
Liquid Biopsies Poorly miRror Renal Ischemia-Reperfusion Injury

Abstract
:1. Introduction
2. Results
2.1. Reduced Renal Clearance in This IR Model
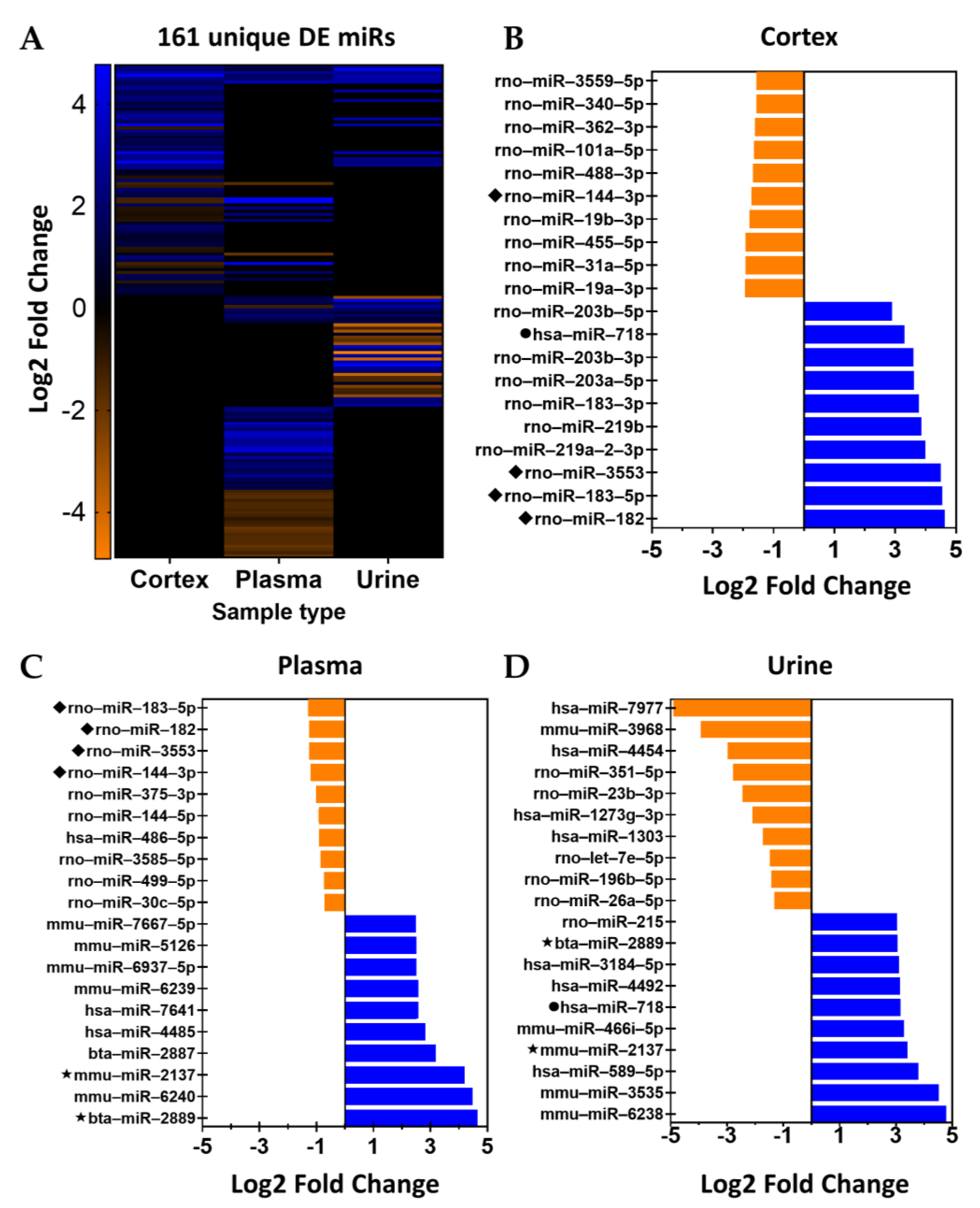
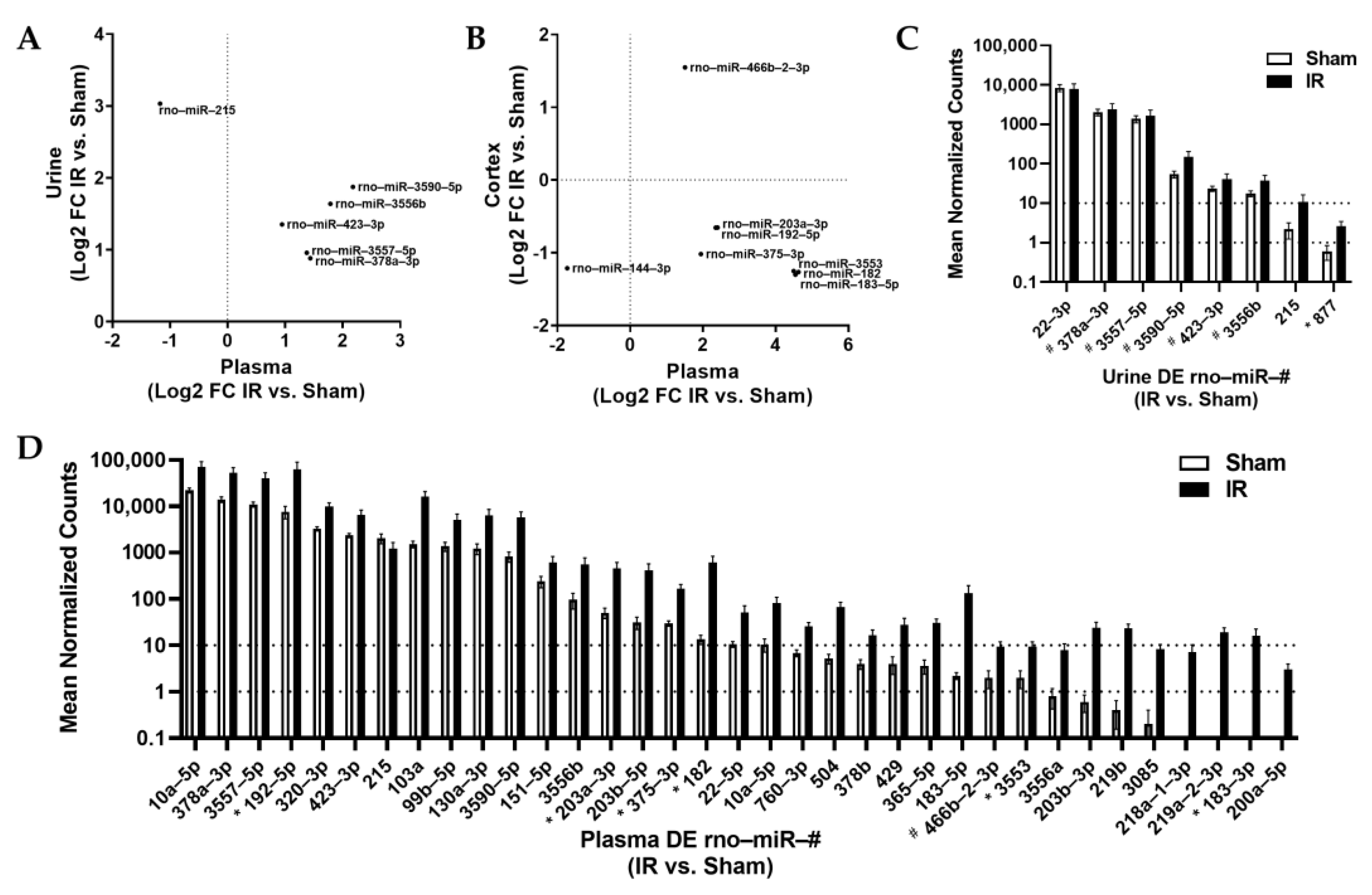
2.2. Small-RNA Sequencing Results
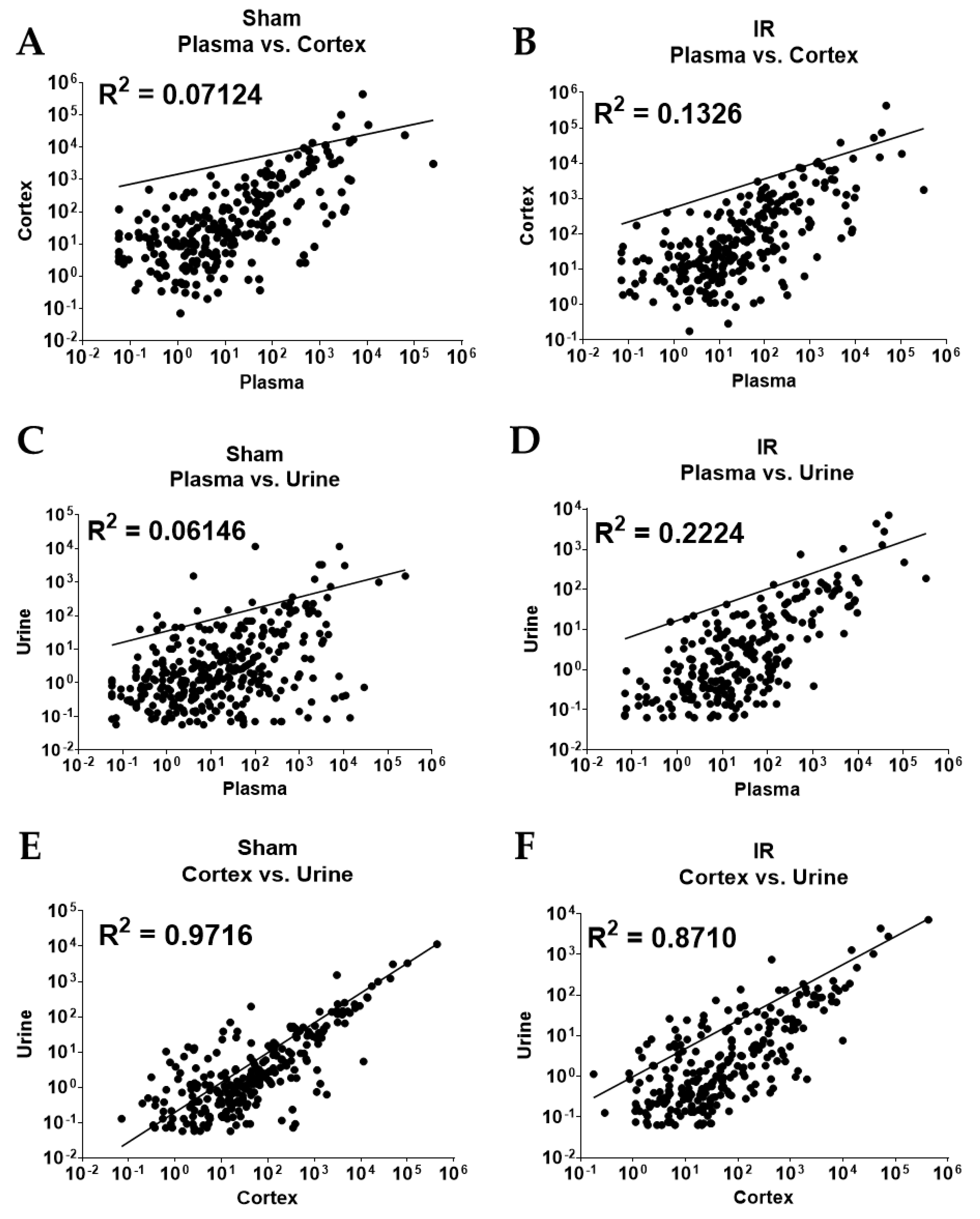
2.3. Overall Correlation of Abundance between Sample Types
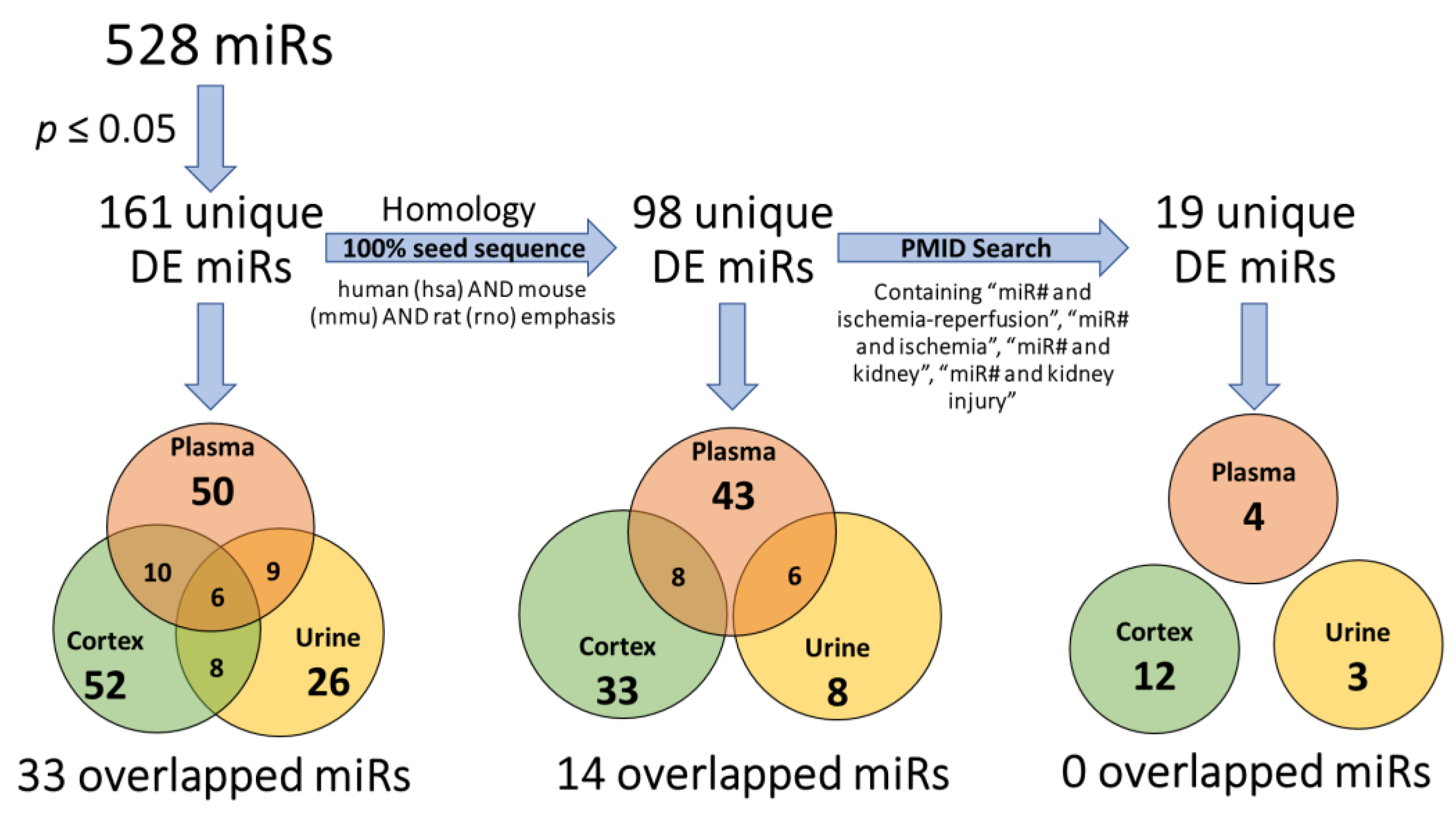
2.4. Analysis by Nomenclature
2.5. Homology and Abundance
2.6. Prior Identification in Related Studies
3. Discussion
3.1. Reflection of Organ-Specific miR Changes in Biofluids
3.2. Alignment of Biofluid miRs with Established Biomarkers
3.3. Importance of Study Design
3.4. Limitations
4. Materials and Methods
4.1. Ischemia-Reperfusion (IR) Model
4.2. RNA Extraction and MiR Library Prep
4.3. Library Construction, Sequencing, and Analysis
4.4. Post-Hoc Analysis of miR Profile Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, Y.L.; Li, H.F.; Chen, H.H.; Lin, H. MicroRNAs as Biomarkers and Therapeutic Targets in Inflammation- and Ischemia-Reperfusion-Related Acute Renal Injury. Int. J. Mol. Sci. 2020, 21, 6738. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, F.; Zhang, J.; Sun, Z.; Wang, W. Advances of miRNAs in kidney graft injury. Transpl. Rev. 2021, 35, 100591. [Google Scholar] [CrossRef]
- Zankar, S.; Rodriguez, R.A.; Vinas, J.L.; Burns, K.D. The therapeutic effects of microRNAs in preclinical studies of acute kidney injury: A systematic review protocol. Syst. Rev. 2019, 8, 235. [Google Scholar] [CrossRef] [PubMed]
- Banaei, S. Novel role of microRNAs in renal ischemia reperfusion injury. Ren. Fail. 2015, 37, 1073–1079. [Google Scholar] [CrossRef]
- Lorenzen, J.M.; Batkai, S.; Thum, T. Regulation of cardiac and renal ischemia-reperfusion injury by microRNAs. Free Radic. Biol. Med. 2013, 64, 78–84. [Google Scholar] [CrossRef]
- Lorenzen, J.M. Vascular and circulating microRNAs in renal ischaemia-reperfusion injury. J. Physiol. 2015, 593, 1777–1784. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilad, S.; Meiri, E.; Yogev, Y.; Benjamin, S.; Lebanony, D.; Yerushalmi, N.; Benjamin, H.; Kushnir, M.; Cholakh, H.; Melamed, N.; et al. Serum microRNAs are promising novel biomarkers. PLoS ONE 2008, 3, e3148. [Google Scholar] [CrossRef] [Green Version]
- Bagnasco, S.; Good, D.; Balaban, R.; Burg, M. Lactate production in isolated segments of the rat nephron. Am. J. Physiol. 1985, 248, F522–F526. [Google Scholar] [CrossRef]
- Hall, A.M.; de Seigneux, S. Metabolic mechanisms of acute proximal tubular injury. Pflugers Arch. 2022, 474, 813–827. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, N.; Hosford, M.; Sandoval, R.M.; Wagner, M.C.; Atkinson, S.J.; Bamburg, J.; Molitoris, B.A. Ischemia activates actin depolymerizing factor: Role in proximal tubule microvillar actin alterations. Am. J. Physiol. 1999, 276, F544–F551. [Google Scholar] [CrossRef]
- Tingle, S.J.; Sewpaul, A.; Bates, L.; Thompson, E.R.; Shuttleworth, V.; Figueiredo, R.; Ibrahim, I.K.; Ali, S.; Wilson, C.; Sheerin, N.S. Dual MicroRNA Blockade Increases Expression of Antioxidant Protective Proteins: Implications for Ischemia-Reperfusion Injury. Transplantation 2020, 104, 1853–1861. [Google Scholar] [CrossRef]
- Liu, D.; Liu, Y.; Zheng, X.; Liu, N. c-MYC-induced long noncoding RNA MEG3 aggravates kidney ischemia-reperfusion injury through activating mitophagy by upregulation of RTKN to trigger the Wnt/β-catenin pathway. Cell Death Dis. 2021, 12, 191. [Google Scholar] [CrossRef] [PubMed]
- Ou, H.; Teng, H.; Qin, Y.; Luo, X.; Yang, P.; Zhang, W.; Chen, W.; Lv, D.; Tang, H. Extracellular vesicles derived from microRNA-150-5p-overexpressing mesenchymal stem cells protect rat hearts against ischemia/reperfusion. Aging 2020, 12, 12669–12683. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhao, S.; Li, W.; Ruan, Y.; Yuan, R.; Ning, J.; Jiang, K.; Xie, J.; Yao, X.; Li, H.; et al. Tubular cell-derived exosomal miR-150-5p contributes to renal fibrosis following unilateral ischemia-reperfusion injury by activating fibroblast in vitro and in vivo. Int. J. Biol. Sci. 2021, 17, 4021–4033. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Li, Y.; Zhang, L.; Chen, X.; Xu, H. Foxp3 attenuates cerebral ischemia/reperfusion injury through microRNA-150-5p-modified NCS1. Exp. Cell Res. 2021, 2021, 112942. [Google Scholar] [CrossRef]
- Li, X.; Bi, T.; Yang, S. Exosomal microRNA-150-5p from bone marrow mesenchymal stromal cells mitigates cerebral ischemia/reperfusion injury via targeting toll-like receptor 5. Bioengineered 2022, 13, 3030–3043. [Google Scholar] [CrossRef]
- Godwin, J.G.; Ge, X.; Stephan, K.; Jurisch, A.; Tullius, S.G.; Iacomini, J. Identification of a microRNA signature of renal ischemia reperfusion injury. Proc. Natl. Acad. Sci. USA 2010, 107, 14339–14344. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Pei, L.; Lin, F.; Yin, H.; Li, X.; He, W.; Liu, N.; Gou, X. Exosomes from human-bone-marrow-derived mesenchymal stem cells protect against renal ischemia/reperfusion injury via transferring miR-199a-3p. J. Cell. Physiol. 2019, 234, 23736–23749. [Google Scholar] [CrossRef]
- Yu, L.; Gu, T.; Shi, E.; Wang, Y.; Fang, Q.; Wang, C. Dysregulation of renal microRNA expression after deep hypothermic circulatory arrest in rats. Eur. J. Cardio-Thorac. Surg. Off. J. Eur. Assoc. Cardio-Thorac. Surg. 2016, 49, 1725–1731. [Google Scholar] [CrossRef]
- Papadopoulos, T.; Casemayou, A.; Neau, E.; Breuil, B.; Caubet, C.; Calise, D.; Thornhill, B.A.; Bachvarova, M.; Belliere, J.; Chevalier, R.L.; et al. Systems biology combining human- and animal-data miRNA and mRNA data identifies new targets in ureteropelvic junction obstruction. BMC Syst. Biol. 2017, 11, 31. [Google Scholar] [CrossRef] [Green Version]
- Gan, J.; Yuan, J.; Liu, Y.; Lu, Z.; Xue, Y.; Shi, L.; Zeng, H. Circular RNA_101237 mediates anoxia/reoxygenation injury by targeting let-7a-5p/IGF2BP3 in cardiomyocytes. Int. J. Mol. Med. 2020, 45, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Viñas, J.L.; Burger, D.; Zimpelmann, J.; Haneef, R.; Knoll, W.; Campbell, P.; Gutsol, A.; Carter, A.; Allan, D.S.; Burns, K.D. Transfer of microRNA-486-5p from human endothelial colony forming cell-derived exosomes reduces ischemic kidney injury. Kidney Int. 2016, 90, 1238–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinas, J.L.; Spence, M.; Gutsol, A.; Knoll, W.; Burger, D.; Zimpelmann, J.; Allan, D.S.; Burns, K.D. Receptor-Ligand Interaction Mediates Targeting of Endothelial Colony Forming Cell-derived Exosomes to the Kidney after Ischemic Injury. Sci. Rep. 2018, 8, 16320. [Google Scholar] [CrossRef] [Green Version]
- Vinas, J.L.; Spence, M.; Porter, C.J.; Douvris, A.; Gutsol, A.; Zimpelmann, J.A.; Campbell, P.A.; Burns, K.D. micro-RNA-486-5p protects against kidney ischemic injury and modifies the apoptotic transcriptome in proximal tubules. Kidney Int. 2021, 100, 597–612. [Google Scholar] [CrossRef]
- Zou, Y.F.; Wen, D.; Zhao, Q.; Shen, P.Y.; Shi, H.; Zhao, Q.; Chen, Y.X.; Zhang, W. Urinary MicroRNA-30c-5p and MicroRNA-192-5p as potential biomarkers of ischemia-reperfusion-induced kidney injury. Exp. Biol. Med. 2017, 242, 657–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Yu, S.; Zheng, B.; Liu, D.; Wan, F.; Ma, Y.; Wang, J.; Gao, Z.; Shan, Z. miR-30c-5p Reduces Renal Ischemia-Reperfusion Involving Macrophage. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 4362–4369. [Google Scholar] [CrossRef]
- Pellegrini, K.L.; Gerlach, C.V.; Craciun, F.L.; Ramachandran, K.; Bijol, V.; Kissick, H.T.; Vaidya, V.S. Application of small RNA sequencing to identify microRNAs in acute kidney injury and fibrosis. Toxicol. Appl. Pharmacol. 2016, 312, 42–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Q.; Liu, Y.; Lv, X.W.; Dai, R.X.; Yang, X.H.; Kong, B.H. LncRNA TUG1 mediates ischemic myocardial injury by targeting miR-132-3p/HDAC3 axis. Am. J. Physiol. Heart Circ. Physiol. 2020, 318, H729. [Google Scholar] [CrossRef]
- Liu, R.R.; Li, J.; Gong, J.Y.; Kuang, F.; Liu, J.Y.; Zhang, Y.S.; Ma, Q.L.; Song, C.J.; Truax, A.D.; Gao, F.; et al. MicroRNA-141 regulates the expression level of ICAM-1 on endothelium to decrease myocardial ischemia-reperfusion injury. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1303–H1313. [Google Scholar] [CrossRef]
- Chu, C.P.; Liu, S.; Song, W.; Xu, E.Y.; Nabity, M.B. Small RNA sequencing evaluation of renal microRNA biomarkers in dogs with X-linked hereditary nephropathy. Sci. Rep. 2021, 11, 17437. [Google Scholar] [CrossRef]
- Connor, K.L.; Teenan, O.; Cairns, C.; Banwell, V.; Thomas, R.A.; Rodor, J.; Finnie, S.; Pius, R.; Tannahill, G.M.; Sahni, V.; et al. Identifying cell-enriched miRNAs in kidney injury and repair. JCI Insight 2020, 5, e140399. [Google Scholar] [CrossRef]
- Wang, I.K.; Sun, K.T.; Tsai, T.H.; Chen, C.W.; Chang, S.S.; Yu, T.M.; Yen, T.H.; Lin, F.Y.; Huang, C.C.; Li, C.Y. MiR-20a-5p mediates hypoxia-induced autophagy by targeting ATG16L1 in ischemic kidney injury. Life Sci. 2015, 136, 133–141. [Google Scholar] [CrossRef]
- Yu, W.; Zeng, H.; Chen, J.; Fu, S.; Huang, Q.; Xu, Y.; Xu, A.; Lan, H.Y.; Tang, Y. miR-20a-5p is enriched in hypoxia-derived tubular exosomes and protects against acute tubular injury. Clin. Sci. 2020, 134, 2223–2234. [Google Scholar] [CrossRef]
- Lee, M.S.; Yip, H.K.; Yang, C.C.; Chiang, J.Y.; Huang, T.H.; Li, Y.C.; Chen, K.H.; Sung, P.H. Overexpression of miR-19a and miR-20a in iPS-MSCs preserves renal function of chronic kidney disease with acute ischaemia-reperfusion injury in rat. J. Cell. Mol. Med. 2021, 25, 7675–7689. [Google Scholar] [CrossRef]
- Zaman, A.; Banday, A.A. Angiotensin (1–7) protects against renal ischemia-reperfusion injury via regulating expression of NRF2 and microRNAs in Fisher 344 rats. Am. J. Physiol.-Ren. Physiol. 2022, 323, F33–F47. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Deng, X.; Kang, Z.; Wang, Y.; Xia, T.; Ding, N.; Yin, Y. Elevation of miR-21, through targeting MKK3, may be involved in ischemia pretreatment protection from ischemia-reperfusion induced kidney injury. J. Nephrol. 2016, 29, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Cao, Y.; Wang, H.; Wang, Q.; Ji, J.; Sun, X.; Dong, Z. Circular RNA YAP1 acts as the sponge of microRNA-21-5p to secure HK-2 cells from ischaemia/reperfusion-induced injury. J. Cell. Mol. Med. 2020, 24, 4707–4715. [Google Scholar] [CrossRef] [Green Version]
- Cabral, A.; da Silva Cândido, D.; Monteiro, S.M.; Lemos, F.; Saitovitch, D.; Noronha, I.L.; Alves, L.F.; Geraldo, M.V.; Kalil, J.; Cunha-Neto, E.; et al. Differential microRNA Profile in Operational Tolerance: A Potential Role in Favoring Cell Survival. Front. Immunol. 2019, 10, 740. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Qiu, W.; Ma, J.; Wang, Y.; Hu, Z.; Long, K.; Wang, X.; Jin, L.; Tang, Q.; Tang, G.; et al. miR-27a-5p Attenuates Hypoxia-induced Rat Cardiomyocyte Injury by Inhibiting Atg7. Int. J. Mol. Sci. 2019, 20, 2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glineur, S.F.; Hanon, E.; Dremier, S.; Snelling, S.; Berteau, C.; De Ron, P.; Nogueira da Costa, A. Assessment of a Urinary Kidney MicroRNA Panel as Potential Nephron Segment-Specific Biomarkers of Subacute Renal Toxicity in Preclinical Rat Models. Toxicol. Sci. Off. J. Soc. Toxicol. 2018, 166, 409–419. [Google Scholar] [CrossRef]
- Wang, J.; Li, X.; Wu, X.; Wang, Z.; Zhang, C.; Cao, G.; Yan, T. Expression Profiling of Exosomal miRNAs Derived from the Peripheral Blood of Kidney Recipients with DGF Using High-Throughput Sequencing. BioMed. Res. Int. 2019, 2019, 1759697. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Xiong, J.; Wen, H.; Wei, H.; Zeng, X. MiR-98-5p promotes ischemia/reperfusion-induced microvascular dysfunction by targeting NGF and is a potential biomarker for microvascular reperfusion. Microcirculation 2021, 28, e12657. [Google Scholar] [CrossRef]
- Takuma, A.; Abe, A.; Saito, Y.; Nito, C.; Ueda, M.; Ishimaru, Y.; Harada, H.; Abe, K.; Kimura, K.; Asakura, T. Gene Expression Analysis of the Effect of Ischemic Infarction in Whole Blood. Int. J. Mol. Sci. 2017, 18, 2335. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhou, X.; Wu, Y. The miR-26a-5p/IL-6 axis alleviates sepsis-induced acute kidney injury by inhibiting renal inflammation. Ren. Fail. 2022, 44, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Bellinger, M.A.; Bean, J.S.; Rader, M.A.; Heinz-Taheny, K.M.; Nunes, J.S.; Haas, J.V.; Michael, L.F.; Rekhter, M.D. Concordant changes of plasma and kidney microRNA in the early stages of acute kidney injury: Time course in a mouse model of bilateral renal ischemia-reperfusion. PLoS ONE 2014, 9, e93297. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Luo, S.; Zhang, J.; Yuan, Y.; Jiang, W.; Zhu, H.; Ding, X.; Zhan, L.; Wu, H.; Xie, Y.; et al. lncRNA H19 Alleviated Myocardial I/RI via Suppressing miR-877-3p/Bcl-2-Mediated Mitochondrial Apoptosis. Mol. Ther.-Nucleic Acids 2019, 17, 297–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saikumar, J.; Hoffmann, D.; Kim, T.M.; Gonzalez, V.R.; Zhang, Q.; Goering, P.L.; Brown, R.P.; Bijol, V.; Park, P.J.; Waikar, S.S.; et al. Expression, circulation, and excretion profile of microRNA-21, -155, and -18a following acute kidney injury. Toxicol. Sci. Off. J. Soc. Toxicol. 2012, 129, 256–267. [Google Scholar] [CrossRef] [Green Version]
- Marrone, A.K.; Stolz, D.B.; Bastacky, S.I.; Kostka, D.; Bodnar, A.J.; Ho, J. MicroRNA-17~92 is required for nephrogenesis and renal function. J. Am. Soc. Nephrol. 2014, 25, 1440–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiba, T.; Cerqueira, D.M.; Li, Y.; Bodnar, A.J.; Mukherjee, E.; Pfister, K.; Phua, Y.L.; Shaikh, K.; Sanders, B.T.; Hemker, S.L.; et al. Endothelial-Derived miR-17∼92 Promotes Angiogenesis to Protect against Renal Ischemia-Reperfusion Injury. J. Am. Soc. Nephrol. 2021, 32, 553–562. [Google Scholar] [CrossRef]
- Cai, Q.; Wang, C.; Huang, L.; Wu, C.; Yan, B.; Chen, T.; Li, Q.; Wang, L. Long Non-Coding RNA Small Nucleolar RNA Host Gene 5 (SNHG5) Regulates Renal Tubular Damage in Diabetic Nephropathy via Targeting MiR-26a-5p. Horm. Metab. Res. 2021, 53, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Chuppa, S.; Liang, M.; Liu, P.; Liu, Y.; Casati, M.C.; Cowley, A.W.; Patullo, L.; Kriegel, A.J. MicroRNA-21 regulates peroxisome proliferator-activated receptor alpha, a molecular mechanism of cardiac pathology in Cardiorenal Syndrome Type 4. Kidney Int. 2018, 93, 375–389. [Google Scholar] [CrossRef] [Green Version]
- Church, R.J.; McDuffie, J.E.; Sonee, M.; Otieno, M.; Ma, J.Y.; Liu, X.; Watkins, P.B.; Harrill, A.H. MicroRNA-34c-3p is an early predictive biomarker for doxorubicin-induced glomerular injury progression in male Sprague-Dawley rats. Toxicol. Res. 2014, 3, 384–394. [Google Scholar] [CrossRef]
- Kito, N.; Endo, K.; Ikesue, M.; Weng, H.; Iwai, N. miRNA Profiles of Tubular Cells: Diagnosis of Kidney Injury. BioMed Res. Int. 2015, 2015, 465479. [Google Scholar] [CrossRef] [Green Version]
- Mompeón, A.; Pérez-Cremades, D.; Paes, A.B.; Sanchis, J.; Ortega-Paz, L.; Andrea, R.; Brugaletta, S.; Sabate, M.; Novella, S.; Dantas, A.P.; et al. Circulating miRNA Fingerprint and Endothelial Function in Myocardial Infarction: Comparison at Acute Event and One-Year Follow-Up. Cells 2022, 11, 1823. [Google Scholar] [CrossRef] [PubMed]
- Han, W.K.; Bailly, V.; Abichandani, R.; Thadhani, R.; Bonventre, J.V. Kidney Injury Molecule-1 (KIM-1): A novel biomarker for human renal proximal tubule injury. Kidney Int. 2002, 62, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Cui, R.; Xu, J.; Chen, X.; Zhu, W. Global miRNA expression is temporally correlated with acute kidney injury in mice. PeerJ 2016, 4, e1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, J.H.; Kim, G.B.; Seok, H. Implication of microRNA as a potential biomarker of myocarditis. Clin. Exp. Pediatr. 2022, 65, 230–238. [Google Scholar] [CrossRef]
- Liang, Y.; Fang, D.; Gao, X.; Deng, X.; Chen, N.; Wu, J.; Zeng, M.; Luo, M. Circulating microRNAs as emerging regulators of COVID-19. Theranostics 2023, 13, 125–147. [Google Scholar] [CrossRef]
- Megahed, F.; Tabll, A.; Atta, S.; Ragheb, A.; Smolic, R.; Petrovic, A.; Smolic, M. MicroRNAs: Small Molecules with Significant Functions, Particularly in the Context of Viral Hepatitis B and C Infection. Medicina 2023, 59, 173. [Google Scholar] [CrossRef] [PubMed]
- Chorley, B.N.; Ellinger-Ziegelbauer, H.; Tackett, M.; Simutis, F.J.; Harrill, A.H.; McDuffie, J.; Atabakhsh, E.; Nassirpour, R.; Whiteley, L.O.; Leonard, J.F.; et al. Urinary miRNA Biomarkers of Drug-Induced Kidney Injury and Their Site Specificity Within the Nephron. Toxicol. Sci. Off. J. Soc. Toxicol. 2021, 180, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Cao, Y.; Sun, M.; Feng, H. Expression, regulation, and function of exosome-derived miRNAs in cancer progression and therapy. FASEB J. 2021, 35, e21916. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, B.; Li, Q.; Wei, S.; He, Z.; Huang, X.; Wang, L.; Xia, Y.; Xu, Z.; Li, Z.; et al. Exosomal miR-21-5p derived from gastric cancer promotes peritoneal metastasis via mesothelial-to-mesenchymal transition. Cell Death Dis. 2018, 9, 854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Gao, X.; Wei, F.; Zhang, X.; Yu, J.; Zhao, H.; Sun, Q.; Yan, F.; Yan, C.; Li, H.; et al. Diagnostic and prognostic value of circulating miR-21 for cancer: A systematic review and meta-analysis. Gene 2014, 533, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Kumarswamy, R.; Volkmann, I.; Thum, T. Regulation and function of miRNA-21 in health and disease. RNA Biol. 2011, 8, 706–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Li, W.; Yu, W.; Rao, T.; Li, H.; Ruan, Y.; Yuan, R.; Li, C.; Ning, J.; Li, S.; et al. Exosomal miR-21 from tubular cells contributes to renal fibrosis by activating fibroblasts via targeting PTEN in obstructed kidneys. Theranostics 2021, 11, 8660–8673. [Google Scholar] [CrossRef]
- Loboda, A.; Sobczak, M.; Jozkowicz, A.; Dulak, J. TGF-beta1/Smads and miR-21 in Renal Fibrosis and Inflammation. Mediat. Inflamm. 2016, 2016, 8319283. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kriegel, A.J.; Liu, Y.; Cohen, B.; Usa, K.; Liu, Y.; Liang, M. MiR-382 targeting of kallikrein 5 contributes to renal inner medullary interstitial fibrosis. Physiol. Genom. 2012, 44, 259–267. [Google Scholar] [CrossRef]
- Kriegel, A.J.; Liu, Y.; Liu, P.; Baker, M.A.; Hodges, M.R.; Hua, X.; Liang, M. Characteristics of microRNAs enriched in specific cell types and primary tissue types in solid organs. Physiol. Genom. 2013, 45, 1144–1156. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sham Group | IR Group | |||
|---|---|---|---|---|
| Test | Units | Mean ± SD | Mean ± SD | p-Value * |
| Urea | mg/dL | 17.6 ± 3.9 | 96.5 ± 25.1 | <0.001 * |
| Creatinine | mg/dL | 0.32 ± 0.04 | 3.80 ± 1.42 | 0.001 * |
| Total Protein | g/dL | 5.54 ± 0.21 | 5.47 ± 0.35 | 0.678 |
| Albumin | g/dL | 2.76 ± 0.11 | 2.63 ± 0.15 | 0.115 |
| Glucose | mg/dL | 175.4 ± 19.0 | 220.0 ± 38.1 | 0.025 * |
| Phosphorous | mg/dL | 8.32 ± 0.60 | 12.84 ± 2.96 | 0.006 * |
| Calcium | mg/dL | 9.62 ± 0.08 | 9.56 ± 0.34 | 0.654 |
| Sodium | mmol/L | 145.8 ± 0.8 | 140.4 ± 5.3 | 0.037 * |
| Potassium | mmol/L | 5.24 ± 0.35 | 6.59 ± 1.81 | 0.099 |
| Chloride | mmol/L | 102.6 ± 1.1 | 104.0 ± 19.1 | 0.853 |
| Bicarbonate | mmol/L | 25.8 ± 0.5 | 18.0 ± 5.9 | 0.012 * |
| Anion Gap | mmol/L | 22.6 ± 0.9 | 39.0 ± 27.5 | 0.166 |
| AST(GOT) | U/L | 266.2 ± 111.4 | 344.6 ± 60.7 | 0.205 |
| ALT(GPT) | U/L | 54.8 ± 16.2 | 81.7 ± 19.3 | 0.027 * |
| Alk. Ptase | U/L | 93.0 ± 4.5 | 128.0 ± 19.1 | 0.002 * |
| Globulin | g/dL | 2.78 ± 0.13 | 2.84 ± 0.21 | 0.544 |
| Gamma-GT | U/L | 0.2 ± 0.5 | 18.0 ± 3.3 | <0.001 * |
| Triglycerides | mg/dL | 38.2 ± 3.1 | 39.7 ± 5.4 | 0.551 |
| Cholesterol | mg/dL | 96.8 ± 14.4 | 87.3 ± 5.2 | 0.219 |
| Sample Type | miR and IR Studies References | p Value | adj. p Value | log2FC |
|---|---|---|---|---|
| Plasma | rno–miR–145–5p [12,13] | 0.0057 | 0.0401 | −0.7914 |
| rno–miR–150–5p [14,15,16,17] | 0.0025 | 0.0221 | −1.1176 | |
| rno–miR–199a–3p [18,19] | 0.0058 | 0.0401 | −1.2610 | |
| rno–miR–10a–3p [20] | <0.0001 | <0.0001 | 2.2347 | |
| rno–let–7a–5p [21,22] | 0.0055 | 0.0381 | 0.3956 | |
| Cortex | hsa–miR–486–5p [23,24,25] | 0.0045 | 0.0326 | −0.9168 |
| rno–miR–30c–5p [26,27] | 0.0018 | 0.0149 | −0.7125 | |
| rno–miR–132–3p [28,29] | <0.0001 | <0.0001 | 2.2297 | |
| rno–miR–146b–5p [28,30,31] | <0.0001 | <0.0001 | 2.1612 | |
| rno–miR–147 [31] | 0.0022 | 0.0180 | 1.3324 | |
| rno–miR–18a–5p [28,32] | <0.0001 | 0.0001 | 1.8245 | |
| rno–miR–20a–5p [33,34,35,36] | 0.0010 | 0.0098 | 0.8320 | |
| rno–miR–21–5p [36,37,38] | <0.0001 | <0.0001 | 1.3052 | |
| rno–miR–27a–5p [39,40] | 0.0001 | 0.0011 | 1.3213 | |
| rno–miR–34c–5p [41] | <0.0001 | <0.0001 | 1.2235 | |
| rno–miR–98–5p [42,43] | 0.0002 | 0.0026 | 0.5486 | |
| Urine | rno–miR–196b–5p [44] | 0.0037 | 0.0363 | −1.4190 |
| rno–miR–26a–5p [45] | 0.0005 | 0.0084 | −1.3099 | |
| rno–miR–877 [46,47] | 0.0031 | 0.0320 | 2.4805 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, A.C.; Singh, V.; Liu, P.; Kriegel, A.J. Liquid Biopsies Poorly miRror Renal Ischemia-Reperfusion Injury. Non-Coding RNA 2023, 9, 24. https://doi.org/10.3390/ncrna9020024
Williams AC, Singh V, Liu P, Kriegel AJ. Liquid Biopsies Poorly miRror Renal Ischemia-Reperfusion Injury. Non-Coding RNA. 2023; 9(2):24. https://doi.org/10.3390/ncrna9020024
Chicago/Turabian StyleWilliams, Adaysha C., Vaishali Singh, Pengyuan Liu, and Alison J. Kriegel. 2023. "Liquid Biopsies Poorly miRror Renal Ischemia-Reperfusion Injury" Non-Coding RNA 9, no. 2: 24. https://doi.org/10.3390/ncrna9020024