
FLVCR1-AS1 and FBXL19-AS1: Two Putative lncRNA Candidates in Multiple Human Cancers
 ,
,
Abstract
:1. Introduction
2. Search Strategy and Literature Selection
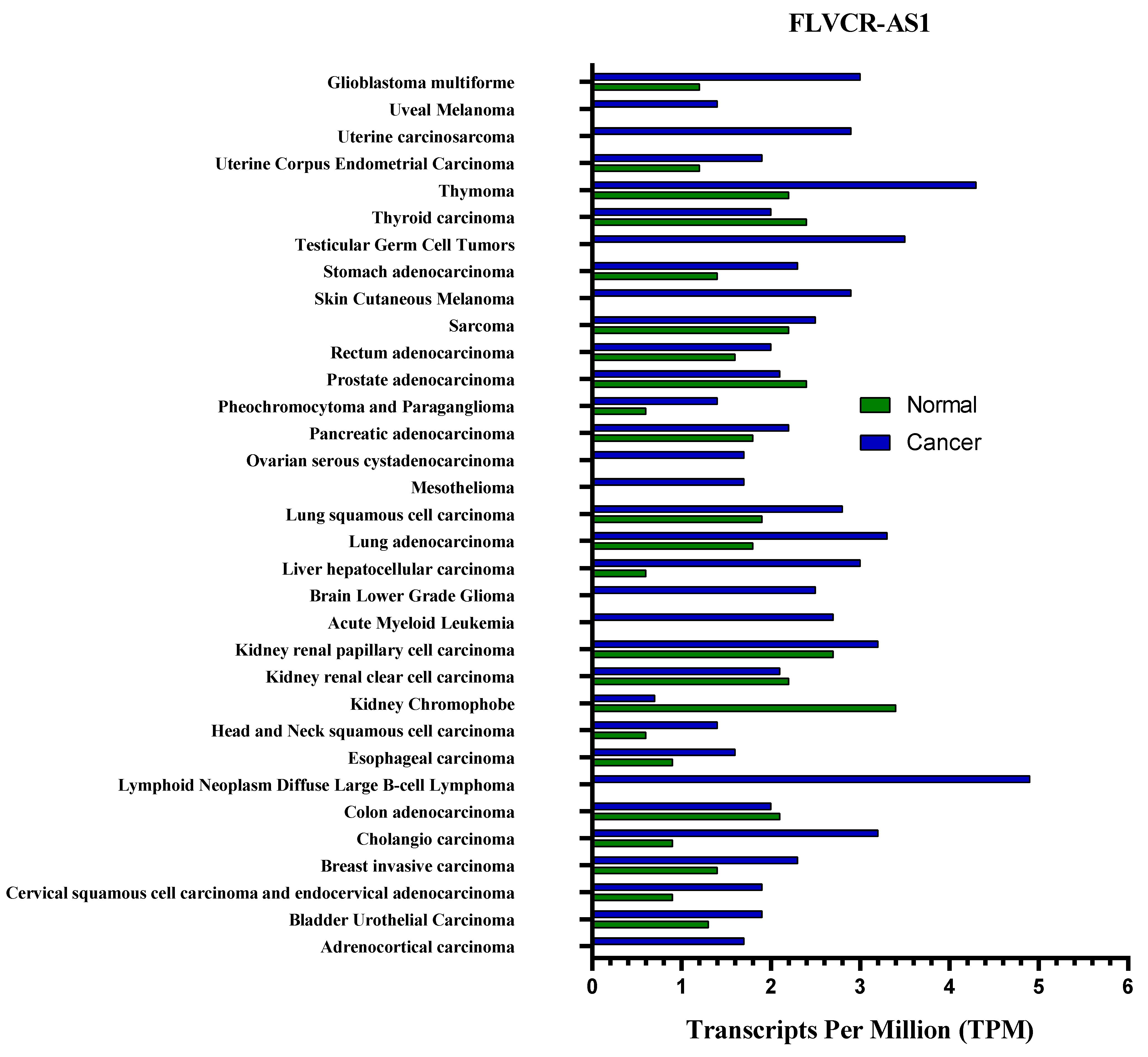
3. The Effect of FLVCR1-AS1 on Several Cancers
3.1. Cholangiocarcinoma
3.2. Hepatocellular Carcinoma
3.3. Gastric Cancer
3.4. Colorectal Cancer
3.5. Glioma and Glioblastoma
3.6. Non-Small Cell Lung Cancer
3.7. Ovarian Cancer
3.8. Breast Cancer
3.9. Osteosarcoma
3.10. Pancreatic Cancer
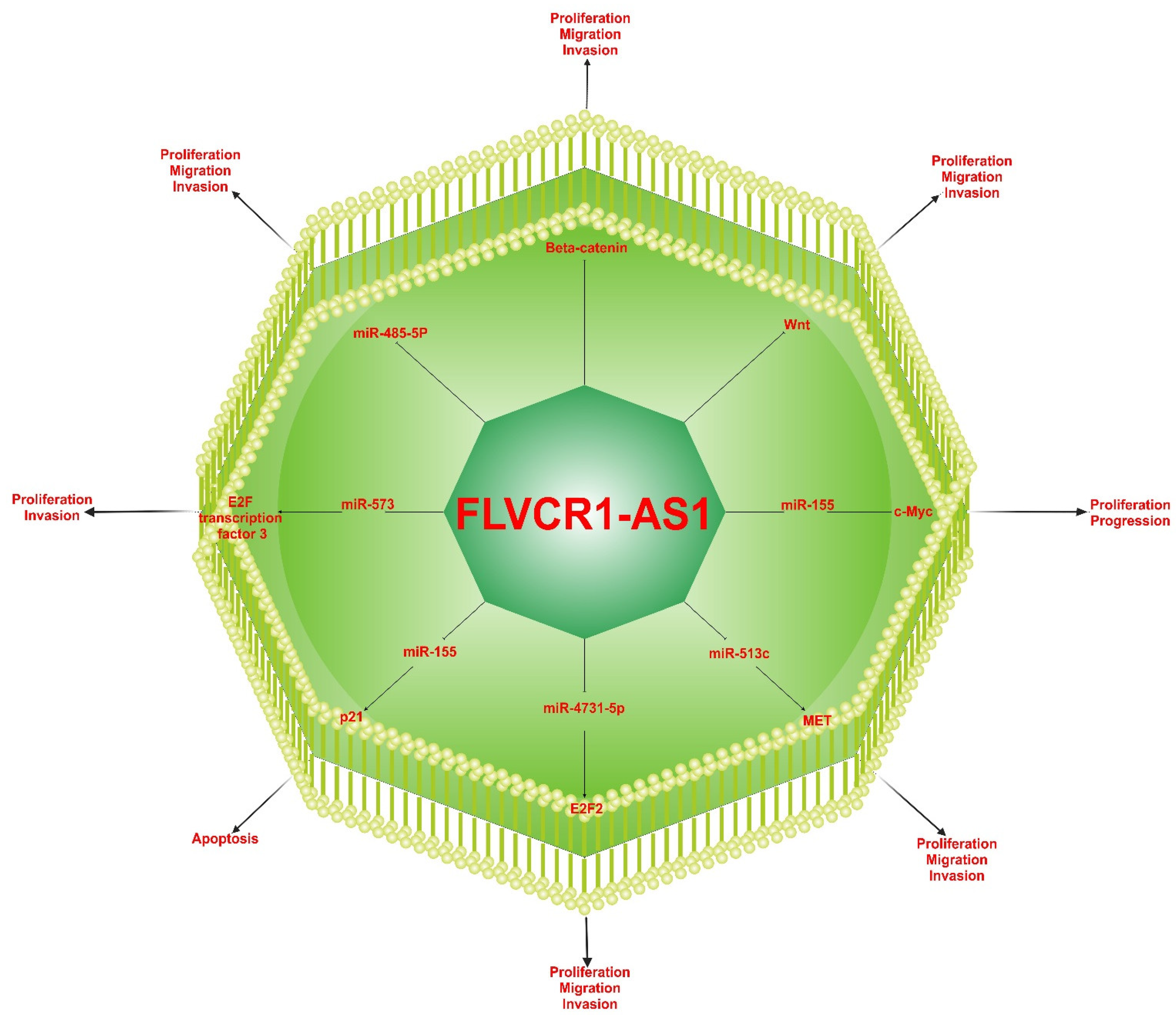
4. Regulating Mechanisms of FLVCR1-AS1
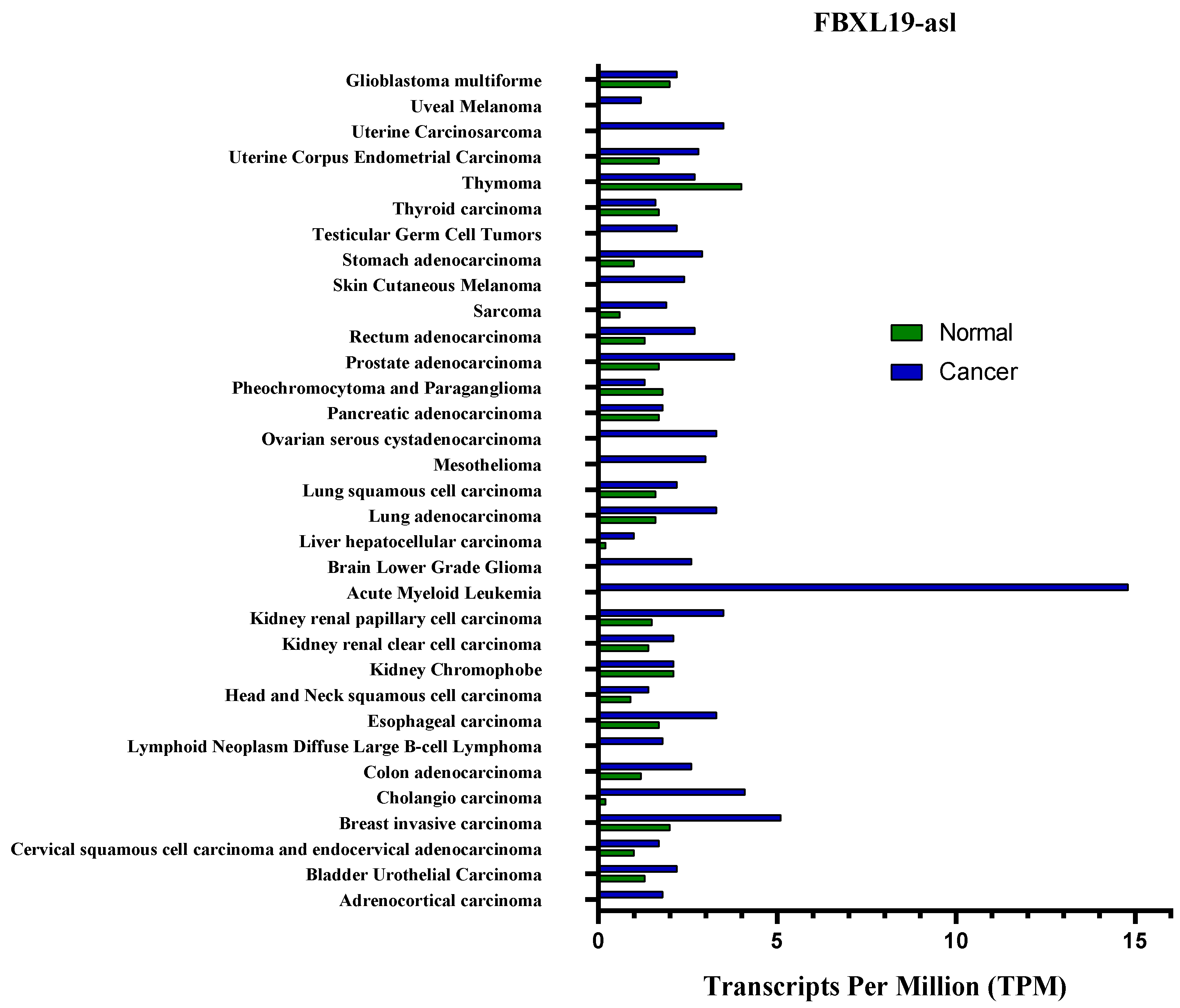
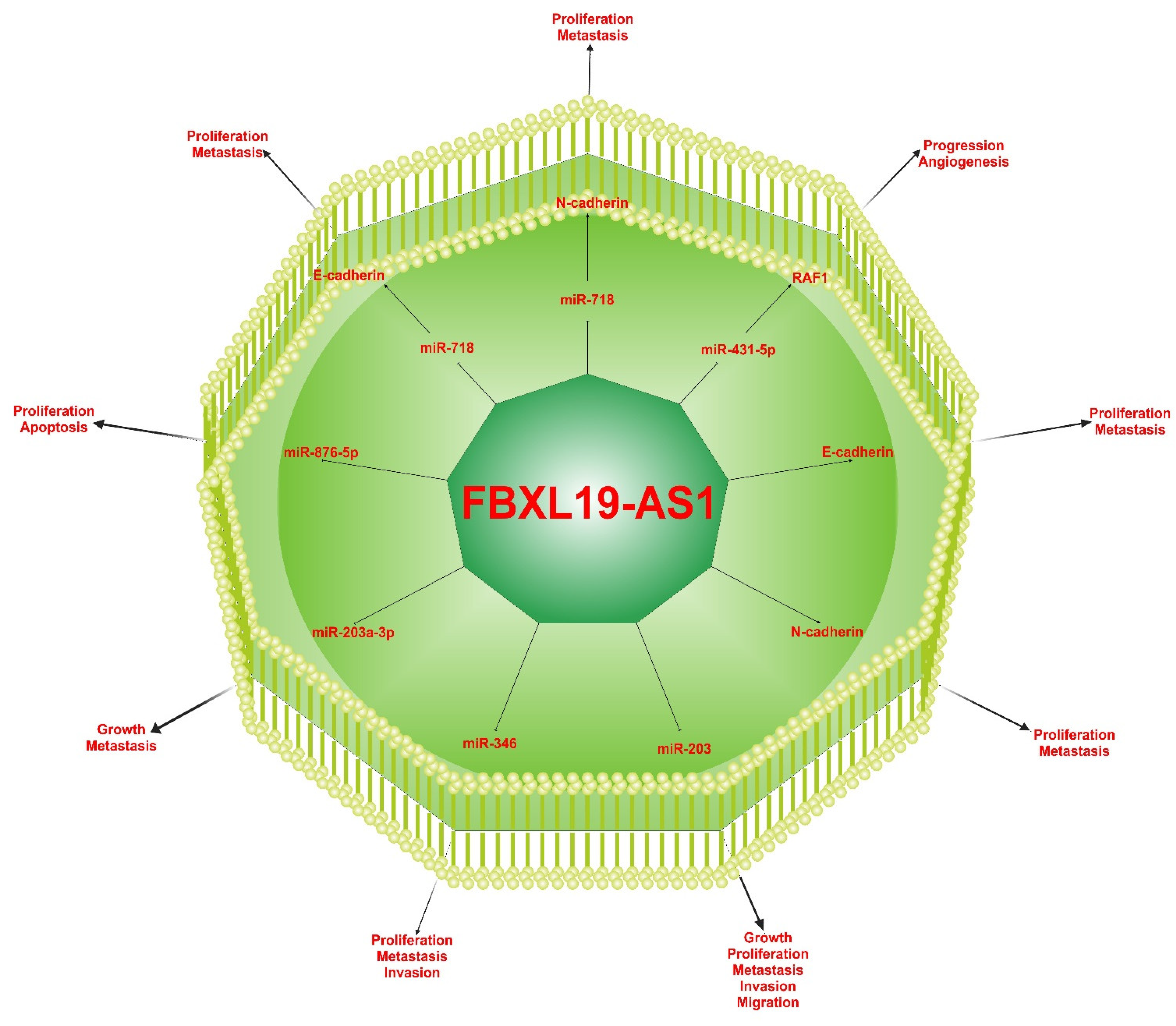
5. The Effect of FBXL19-AS1 on Several Cancers
5.1. Hepatocellular Carcinoma
5.2. Gastric Cancer
5.3. Colorectal Cancer
5.4. Glioma
5.5. Lung Cancer
5.6. Cervical Cancer
5.7. Breast Cancer
5.8. Osteosarcoma
5.9. Nasopharyngeal Carcinoma
5.10. Acute Myeloid Leukemia
6. Molecular Mechanisms of FBXL19-AS1
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Wapinski, O.; Chang, H.Y. Long noncoding RNAs and human disease. Trends Cell Biol. 2011, 21, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, Z.; Wang, X.; Huang, Z.; He, Z.; Chen, Y. Long non-coding RNA: A new player in cancer. J. Hematol. Oncol. 2013, 6, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-Coding RNAs and their Integrated Networks. J. Integr. Bioinform. 2019, 16, 20190027. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, N.F.; Manoochehri, H.; Khoei, S.G.; Sheykhhasan, M. The Functional Role of Long Non-coding RNA UCA1 in Human Multiple Cancers: A Review Study. Curr. Mol. Med. 2021, 21, 96–110. [Google Scholar] [CrossRef]
- Fazaeli, H.; Sheikholeslami, A.; Ghasemian, F.; Amini, E.; Sheykhhasan, M. The emerging role of LncRNA FENDRR in multiple cancer: A review study. Curr. Mol. Med. 2022. [Google Scholar] [CrossRef]
- Akhbari, M.H.; Zafari, Z.; Sheykhhasan, M. Competing Endogenous RNAs (ceRNAs) in Colorectal Cancer: A Review. Expert Rev. Mol. Med. 2022, 24, e27. [Google Scholar] [CrossRef]
- Clark, M.B.; Mattick, J.S. (Eds.) Long Noncoding RNAs in Cell Biology. Seminars in Cell & Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Bhat, S.A.; Ahmad, S.M.; Mumtaz, P.T.; Malik, A.A.; Dar, M.A.; Urwat, U.; Shah, R.A.; Ganai, N.A. Long non-coding RNAs: Mechanism of action and functional utility. Non-Coding RNA Res. 2016, 1, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Mao, Y.S.; Sunwoo, H.; Zhang, B.; Spector, D.L. Direct visualization of the co-transcriptional assembly of a nuclear body by noncoding RNAs. Nat. Cell Biol. 2010, 13, 95–101. [Google Scholar] [CrossRef]
- Zhao, J.; Sun, B.K.; Erwin, J.A.; Song, J.-J.; Lee, J.T. Polycomb Proteins Targeted by a Short Repeat RNA to the Mouse X Chromosome. Science 2008, 322, 750–756. [Google Scholar] [CrossRef]
- Sado, T.; Hoki, Y.; Sasaki, H. Tsix Silences Xist through Modification of Chromatin Structure. Dev. Cell 2005, 9, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Hogg, J.R.; Collins, K. Structured non-coding RNAs and the RNP Renaissance. Curr. Opin. Chem. Biol. 2008, 12, 684–689. [Google Scholar] [CrossRef] [Green Version]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.C.; Hänzelmann, S.; Sentürk Cetin, N.; Frank, S.; Zajzon, B.; Derks, J.P.; Akhade, V.S.; Ahuja, G.; Kanduri, C.; Grummt, I.; et al. Detection of RNA-DNA binding sites in long noncoding RNAs. Nucleic Acids Res. 2019, 47, e32. [Google Scholar] [CrossRef] [Green Version]
- Gao, N.; Li, Y.; Li, J.; Gao, Z.; Yang, Z.; Li, Y.; Liu, H.; Fan, T. Long Non-Coding RNAs: The Regulatory Mechanisms, Research Strategies, and Future Directions in Cancers. Front. Oncol. 2020, 10, 598817. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118, Correction in 2021, 22, 159. https://doi.org/10.1038/s41580-021-00330-4. [Google Scholar] [CrossRef]
- Yan, H.; Bu, P. Non-coding RNA in cancer. Essays Biochem. 2021, 65, 625–639. [Google Scholar] [CrossRef]
- Yang, G.; Lu, X.; Yuan, L. LncRNA: A link between RNA and cancer. Biochimica et Biophysica Acta (BBA)-Gene Regulatory Mechanisms. 2014, 1839, 1097–1109. [Google Scholar] [CrossRef]
- Martens-Uzunova, E.S.; Böttcher, R.; Croce, C.M.; Jenster, G.; Visakorpi, T.; Calin, G.A. Long Noncoding RNA in Prostate, Bladder, and Kidney Cancer. Eur. Urol. 2013, 65, 1140–1151. [Google Scholar] [CrossRef]
- Li, G.; Shi, H.; Wang, X.; Wang, B.; Qu, Q.; Geng, H.; Sun, H. Identification of diagnostic long non-coding RNA biomarkers in patients with hepatocellular carcinoma. Mol. Med. Rep. 2019, 20, 1121–1130. [Google Scholar] [CrossRef]
- Liu, H.; Ye, D.; Chen, A.; Tan, D.; Zhang, W.; Jiang, W.; Wang, M.; Zhang, X. A pilot study of new promising non-coding RNA diagnostic biomarkers for early-stage colorectal cancers. Clin. Chem. Lab. Med. (CCLM) 2019, 57, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Vencken, S.F.; Greene, C.M.; McKiernan, P.J. Non-coding RNA as lung disease biomarkers. Thorax 2015, 70, 501–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, T.; Gao, G.; Cao, Y. Long Noncoding RNAs as Novel Biomarkers Have a Promising Future in Cancer Diagnostics. Dis. Markers 2016, 2016, 9085195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Xiao, T.; Ou, W.; Wu, Z.; Wu, J.; Tang, J.; Tian, B.; Zhou, Y.; Su, M.; Wang, W. LncRNA SNHG16 as a potential biomarker and therapeutic target in human cancers. Biomark. Res. 2020, 8, 41. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Jeong, S.; Zhu, Y.; Chen, L.; Xia, Q. miRNA and lncRNA as biomarkers in cholangiocarcinoma(CCA). Oncotarget 2017, 8, 100819–100830. [Google Scholar] [CrossRef] [Green Version]
- Sideris, N.; Dama, P.; Bayraktar, S.; Stiff, T.; Castellano, L. LncRNAs in breast cancer: A link to future approaches. Cancer Gene Ther. 2022, 29, 1866–1877. [Google Scholar] [CrossRef]
- Bao, W.; Cao, F.; Ni, S.; Yang, J.; Li, H.; Su, Z.; Zhao, B. lncRNA FLVCR1-AS1 regulates cell proliferation, migration and invasion by sponging miR-485-5p in human cholangiocarcinoma. Oncol. Lett. 2019, 18, 2240–2247. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Meng, X.-M.; Huang, C.; Wu, B.-M.; Zhang, L.; Lv, X.-W.; Li, J. Long noncoding RNAs: Novel insights into hepatocelluar carcinoma. Cancer Lett. 2014, 344, 20–27. [Google Scholar] [CrossRef]
- Zhang, K.; Zhao, Z.; Yu, J.; Chen, W.; Xu, Q.; Chen, L. LncRNA FLVCR1-AS1 acts as miR-513c sponge to modulate cancer cell proliferation, migration, and invasion in hepatocellular carcinoma. J. Cell. Biochem. 2018, 119, 6045–6056. [Google Scholar] [CrossRef]
- Liu, Y.; Guo, G.; Zhong, Z.; Sun, L.; Liao, L.; Wang, X.; Cao, Q.; Chen, H. Long non-coding RNA FLVCR1-AS1 sponges miR-155 to promote the tumorigenesis of gastric cancer by targeting c-Myc. Am. J. Transl. Res. 2019, 11, 793–805. [Google Scholar]
- Han, Y.; Wang, X.; Mao, E.; Shen, B.; Huang, L. lncRNA FLVCR1-AS1 drives colorectal cancer progression via modulation of the miR-381/RAP2A axis. Mol. Med. Rep. 2020, 23, 139. [Google Scholar] [CrossRef]
- Cui, T.; Gui, D.; Gu, C.; Yan, S.; Yue, Y.; Zhang, J.; Sun, H.; Fang, Y.; Jiang, N. Long Non-Coding RNA FLVCR1-AS1 Acts as miR-493-3p Sponge to Modulate Cancer Cell Proliferation, Invasion and Migration in Colorectal Cancer. J. Biomater. Tissue Eng. 2020, 10, 306–314. [Google Scholar] [CrossRef]
- Yan, Z.; Zhang, W.; Xiong, Y.; Wang, Y.; Li, Z. Long noncoding RNA FLVCR1-AS1 aggravates biological behaviors of glioma cells via targeting miR-4731-5p/E2F2 axis. Biochem. Biophys. Res. Commun. 2019, 521, 716–720. [Google Scholar] [CrossRef]
- Gao, W.; Li, H.; Liu, Y.; Zhang, Y.; Zhao, H.; Liu, F. Long non-coding RNA FLVCR1-AS1 promotes glioma cell proliferation and invasion by negatively regulating miR-30b-3p. Mol. Med. Rep. 2020, 22, 723–732. [Google Scholar] [CrossRef]
- Gao, X.; Zhao, S.; Yang, X.; Zang, S.; Yuan, X. Long non-coding RNA FLVCR1-AS1 contributes to the proliferation and invasion of lung cancer by sponging miR-573 to upregulate the expression of E2F transcription factor 3. Biochem. Biophys. Res. Commun. 2018, 505, 931–938. [Google Scholar] [CrossRef]
- Lin, H.; Shangguan, Z.; Zhu, M.; Bao, L.; Zhang, Q.; Pan, S. lncRNA FLVCR1-AS1 silencing inhibits lung cancer cell proliferation, migration, and invasion by inhibiting the activity of the Wnt/beta-catenin signaling pathway. J. Cell. Biochem. 2019, 120, 10625–10632. [Google Scholar] [CrossRef]
- Yan, H.; Li, H.; Silva, M.A.; Guan, Y.; Yang, L.; Zhu, L.; Zhang, Z.; Li, G.; Ren, C. LncRNA FLVCR1-AS1 mediates miR-513/YAP1 signaling to promote cell progression, migration, invasion and EMT process in ovarian cancer. J. Exp. Clin. Cancer Res. 2019, 38, 356. [Google Scholar] [CrossRef] [Green Version]
- Pan, Z.; Ding, J.; Yang, Z.; Li, H.; Ding, H.; Chen, Q. LncRNA FLVCR1-AS1 promotes proliferation, migration and activates Wnt/β-catenin pathway through miR-381-3p/CTNNB1 axis in breast cancer. Cancer Cell Int. 2020, 20, 214. [Google Scholar] [CrossRef]
- Jiang, S.; Kong, P.; Liu, X.; Yuan, C.; Peng, K.; Liang, Y. LncRNA FLVCR1-AS1 accelerates osteosarcoma cells to proliferate, migrate and invade via activating wnt/β-catenin pathway. J. BUON Off. J. Balk. Union Oncol. 2020, 25, 2078–2085. [Google Scholar]
- Lin, J.; Zhai, S.; Zou, S.; Xu, Z.; Zhang, J.; Jiang, L.; Deng, X.; Chen, H.; Peng, C.; Zhang, J.; et al. Positive feedback between lncRNA FLVCR1-AS1 and KLF10 may inhibit pancreatic cancer progression via the PTEN/AKT pathway. J. Exp. Clin. Cancer Res. 2021, 40, 316. [Google Scholar] [CrossRef]
- Hao, J.; Jia, Y.U.; Zi-Yu, W.; Ou, Q.; Jun, W.; Jiang, H.; Zhao-Lian, L.I. Mechanism of flavonoids of Sophorae Fructus in inhibiting proliferation, migration and invasion of hepatocellular carcinoma cells by regulating LncRNA FBXL19-AS1/miR-342-3p pathway. Zhongguo Zhong Yao Za Zhi 2020, 45, 4440–4447. (In Chinese) [Google Scholar] [CrossRef] [PubMed]
- He, D.; Zhang, X.; Zhu, X.; Maharjan, N.; Wang, Y.; Luo, P.; Liang, C.; Tu, J. Role of FBXL19-AS1 in hepatocellular carcinoma by lncRNA–miRNA–mRNA network analysis and its diagnostic and prognostic value. Res. Sq. 2020. [Google Scholar] [CrossRef]
- He, D.; Zhang, X.; Zhu, X.; Maharjan, N.; Wang, Y.; Luo, P.; Liang, C.; Tu, J. Identify and Validate the Transcriptomic, Functional Network, and Predictive Validity of FBXL19-AS1 in Hepatocellular Carcinoma. Front. Oncol. 2020, 10, 609601. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, L. FBXL19-AS1 aggravates the progression of hepatocellular cancer by downregulating KLF2. J. BUON 2021, 26, 1333–1339. [Google Scholar]
- Wang, X.L.; Wang, J.L.; Ma, W.; Jiang, Y.; Wan, J.H.; Jiang, S.J.; He, Z.K.; Wang, X.J.; An, Z.Y.; Liu, X.X.; et al. Long non-coding RNA FBXL19-AS1 serves as a competing endogenous RNA to regulate ZEB1 expression by sponging miR-431 in gastric cancer. J. Biol. Regul. Homeost. Agents 2020, 34, 1847–1855. [Google Scholar]
- Ji, R.X.; Ren, F.; Liu, X.Q.; Yuan, X. LncRNA FBXL19-AS1 promotes the development of gastric cancer by regulating miR-876-5p/HMGB3 axis. J. Biol. Regul. Homeost. Agents 2020, 34, 1513–1518. [Google Scholar] [CrossRef]
- Shen, B.; Yuan, Y.; Zhang, Y.; Yu, S.; Peng, W.; Huang, X.; Feng, J. Long non-coding RNA FBXL19-AS1 plays oncogenic role in colorectal cancer by sponging miR-203. Biochem. Biophys. Res. Commun. 2017, 488, 67–73. [Google Scholar] [CrossRef]
- Liu, X.; Wu, P.; Su, R.; Xue, Y.; Yang, C.; Wang, D.; Ruan, X.; Zheng, J.; Yang, Y.; Li, Z.; et al. IGF2BP2 stabilized FBXL19-AS1 regulates the blood-tumour barrier permeability by negatively regulating ZNF765 by STAU1-mediated mRNA decay. RNA Biol. 2020, 17, 1777–1788. [Google Scholar] [CrossRef]
- Yu, D.-J.; Li, Y.-H.; Zhong, M. LncRNA FBXL19-AS1 promotes proliferation and metastasis via regulating epithelial-mesenchymal transition in non-small cell lung cancer. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4800–4806. [Google Scholar]
- Jiang, Q.; Cheng, L.; Ma, D.; Zhao, Y. FBXL19-AS1 exerts oncogenic function by sponging miR-431-5p to regulate RAF1 expression in lung cancer. Biosci. Rep. 2019, 39, BSR20181804. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, X.; Liu, Y.; Xu, S. Long noncoding RNA FBXL19—AS1 induces tumor growth and metastasis by sponging miR—203a—3p in lung adenocarcinoma. J. Cell. Physiol. 2019, 235, 3612–3625. [Google Scholar] [CrossRef]
- Huang, X.; Shi, H.; Shi, X.; Jiang, X. LncRNA FBXL19-AS1 promotes proliferation and metastasis of cervical cancer through upregulating COL1A1 as a sponge of miR-193a-5p. J. Biol. Res. 2021, 28, 20. [Google Scholar] [CrossRef]
- Wan, S.; Ni, G.; Ding, J.; Huang, Y. Long Noncoding RNA FBXL19-AS1 Expedites Cell Growth, Migration and Invasion in Cervical Cancer by miR-193a-5p/PIN1 Signaling. Cancer Manag. Res. 2020, 12, 9741–9752. [Google Scholar] [CrossRef]
- Ding, Z.; Ye, P.; Yang, X.; Cai, H. LncRNA FBXL19-AS1 promotes breast cancer cells proliferation and invasion via acting as a molecular sponge to miR-718. Biosci. Rep. 2019, 39, BSR20182018. [Google Scholar] [CrossRef] [Green Version]
- Dong, G.; Pan, T.; Zhou, D.; Li, C.; Liu, J.; Zhang, J. FBXL19-AS1 promotes cell proliferation and inhibits cell apoptosis via miR-876-5p/FOXM1 axis in breast cancer. Acta Biochim. Et Biophys. Sin. 2019, 51, 1106–1113. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, X.; Zhou, W.; Hu, J.; Zhou, D. LIN28A-stabilized FBXL19-AS1 promotes breast cancer migration, invasion and EMT by regulating WDR66. Vitr. Cell. Dev. Biol.-Animal 2019, 55, 426–435. [Google Scholar] [CrossRef]
- Pan, R.; He, Z.; Ruan, W.; Li, S.; Chen, H.; Chen, Z.; Liu, F.; Tian, X.; Nie, Y. lncRNA FBXL19-AS1 regulates osteosarcoma cell proliferation, migration and invasion by sponging miR-346. OncoTargets Ther. 2018, 11, 8409–8420. [Google Scholar] [CrossRef] [Green Version]
- Dong, H.; Huang, C.; Huang, J. FBXL19-AS1 promotes the progression of nasopharyngeal carcinoma by acting as a competing endogenous RNA to sponge miR-431 and upregulate PBOV1. Mol. Med. Rep. 2021, 24, 647. [Google Scholar] [CrossRef]
- Sheng, H.; Zhang, J.; Ma, Y.; Zhang, Y.; Dai, Y.; Jiang, R. lncRNA FBXL19—AS1 is a diagnosis biomarker for paediatric patients with acute myeloid leukemia. J. Gene Med. 2021, 23, e3317. [Google Scholar] [CrossRef]
- Arnold, M.; Abnet, C.C.; Neale, R.E.; Vignat, J.; Giovannucci, E.L.; McGlynn, K.A.; Bray, F. Global Burden of 5 Major Types of Gastrointestinal Cancer. Gastroenterology 2020, 159, 335–349. [Google Scholar] [CrossRef]
- Chiang, T.H.; Chang, W.J.; Chen, S.L.S.; Yen, A.M.F.; Fann, J.C.Y.; Chiu, S.Y.H.; Chen, Y.R.; Chuang, S.L.; Shieh, C.F.; Liu, C.Y.; et al. Mass eradication of Helicobacter pylori to reduce gastric cancer incidence and mortality: A long-term cohort study on Matsu Islands. Gut 2021, 70, 243–250. [Google Scholar] [PubMed]
- Wang, Q.-X.; Zhu, Y.-Q.; Zhang, H.; Xiao, J. Altered MiRNA Expression in Gastric Cancer: A Systematic Review and Meta-Analysis. Cell. Physiol. Biochem. 2015, 35, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Shin, V.Y.; Chu, K.M. MiRNA as potential biomarkers and therapeutic targets for gastric cancer. World J. Gastroenterol. 2014, 20, 10432–10439. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.; Min, B.-H.; Jang, J.; Kang, S.Y.; Bae, H.; Jang, S.S.; Kim, J.-I.; Kim, K.-M. MicroRNA Expression Profiles in Gastric Carcinogenesis. Sci. Rep. 2018, 8, 14393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, N.-B.; He, Y.-F.; Li, X.-Q.; Wang, K.; Wang, R.-L. The role of miRNA and lncRNA in gastric cancer. Oncotarget 2017, 8, 81572–81582. [Google Scholar] [CrossRef]
- Sheykhhasan, M.; Ahmadyousefi, Y.; Seyedebrahimi, R.; Tanzadehpanah, H.; Manoochehri, H.; Dama, P.; Hosseini, N.F.; Akbari, M.; Farsani, M.E. DLX6-AS1: A putative lncRNA candidate in multiple human cancers. Expert Rev. Mol. Med. 2021, 23, E17. [Google Scholar] [CrossRef]
- Mesfin, F.B.; Al-Dhahir, M.A. Gliomas. 2022 Apr 9. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- DeSantis, C.; Ma, J.; Bryan, L.; Jemal, A. Breast cancer statistics, 2013. CA A Cancer J. Clin. 2014, 64, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Stuckey, A. Breast cancer: Epidemiology and risk factors. Clin. Obstet. Gynecol. 2011, 54, 96–102. [Google Scholar] [CrossRef]
- Xu, S.; Gong, Y.; Yin, Y.; Xing, H.; Zhang, N. The multiple function of long noncoding RNAs in osteosarcoma progression, drug resistance and prognosis. Biomed. Pharmacother. 2020, 127, 110141. [Google Scholar] [CrossRef]
- Li, Z.; Hong, S.; Liu, Z. LncRNA LINC00641 predicts prognosis and inhibits bladder cancer progression through miR-197-3p/KLF10/PTEN/PI3K/AKT cascade. Biochem. Biophys. Res. Commun. 2018, 503, 1825–1829. [Google Scholar] [CrossRef]
- Yang, N.; Chen, J.; Zhang, H.; Wang, X.; Yao, H.; Peng, Y.; Zhang, W. LncRNA OIP5-AS1 loss-induced microRNA-410 accumulation regulates cell proliferation and apoptosis by targeting KLF10 via activating PTEN/PI3K/AKT pathway in multiple myeloma. Cell Death Dis. 2017, 8, e2975. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Yang, X.; Ni, H. Long non-coding RNA FLVCR1-AS1 functions as a ceRNA to aggravate cervical cancer cell growth by the miR-381-3p/MAGT1 axis. Arch. Gynecol. Obstet. 2022, 306, 2093–2103. [Google Scholar] [CrossRef]
- Kent, L.N.; Leone, G. The broken cycle: E2F dysfunction in cancer. Nat. Rev. Cancer 2019, 19, 326–338. [Google Scholar] [CrossRef]
- Shi, Y.-B.; Liu, S.-L.; Mou, X.-R.; Liao, J.; Che, J.-P.; Fei, X.-Q.; Wang, A.-R. Long noncoding RNA HOXA-AS2 acts as an oncogene by targeting miR-145-3p in human non-small cell lung cancer. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 1243–1249. [Google Scholar]
- Lu, W.; Wang, Q.; Fu, C. Interfering with Long Non-Coding RNA FBXL19-AS1 Inhibits the Proliferation, Invasion and Migration of Hepatoma Carcinoma Cells via Targeting miR-541-5p. J. Biomater. Tissue Eng. 2021, 11, 2120–2127. [Google Scholar] [CrossRef]
- Dong, N.; Guo, R.; Fu, A. Effects of long non-coding RNA FBXL19-AS1 targeting miR-339-3p on the proliferation, migration and invasion of pancreatic cancer cells. Chin. J. Pancreatol. 2021, 6, 187–194. [Google Scholar]







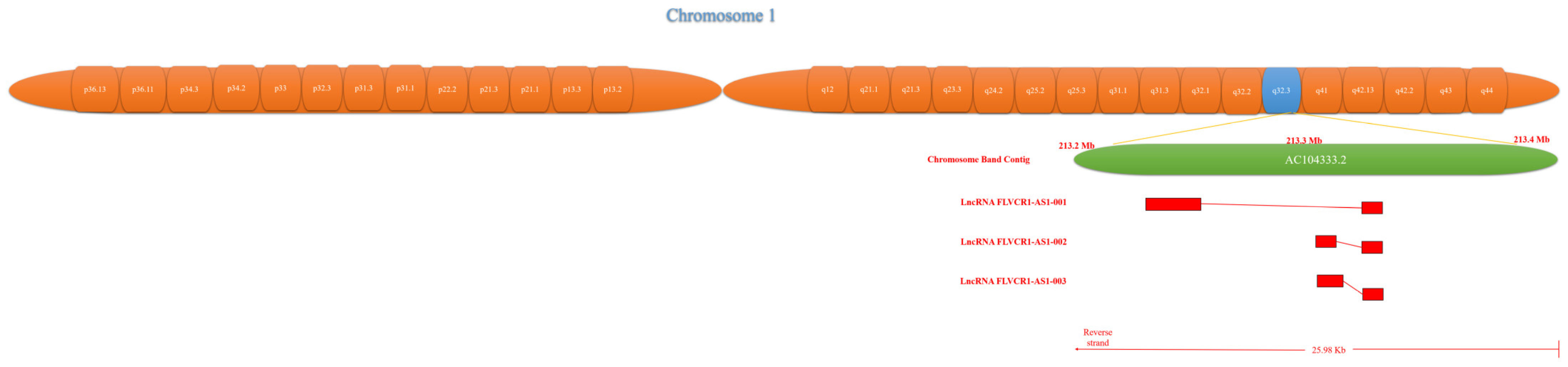{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subcellular Locations | Score |
|---|---|
| Cytoplasm | 0.0682472744361 |
| Nucleus | 0.0229142140504 |
| Ribosome | 0.0409735673257 |
| Cytosol | 0.828903123455 |
| Exosome | 0.0389618207329 |
| Subcellular Locations | Score |
|---|---|
| Cytoplasm | 0.323765378524 |
| Nucleus | 0.170541522227 |
| Ribosome | 0.275374297237 |
| Cytosol | 0.075880246739 |
| Exosome | 0.154438555273 |
| Cancer Type | Number of Case | Assessed Cell Lines | Interacting Genes and Proteins | Novel Therapeutic/Diagnostic | Expression | Function | Years Reference |
|---|---|---|---|---|---|---|---|
| Gastric cancer | A total of 30 Gastric cancer tissues and their adjacent normal tissues | The human gastric epithelial cell line GES-1 and three human gastric cancer cell lines (AGS, MNK-45, and MGC-803) | miR-155, c-Myc | A novel therapeutic target for treatment of patients with GC | ↑ | Promote cell proliferation and invasion | 2019 [31] |
| Hepatocellular carcinoma | A total of 60 HCC and matched normal tissues | The normal liver cell line LO2 and the HCC cell lines(Hep3B, HepG2, Huh7, and PLC/PRF-5) | miR-513c | A new target in HCC prevention and treatment | ↑ | Promote cell proliferation, migration, and invasion | 2018 [30] |
| Lung cancer | A total of 62 NSCLC and adjacent normal lung tissues | Five NSCLC cell lines (H358, A549, H520, H1299, and SKMES1), human bronchial epithelial cell line (BEAS-2B), and human embryonic kidney (HEK) 293T cell line | miR-573, E2F transcription factor 3 | A novel diagnostic biomarker and therapeutic target for NSCLC | ↑ | Promote cell proliferation and invasion | 2018 [36] |
| Lung cancer | A total of 29 lung cancer tissues and their adjacent normal tissues | Human bronchial epithelial cell line (16HBE) and human lung cancer cell lines (SPCA1, A549, and H1299) | Wnt/β-catenin signaling pathway | A novel targets for the treatment of lung cancer | ↑ | Promote cell proliferation, migration, and invasion | 2019 [37] |
| Glioma | A total of 51 glioma tissues and adjacent normal tissues | Glioma cell lines and human astrocyte cell line (NHA) | miR-4731-5p/E2F2 | A potential target for glioma tumor therapy | ↑ | Promote cell proliferation, migration, and invasion | 2020 [34] |
| Glioma | A total of 50 glioblastoma tissues and adjacent normal tissues | The human GBM cell lines U251, T98G, LN229 and SHG44 and normal human astrocyte (NHA) cells | miR-30b-3p | A novel therapeutic target and diagnostic biomarker for glioblastoma | ↑ | Promote cell proliferation and invasion | 2020 [35] |
| Breast cancer | - | Human normal breast epithelial cell (MCF-10A) and BC cells (MDA-MB-231, T47D, BT-474, SKBR3, MCF7) | Wnt/β-catenin pathway, miR-381-3p, CTNNB1 | A promising target for breast cancer therapy | ↑ | Promote cell proliferation, migration, invasion | 2020 [39] |
| Ovarian cancer | A total of 50 ovarian serous cancer tissues and adjacent normal tissues | SKOV3 and OVCAR3 cells | miR-513, YAP1 | A potential therapeutic target for human ovarian cancer | ↑ | Promote cell progression, migration, invasion, and EMT process | 2019 [38] |
| Cholangiocarcinoma | A total of 22 cholangiocarcinoma and adjacent normal tissues | Human cell lines, including the noncancerous cholangiocyte cell line HIBEC and the CCA cell lines RBE, CCLP1, HuCCT1, and HCCC-9810 | miR-485-5p | A novel therapeutic target and a potential diagnostic marker for cholangiocarcinoma | ↑ | Promote cell proliferation, migration, and invasion | 2019 [28] |
| Colorectal cancer | A total of 26 pairs of colorectal cancer tissues and adjacent non-tumor tissues | Four human CRC cell lines, namely Caco-2, SW480, LoVo, and SW1116, the NCM460 normal colonic epithelial cell line and 293T cells | miR-381, RAP2A | - | ↑ | Promote cell viability, apoptosis, migration, and invasion | 2020 [32] |
| Colorectal cancer | - | Human CRC cell lines, namely Caco-2, SW480 | miR-493-3p | - | ↑ | Promote cell proliferation, invasion, and migration | 2020 [33] |
| Osteosarcoma cells | A total of 48 osteosarcoma tissues and adjacent non-tumor tissues | Normal osteoblasts (hFOB1.19) and U2OS and MG63 osteosarcoma cell lines | Wnt/β-catenin pathway, CTNNB1, SOX4, CCND1, CCND2, MYC and nucleus β-catenin | - | ↑ | Promote cell proliferation, invasion, and migration | 2020 [40] |
| pancreatic cancer | A total of 77 samples of human pancreatic cancer tissues and corresponding normal tissues | Human PC cell lines (Bxpc-3, CFPAC-1, MIA PaCa-2,PANC-1, and PATU-8988) and human normal pancreatic ductal epithelial cells (HPNE) | KLF10 and PTEN/AKT pathway | A noveltherapeutic strategy for PC treatment | ↑ | Inhibit proliferation, cell cycle, and migration | 2021 [41] |
| Cervical cancer | - | Cervical cancer cell | miR-381, MAGT1 | - | ↑ | Promote cell growth | 2022 [74] |
| Cancer Type | Number of Case | Assessed Cell Lines | Interacting Genes and Proteins | Novel Therapeutic/Diagnostic | Expression | Function | Years Reference |
|---|---|---|---|---|---|---|---|
| Gastric cancer | - | - | miR-876-5p, HMGB3 | - | ↑ | Promote cell development | 2020 [47] |
| Gastric cancer | - | - | miR-431, ZEB1 | - | ↑ | - | 2020 [76] |
| Breast cancer | A total of 49 breast cancer tissues and adjacent normal tissues | Human BC cell lines (MDA-MB-231, ZR-75-1, MCF-7, BT-549, MDA-MB-468, and T47D) and the normal mammary fibroblast cell line (Hs578Bst) | miR-718 | A potential therapeutic target for breast cancer treatment | ↑ | Promote cell proliferation and invasion | 2019 [55] |
| Breast cancer | - | Non-carcinogenic epithelial cells (MCF-10A) and breast cancer cells (MCF-7, BT-549, MDA-MB-231, and SKBR3) | miR-876-5p, Forkhead box M1 (FOXM1) | A therapeutic approach for treating breast cancer | ↑ | Promote cell proliferation and apoptosis | 2019 [56] |
| Breast cancer | - | Normal cell line HS578Bst, breast cancer cell lines (SK-BR3, BT474, MCF-7, and MDA-MB-231) | lin-28 homolog A (LIN28A), WD repeat domain 66 (WDR66) | A new biological marker in breast cancer | ↑ | Promote cell migration, invasion, and EMT | 2019 [57] |
| Colorectal cancer | A total of 50 human colorectal cancer tissues and their adjacent non-tumor tissues | CRC cell lines (LoVo,HT29,HCT116, and SW620) and normal colon epithelial FHC cel | miR-203 | A new insight for understanding CRC development | ↑ | Promote cell proliferation, migration, and invasion | 2017 [48] |
| Lung cancer | A total of 84 lung cancer tissues and adjacent non-tumor tissues | Lung cancer cell lines (A549, H1975, SPC-A-1, H125, and H1299) and normal human lung cells (MRC-5) | miR-431-5p, RAF1 | A new insight into the therapeutic strategies of lung cancer | ↑ | Promote cell proliferation, migration, invasion, and angiogenesis | 2019 [51] |
| Non-small cell lung cancer | A total of 52 lung cancer tissues and adjacent non-tumor tissues | Five kinds of NSCLC cell lines (A549, H1299, H520, SPCA1, and H358) and normal lung epithelial cells (16HBE) | E-cadherin, N-cadherin, vimentin | - | ↑ | Promote cell proliferation and metastasis | 2019 [50] |
| Lung adenocarcinoma | The lung adenocarcinoma and matched normal adjacent tissue samples | Two human lung adenocarcinoma cells, NCI-H1975 and SPC | miR-203a-3p | A potential prognostic marker and a therapeutic target for patients with lung adenocarcinoma | ↑ | Promote tumor growth and metastasis | 2020 [52] |
| Osteosarcoma | - | Human osteosarcomacell lines MG63, U2OS, SAOS2, HOS, 143B, and the normal osteoblast cell line hFOB1.19 | miR-346 | A novel therapeutic target for osteosarcoma | ↑ | Promote cell proliferation, migration, and invasion | 2018 [58] |
| Cervical cancer | A total of 100 cervical cancer tissues and adjacent non-tumor tissues | Human normal cervical cell line (Ect1/E6E7) and cervical cancer cell lines (C-4-I, SiHa, C-33A and HeLa) | miR-193a-5p, PIN1 | A new direction for treating patients with cervical cancer | ↑ | Promote cell growth, migration, and invasion | 2020 [54] |
| Cervical cancer | A total of 46 cervical cancer tissues and adjacent non-tumor tissues | Human healthy cervical cells (HUCEC) and human cervical cancer cells (HeLa, Caski, C-33 A, AV3) | COL1A1, miR-193a-5p | - | ↑ | Promote cell proliferation and metastasis | 2021 [53] |
| Hepatocellular carcinoma | A total of 57 hepatocellular carcinoma tissues and adjacent non-tumor tissues, and whole blood samples of 92 patients with hepatocellular carcinoma | - | - | A potential biomarker for HCC diagnosis and prognosis | ↑ | Promote cell occurrence and development | 2020 [44] |
| Hepatocellular carcinoma | - | - | miR-342-3p | - | ↑ | Promote cell proliferation, migration, and invasion | 2020 [42] |
| Hepatocellular carcinoma | A total of 60 hepatocellular carcinoma tissues and adjacent non-tumor tissues | Normal liver cell line and HCC cell lines | KLF2 | - | ↑ | Promote tumor progression | 2021 [45] |
| Hepatocellular carcinoma | A total of hepatocellular carcinoma tissues and adjacent non-tumor tissues | Normal liver cell line and HCC cell lines | miR-541-5p | - | ↑ | Promote cell proliferation, invasion, and migration | 2021 [77] |
| Glioma | - | The human brain microvascular endothelial cell line hcMEC/D3 (ECs) | ZNF765, STAU1 | A new potential therapeutic strategy for glioma | ↑ | Promote cell apoptosis | 2020 [49] |
| Nasopharyngeal carcinoma | A total of 30 nasopharyngeal carcinoma tissues and adjacent non-tumor tissues | Human NPC cell lines (C666-1, SUNE1, 5–8F and 6–10B) and nasopharyngeal epithelial cells (NP69) | miR-431, PBOV1 | A novel therapeutic target for nasopharyngeal carcinoma | ↑ | Promote tumor progression | 2021 [59] |
| Acute myeloid leukemia | Serum samples of 137 acute myeloid leukemia patients and 43 healthy controls | - | - | A novel prognostic and diagnostic biomarker for acute myeloid leukemia patients | ↑ | - | 2021 [60] |
| Pancreatic cancer | A total of 73 pancreatic cancer tissues and adjacent non-tumor tissues | Normal pancreatic epithelial cells (hTERT-HPNE) and pancreatic cancer cell lines (Capan-1, SW1990, PaTu8988) | miR-339-3p | - | ↑ | Promote cell proliferation, migration, and invasion | 2021 [78] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheykhhasan, M.; Tanzadehpanah, H.; Ahmadieh Yazdi, A.; Mahaki, H.; Seyedebrahimi, R.; Akbari, M.; Manoochehri, H.; Kalhor, N.; Dama, P. FLVCR1-AS1 and FBXL19-AS1: Two Putative lncRNA Candidates in Multiple Human Cancers. Non-Coding RNA 2023, 9, 1. https://doi.org/10.3390/ncrna9010001
Sheykhhasan M, Tanzadehpanah H, Ahmadieh Yazdi A, Mahaki H, Seyedebrahimi R, Akbari M, Manoochehri H, Kalhor N, Dama P. FLVCR1-AS1 and FBXL19-AS1: Two Putative lncRNA Candidates in Multiple Human Cancers. Non-Coding RNA. 2023; 9(1):1. https://doi.org/10.3390/ncrna9010001
Chicago/Turabian StyleSheykhhasan, Mohsen, Hamid Tanzadehpanah, Amirhossein Ahmadieh Yazdi, Hanie Mahaki, Reihaneh Seyedebrahimi, Mohammad Akbari, Hamed Manoochehri, Naser Kalhor, and Paola Dama. 2023. "FLVCR1-AS1 and FBXL19-AS1: Two Putative lncRNA Candidates in Multiple Human Cancers" Non-Coding RNA 9, no. 1: 1. https://doi.org/10.3390/ncrna9010001