomas, S.; Nicaud, J.M.; Leplat, C. Using a vector pool containing variable-strength promoters to optimize protein production in Yarrowia lipolytica. Microb. Cell Fact. 2017, 16, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larroude, M.; Celinska, E.; Back, A.; Thomas, S.; Nicaud, J.-M.; Ledesma-Amaro, R. A synthetic biology approach to transform Yarrowia lipolytica into a competitive biotechnological producer of β-carotene. Biotechnol. Bioeng. 2018, 115, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Turner, G.; Gershenzon, J.; Nielson, E.E.; Froehlich, J.E.; Croteau, R. Limonene synthase, the enzyme responsible for monoterpene biosynthesis in peppermint, is localized to leucoplasts of oil gland secretory cells. Plant Phys. 1999, 120, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.C.; McGarvey, D.J.; Katahira, E.J.; Croteau, R. Truncation of Limonene Synthase Preprotein Provides a Fully Active ‘Pseudomature’ Form of This Monoterpene Cyclase and Reveals the Function of the Amino-Terminal Arginine Pair. Biochemistry 1998, 37, 12213–12220. [Google Scholar] [CrossRef]
- Jia, H.; Chen, T.; Qu, J.; Yao, M.; Xiao, W.; Wang, Y.; Li, C.; Yuan, Y. Collaborative subcellular compartmentalization to improve GPP utilization and boost sabinene accumulation in Saccharomyces cerevisiae. Biochem. Eng. J. 2020, 164, 107768. [Google Scholar] [CrossRef]
- Jongedijk, E.; Cankar, K.; Buchhaupt, M.; Schrader, J.; Bouwmeester, H.; Beekwilder, J. Biotechnological production of limonene in microorganisms. Appl. Microbiol. Biotechnol. 2016, 100, 2927–2938. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Lv, Y.-B.; Chen, J.; Imanaka, T.; Wei, L.-J.; Hua, Q. Metabolic engineering of oleaginous yeast Yarrowia lipolytica for limonene overproduction. Biotechnol. Biofuels 2016, 9, 214. [Google Scholar] [CrossRef] [Green Version]
- Tianhua, C.; Ruosi, Z.; Guozhen, J.; Mingdong, Y.; Hong, L.; Ying, W.; Wenhai, X.; Yingjin, Y. Metabolic engineering of Saccharomyces cerevisiae for pinene production. CIESC J. 2019, 70, 179–188. [Google Scholar]
- Stermer, B.A.; Bostock, R.M. Involvement of 3-hydroxy-3-methylglutaryl coenzyme A reductase in the regulation of sesquiterpenoid phytoalexin synthesis in potato. Plant Phys. 1987, 84, 404–408. [Google Scholar] [CrossRef] [PubMed]
- Matthäus, F.; Ketelhot, M.; Gatter, M.; Barth, G. Production of Lycopene in the Non-Carotenoid-Producing Yeast Yarrowia lipolytica. Appl. Environ. Microb. 2014, 80, 1660–1669. [Google Scholar] [CrossRef] [Green Version]
- Stermer, B.A.; Bianchini, G.M.; Korth, K.L. Regulation of HMG-CoA reductase activity in plants. J. Lipid Res. 1994, 35, 1133–1140. [Google Scholar] [CrossRef]
- Nielsen, J. Systems biology of lipid metabolism: From yeast to human. FEBS Lett. 2009, 583, 3905–3913. [Google Scholar] [CrossRef] [Green Version]
- Donald, K.A.; Hampton, R.Y.; Fritz, I.B. Effects of overproduction of the catalytic domain of 3-hydroxy-3-methylglutaryl coenzyme A reductase on squalene synthesis in Saccharomyces cerevisiae. Appl. Environ. Microb. 1997, 63, 3341–3344. [Google Scholar] [CrossRef] [Green Version]
- Polakowski, T.; Stahl, U.; Lang, C. Overexpression of a cytosolic hydroxymethylglutaryl-CoA reductase leads to squalene accumulation in yeast. Appl. Microbiol. Biotechnol. 1998, 49, 66–71. [Google Scholar] [CrossRef]
- Ro, D.-K.; Paradise, E.M.; Ouellet, M.; Fisher, K.J.; Newman, K.L.; Ndungu, J.M.; Ho, K.A.; Eachus, R.A.; Ham, T.S.; Kirby, J.; et al. Production of the antimalarial drug precursor artemisinic acid in engineered yeast. Nature 2006, 440, 940–943. [Google Scholar] [CrossRef]
- Verwaal, R.; Wang, J.; Meijnen, J.-P.; Visser, H.; Sandmann, G.; van den Berg Johan, A.; van Ooyen Albert, J.J. High-Level Production of Beta-Carotene in Saccharomyces cerevisiae by Successive Transformation with Carotenogenic Genes from Xanthophyllomyces dendrorhous. Appl. Environ. Microb. 2007, 73, 4342–4350. [Google Scholar] [CrossRef] [Green Version]
- Rasool, A.; Ahmed, M.S.; Li, C. Overproduction of squalene synergistically downregulates ethanol production in Saccharomyces cerevisiae. Chem. Eng. Sci. 2016, 152, 370–380. [Google Scholar] [CrossRef]
- Sivy, T.L.; Fall, R.; Rosenstiel, T.N. Evidence of Isoprenoid Precursor Toxicity in Bacillus subtilis. Biosci. Biotech. Biochem. 2011, 75, 2376–2383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthelot, K.; Estevez, Y.; Deffieux, A.; Peruch, F. Isopentenyl diphosphate isomerase: A checkpoint to isoprenoid biosynthesis. Biochimie 2012, 94, 1621–1634. [Google Scholar] [CrossRef] [PubMed]
- Ladevèze, V.; Marcireau, C.; Delourme, D.; Karst, F. General resistance to sterol biosynthesis inhibitors in Saccharomyces cerevisiae. Lipids 1993, 28, 907–912. [Google Scholar] [CrossRef] [PubMed]
- Xiaohe, C.; Wenzhi, B.; Lijuan, L.; Haifeng, X. Excavation of metabolic pathways for increasing the production of isoprene in Saccharomyces cerevisiae. Food Ferment. Ind. 2020, 46, 1–7. [Google Scholar]
- Ignea, C.; Cvetkovic, I.; Loupassaki, S.; Kefalas, P.; Johnson, C.B.; Kampranis, S.C.; Makris, A.M. Improving yeast strains using recyclable integration cassettes, for the production of plant terpenoids. Microb. Cell Fact. 2011, 10, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Xia, F.; Du, J.; Wang, K.; Liu, L.; Ba, L.; Liu, H.; Liu, Y. Application of Multiple Strategies To Debottleneck the Biosynthesis of Longifolene by Engineered Saccharomyces cerevisiae. J. Agric. Food Chem. 2022, 70, 11336–11343. [Google Scholar] [CrossRef]
- Marsafari, M.; Xu, P. Debottlenecking mevalonate pathway for antimalarial drug precursor amorphadiene biosynthesis in Yarrowia lipolytica. Metab. Eng. Commun. 2020, 10, e00121. [Google Scholar] [CrossRef]
- Kwak, S.; Kim, S.R.; Xu, H.; Zhang, G.-C.; Lane, S.; Kim, H.; Jin, Y.-S. Enhanced isoprenoid production f rom xylose by engineered Saccharomyces cerevisiae. Biotechnol. Bioeng. 2017, 114, 2581–2591. [Google Scholar] [CrossRef]
- Kim, T.Y.; Park, H.; Kim, S.-K.; Kim, S.-J.; Park, Y.-C. Production of (−)-α-bisabolol in metabolically engineered Saccharomyces cerevisiae. J. Biotechnol. 2021, 340, 13–21. [Google Scholar] [CrossRef]
- Liu, H.; Marsafari, M.; Deng, L.; Xu, P. Understanding lipogenesis by dynamically profiling transcriptional activity of lipogenic promoters in Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2019, 103, 3167–3179. [Google Scholar] [CrossRef]
- Liu, H.; Wang, F.; Deng, L.; Xu, P. Genetic and bioprocess engineering to improve squalene production in Yarrowia lipolytica. Bioresour. Technol. 2020, 317, 123991. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Shi, F.; Zhan, W. Overexpression of ZWF1 and POS5 improves carotenoid biosynthesis in recombinant Saccharomyces cerevisiae. Lett. Appl. Microbiol. 2015, 61, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Kwak, S.; Yun, E.J.; Lane, S.; Oh, E.J.; Kim, K.H.; Jin, Y.-S. Redirection of the Glycolytic Flux Enhances Isoprenoid Production in Saccharomyces cerevisiae. Biotechnol. J. 2020, 15, 1900173. [Google Scholar] [CrossRef]
- Moreira dos Santos, M.; Thygesen, G.; Kötter, P.; Olsson, L.; Nielsen, J. Aerobic physiology of redox-engineered Saccharomyces cerevisiae strains modified in the ammonium assimilation for increased NADPH availability. FEMS Yeast Res. 2003, 4, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Asadollahi, M.A.; Maury, J.; Patil, K.R.; Schalk, M.; Clark, A.; Nielsen, J. Enhancing sesquiterpene production in Saccharomyces cerevisiae through in silico driven metabolic engineering. Metab. Eng. 2009, 11, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Boles, E.; Lehnert, W.; Zimmermann, F.K. The role of the NAD-dependent glutamate dehydrogenase in restoring growth on glucose of a Saccharomyces cerevisiae phosphoglucose isomerase mutant. Eur. J. Biochem. 1993, 217, 469–477. [Google Scholar] [CrossRef]
- Asadollahi, M.A.; Maury, J.; Schalk, M.; Clark, A.; Nielsen, J. Enhancement of farnesyl diphosphate pool as direct precursor of sesquiterpenes through metabolic engineering of the mevalonate pathway in Saccharomyces cerevisiae. Biotechnol. Bioeng. 2010, 106, 86–96. [Google Scholar]
- Scalcinati, G.; Knuf, C.; Partow, S.; Chen, Y.; Maury, J.; Schalk, M.; Daviet, L.; Nielsen, J.; Siewers, V. Combined metabolic engineering of precursor and co-factor supply to increase α-santalene production by Saccharomyces cerevisiae. Microb. Cell Fact. 2012, 11, 117. [Google Scholar] [CrossRef] [Green Version]
- Oswald, M.; Fischer, M.; Dirninger, N.; Karst, F. Monoterpenoid biosynthesis in Saccharomyces cerevisiae. FEMS Yeast Res. 2007, 7, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Ignea, C.; Pontini, M.; Maffei, M.E.; Makris, A.M.; Kampranis, S.C. Engineering Monoterpene Production in Yeast Using a Synthetic Dominant Negative Geranyl Diphosphate Synthase. ACS Synth. Biol. 2014, 3, 298–306. [Google Scholar] [CrossRef]
- Kildegaard, K.R.; Adiego-Pérez, B.; Doménech Belda, D.; Khangura, J.K.; Holkenbrink, C.; Borodina, I. Engineering of Yarrowia lipolytica for production of astaxanthin. Syn. Syst. Biotechnol. 2017, 2, 287–294. [Google Scholar] [CrossRef] [Green Version]
- Asadollahi, M.A.; Maury, J.; Møller, K.; Nielsen, K.F.; Schalk, M.; Clark, A.; Nielsen, J. Production of plant sesquiterpenes in Saccharomyces cerevisiae: Effect of ERG9 repression on sesquiterpene biosynthesis. Biotechnol. Bioeng. 2008, 99, 666–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Daviet, L.; Schalk, M.; Siewers, V.; Nielsen, J. Establishing a platform cell factory through engineering of yeast acetyl-CoA metabolism. Metab. Eng. 2013, 15, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Kozak Barbara, U.; van Rossum Harmen, M.; Luttik Marijke, A.H.; Akeroyd, M.; Benjamin, K.R.; Wu, L.; de Vries, S.; Daran, J.-M.; Pronk Jack, T.; van Maris Antonius, J.A. Engineering Acetyl Coenzyme A Supply: Functional Expression of a Bacterial Pyruvate Dehydrogenase Complex in the Cytosol of Saccharomyces cerevisiae. mBio 2014, 5, e01696-14. [Google Scholar]
- Shiba, Y.; Paradise, E.M.; Kirby, J.; Ro, D.-K.; Keasling, J.D. Engineering of the pyruvate dehydrogenase bypass in Saccharomyces cerevisiae for high-level production of isoprenoids. Metab. Eng. 2007, 9, 160–168. [Google Scholar] [CrossRef]
- Zhang, Q.; Zeng, W.; Xu, S.; Zhou, J. Metabolism and strategies for enhanced supply of acetyl-CoA in Saccharomyces cerevisiae. Bioresour. Technol. 2021, 342, 125978. [Google Scholar] [CrossRef]
- Malina, C.; Larsson, C.; Nielsen, J. Yeast mitochondria: An overview of mitochondrial biology and the potential of mitochondrial systems biology. FEMS Yeast Res. 2018, 18, 040. [Google Scholar] [CrossRef]
- Avalos, J.L.; Fink, G.R.; Stephanopoulos, G. Compartmentalization of metabolic pathways in yeast mitochondria improves the production of branched-chain alcohols. Nat. Biotechnol. 2013, 31, 335–341. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Ching, C.-B. Mitochondrial acetyl-CoA utilization pathway for terpenoid productions. Metab. Eng. 2016, 38, 303–309. [Google Scholar] [CrossRef]
- Lv, X.; Wang, F.; Zhou, P.; Ye, L.; Xie, W.; Xu, H.; Yu, H. Dual regulation of cytoplasmic and mitochondrial acetyl-CoA utilization for improved isoprene production in Saccharomyces cerevisiae. Nat. Commun. 2016, 7, 12851. [Google Scholar] [CrossRef] [Green Version]
- Yee, D.A.; DeNicola, A.B.; Billingsley, J.M.; Creso, J.G.; Subrahmanyam, V.; Tang, Y. Engineered mitochondrial production of monoterpenes in Saccharomyces cerevisiae. Metab. Eng. 2019, 55, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Farhi, M.; Marhevka, E.; Masci, T.; Marcos, E.; Eyal, Y.; Ovadis, M.; Abeliovich, H.; Vainstein, A. Harnessing yeast subcellular compartments for the production of plant terpenoids. Metab. Eng. 2011, 13, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Kohlwein, S.D.; Veenhuis, M.; van der Klei, I.J. Lipid droplets and peroxisomes: Key players in cellular lipid homeostasis or a matter of fat—store’em up or burn’em down. Genetics 2013, 193, 1–50. [Google Scholar] [CrossRef] [Green Version]
- Joshi, A.S.; Cohen, S. Biology, Lipid droplet and peroxisome biogenesis: Do they go hand-in-hand? Front. Cell Dev. Biol. 2019, 7, 92. [Google Scholar] [CrossRef] [Green Version]
- van der Klei, I.J.; Veenhuis, M. Yeast peroxisomes: Function and biogenesis of a versatile cell organelle. Trends Microbiol. 1997, 5, 502–509. [Google Scholar] [CrossRef]
- Saraya, R.; Veenhuis, M.; van der Klei, I.J. Peroxisomes as dynamic organelles: Peroxisome abundance in yeast. FEBS J. 2010, 277, 3279–3288. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.-S.; Li, T.; Zhou, W.; Jiang, M.; Tao, X.-Y.; Liu, M.; Zhao, M.; Ren, Y.-H.; Gao, B.; Wang, F.-Q.; et al. The yeast peroxisome: A dynamic storage depot and subcellular factory for squalene overproduction. Metab. Eng. 2020, 57, 151–161. [Google Scholar] [CrossRef]
- Dusséaux, S.; Wajn William, T.; Liu, Y.; Ignea, C.; Kampranis Sotirios, C. Transforming yeast peroxisomes into microfactories for the efficient production of high-value isoprenoids. Proc. Natl. Acad. Sci. USA 2020, 117, 31789–31799. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Shi, T.-Q.; Peng, Q.-Q.; Sun, X.-M.; Ji, X.-J.; Huang, H. Harnessing Yarrowia lipolytica Peroxisomes as a Subcellular Factory for α-Humulene Overproduction. J. Agric. Food Chem. 2021, 69, 13831–13837. [Google Scholar] [CrossRef]
- Zhu, J.; Schwartz, C.; Wheeldon, I. Controlled intracellular trafficking alleviates an expression bottleneck in S. cerevisiae ester biosynthesis. Metab. Eng. Commun. 2019, 8, e00085. [Google Scholar] [CrossRef]
- Ma, T.; Shi, B.; Ye, Z.; Li, X.; Liu, M.; Chen, Y.; Xia, J.; Nielsen, J.; Deng, Z.; Liu, T. Lipid engineering combined with systematic metabolic engineering of Saccharomyces cerevisiae for high-yield production of lycopene. Metab. Eng. 2019, 52, 134–142. [Google Scholar] [CrossRef] [PubMed]
- de Ruijter, J.C.; Koskela, E.V.; Frey, A.D. Enhancing antibody folding and secretion by tailoring the Saccharomyces cerevisiae endoplasmic reticulum. Microb. Cell Fact. 2016, 15, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arendt, P.; Miettinen, K.; Pollier, J.; De Rycke, R.; Callewaert, N.; Goossens, A. An endoplasmic reticulum-engineered yeast platform for overproduction of triterpenoids. Metab. Eng. 2017, 40, 165–175. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Terpenes | Applications | Market Value | Current Production by Microbes (g/L) | Reference |
|---|---|---|---|---|
| Squalene | Pharmaceuticals, health products, and vaccine additives | 241.9 million USD | 21.2 | [6,7,8,9] |
| Limonene | Food additives, pharmaceuticals, cosmetics, biomaterials, and biofuels | 1.9 billion USD | 2.23 | [10,11,12,13,14,15] |
| Farnesene | Food additives, pharmaceuticals, cosmetics, biofuels | 315.8 million USD | 130 | [16,17,18,19,20] |
| β-carotene | Food additives, health products, pharmaceutical, and cosmetics | 300 billion USD | 39.5 | [21,22,23,24] |
| Astaxanthin | Health products cosmetics, and pharmaceuticals | 663.89 million USD | 1.18 | [25,26,27] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Liu, H.; Jin, Y.; Liu, J.; Deng, L.; Wang, F. Recent Advances in Multiple Strategies for the Synthesis of Terpenes by Engineered Yeast. Fermentation 2022, 8, 615. https://doi.org/10.3390/fermentation8110615
Yang L, Liu H, Jin Y, Liu J, Deng L, Wang F. Recent Advances in Multiple Strategies for the Synthesis of Terpenes by Engineered Yeast. Fermentation. 2022; 8(11):615. https://doi.org/10.3390/fermentation8110615
Chicago/Turabian StyleYang, Limeng, Huan Liu, Yuhan Jin, Junfeng Liu, Li Deng, and Fang Wang. 2022. "Recent Advances in Multiple Strategies for the Synthesis of Terpenes by Engineered Yeast" Fermentation 8, no. 11: 615. https://doi.org/10.3390/fermentation8110615