
Microbial Astaxanthin Production from Agro-Industrial Wastes—Raw Materials, Processes, and Quality
 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
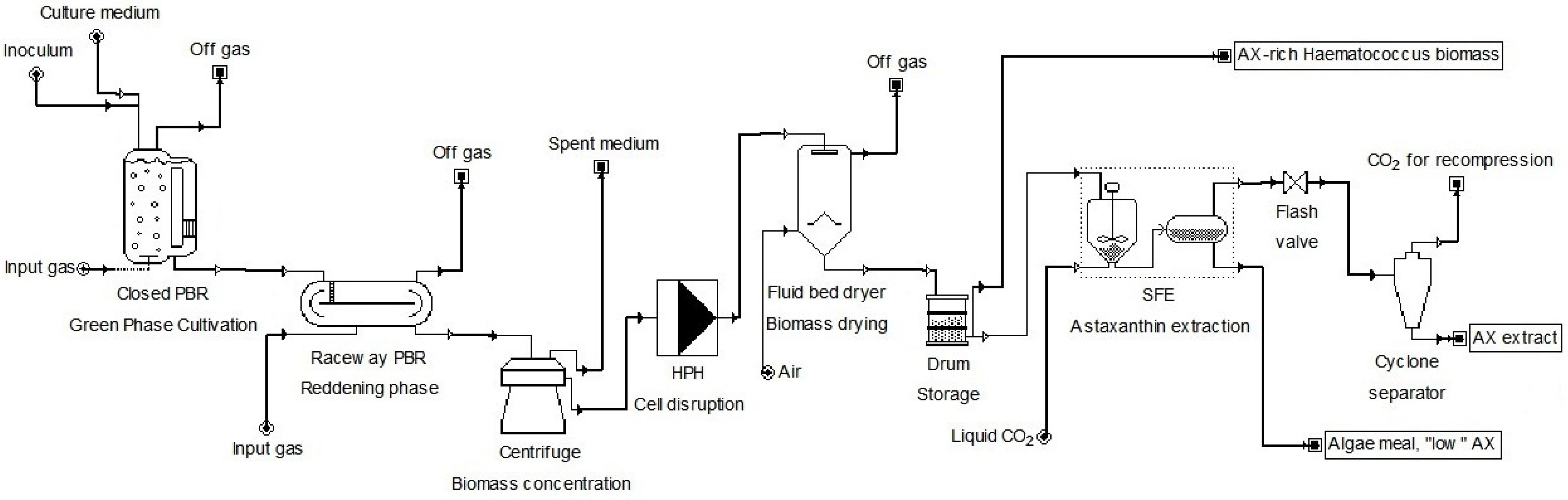
1.1. Current Production
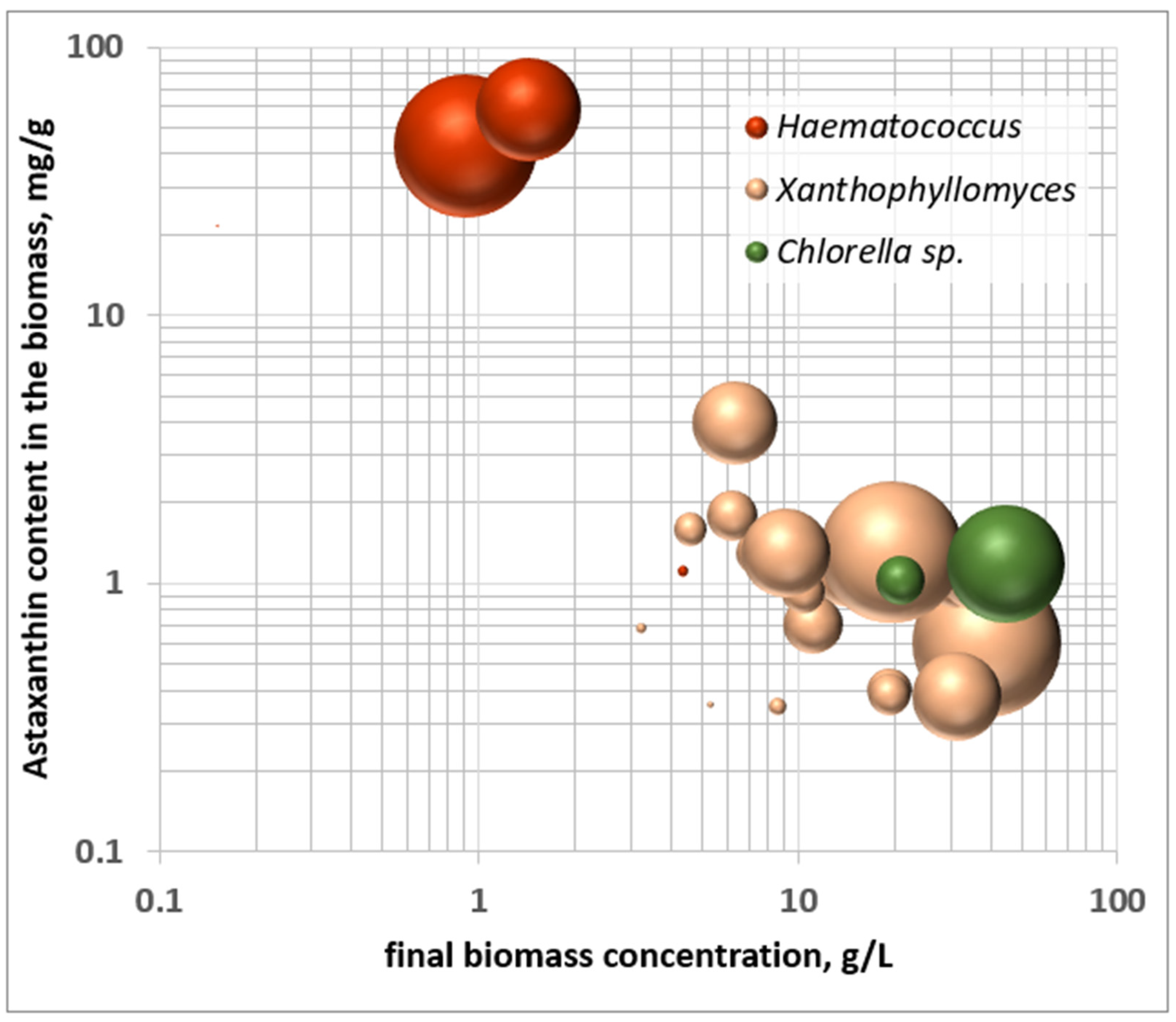
1.2. The Importance of Intracellular Concentration
- (a)
- 25 kg of Haematococcus with 40 mg/g of astaxanthin, which would translate into 25 m3 of culture harvested each day, with a concentration of 1 g/L biomass.
- (b)
- 1 ton of Xanthophyllomyces sp. with 1 mg/g of astaxanthin, which requires a volume of fermentation of 50 m3 for a culture reaching 20 g/L of yeast biomass.
1.3. Culture Media Costs: Nutrients and Water
2. Microbial AX Production from Agro-Industry Wastes
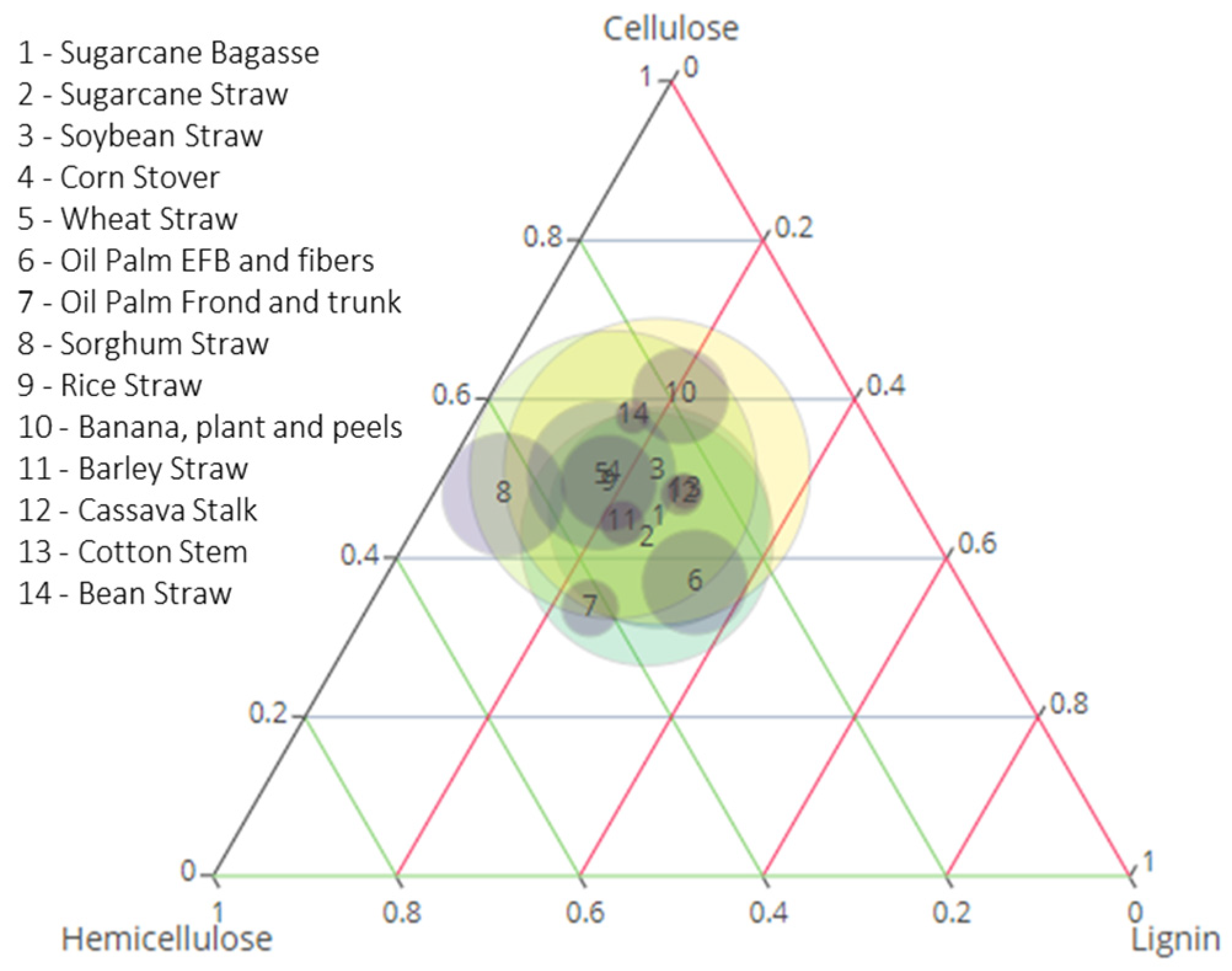
2.1. Production and Productivity in Residues

{kind=link}
{kind=link}
{kind=link}
| Ref. | Microorganism | AX Content, mg·g−1 | Time, Days | Biomass, g·L−1 | Volumetric AX Productivity, mg·L−1·d−1 | Waste/Residue |
|---|---|---|---|---|---|---|
| [35] | Chlorella protothecoides | 0.096 | 7 | 3.6 | 0.049 | Ricotta production waste, scotta |
| [36] | Chlorella zofingiensis | 1.03 | 10 | 21 | 2.163 | Pretreated cane molasses |
| 1.19 | 10 | 44.6 | 5.307 | |||
| [37] | Haematococcus pluvialis | 43 | 6 | 0.906 | 6.493 | Minkery wastewater |
| [28] | 1.11 | 11 | 4.37 | 0.441 | Bioethanol plant wastewater | |
| [27] | 0.4 | 9 | 0.7 | 0.031 | ||
| [38] | 0.011 | 13 | 0.97 | 0.001 | Swine slurry | |
| [39] | 21.59 | 35 | 0.151 | 0.093 | Domestic wastewater | |
| [29] | 58.7 | 18 | 1.43 | 4.663 | Treated piggery wastewater | |
| [40] | 98 | 8 | Synthetic medium (for reference) | |||
| [41] | Paracoccus NBRC 101723 | 0.963 | 5 | Synthetic medium (for reference) | ||
| [42] | Thraustochytrium striatum | 0.6 | 8 | 4.2 | 0.315 | Corn stover hydrolysate |
| [43] | Xanthophyllomyces dendrorhous | 1.448 | 7 | 9.21 | 1.905 | Low-cost agro products |
| [44] | 0.35 | 4 | 8.6 | 0.753 | Cassava residues substrate | |
| [45] | 0.355 | 7 | 5.3 | 0.269 | Fruit and vegetable waste | |
| [46] | 3.962 | 2 | 6.35 | 12.579 | Mussel processing wastewater | |
| [30] | 1.6 | 5 | 4.6 | 1.472 | Peat hydrolysate | |
| 0.4 | 4 | 19.3 | 1.930 | Sugar cane juice and urea | ||
| 1.3 | 5 | 7.5 | 1.950 | Grape juice | ||
| 1.8 | 5 | 6.2 | 2.232 | Coconut milk | ||
| 1.1 | 5 | 14.1 | 3.102 | Molasses | ||
| 0.6 | 3.5 | 39 | 6.686 | Date juice | ||
| 1.3 | 4 | 19.6 | 6.370 | Mustard waste isolates | ||
| 1 | 14 | 30.6 | 2.186 | Eucalyptus hydrolysate | ||
| 1.1 | 12 | 36 | 3.300 | Sugar beet molasses | ||
| [47] | 1.141 | 6 | 18.43 | 3.505 | Citrus waste isolates | |
| [48] | 1.31 | 4 | 19.6 | 6.419 | Mustard waste isolates | |
| [34] | 0.03 | 7 | 7.5 | 0.032 | Enzymatic hydrolysates of pre-hydrolyzed wood | |
| [49] | 0.68 | 5 | 3.23 | 0.439 | Thai traditional rice vermicelli plant | |
| 0.93 | 5 | 10.3 | 1.916 | |||
| [33] | 4 | 5.750 | Barley straw | |||
| [50] | 0.018 | 5 | 5.5 | 0.020 | Residual-brewery yeast extract | |
| [31] | 0.383 | 4 | 19.35 | 1.853 | Sugarcane juice | |
| [32] | 0.38 | 3 | 31.4 | 3.977 | ||
| [51] | 1.3 | 3 | 9.2 | 3.987 | Vinasse supplementation of sugarcane juice | |
| [52] | 0.702 | 3 | 11.16 | 2.611 | Synthetic medium (for reference) |
2.2. Transport, Handling, and Pretreatment of Agro-Industry Wastes
2.3. Products and Quality Control
3. Trends and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kumar, S.; Kumar, R.; Diksha; Kumari, A.; Panwar, A. Astaxanthin: A super antioxidant from microalgae and its therapeutic potential. J. Basic Microbiol. 2021, 62, 1064–1082. [Google Scholar] [CrossRef] [PubMed]
- Naguib, Y.M.A. Antioxidant activities of astaxanthin and related carotenoids. J. Agric. Food Chem. 2000, 48, 1150–1154. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Sintara, M.; Chang, T. Multi-radical (ORAC MR5) antioxidant capacity of selected berries and effects of food processing. J. Berry Res. 2016, 6, 159–173. [Google Scholar] [CrossRef]
- Ahuja, K.; Rawat, A. Astaxanthin Market Size by Source (Synthetic, Natural), by Application (Dietary Supplement, Personal Care, Pharmaceuticals, Food & Beverages, Animal Feed {Aquaculture, Livestock, Pets}) Industry Outlook Report, Regional Analysis, Application Potential, Price Trends, Competitive Market Share & Forecast, 2019–2026; Global Market Insights: Selbyville, DE, USA, 2019. [Google Scholar]
- Markets&Markets. Astaxanthin Market Share, Size (2021–2026) MarketsandMarkets. 2021. Available online: https://www.marketsandmarkets.com/Market-Reports/astaxanthin-market-162119410.html?gclid=Cj0KCQjwuuKXBhCRARIsAC-gM0gvdQQSiFavi31XlrEtigZRktVejiXvdSvI7txY42Wxdv-571UGEgMaAnjGEALw_wcB (accessed on 13 August 2022).
- Grand View Research. Astaxanthin Market Size|Industry Report, 2021–2028. Astaxanthin Market Size, Share & Trends Report 2021–2028; Grand View Research: San Francisco, CA, USA, 2021; Available online: https://www.grandviewresearch.com/industry-analysis/global-astaxanthin-market (accessed on 13 August 2022).
- Jannel, S.; Caro, Y.; Bermudes, M.; Petit, T. Novel insights into the biotechnological production of Haematococcus pluvialis-derived astaxanthin: Advances and key challenges to allow its industrial use as novel food ingredient. J. Mar. Sci. Eng. 2020, 8, 789. [Google Scholar] [CrossRef]
- Angell, A.; de Nys, R.; Mangott, A.; Vucko, M.J. The effects of concentration and supplementation time of natural and synthetic sources of astaxanthin on the colouration of the prawn Penaeus monodon. Algal Res. 2018, 35, 577–585. [Google Scholar] [CrossRef]
- Su, F.; Yu, W.; Liu, J. Comparison of effect of dietary supplementation with Haematococcus pluvialis powder and synthetic astaxanthin on carotenoid composition, concentration, esterification degree and astaxanthin isomers in ovaries, hepatopancreas, carapace, epithelium of adult female Chinese mitten crab (Eriocheir sinensis). Aquaculture 2020, 523, 735146. [Google Scholar] [CrossRef]
- Aneesh, P.A.; Ajeeshkumar, K.K.; Lekshmi, R.G.K.; Anandan, R.; Ravishankar, C.N.; Mathew, S. Bioactivities of astaxanthin from natural sources, augmenting its biomedical potential: A review. Trends Food Sci. Technol. 2022, 125, 81–90. [Google Scholar] [CrossRef]
- Capelli, B.; TalbFott, S.; Ding, L. Astaxanthin Sources: Suitability for Human Health and Nutrition. Funct. Foods Health Dis. 2019, 9, 430–445. Available online: https://www.ffhdj.com/index.php/ffhd/article/view/584/1139 (accessed on 13 August 2022). [CrossRef]
- De Carvalho, J.C.; Cardoso, L.C.; Ghiggi, V.; Woiciechowski, A.L.; de Souza Vandenberghe, L.P.; Soccol, C.R. Microbial pigments. In Biotransformation of Waste Biomass into High Value Biochemicals; Springer: New York, NY, USA, 2014; pp. 73–97. ISBN 9781461480051. [Google Scholar] [CrossRef]
- Stachowiak, B.; Szulc, P. Astaxanthin for the Food Industry. Molecules 2021, 26, 2666. [Google Scholar] [CrossRef]
- García-Vaquero, M.; Brunton, N.; Lafarga, T. Microalgae as a source of pigments for food applicati ons. In Cultured Microalgae for the Food Industry: Current and Potential Applications; Academic Press: Cambridge, MA, USA, 2021; pp. 177–198. [Google Scholar] [CrossRef]
- Oslan, S.N.H.; Shoparwe, N.F.; Yusoff, A.H.; Rahim, A.A.; Chang, C.S.; Tan, J.S.; Oslan, S.N.; Arumugam, K.; Ariff, A.B.; Sulaiman, A.Z.; et al. A Review on Haematococcus pluvialis Bioprocess Optimization of Green and Red Stage Culture Conditions for the Production of Natural Astaxanthin. Biomolecules 2021, 11, 256. [Google Scholar] [CrossRef]
- De Carvalho, J.C.; Magalhães, A.I.; de Melo Pereira, G.V.; Medeiros, A.B.P.; Sydney, E.B.; Rodrigues, C.; Aulestia, D.T.M.; de Souza Vandenberghe, L.P.; Soccol, V.T.; Soccol, C.R. Microalgal biomass pretreatment for integrated processing into biofuels, food, and feed. Bioresour. Technol. 2020, 300, 122719. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.; Mahfuzur, R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-producing green microalga Haematococcus pluvialis: From single cell to high value commercial products. Front. Plant Sci. 2016, 7, 531. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.N.; Ngo, D.-H.; Tran, Q.T.; Nguyen, H.C.; Su, C.-H.; Ngo, D.-N. Enhancing Astaxanthin Biosynthesis by Rhodosporidium toruloides Mutants and Optimization of Medium Compositions Using Response Surface Methodology. Processes 2020, 8, 497. [Google Scholar] [CrossRef]
- Colusse, G.A.; Duarte, M.E.R.; de Carvalho, J.C.; Noseda, M.D. Media effects on laboratory scale production costs of Haematococcus pluvialis biomass. Bioresour. Technol. Rep. 2019, 7, 100236. [Google Scholar] [CrossRef]
- Williams, P.; Rickards, S. Waterfund/IBM the true cost of water. In ICSI 2014: Creating Infrastructure for a Sustainable World; American Society of Civil Engineers: Reston, VA, USA, 2014; pp. 54–68. [Google Scholar]
- Magalhães, A.I., Jr.; de Carvalho, J.C.; de Carvalho, N.D.P.; Soccol, C.R. Are Sugarcane Molasses Competitive Substrates for Bio-based Platform Chemicals? J. Agric. Food Chem. 2020, 68, 4073. [Google Scholar] [CrossRef]
- Colusse, G.A.; Santos, A.O.; Rodrigues, J.M.; Barga, M.C.; Duarte, M.E.R.; de Carvalho, J.C.; Noseda, M.D. Rice vinasse treatment by immobilized Synechococcus pevalekii and its effect on Dunaliella salina cultivation. Bioprocess Biosyst. Eng. 2021, 44, 1477–1490. [Google Scholar] [CrossRef]
- De Carvalho, J.C.; Borghetti, I.A.; Cartas, L.C.; Woiciechowski, A.L.; Soccol, V.T.; Soccol, C.R. Biorefinery integration of microalgae production into cassava processing industry: Potential and perspectives. Bioresour. Technol. 2018, 247, 1165–1172. [Google Scholar] [CrossRef]
- Magalhães, A.I.; Carvalho, J.C.; Melo Pereira, G.V.; Karp, S.G.; Câmara, M.C.; Medina, J.D.C.; Soccol, C.R. Lignocellulosic biomass from agro-industrial residues in South America: Current developments and perspectives. Biofuels Bioprod. Biorefin. 2019, 13, 1505–1519. [Google Scholar] [CrossRef]
- De Carvalho, J.C.; Sydney, E.B.; Tessari, L.F.A.; Soccol, C.R. Culture media for mass production of microalgae. In Biofuels from Algae, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 33–50. [Google Scholar] [CrossRef]
- Tessari, L.F.A.; Soccol, C.R.; Rodrigues, C.; González, E.G.; Tanobe, V.O.d.A.; Kirnev, P.C.d.S.; de Carvalho, J.C. Development of a Culture Medium for Microalgae Production Based on Minimal Processing of Oil Palm Biomass Ash. Fermentation 2022, 8, 55. [Google Scholar] [CrossRef]
- Haque, F.; Dutta, A.; Thimmanagari, M.; Chiang, Y.W. Intensified green production of astaxanthin from Haematococcus pluvialis. Food Bioprod. Process. 2016, 99, 1–11. [Google Scholar] [CrossRef]
- Haque, F.; Dutta, A.; Thimmanagari, M.; Chiang, Y.W. Integrated Haematococcus pluvialis biomass production and nutrient removal using bioethanol plant waste effluent. Process Saf. Environ. Prot. 2017, 111, 128–137. [Google Scholar] [CrossRef]
- Kang, C.D.; An, J.Y.; Park, T.H.; Sim, S.J. Astaxanthin biosynthesis from simultaneous N and P uptake by the green alga Haematococcus pluvialis in primary-treated wastewater. Biochem. Eng. J. 2006, 31, 234–238. [Google Scholar] [CrossRef]
- Rodríguez-Sáiz, M.; de la Fuente, J.L.; Barredo, J.L. Xanthophyllomyces dendrorhous for the industrial production of astaxanthin. Appl. Microbiol. Biotechnol. 2010, 88, 645–658. [Google Scholar] [CrossRef] [PubMed]
- Moriel, D.G.; Chociai, M.B.; Machado, I.M.P.; Fontana, J.D.; Bonfim, T.M.B. Effect of feeding methods on the astaxanthin production by Phaffia rhodozyma in fed-batch process. Braz. Arch. Biol. Technol. 2005, 48, 397–401. [Google Scholar] [CrossRef]
- Moriel, D.G.; Machado, I.M.P.; Fontana, J.D.; Bonfim, T.M.B. Optimization of biomass and astaxanthin production by the yeast Phaffia rhodozyma. Rev. Bras. Ciênc. Farm. 2004, 40, 421–424. [Google Scholar] [CrossRef]
- Nghiem, N.P.; Kim, T.H.; Yoo, C.G.; Hicks, K.B. Enzymatic Fractionation of SAA-Pretreated Barley Straw for Production of Fuel Ethanol and Astaxanthin as a Value-Added Co-Product. Appl. Biochem. Biotechnol. 2013, 171, 341–351. [Google Scholar] [CrossRef]
- Parajo, J.C.; Santos, V.; Vazquez, M.; Cruz, J.M.; Parajó, J.C.; Santos, V.; Vázquez, M.; Cruz, J.M. Production of carotenoids by Xanthophyllomyces dendrorhous growing on enzymatic hydrolysates of prehydrolysed wood. Food Chem. 1997, 60, 347–355. [Google Scholar] [CrossRef]
- Ribeiro, J.E.S.; Martini, M.; Altomonte, I.; Salari, F.; Nardoni, S.; Sorce, C.; da Silva, F.L.H.; Andreucci, A. Production of Chlorella protothecoides biomass, chlorophyll and carotenoids using the dairy industry by-product scotta as a substrate. Biocatal. Agric. Biotechnol. 2017, 11, 207–213. [Google Scholar] [CrossRef]
- Liu, J.; Sun, Z.; Zhong, Y.; Gerken, H.; Huang, J.; Chen, F. Utilization of cane molasses towards cost-saving astaxanthin production by a Chlorella zofingiensis mutant. J. Appl. Phycol. 2013, 25, 1447–1456. [Google Scholar] [CrossRef]
- Liu, Y.; Yildiz, I. Bioremediation of minkery wastewater and astaxanthin production by Haematococcus pluvialis. Int. J. Glob. Warm. 2019, 19, 145–157. [Google Scholar] [CrossRef]
- Ledda, C.; Tamiazzo, J.; Borin, M.; Adani, F. A simplified process of swine slurry treatment by primary filtration and Haematococcus pluvialis culture to produce low cost astaxanthin. Ecol. Eng. 2016, 90, 244–250. [Google Scholar] [CrossRef]
- Sato, H.; Nagare, H.; Huynh, T.N.C.; Komatsu, H. Development of a new wastewater treatment process for resource recovery of carotenoids. Water Sci. Technol. 2015, 72, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Domınguez-Bocanegra, A.R.; Legarreta, I.G.; Jeronimo, F.M.; Campocosio, A.T. Influence of environmental and nutritional factors in the production of astaxanthin from Haematococcus pluvialis. Bioresour. Technol. 2004, 92, 209–214. [Google Scholar] [CrossRef]
- Chougle, J.A.; Bankar, S.B.; Chavan, P.V.; Patravale, V.B.; Singhal, R.S. Supercritical carbon dioxide extraction of astaxanthin from Paracoccus NBRC 101723: Mathematical modelling study. Sep. Sci. Technol. 2016, 51, 2164–2173. [Google Scholar] [CrossRef]
- Xiao, R.; Li, X.; Leonard, E.; Tharayil, N.; Zheng, Y. Investigation on the effects of cultivation conditions, fed-batch operation, and enzymatic hydrolysate of corn stover on the astaxanthin production by Thraustochytrium striatum. Algal Res. 2019, 39, 101475. [Google Scholar] [CrossRef]
- Bhatt, P.C.; Ahmad, M.; Panda, B.P. Enhanced bioaccumulation of astaxanthin in Phaffia rhodozyma by utilising low-cost agro products as fermentation substrate. Biocatal. Agric. Biotechnol. 2013, 2, 58–63. [Google Scholar] [CrossRef]
- Yang, J.; Tan, H.; Yang, R.; Sun, X.; Zhai, H.; Li, K. Astaxanthin production by Phaffia rhodozyma fermentation of cassava residues substrate. Agric. Eng. Int. CIGR J. 2011, 13, 1–6. [Google Scholar]
- Gervasi, T.; Santini, A.; Daliu, P.; Salem, A.Z.M.; Gervasi, C.; Pellizzeri, V.; Barrega, L.; De Pasquale, P.; Dugo, G.; Cicero, N. Astaxanthin production by Xanthophyllomyces dendrorhous growing on a low cost substrate. Agrofor. Syst. 2019, 94, 1229–1234. [Google Scholar] [CrossRef]
- Rodriguez Amado, I.; Antonio Vazquez, J.; Amado, I.R.; Vázquez, J.A. Mussel processing wastewater: A low-cost substrate for the production of astaxanthin by Xanthophyllomyces dendrorhous. Microb. Cell Factories 2015, 14, 177. [Google Scholar] [CrossRef]
- Wu, W.; Lu, M.; Yu, L. Citrus Residues Isolates Improve Astaxanthin Production by Xanthophyllomyces dendrorhous. Z. Fur. Nat. Sect. C-J. Biosci. 2010, 65, 594–598. [Google Scholar] [CrossRef]
- Tinoi, J.; Rakariyatham, N.; Deming, R.L. Utilization of mustard waste isolates for improved production of astaxanthin by Xanthophyllomyces dendrorhous. J. Ind. Microbiol. Biotechnol. 2006, 33, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Sujarit, C.; Rittirut, W.; Amornlerdpison, D.; Siripatana, C. Astaxanthin production from sewage of traditional Thai rice vermicelli. J. Phys. Conf. Ser. 2017, 820, 12011. [Google Scholar] [CrossRef]
- Irtiza, A.; Shatunova, S.; Glukhareva, T.; Kovaleva, E. Production of astaxanthin rich feed supplement for animals from Phaffia rhodozyma yeast at low cost. In Proceedings of the IV International Young Researchers’ Conference: Physics, Technologies and Innovation (PTI-2017), Ekaterinburg, Russia, 15–19 May 2017; Valeeva, A.A., Volkovich, V.A., Eds.; AIP Publishing: Woodbury, NY, USA, 2017; Volume 1886. [Google Scholar] [CrossRef]
- Fontana, J.D.; Baron, M.; Guimaraes, M.F.; Maraschin, M.; Florêncio, J.A.; Bonfim, T.M.B.; Chocial, M.B.; Ulhoa, C. Astaxanthinogenesis in the yeast Phaffia rhodozyma. In Biotechnology for Fuels and Chemicals; Springer: Cham, Switzerland, 1997; pp. 305–314. [Google Scholar]
- Guo, X.; Li, X.; Xiao, D. Optimization of culture conditions for production of astaxanthin by Phaffia rhodozyma. In Proceedings of the 4th International Conference on Bioinformatics and Biomedical Engineering, Chengdu, China, 18–20 June 2010; pp. 1–4. [Google Scholar]
- Dahiya, S.; Kumar, A.N.; Sravan, J.S.; Chatterjee, S.; Sarkar, O.; Mohan, S.V. Food waste biorefinery: Sustainable strategy for circular bioeconomy. Bioresour. Technol. 2018, 248, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Maina, S.; Kachrimanidou, V.; Koutinas, A. A roadmap towards a circular and sustainable bioeconomy through waste valorization. Curr. Opin. Green Sustain. Chem. 2017, 8, 18–23. [Google Scholar] [CrossRef]
- Woiciechowski, A.L.; Karp, S.G.; Sobral, K.; De Carvalho, J.C.; Letti, L.A.J.; Soccol, V.T.; Soccol, C.R. Pretreatment strategies to enhance value addition of agro-industrial wastes. In Biotransformation of Waste Biomass into High Value Biochemicals; Springer: Cham, Switzerland, 2014; ISBN 9781461480. [Google Scholar] [CrossRef]
- Iglesias, H. Handbook of Food Isotherms: Water Sorption Parameters for Food and Food Components; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Ribas, M.M.F.; Cereda, M.P.; Bôas, V.; Lyra, R. Use of cassava wastewater treated anaerobically with alkaline agents as fertilizer for maize (Zea mays L.). Braz. Arch. Biol. Technol. 2010, 53, 55–62. [Google Scholar] [CrossRef]
- Marques, S.S.I.; Nascimento, I.A.; de Almeida, P.F.; Chinalia, F.A. Growth of Chlorella vulgaris on sugarcane vinasse: The effect of anaerobic digestion pretreatment. Appl. Biochem. Biotechnol. 2013, 171, 1933–1943. [Google Scholar] [CrossRef]
- Woiciechowski, A.L.; de Souza Vandenberghe, L.P.; Karp, S.G.; Letti, L.A.J.; de Carvalho, J.C.; Medeiros, A.B.P.; Spier, M.R.; Faraco, V.; Soccol, V.T.; Soccol, C.R. The pretreatment step in lignocellulosic biomass conversion: Current systems and new biological systems. In Lignocellulose Conversion: Enzymatic and Microbial Tools for Bioethanol Production; Springer: Cham, Switzerland, 2013. [Google Scholar] [CrossRef]
- Beltrán-Rocha, J.C.; Guajardo-Barbosa, C.; Barceló-Quinta, I.D.; López-Chuken, U.J. Biotreatment of secondary municipal effluents using microalgae: Effect of pH, nutrients (C, N AND P) and CO2 enrichment. Rev. Biol. Mar. Oceanogr. 2017, 52, 417–427. [Google Scholar] [CrossRef]
- Chavan, R.; Mutnuri, S. Tertiary treatment of domestic wastewater by Spirulina platensis integrated with microalgal biorefinery. Biofuels 2018, 10, 33–34. [Google Scholar] [CrossRef]
- Gouveia, L.; Graça, S.; Sousa, C.; Ambrosano, L.; Ribeiro, B.; Botrel, E.P.; Neto, P.C.; Ferreira, A.F.; Silva, C.M. Microalgae biomass production using wastewater: Treatment and costs. Scale-up considerations. Algal Res. 2016, 16, 167–176. [Google Scholar] [CrossRef]
- Ledda, C.; Romero Villegas, G.I.; Adani, F.; Acién Fernández, F.G.; Molina Grima, E. Utilization of centrate from wastewater treatment for the outdoor production of Nannochloropsis gaditana biomass at pilot-scale. Algal Res. 2015, 12, 17–25. [Google Scholar] [CrossRef]
- Goyzueta-Mamani, L.D.; de Carvalho, J.C.; Magalhães, A.I.; Soccol, C.R. Production of arachidonic acid by Mortierella alpina using wastes from potato chips industry. J. Appl. Microbiol. 2020, 130, 1592–1601. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhou, W.; Chen, H.; Zhan, J.; He, C.; Wang, Q. Ammonium nitrogen tolerant chlorella strain screening and its damaging effects on photosynthesis. Front. Microbiol. 2019, 9, 3250. [Google Scholar] [CrossRef] [PubMed]
- Molina, D.; de Carvalho, J.C.; Júnior, A.I.M.; Faulds, C.; Bertrand, E.; Soccol, C.R. Biological contamination and its chemical control in microalgal mass cultures. Appl. Microbiol. Biotechnol. 2019, 103, 9345–9358. [Google Scholar] [CrossRef] [PubMed]
- Ambati, R.R.; Moi, P.S.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, Extraction, Stability, Biological Activities and Its Commercial Applications—A Review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration. GRAS Notice Inventory. Generally Recognized as Safe (GRAS) Notice Inventory. 2021. Available online: https://www.fda.gov/food/generally-recognized-safe-gras/gras-notice-inventory (accessed on 29 April 2021).
- FDA. Dietary Supplement Products & Ingredients. Available online: https://www.fda.gov/food/dietary-supplements/dietary-supplement-products-ingredients (accessed on 16 September 2022).



| Component | Concentration, g/L or kg/m3 of Medium | Component Price, US$/ton FOB, China * | |||
|---|---|---|---|---|---|
| NaNO3 | 0.250 | 350 | Medium cost per cubic meter | US$ 0.35 | |
| CaCl2•2H2O | 0.025 | 120 | Biomass production **, kg biomass per cubic meter of medium | 0.643 kg | |
| MgSO4•7H2O | 0.074 | 100 | Contribution of media components to biomass cost, US$/kg biomass | US$ 0.54 | |
| K2HPO4 | 0.075 | 1100 | Astaxanthin production, kg per cubic meter of medium | 0.0257 kg | |
| KH2PO4 | 0.175 | 950 | Contribution of media components to astaxanthin cost, US$/kg astaxanthin | US$ 13.55 | |
| NaCl | 0.025 | 65 |
| Components | Cassava Wastewater (Average) [23] | Secondary Effluent (Treated Piggery Wastewater) [29] | Digested Cassava Wastewater [57] | Cane Molasses, Diluted to 10% w/v Sugars [36,37] | Anaerobically Digested Vinasse [58] | Mixed Scotta (Cheese Whey) [35] |
|---|---|---|---|---|---|---|
| Total carbohydrates (g/L) | 32.9 | 100 | 40.7 | |||
| Total lipids (g/L) | 0.12 | 1.8 | ||||
| Protein (g/L) | 1.53 | 9.1 | ||||
| Total nitrogen (g/L) | 0.84 | 2.38 | 0.02 | 1.1 | ||
| Phosphorus (mg/L) | 485.6 | 98.7 | 87 | 14 | 930 | |
| Potassium (mg/L) | 1324.9 | 1680 | 38 | 1190 | ||
| Calcium (mg/L) | 162.63 | 190 | 480 | |||
| Magnesium (mg/L) | 242.96 | 420 | 300 | |||
| Sulfur (mg/L) | 50.33 | 660 | 40 | |||
| Iron (mg/L) | 7.57 | 0.4 | 164 | |||
| Zinc (mg/L) | 3.83 | 9.2 | 73 | |||
| Manganese (mg/L) | 1.21 | 0.4 | 3.13 | |||
| Copper (mg/L) | 0.48 | 0.8 | ||||
| Sodium (mg/L) | 51.7 | 9.6 | 2000 | |||
| pH | 5.46 | 7.85 | 8.05 | 6.7 | 6.02 | |
| COD (g O2/L) | 20.21 | 800 | 17.1 | 0.3 | ||
| Total Solids | 15.49 | 44 | 157.9 | 77.5 | ||
| Ammonium, mg/L | 41.42 | 12.9 | 20 | |||
| Nitrate, mg/L | 24.27 | 469 | 2 | |||
| C:N ratio | 15.8 | 24.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Carvalho, J.C.; Goyzueta-Mamani, L.D.; Molina-Aulestia, D.T.; Magalhães Júnior, A.I.; Iwamoto, H.; Ambati, R.; Ravishankar, G.A.; Soccol, C.R. Microbial Astaxanthin Production from Agro-Industrial Wastes—Raw Materials, Processes, and Quality. Fermentation 2022, 8, 484. https://doi.org/10.3390/fermentation8100484
de Carvalho JC, Goyzueta-Mamani LD, Molina-Aulestia DT, Magalhães Júnior AI, Iwamoto H, Ambati R, Ravishankar GA, Soccol CR. Microbial Astaxanthin Production from Agro-Industrial Wastes—Raw Materials, Processes, and Quality. Fermentation. 2022; 8(10):484. https://doi.org/10.3390/fermentation8100484
Chicago/Turabian Stylede Carvalho, Júlio Cesar, Luis Daniel Goyzueta-Mamani, Denisse Tatiana Molina-Aulestia, Antônio Irineudo Magalhães Júnior, Hissashi Iwamoto, RangaRao Ambati, Gokare A. Ravishankar, and Carlos Ricardo Soccol. 2022. "Microbial Astaxanthin Production from Agro-Industrial Wastes—Raw Materials, Processes, and Quality" Fermentation 8, no. 10: 484. https://doi.org/10.3390/fermentation8100484