
Cultivation of Inonotus hispidus in Stirred Tank and Wave Bag Bioreactors to Produce the Natural Colorant Hispidin
 , , and
, , and
Abstract
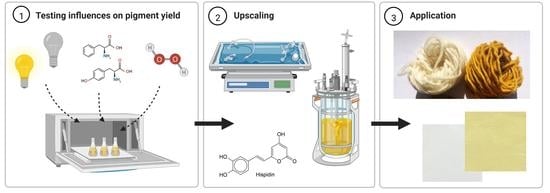
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cultivation Parameters and Media
2.2. Investigation of Different Influences on Pigment Yield
2.3. Fermentation Experiments
2.4. Analytics
2.5. Application of Cultivation Extracts
3. Results
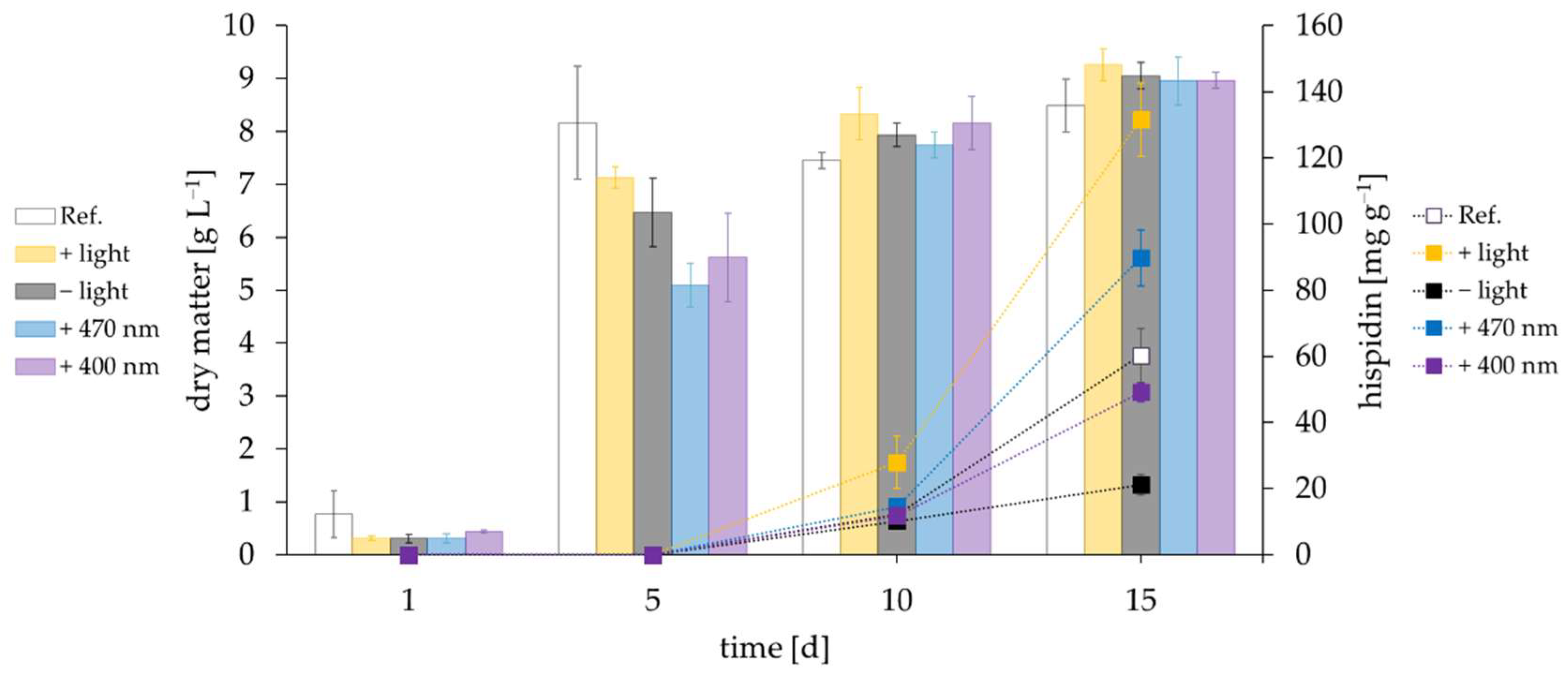
3.1. Influence of Light
3.2. Influence of Oxidative Stress
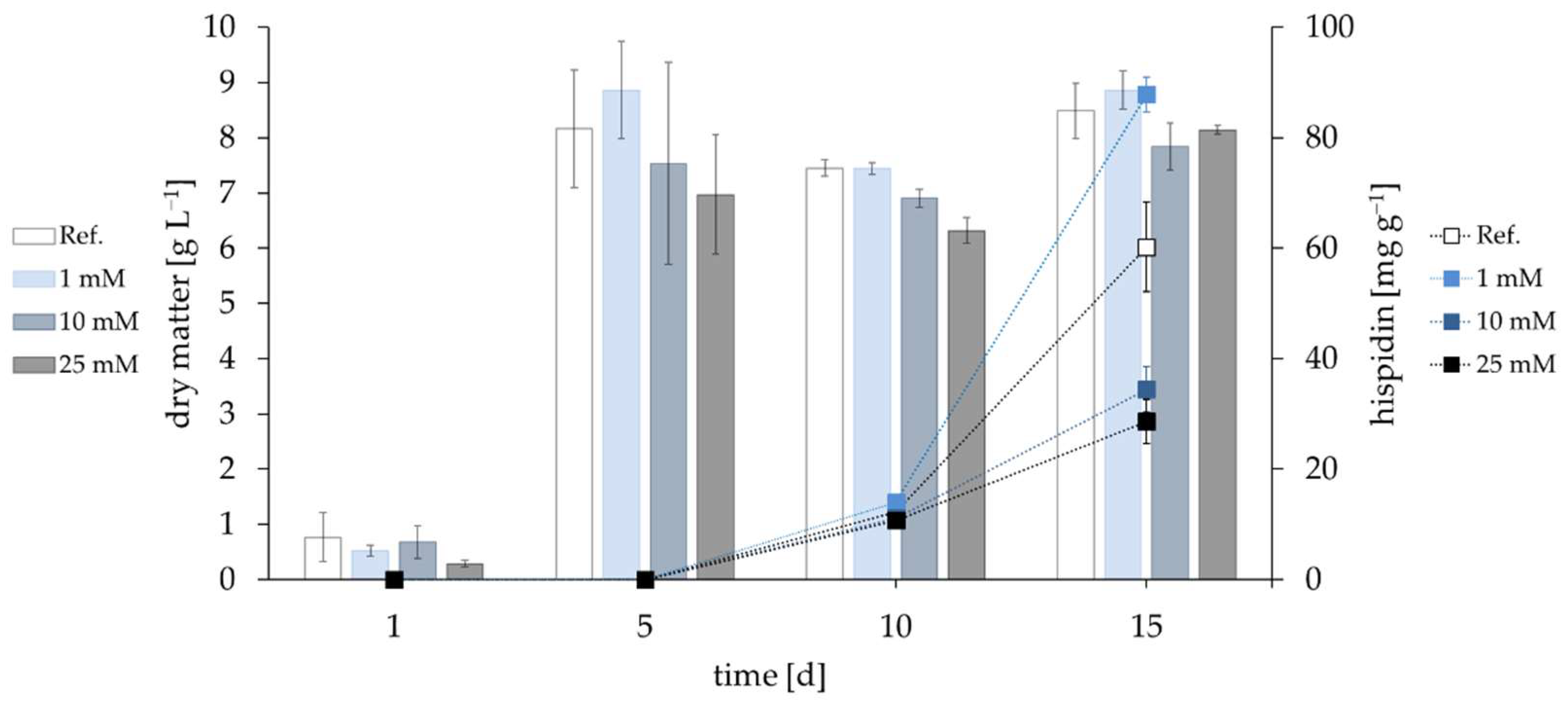
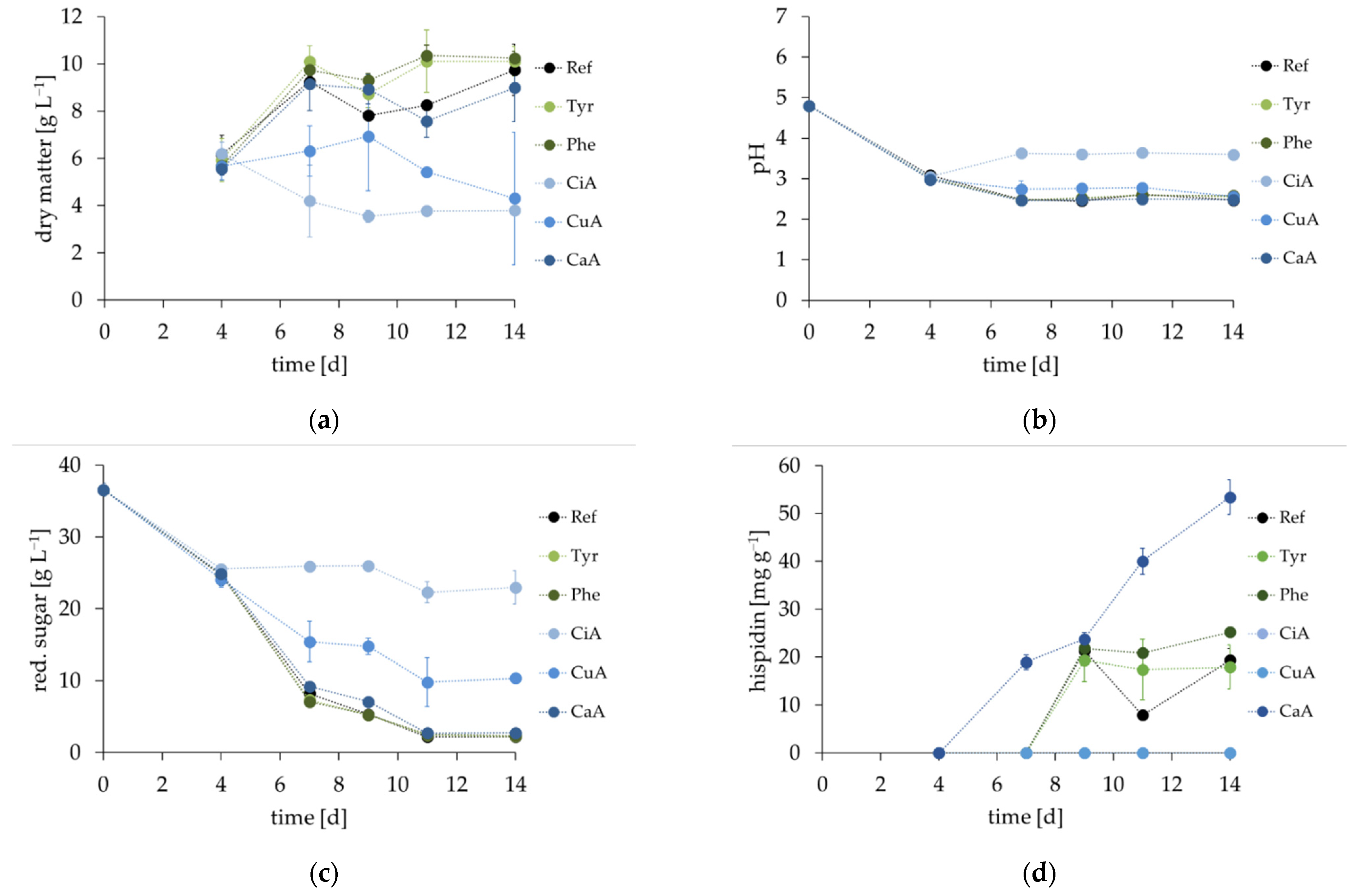
3.3. Influence of Precursors
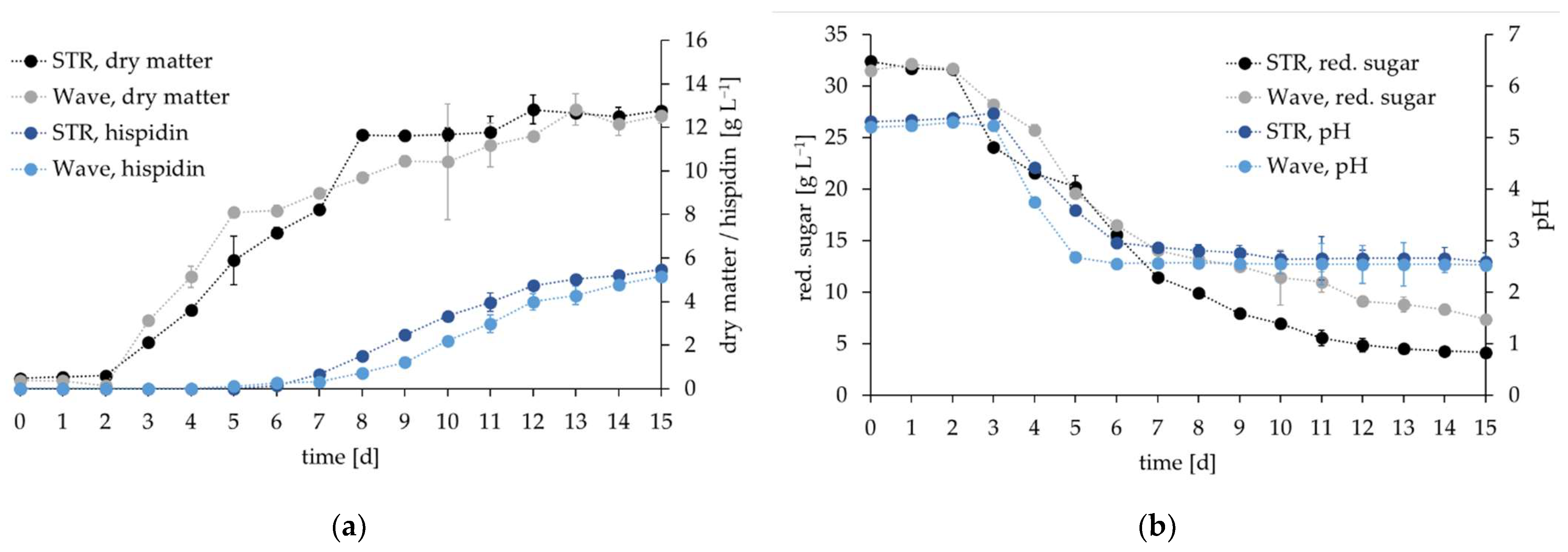
3.4. Comparative Cultivation in Two Types of Bioreactors
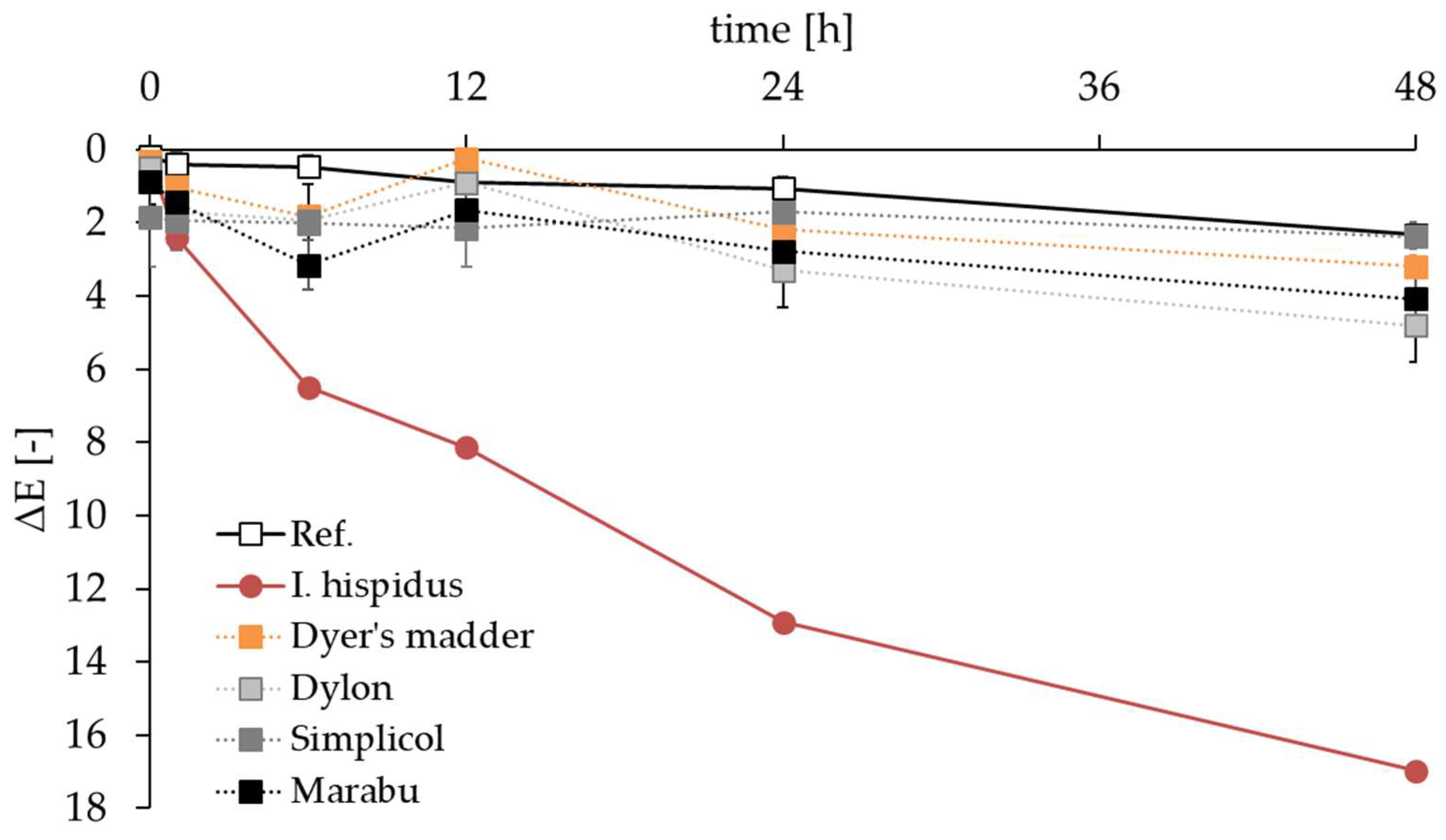
3.5. Application of Hispidin as a Colorant
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palkina, K.A.; Ipatova, D.A.; Shakhova, E.S.; Balakireva, A.V.; Markina, N.M. Therapeutic Potential of Hispidin-Fungal and Plant Polyketide. J. Fungi 2021, 7, 323. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.-K.; Yun, B.-S. Styrylpyrone-class compounds from medicinal fungi Phellinus and Inonotus spp., and their medicinal importance. J. Antibiot. 2011, 64, 349–359. [Google Scholar] [CrossRef]
- Angelini, P.; Girometta, C.; Tirillini, B.; Moretti, S.; Covino, S.; Cipriani, M.; D’Ellena, E.; Angeles, G.; Federici, E.; Savino, E.; et al. A comparative study of the antimicrobial and antioxidant activities of Inonotus hispidus fruit and their mycelia extracts. Int. J. Food Prop. 2019, 22, 768–783. [Google Scholar] [CrossRef] [Green Version]
- Gründemann, C.; Arnhold, M.; Meier, S.; Bäcker, C.; Garcia-Käufer, M.; Grunewald, F.; Steinborn, C.; Klemd, A.M.; Wille, R.; Huber, R.; et al. Effects of Inonotus hispidus Extracts and Compounds on Human Immunocompetent Cells. Planta Med. 2016, 82, 1359–1367. [Google Scholar] [CrossRef]
- Smolskaitė, L.; Venskutonis, P.R.; Talou, T. Comprehensive evaluation of antioxidant and antimicrobial properties of different mushroom species. LWT—Food Sci. Technol. 2015, 60, 462–471. [Google Scholar] [CrossRef] [Green Version]
- El Hassane, A.; Shah, S.A.A.; Hassan, N.B.; El Moussaoui, N.; Ahmad, R.; Zulkefeli, M.; Weber, J.-F.F. Antioxidant activity of hispidin oligomers from medicinal fungi: A DFT study. Molecules 2014, 19, 3489–3507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu’Lock, J.D.; Leeming, P.R.; Smith, H.G. 400. Pyrones. Part II. Hispidin, a new pigment and precursor of a fungus “lignin”. J. Chem. Soc. 1962, 2085–2089. [Google Scholar] [CrossRef]
- Park, I.-H.; Chung, S.-K.; Lee, K.-B.; Yoo, Y.-C.; Kim, S.-K.; Kim, G.-S.; Song, K.-S. An antioxidant hispidin from the mycelial cultures of Phellinus linteus. Arch. Pharm. Res. 2004, 27, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.C.; Tudor, D.; Zhang, W.R.; Ng, S.; Cooper, P.A. Ability of three yellow pigment producing fungi to colour wood under controlled conditions. Int. Wood Prod. J. 2014, 5, 103–107. [Google Scholar] [CrossRef]
- Friese, W.; Wähnert, V. Einfach färben mit Pilzen und Pflanzen; Freya Verlag GmbH: Engerwitzdorf, Austria, 2021; ISBN 9783990254141. [Google Scholar]
- Sundström, C.; Sundström, E. Mit Pilzen färben: Eine Fundgrube für Kunstgewerbler, Pilzsammler und Naturfreunde; Orell Füssli: Zürich, Switzerland; Schwäbisch Hall, Germany, 1984; ISBN 3280014735. [Google Scholar]
- Tegeler, K.; Deutsche Gesellschaft für Mykologie e. V. Leitfaden zum Färben mit Pilzen, 2nd ed.; Josef Maria Christan: München, Germany, 2016. [Google Scholar]
- Nambudiri, A.M.D.; Vance, C.P.; Towers, G.H.N. Styrylpyrone biosynthesis in Polyporus hispidus: II. Enzymic hydroxylation of p-coumaric acid and bis-noryangonin. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1974, 343, 148–155. [Google Scholar] [CrossRef]
- Velíšek, J.; Cejpek, K. Pigments of higher fungi—A review. Czech J. Food Sci. 2011, 29, 87–102. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.-Y.; Liu, J.-K. Pigments of fungi (macromycetes). Nat. Prod. Rep. 2010, 27, 1531–1570. [Google Scholar] [CrossRef] [PubMed]
- Perrin, P.W.; Towers, G. Hispidin biosynthesis in cultures of Polyporus hispidus. Phytochemistry 1973, 12, 589–592. [Google Scholar] [CrossRef]
- Zheng, W.; Zhao, Y.; Zhang, M.; Wei, Z.; Miao, K.; Sun, W. Oxidative stress response of Inonotus obliquus induced by hydrogen peroxide. Med. Mycol. 2009, 47, 814–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nambudiri, A.M.D.; Vance, C.P.; Towers, G.H.N. Effect of light on enzymes of phenylpropanoid metabolism and hispidin biosynthesis in Polyporus hispidus. Biochem. J. 1973, 134, 891–897. [Google Scholar] [CrossRef] [Green Version]
- Vance, C.P.; Tregunna, E.B.; Nambudiri, A.M.D.; Towers, G.H.N. Styrylpyrone biosynthesis in Polyporus hispidus: I. Action spectrum and photoregulation of pigment and enzyme formation. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1974, 343, 138–147. [Google Scholar] [CrossRef]
- Huang, S.-Y.; Chang, S.-F.; Chau, S.-F.; Chiu, S.-C. The Protective Effect of Hispidin against Hydrogen Peroxide-Induced Oxidative Stress in ARPE-19 Cells via Nrf2 Signaling Pathway. Biomolecules 2019, 9, 380. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.S.; Lee, J.S.; Lee, J.H.; Kwon, D.S.; Lee, K.E.; Lee, S.Y.; Hong, E.K. Hispidin produced from Phellinus linteus protects pancreatic beta-cells from damage by hydrogen peroxide. Arch. Pharm. Res. 2010, 33, 853–861. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, J.S.; Kim, Y.R.; Jung, W.C.; Lee, K.E.; Lee, S.Y.; Hong, E.K. Hispidin isolated from Phellinus linteus protects against hydrogen peroxide-induced oxidative stress in pancreatic MIN6N β-cells. J. Med. Food 2011, 14, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Jonczyk, P.; Takenberg, M.; Hartwig, S.; Beutel, S.; Berger, R.G.; Scheper, T. Cultivation of shear stress sensitive microorganisms in disposable bag reactor systems. J. Biotechnol. 2013, 167, 370–376. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. ISBN 9780123721808. [Google Scholar]
- Kavitha, S.; Chandra, T.S. Oxidative stress protection and glutathione metabolism in response to hydrogen peroxide and menadione in riboflavinogenic fungus Ashbya gossypii. Appl. Biochem. Biotechnol. 2014, 174, 2307–2325. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Rottmann, E.; Hauke, K.F.; Krings, U.; Berger, R.G. Enzymatic acrylamide mitigation in French fries—An industrial-scale case study. Food Control 2021, 123, 107739. [Google Scholar] [CrossRef]
- Takeda, J.; Ozeki, Y.; Yoshida, K. Action spectrum for induction of promoter activity of phenylalanine ammonia-lyase gene by UV in carrot suspension cells. Photochem. Photobiol. 1997, 66, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Gornostai, T.G.; Borovskii, G.G.; Kashchenko, N.I.; Olennikov, D.N. Phenolic Compounds of Inonotus rheades (Agaricomycetes) Mycelium: RP-UPLC-DAD-ESI/MS Profile and Effect of Light Wavelength on Styrylpyrone Content. Int. J. Med. Mushrooms 2018, 20, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Oba, Y.; Suzuki, Y.; Martins, G.N.R.; Carvalho, R.P.; Pereira, T.A.; Waldenmaier, H.E.; Kanie, S.; Naito, M.; Oliveira, A.G.; Dörr, F.A.; et al. Identification of hispidin as a bioluminescent active compound and its recycling biosynthesis in the luminous fungal fruiting body. Photochem. Photobiol. Sci. 2017, 16, 1435–1440. [Google Scholar] [CrossRef] [PubMed]
- Purtov, K.V.; Petushkov, V.N.; Baranov, M.S.; Mineev, K.S.; Rodionova, N.S.; Kaskova, Z.M.; Tsarkova, A.S.; Petunin, A.I.; Bondar, V.S.; Rodicheva, E.K.; et al. The Chemical Basis of Fungal Bioluminescence. Angew. Chem. Int. Ed Engl. 2015, 54, 8124–8128. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.-H.; Lee, Y.-M.; Park, S.R.; Da Kim, H.; Lim, B.O. Anticancer activity of hispidin via reactive oxygen species-mediated apoptosis in colon cancer cells. Anticancer. Res. 2014, 34, 4087–4093. [Google Scholar]
- Liang, C.-H.; Wu, C.-Y.; Li, P.-H.; Liang, Z.-C. Optimal Liquid Inoculum Conditions and Grain Medium Enhanced Hispidin Production by Species of Genus Phellinus (Agaricomycetes) in Solid-State Fermentation. Int. J. Med. Mushrooms 2022, 24, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Li, I.-C.; Chen, C.C.; Sheu, S.-J.; Huang, I.-H.; Chen, C.-C. Optimized production and safety evaluation of hispidin-enriched Sanghuangporus sanghuang mycelia. Food Sci. Nutr. 2020, 8, 1864–1873. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bergmann, P.; Takenberg, M.; Frank, C.; Zschätzsch, M.; Werner, A.; Berger, R.G.; Ersoy, F. Cultivation of Inonotus hispidus in Stirred Tank and Wave Bag Bioreactors to Produce the Natural Colorant Hispidin. Fermentation 2022, 8, 541. https://doi.org/10.3390/fermentation8100541
Bergmann P, Takenberg M, Frank C, Zschätzsch M, Werner A, Berger RG, Ersoy F. Cultivation of Inonotus hispidus in Stirred Tank and Wave Bag Bioreactors to Produce the Natural Colorant Hispidin. Fermentation. 2022; 8(10):541. https://doi.org/10.3390/fermentation8100541
Chicago/Turabian StyleBergmann, Pia, Meike Takenberg, Christina Frank, Marlen Zschätzsch, Anett Werner, Ralf G. Berger, and Franziska Ersoy. 2022. "Cultivation of Inonotus hispidus in Stirred Tank and Wave Bag Bioreactors to Produce the Natural Colorant Hispidin" Fermentation 8, no. 10: 541. https://doi.org/10.3390/fermentation8100541