
Exploring Differences in Culturable Fungal Diversity Using Standard Freezing Incubation—A Case Study in the Limestones of Lemos Pantheon (Portugal)

 ,
,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Sampling Strategy
2.2. Isolation, Morphological and Molecular Identification
2.3. Data Analysis
3. Results and Discussion
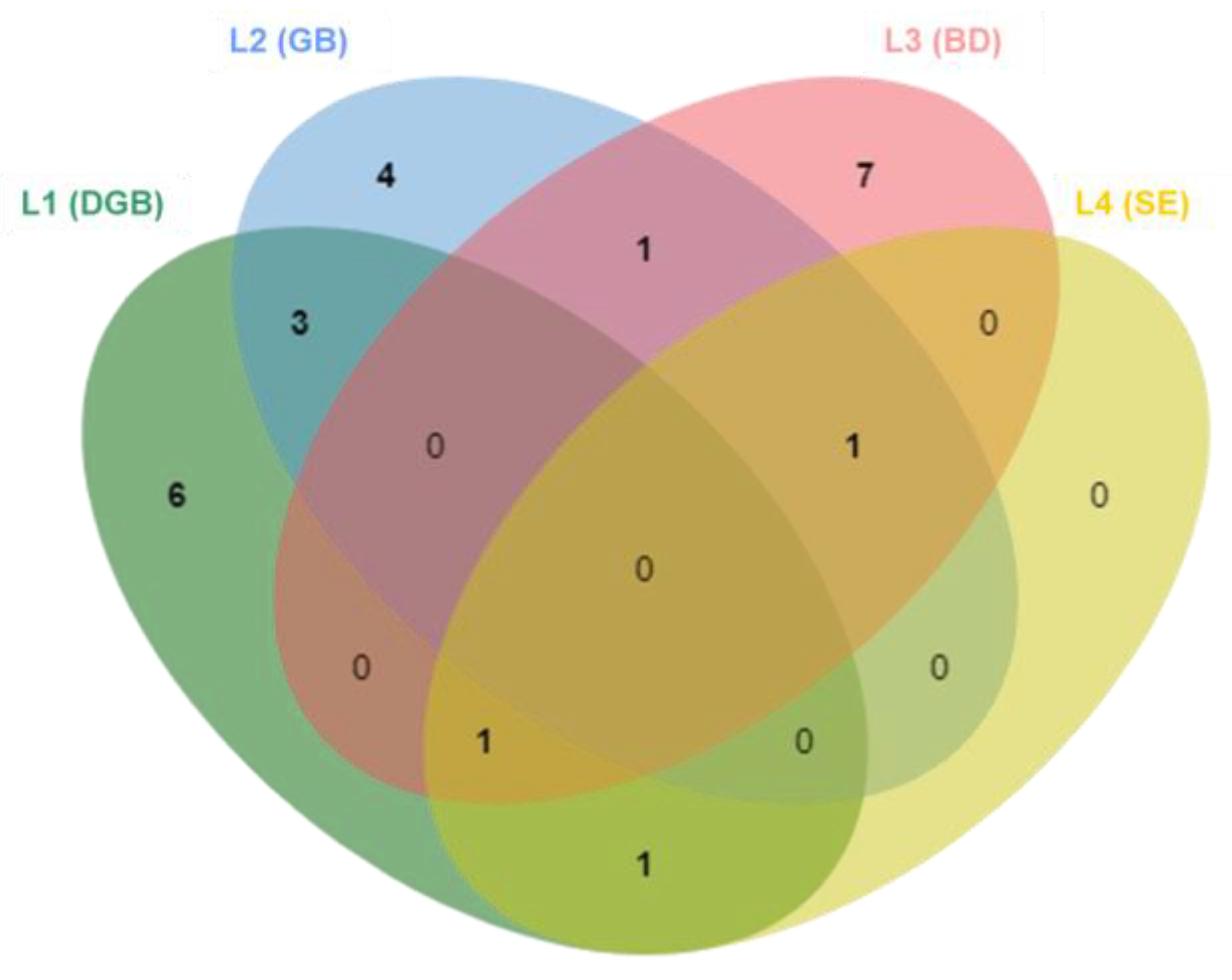
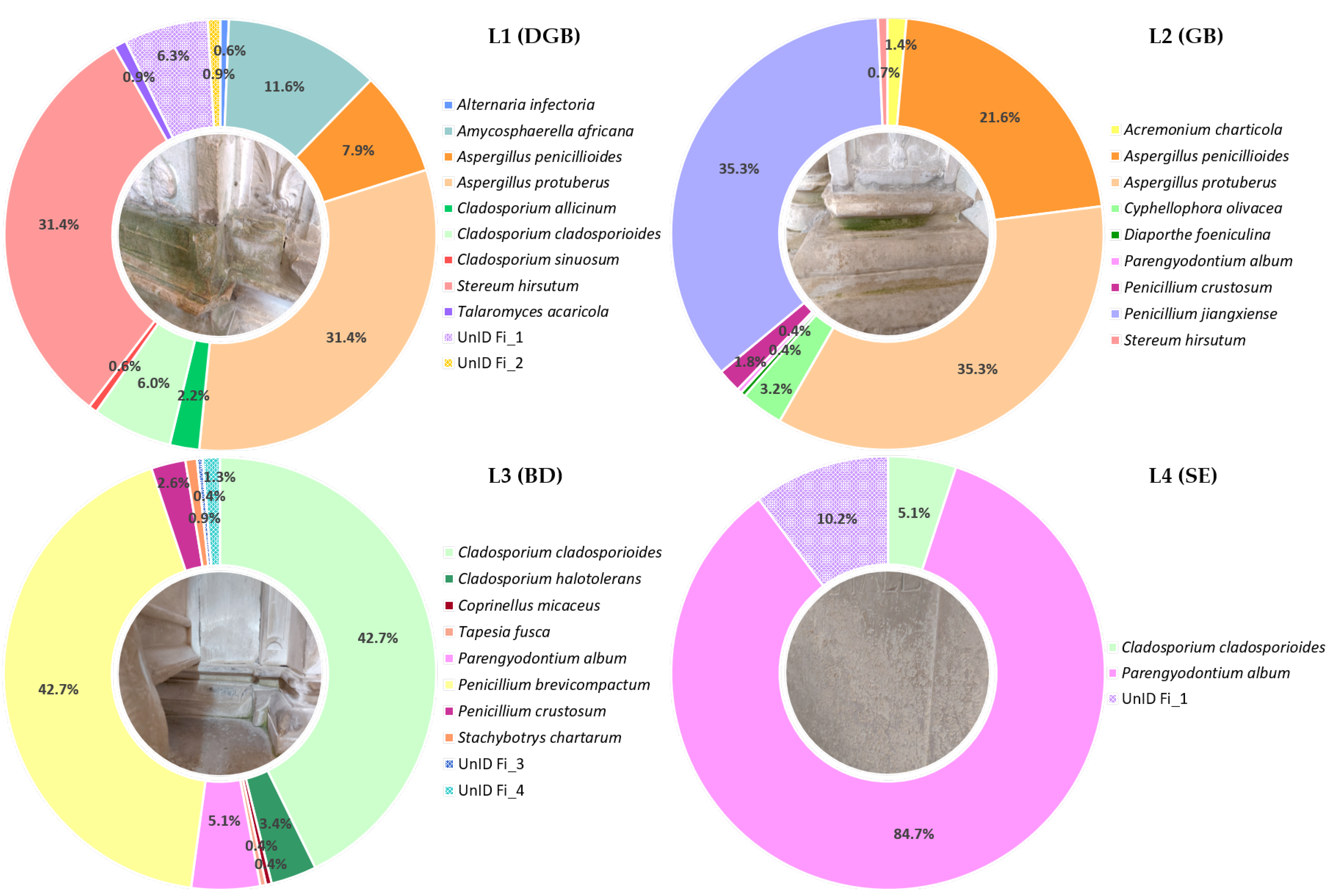
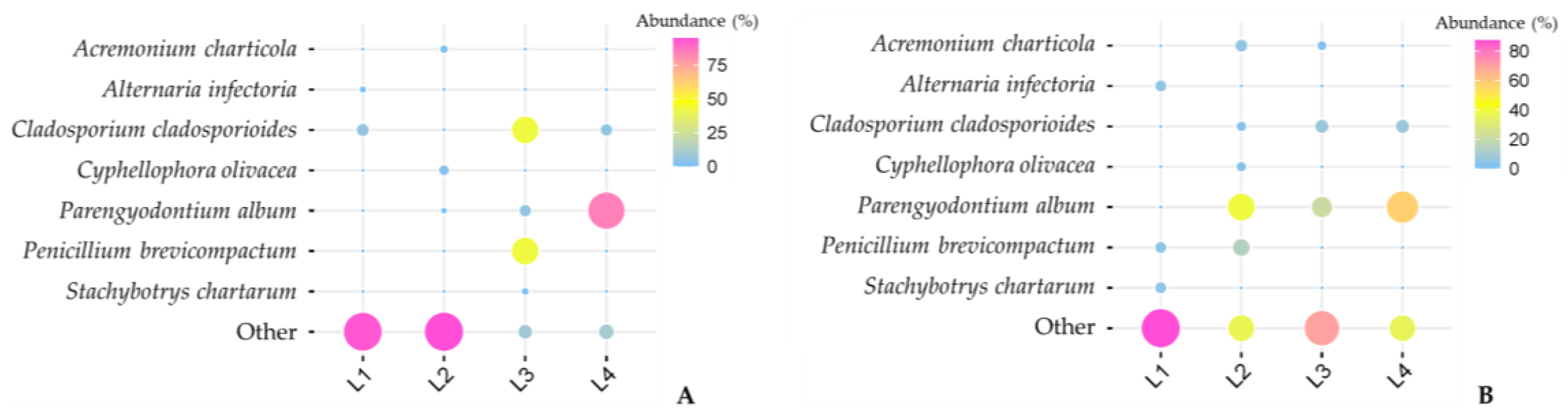
3.1. Frozen Diversity and Its Biodeteriorative Potential
3.2. Diversity vs. Isolation Method
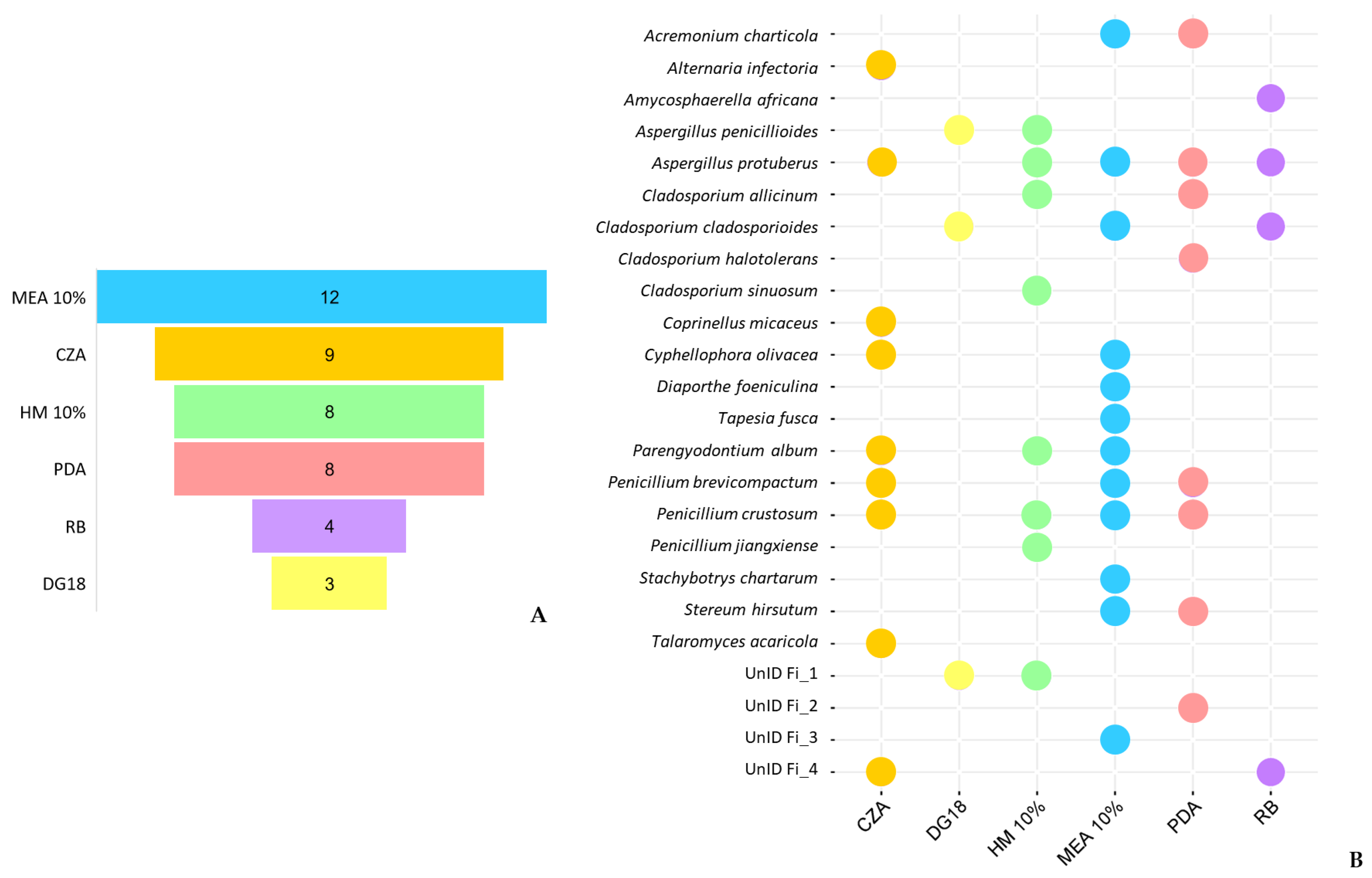
3.3. Effect of Isolation Media
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Sample | Medium | Isolate ID | GenBank Accession Numbers | Notes | ||||
|---|---|---|---|---|---|---|---|---|---|
| ITS | Translation Elongation Factor 1α (tef1) | β-Tubulin (benA) | Calmodulin (CaM) | Partial Actin (Act) | |||||
| PL133 | L4 | MEA 10% | Parengyodontium album | OQ211122 | OQ330805 | – | – | – | a |
| PL134 | L3 | RB | Cladosporium cladosporioides | OQ211118 | – | – | – | OQ330822 | a |
| PL135 | L2 | PDA | Acremonium charticola | OQ211119 | OQ330803 | – | – | – | a |
| PL136 | L2 | PDA | Acremonium charticola | OQ211123 | OQ330806 | – | – | – | a |
| PL137 | L3 | MEA 10% | Cladosporium cladosporioides | – | – | – | – | – | b |
| PL138 | L3 | PDA | Penicillium brevicompactum | OQ211120 | – | OQ330764 | – | – | a |
| PL139 | L3 | CZA | Parengyodontium album | – | – | – | – | – | b |
| PL140 | L1 | PDA | Cladosporium allicinum | OQ211114 | – | – | – | OQ330820 | a |
| PL141 | L4 | HM 10% | Parengyodontium album | – | – | – | – | – | b |
| PL142 | L4 | HM 10% | UnID Fi_1 | – | – | – | – | – | d |
| PL143 | L4 | HM 10% | UnID Fi_1 | – | – | – | – | – | d |
| PL144 | L3 | MEA 10% | Tapesia fusca | OQ211115 | OQ330802 | – | – | – | c |
| PL145 | L2 | HM 10% | Penicillium jiangxiense | OQ211124 | – | OQ330765 | – | – | a |
| PL146 | L2 | MEA 10% | Acremonium charticola | – | – | – | – | – | b |
| PL147 | L1 | HM 10% | UnID Fi_1 | – | – | – | – | – | d |
| PL148 | L1 | MEA 10% | Aspergillus protuberus | – | – | – | – | – | b |
| PL149 | L2 | MEA 10% | Acremonium charticola | OQ211126 | OQ330807 | – | – | – | a |
| PL150 | L3 | CZA | Penicillium brevicompactum | OQ211125 | – | OQ330766 | – | – | a |
| PL151 | L3 | CZA | Coprinellus micaceus | OQ211121 | OQ330804 | – | – | – | c |
| PL152 | L3 | MEA 10% | Stachybotrys chartarum | OQ211138 | OQ330811 | – | – | – | a |
| PL153 | L3 | MEA 10% | Stachybotrys chartarum | – | – | – | – | – | b |
| PL154 | L2 | MEA 10% | Cyphellophora olivacea | OQ211128 | OQ330808 | – | – | – | a |
| PL155 | L2 | MEA 10% | Parengyodontium album | – | – | – | – | – | b |
| PL156 | L2 | MEA 10% | Cyphellophora olivacea | OQ211135 | OQ330810 | – | – | – | a |
| PL157 | L2 | HM 10% | Aspergillus protuberus | OQ211136 | – | OQ330771 | OQ330788 | – | a |
| PL158 | L2 | HM 10% | Aspergillus penicillioides | OQ211141 | – | OQ330774 | OQ330798 | – | a |
| PL159 | L2 | HM 10% | Penicillium crustosum | OQ211116 | – | OQ330763 | – | – | a |
| PL160 | L2 | CZ | Penicillium crustosum | OQ211142 | – | OQ330775 | – | – | a |
| PL161 | L2 | HM 10% | Penicillium crustosum | OQ211143 | – | – | – | – | a |
| PL162 | L2 | HM 10% | Aspergillus penicillioides | OQ211131 | – | OQ330769 | – | – | a |
| PL163 | L2 | HM 10% | Aspergillus penicillioides | – | – | – | – | – | b |
| PL164 | L1 | MEA 10% | Cladosporium cladosporioides | OQ211117 | – | – | – | OQ330821 | a |
| PL165 | L1 | CZA | Alternaria infectoria | OQ211140 | OQ330812 | – | – | – | a |
| PL166 | L2 | RB | Aspergillus protuberus | OQ211148 | – | OQ330779 | OQ330789 | – | a |
| PL167 | L2 | RB | Aspergillus protuberus | OQ211137 | – | OQ330772 | OQ330790 | – | a |
| PL168 | L2 | RB | Aspergillus protuberus | OQ211145 | – | OQ330776 | OQ330791 | – | a |
| PL169 | L1 | RB | Amycosphaerella africana | OQ211144 | OQ330814 | – | – | – | a |
| PL170 | L2 | CZA | Aspergillus protuberus | OQ211139 | – | OQ330773 | OQ330792 | – | a |
| PL171 | L2 | CZA | Aspergillus protuberus | OQ211147 | – | OQ330778 | OQ330793 | – | a |
| PL172 | L2 | CZA | Streptomyces | – | – | – | – | – | e |
| PL173 | L2 | CZA | Streptomyces | – | – | – | – | – | e |
| PL174 | L2 | MEA 10% | Cyphellophora olivacea | OQ211156 | OQ330816 | – | – | – | a |
| PL175 | L3 | PDA | Cladosporium halotolerans | OQ211149 | – | – | – | OQ330823 | a |
| PL176 | L2 | PDA | Streptomyces | – | – | – | – | – | e |
| PL177 | L2 | PDA | Penicillium crustosum | OQ211152 | – | OQ330781 | – | – | a |
| PL178 | L2 | PDA | Streptomyces | – | – | – | – | – | e |
| PL179 | L2 | RB | Aspergillus protuberus | – | – | – | – | – | b |
| PL180 | L1 | RB | Amycosphaerella africana | – | – | – | – | – | b |
| PL181 | L1 | HM 10% | UnID Fi_1 | – | – | – | – | – | d |
| PL182 | L2 | HM 10% | Penicillium crustosum | OQ211151 | – | OQ330780 | – | – | a |
| PL183 | L3 | RB | Cladosporium cladosporioides | OQ211150 | – | – | – | OQ330824 | a |
| PL184 | L3 | RB | Cladosporium cladosporioides | – | – | – | – | – | b |
| PL185 | L1 | RB | Amycosphaerella africana | OQ211154 | – | – | – | – | a |
| PL186 | L1 | RB | Amycosphaerella africana | OQ211155 | OQ330815 | – | – | – | a |
| PL187 | L3 | PDA | Penicillium brevicompactum | OQ211153 | – | OQ330782 | – | – | a |
| PL188 | L3 | MEA 10% | Penicillium crustosum | OQ211162 | – | – | – | – | a |
| PL189 | L3 | CZA | Parengyodontium album | OQ211129 | OQ330809 | – | – | – | a |
| PL190 | L2 | MEA 10% | Cyphellophora olivacea | OQ211161 | OQ330818 | – | – | – | a |
| PL191 | L2 | MEA 10% | Streptomyces | – | – | – | – | – | e |
| PL192 | L2 | MEA 10% | Streptomyces | – | – | – | – | – | e |
| PL193 | L3 | MEA 10% | Penicillium brevicompactum | OQ211163 | – | OQ330783 | – | a | |
| PL194 | L2 | MEA 10% | Streptomyces | – | – | – | – | – | e |
| PL195 | L2 | MEA 10% | Cyphellophora olivacea | OQ211165 | – | – | – | – | a |
| PL196 | L1 | PDA | Cladosporium allicinum | OQ211166 | – | – | – | OQ330826 | a |
| PL197 | L2 | CZA | Streptomyces | – | – | – | – | – | e |
| PL198 | L2 | CZA | Cyphellophora olivacea | OQ211164 | OQ330819 | – | – | – | a |
| PL199 | L2 | CZA | Streptomyces | – | – | – | – | – | e |
| PL200 | L1 | PDA | UnID Fi_2 | – | – | – | – | – | d |
| PL201 | L1 | MEA 10% | Aspergillus protuberus | OQ211127 | – | OQ330767 | OQ330794 | – | a |
| PL202 | L1 | HM 10% | Aspergillus penicillioides | OQ211132 | – | OQ330770 | – | – | a |
| PL203 | L2 | HM 10% | Aspergillus penicillioides | OQ211133 | – | – | – | – | a |
| PL204 | L3 | MEA 10% | UnID Fi_3 | – | – | – | – | – | d |
| PL205 | L4 | DG18 | UnID Fi_1 | – | – | – | – | – | d |
| PL206 | L4 | DG18 | Cladosporium cladosporioides | OQ211168 | – | – | – | OQ330828 | a |
| PL207 | L1 | HM 10% | Aspergillus penicillioides | OQ211130 | – | OQ330768 | – | – | a |
| PL208 | L1 | PDA | Stereum hirsutum | OQ211160 | – | – | – | – | c |
| PL209 | L3 | RB | Cladosporium cladosporioides | – | – | – | – | – | b |
| PL210 | L1 | PDA | Stereum hirsutum | – | – | – | – | – | b |
| PL211 | L2 | RB | Aspergillus protuberus | OQ211146 | – | OQ330777 | OQ330795 | – | a |
| PL212 | L2 | MEA 10% | Stereum hirsutum | OQ211157 | OQ330817 | – | – | – | c |
| PL213 | L2 | MEA 10% | Diaporthe foeniculina | OQ211158 | – | – | – | – | a |
| PL214 | L3 | CZA | UnID Fi_4 | – | – | – | – | – | d |
| PL215 | L1 | HM 10% | Cladosporium sinuosum | OQ211159 | – | – | – | OQ330825 | a |
| PL216 | L3 | RB | UnID Fi_4 | – | – | – | – | d | |
| PL217 | L1 | HM 10% | Cladosporium allicinum | OQ211167 | – | – | – | OQ330827 | a |
| PL218 | L2 | CZA | Streptomyces | – | – | – | – | – | e |
| PL219 | L1 | DG18 | Aspergillus penicillioides | OQ211169 | – | OQ330784 | OQ330799 | – | a |
| PL220 | L1 | PDA | Aspergillus protuberus | OQ211170 | – | OQ330785 | OQ330796 | – | a |
| PL221 | L4 | DG18 | UnID Fi_1 | – | – | – | – | – | d |
| PL222 | L1 | PDA | Aspergillus protuberus | OQ211171 | – | OQ330786 | OQ330797 | – | a |
| PL223 | L1 | DG18 | Aspergillus penicillioides | – | – | – | – | – | b |
| PL224 | L1 | PDA | UnID Fi_2 | – | – | – | – | – | d |
| PL225 | L1 | CZA | Talaromyces acaricola | OQ211172 | – | OQ330787 | OQ330800 | – | a |
| PL226 | L1 | CZA | Talaromyces acaricola | OQ211173 | – | – | – | – | a |
| PL227 | L1 | CZA | Talaromyces acaricola | OQ211174 | – | – | OQ330801 | – | a |
References
- Merino, N.; Aronson, H.S.; Bojanova, D.P.; Feyhl-buska, J.; Wong, M.L.; Zhang, S.; Giovannelli, D. Living at the Extremes: Extremophiles and the Limits of Life in a Planetary Context. Front. Microbiol. 2019, 10, 780. [Google Scholar] [CrossRef] [PubMed]
- Onofri, S.; Selbmann, L.; De Hoog, G.S.; Grube, M.; Barreca, D.; Ruisi, S.; Zucconi, L. Evolution and adaptation of fungi at boundaries of life. Adv. Space Res. 2007, 40, 1657–1664. [Google Scholar] [CrossRef]
- Gadd, G.M. Geomicrobiology of the built environment. Nat. Microbiol. 2017, 2, 16275. [Google Scholar] [CrossRef]
- Dedesko, S.; Siegel, J.A. Moisture parameters and fungal communities associated with gypsum drywall in buildings. Microbiome 2015, 3, 71. [Google Scholar] [CrossRef] [PubMed]
- Palmer, F.E.; Emery, D.R.; Stemmler, J.; Staley, J.T. Survival and growth of microcolonial rock fungi as affected by temperature and humidity. New Phytol. 1987, 107, 155–162. [Google Scholar] [CrossRef]
- Gorbushina, A.A.; Kotlova, E.R.; Sherstneva, O.A. Cellular responses of microcolonial rock fungi to long-term desiccation and subsequent rehydration. Stud. Mycol. 2008, 61, 91–97. [Google Scholar] [CrossRef]
- Onofri, S.; Zucconi, l.; Isola, D.; Selbmann, L. Rock-inhabiting fungi and their role in deterioration of stone monuments in the Mediterranean area. Plant Biosyst. 2014, 148, 384–391. [Google Scholar] [CrossRef]
- Saiz-Jimenez, C. Deposition of anthropogenic compounds on monuments and their effect on airborne microorganisms. Aerobiologia 1995, 11, 161–175. [Google Scholar] [CrossRef]
- Urzì, C.; De Leo, F.; Salamone, P.; Criseo, G. Airborne fungal spores colonising marbles exposed in the terrace of Messina Museum, Sicily. Aerobiologia 2001, 17, 11–17. [Google Scholar] [CrossRef]
- Polo, A.; Gulotta, D.; Santo, N.; Di Benedetto, C.; Fascio, U.; Toniolo, L.; Villa, F.; Cappitelli, F. Importance of subaerial biofilms and airborne microflora in the deterioration of stonework: A molecular study. Biofouling 2012, 28, 1093–1106. [Google Scholar] [CrossRef]
- De Leo, F.; Marchetta, A.; Urzì, C. Black Fungi on Stone-Built Heritage: Current Knowledge and Future Outlook. Appl. Sci. 2022, 12, 3969. [Google Scholar] [CrossRef]
- Scheerer, S.; Ortega-Morales, O.; Gaylarde, C. Chapter 5 Microbial Deterioration of Stone Monuments—An Updated Overview. Adv. Appl. Microbiol. 2009, 66, 97–139. [Google Scholar] [CrossRef] [PubMed]
- Dakal, T.C.; Cameotra, S.S. Microbially induced deterioration of architectural heritages: Routes and mechanisms involved. Environ. Sci. Eur. 2012, 24, 36. [Google Scholar] [CrossRef]
- Gadd, G.M. Geomycology: Biogeochemical transformations of rocks, minerals, metals and radionuclides by Fungi, Bioweathering and Bioremediation. Mycol. Res. 2007, 111, 3–49. [Google Scholar] [CrossRef]
- Gadd, G.M. Fungi, Rocks, and Minerals. Elements 2017, 3, 171–176. [Google Scholar] [CrossRef]
- Warscheid, T.; Braams, J. Biodeterioration of stone: A review. Int. Biodeterior. Biodegrad. 2000, 46, 343–368. [Google Scholar] [CrossRef]
- Sterflinger, K. Fungi as geologic agents. Geomicrobiol. J. 2000, 17, 97–124. [Google Scholar] [CrossRef]
- Sterflinger, K. Fungi: Their role in deterioration of cultural heritage. Fungal Biol. Rev. 2010, 24, 47–55. [Google Scholar] [CrossRef]
- Griffin, P.S.; Indictor, N.; Koestler, R.J. The biodeterioration of stone: A review of deterioration mechanisms, conservation case histories, and treatment. Int. Biodeterior. 1991, 28, 187–207. [Google Scholar] [CrossRef]
- Pinheiro, A.C.; Mesquita, N.; Trovão, J.; Soares, F.; Tiago, I.; Coelho, C.; Carvalho, H.P.; Gil, F.; Catarino, L.; Piñar, G.; et al. Limestone biodeterioration: A review on the portuguese cultural heritage scenario. J. Cult. Herit. 2019, 36, 275–285. [Google Scholar] [CrossRef]
- Sterflinger, K.; Piñar, G. Microbial deterioration of cultural heritage and works of art—Tilting at windmills? Appl. Microbiol. Biotechnol. 2013, 97, 9637–9646. [Google Scholar] [CrossRef] [PubMed]
- Adamiak, J.; Otlewska, A.; Tafer, H.; Lopandic, K.; Gutarowska, B.; Sterflinger, K.; Piñar, G. First evaluation of the microbiome of built cultural heritage by using the Ion Torrent next generation sequencing platform. Int. Biodeterior. Biodegrad. 2018, 131, 11–18. [Google Scholar] [CrossRef]
- Brzozowska, I.; Bogdanowicz, A.; Szczęsny, P.; Zielenkiewicz, U.; Laudy, A. Evaluation of bacterial diversity on historical silk velvet textiles from the Museum of King John III’s Palace at Wilanów, Poland. Int. Biodeterior. Biodegrad. 2018, 131, 78–87. [Google Scholar] [CrossRef]
- Duan, Y.; Wu, F.; Wang, W.; Gu, J.D.; Li, Y.; Feng, H.; Chen, T.; Liu, G.; An, L. Differences of Microbial Community on the wall paintings preserved in situ and ex situ of the Tiantishan Grottoes, China. Int. Biodeterior. Biodegrad. 2018, 132, 102–113. [Google Scholar] [CrossRef]
- Dyda, M.; Decewicz, P.; Romaniuk, K.; Wojcieszak, M.; Sklodowska, A.; Dziewit, L.; Drewniak, L.; Laudy, A. Application of metagenomic methods for selection of an optimal growth medium for bacterial diversity analysis of microbiocenoses on historical stone surfaces. Int. Biodeterior. Biodegrad. 2018, 131, 2–10. [Google Scholar] [CrossRef]
- Gaylarde, C.; Ogawa, A.; Beech, I.; Kowalski, M.; Baptista-Neto, J.A. Analysis of dark crusts on the church of Nossa Senhora do Carmo in Rio de Janeiro, Brazil, using chemical, microscope and metabarcoding microbial identification techniques. Int. Biodeterior. Biodegrad. 2017, 117, 60–67. [Google Scholar] [CrossRef]
- Kraková, L.; Šoltys, K.; Otlewska, A.; Pietrzak, K.; Purkrtová, S.; Savická, D.; Puškárová, A.; Bučková, M.; Szemes, T.; Budiš, J.; et al. Comparison of methods for identification of microbial communities in book collections: Culture-dependent (sequencing and MALDI-TOF MS) and culture-in-dependent (Illumina MiSeq). Int. Biodeterior. Biodegrad. 2018, 131, 51–59. [Google Scholar] [CrossRef]
- Lepinay, C.; Mihajlovski, A.; Touron, S.; Seyer, D.; Bousta, F.; Di Martino, P. Bacterial diversity associated with saline efflorescences damaging the walls of a French decorated prehistoric cave registered as a World Cultural Heritage Site. Int. Biodeterior. Biodegrad. 2018, 130, 55–64. [Google Scholar] [CrossRef]
- Li, M.; Fang, C.; Kawasaki, S.; Huang, M.; Achal, V. Bio-consolidation of cracks in masonry cement mortars by Acinetobacter sp. SC4 isolated from a karst cave. Int. Biodeterior. Biodegrad. 2018, 141, 94–100. [Google Scholar] [CrossRef]
- Lors, C.; Aube, J.; Guyoneaud, R.; Vandenbulcke, F.; Damidot, D. Biodeterioration of mortars exposed to sewers in relation to microbial diversity of biofilms formed on the mortars surface. Int. Biodeterior. Biodegrad. 2018, 130, 23–31. [Google Scholar] [CrossRef]
- Martins, C.; Silva Pereira, C.; Plechkova, N.V.; Seddon, K.R.; Wang, J.; Whitfield, S.; Wong, W. Mycobiota of silk faced ancient Mogao Grottoes manuscripts belonging to the Stein collection in the British library. Int. Biodeterior. Biodegrad. 2018, 134, 1–6. [Google Scholar] [CrossRef]
- Ogawa, A.; Celikkol-Aydin, S.; Gaylarde, C.; Baptista-Neto, J.A.; Beech, I. Microbial communities on painted wet and dry external surfaces of a historic fortress in Niterói, Brazil. Int. Biodeterior. Biodegrad. 2017, 123, 164–173. [Google Scholar] [CrossRef]
- Szulc, J.; Otlewska, A.; Ruman, T.; Kubiak, K.; Karbowska-Berent, J.; Kozielec, T.; Gutarowska, B. Analysis of paper foxing by newly available omics techniques. Int. Biodeterior. Biodegrad. 2018, 132, 157–165. [Google Scholar] [CrossRef]
- Trovão, J.; Portugal, A.; Soares, F.; Paiva, D.S.; Mesquita, N.; Coelho, C.; Pinheiro, A.C.; Catarino, L.; Gil, F.; Tiago, I. Fungal diversity and distribution across distinct biodeterioration phenomena in limestone walls of the old cathedral of Coimbra, UNESCO World Heritage Site. Int. Biodeterior. Biodegrad. 2019, 142, 91–102. [Google Scholar] [CrossRef]
- Zhang, X.; Ge, Q.; Zhu, Z.; Deng, Y.; Gu, J.D. Microbiological community of the royal palace in Angkor Thom and Beng Mealea of Cambodia by Illumina sequencing based on 16S rRNA gene. Int. Biodeterior. Biodegrad. 2018, 134, 127–135. [Google Scholar] [CrossRef]
- Selbmann, L.; Stoppiello, G.A.; Onofri, S.; Stajich, J.E.; Coleine, C. Culture-Dependent and Amplicon Sequencing Approaches Reveal Diversity and Distribution of Black Fungi in Antarctic Cryptoendolithic Communities. J. Fungi 2021, 7, 213. [Google Scholar] [CrossRef]
- González, J.M.; Sáiz-Jiménez, C. Application of molecular nucleic acid-based techniques for the study of microbial communities in monuments and artworks. Int. Microbiol. 2005, 8, 189–194. [Google Scholar] [PubMed]
- Mihajlovski, A.; Seyer, D.; Benamara, H.; Bousta, F.; Di Martino, P. An overview of techniques for the characterization and quantification of microbial colonization on stone monuments. Ann. Microbiol. 2015, 65, 1243–1255. [Google Scholar] [CrossRef]
- Dakal, T.C.; Arora, P.K. Evaluation of potential of molecular and physical techniques in studying biodeterioration. Rev. Environ. Sci. Bio Technol. 2012, 11, 71–104. [Google Scholar] [CrossRef]
- Sanmartín, P.; DeAraujo, A.; Vasanthakumar, A. Melding the Old with the New: Trends in Methods Used to Identify, Monitor, and Control Microorganisms on Cultural Heritage Materials. Microb. Ecol. 2018, 76, 64–80. [Google Scholar] [CrossRef]
- Paiva, D.S.; Fernandes, L.; Trovão, J.; Mesquita, N.; Tiago, I.; Portugal, A. Uncovering the Fungal Diversity Colonizing Limestone Walls of a Forgotten Monument in the Central Region of Portugal by High-Throughput Sequencing and Culture-Based Methods. Appl. Sci. 2022, 12, 10650. [Google Scholar] [CrossRef]
- de Nadal, E.; Ammerer, G.; Posas, F. Controlling gene expression in response to stress. Nat. Rev. Genet. 2011, 12, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Mafart, P.; Couvert, O.; Leguérinel, I. Effect of pH on the heat resistance of spores: Comparison of two models. Int. J. Food Microbiol. 2001, 63, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Deegenaars, M.L.; Watson, K. Heat shock response in psychrophilic and psychrotrophic yeast from Antarctica. Extremophiles 1998, 2, 41–50. [Google Scholar] [CrossRef]
- Tesei, D.; Marzban, G.; Zakharova, K.; Isola, D.; Selbmann, L.; Sterflinger, k. Alteration of protein patterns in black rock inhabiting fungi as a response to different temperatures. Fungal Biol. 2012, 116, 932–940. [Google Scholar] [CrossRef]
- Igreja Paroquial de Trofa/Igreja de São Salvador e Panteão dos Lemos. Available online: http://www.monumentos.gov.pt/Site/APP_PagesUser/SIPA.aspx?id=1042 (accessed on 29 January 2023).
- Miller, A.Z.; Leal, N.; Laiz, L.; Candelera, R.; Silva, M.A.; Dionisio, A.; Macedo, M.F.; Sáiz-Jiménez, C. Primary bioreceptivity of limestones used in southern European monuments. Geol. Soc. Lond. Spec. Publ. 2010, 331, 79–92. [Google Scholar] [CrossRef]
- Vergès-Belmin, V. ICOMOS-ISCS: Illustrated Glossary on Stone Deterioration Patterns Glossaire Illustré sur les Formes d’Altération de la Pierre; ICOMOS-MONUMENTS and SITES XV: Paris, France, 2008; ISBN 978-2-918086-00-0. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Stielow, J.; Lévesque, C.; Seifert, K.; Meyer, W.; Irinyi, L.; Smits, D.; Renfurm, R.; Verkley, G.; Groenewald, M.; Chaduli, D. One fungus, which genes? Development and assessment of universal primers for potential secondary fungal DNA barcodes. Persoonia 2015, 35, 242–263. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Yilmaz, N.; Visagie, C.M.; Frisvad, J.C.; Houbraken, J.; Jacobs, K.; Samson, R.A. Taxonomic re-evaluation of species in Talaromyces section Islandici, using a polyphasic approach. Persoonia 2016, 36, 37–56. [Google Scholar] [CrossRef]
- Hong, S.B.; Cho, H.S.; Shin, H.D.; Frisvad, J.C.; Samson, R.A. Novel Neosartorya species isolated from soil in Korea. Int. J. Syst. Evolut. Microbiol. 2006, 56, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Bensch, K.; Groenewald, J.Z.; Braun, U.; Dijksterhuis, J.; de Jesús Yáñez-Morales, M.; Crous, P.W. Common but different: The expanding realm of Cladosporium. Stud. Mycol. 2015, 82, 23–74. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: An online initiative to launch mycology into the 21st century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- Robert, V.; Stegehuis, G.; Stalpers, J. The MycoBank Engine and Related Databases. 2005. Available online: https://www.MycoBank.org/ (accessed on 3 January 2023).
- Hammer, O.; Harper, D.; Ryan, P. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Eevol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Trovão, J.; Tiago, I.; Soares, F.; Paiva, D.S.; Mesquita, N.; Coelho, C.; Catarino, L.; Gil, F.; Portugal, A. Description of Aeminiaceae fam. nov., Aeminium gen. nov. and Aeminium ludgeri sp. nov. (Capnodiales), isolated from a biodeteriorated art-piece in the Old Cathedral of Coimbra. MycoKeys 2019, 45, 57–73. [Google Scholar] [CrossRef]
- Trovão, J.; Gil, F.; Catarino, L.; Soares, F.; Tiago, I.; Portugal, A. Analysis of fungal deterioration phenomena in the first Portuguese King tomb using a multi-analytical approach. Int. Biodeterior. Biodegrad. 2020, 149, 104933. [Google Scholar] [CrossRef]
- Trovão, J.; Portugal, A. Current Knowledge on the Fungal Degradation Abilities Profiled through Biodeteriorative Plate Essays. Appl. Sci. 2021, 11, 4196. [Google Scholar] [CrossRef]
- Dylag, M.; Spychała, K.; Zielinski, J.; Łagowski, D.; Gnat, S. Update on Stachybotrys chartarum—Black Mold Perceived asToxigenic and Potentially Pathogenic to Humans. Biology 2022, 11, 352. [Google Scholar] [CrossRef] [PubMed]
- Scrano, L.; Boccone, L.F.; Bufo, S.A.; Carrieri, R.; Lahoz, E.; Crescenzi, A. Morphological and molecular characterization of fungal populations possibly involved in the biological alteration of stones in historical buildings. Commun. Agric. Appl. Biol. Sci. 2012, 77, 187–195. [Google Scholar]
- Ponizovskaya, V.B.; Rebrikova, N.L.; Kachalkin, A.V.; Antropova, A.B.; Bilanenko, E.N.; Mokeeva, V.L. Micromycetes as Colonizers of Mineral Building Materials in Historic Monuments and Museums. Fungal Biol. 2019, 123, 290–306. [Google Scholar] [CrossRef]
- Isola, D.; Zucconi, L.; Cecchini, A.; Caneva, G. Dark-pigmented biodeteriogenic fungi in Etruscan hypogeal tombs: New data on their culture-dependent diversity, favoring conditions, and resistance to biocidal treatments. Fungal Biol. 2021, 125, 609–620. [Google Scholar] [CrossRef]
- Trovão, J.; Tiago, I.; Catarino, L.; Gil, F.; Portugal, A. In vitro analyses of fungi and dolomitic limestone interactions: Bioreceptivity and biodeterioration assessment. Int. Biodeterior. Biodegrad. 2020, 155, 105107. [Google Scholar] [CrossRef]
- Savković, Ž.; Stupar, M.; Unković, N.; Ivanović, Ž.; Blagojević, J.; Vukojević, J.; Ljaljević Grbić, M. In vitro biodegradation potential of airborne Aspergilli and Penicillia. Sci. Nat. 2019, 106, 8. [Google Scholar] [CrossRef]
- Unković, N.; Erić, S.; Šarić, K.; Stupar, M.; Savković, Ž.; Stanković, S.; Stanojević, O.; Dimkić, I.; Vukojević, J.; Ljaljević Grbić, M. Biogenesis of secondary mycogenic minerals related to wall paintings deterioration process. Micron 2017, 100, 1–9. [Google Scholar] [CrossRef]
- Savković, Ž.; Unković, N.; Stupar, M.; Franković, M.; Jovanović, M.; Erić, S.; Šarić, K.; Stanković, S.; Dimkić, I.; Vukojević, J.; et al. Diversity and biodeteriorative potential of fungal dwellers on ancient stone stela. Int. Biodeterior. Biodegrad. 2016, 115, 212–223. [Google Scholar] [CrossRef]
- Visagie, C.M.; Hirooka, Y.; Tanney, J.B.; Whitfield, E.; Mwange, K.; Meijer, M.; Amend, A.S.; Seifert, K.A.; Samson, R.A. Aspergillus, Penicillium and Talaromyces isolated from house dust samples collected around the world. Stud. Mycol. 2014, 78, 63–139. [Google Scholar] [CrossRef] [PubMed]
- Bensch, K.; Groenewald, J.Z.; Meijer, M.; Dijksterhuis, J.; Jurjević, Ž.; Andersen, B.; Houbraken, J.; Crous, P.W.; Samson, R.A. Cladosporium species in indoor environments. Stud. Mycol. 2018, 89, 177–301. [Google Scholar] [CrossRef] [PubMed]
- Horner, W.E.; Worthan, A.G.; Morey, P.R. Air- and dustborne mycoflora in houses free of water damage and fungal growth. Appl. Environ. Microbiol. 2004, 70, 6394–6400. [Google Scholar] [CrossRef] [PubMed]
- Géry, A.; Rioult, J.-P.; Heutte, N.; Séguin, V.; Bonhomme, J.; Garon, D. First Characterization and Description of Aspergillus Series Versicolores in French Bioaerosols. J. Fungi 2021, 7, 676. [Google Scholar] [CrossRef]
- Nazareth, S.; Gonsalves, V. Aspergillus penicillioides—A true halophile existing in hypersaline and polyhaline econiches. Ann. Microbiol. 2014, 64, 397–402. [Google Scholar] [CrossRef]
- Zammit, G.; Sanchez-Moral, S.; Albertano, P. Bacterially mediated mineralization processes lead to biodeterioration of artworks in Maltese catacombs. Sci. Total Environ. 2011, 409, 2773–2782. [Google Scholar] [CrossRef]
- Li, T.; Hu, Y.; Zhang, B.; Yang, X. Role of fungi in the formation of patinas on Feilaifeng limestone, China. Microb. Ecol. 2018, 76, 352–361. [Google Scholar] [CrossRef]
- Del Monte, M.; Sabbioni, C.; Zappia, G. The Origin of Calcium Oxalates on Historical Buildings, Monuments and Natural Outcrops. Sci. Total Environ. 1987, 67, 17–39. [Google Scholar] [CrossRef]
- Zhang, Y.; Su, M.; Wu, F.; Gu, J.-D.; Li, J.; He, D.; Guo, Q.; Cui, H.; Zhang, Q.; Feng, H. Diversity and Composition of Culturable Microorganisms and Their Biodeterioration Potentials in the Sandstone of Beishiku Temple, China. Microorganisms 2023, 11, 429. [Google Scholar] [CrossRef]
- Aguilera-Cogley, V.A.; Berbegal, M.; Català, S.; Brentu, F.C.; Armengol, J.; Vicent, A. Characterization of Mycosphaerellaceae species associated with citrus greasy spot in Panama and Spain. PLoS ONE 2017, 12, e0189585. [Google Scholar] [CrossRef]
- Arciuolo, R.; Santos, C.; Soares, C.; Castello, G.; Spigolon, N.; Chiusa, G.; Lima, N.; Battilani, P. Molecular Characterization of Diaporthe Species Associated with Hazelnut defects. Front. Plant Sci. 2020, 11, 611655. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, J.; He, Y.; Chang, M.; Menga, J. Characterization of the complete mitochondrial genome of Coprinellus micaceus, a wild saprobic mushroom in China. Mitochondrial DNA Part B 2021, 6, 1979–1981. [Google Scholar] [CrossRef] [PubMed]
- Tanney, J.B.; Seifert, K.A. Mollisiaceae: An overlooked lineage of diverse endophytes. Stud. Mycol. 2020, 95, 293–380. [Google Scholar] [CrossRef] [PubMed]
- Connell, L.; Redman, R.; Craig, S.; Scorzetti, G.; Iszard, M.; Rodriguez, R. Diversity of Soil Yeasts Isolated from South Victoria Land, Antarctica. Microb. Ecol. 2008, 56, 448–459. [Google Scholar] [CrossRef]
- Arenz, B.E.; Blanchette, R.A. Distribution and abundance of soil fungi in Antarctica at sites on the Peninsula, Ross Sea Region and McMurdo dry valleys. Soil Biol. Biochem. 2011, 43, 308–315. [Google Scholar] [CrossRef]
- Robinson, C.H. Cold adaptation in Arctic and Antarctic fungi. New Phytol. 2001, 151, 341–353. [Google Scholar] [CrossRef]
- Wang, M.; Jianqing, T.; Xiang, M.; Liu, X. Living strategy of cold-adapted fungi with the reference to several representative species. Mycology 2017, 8, 178–188. [Google Scholar] [CrossRef]
- Tesei, D. Black Fungi Research: Out-of-This-World Implications. Encyclopedia 2022, 2, 212–229. [Google Scholar] [CrossRef]
- Blanchette, R.A.; Held, B.W.; Jurgens, J.A.; McNew, D.L.; Harrington, T.C.; Duncan, S.M.; Farrell, R.L. Wood destroying soft-rot fungi in the historic expedition huts of Antarctica. Appl. Environ. Microbiol. 2004, 70, 1328–1335. [Google Scholar] [CrossRef]
- Ludley, K.E.; Robinson, C.H. Decomposer, Basidiomycota in Arctic and Antarctic ecosystems. Soil Biol. Biochem. 2008, 40, 11–29. [Google Scholar] [CrossRef]
- Hossain, M.A.; Ahmed, M.S.; Ghannoum, M.A. Attributes of Stachybotrys chartarum and Its Association with Human Disease. J. Allergy Clin. Immunol. 2004, 113, 200–208; Quiz 209. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Choudhry, H.; Asseri, A.H.; Elfaky, M.A.; Mohamed, S.G.A.; Mohamed, G.A. Stachybotrys chartarum—A Hidden Treasure: Secondary Metabolites, Bioactivities, and Biotechnological Relevance. J. Fungi 2022, 8, 504. [Google Scholar] [CrossRef] [PubMed]
- Ruisi, S.; Barreca, D.; Selbmann, L.; Zucconi, L.; Onofri, S. Fungi in Antarctica. Rev. Environ. Sci. Biotechnol. 2007, 6, 127–141. [Google Scholar] [CrossRef]
- Gostinčar, G.; Zalar, P.; Gunde-Cimerman, N. No need for speed: Slow development of fungi in extreme environments. Fungal Biol. Rev. 2022, 39, 1–14. [Google Scholar] [CrossRef]
- Sterflinger, K. Temperature and NaCl- tolerance of rock-inhabiting meristematic fungi. Antonie Van Leeuwenhoek 1998, 74, 271–281. [Google Scholar] [CrossRef]







| Locus | Primers | Sequence (5′-3′) | Annealing (°C) | Amplification/Cycles | Length (bp) | Group | Reference |
|---|---|---|---|---|---|---|---|
| Internal transcribed spacer (ITS) | ITS1F ITS4 | ctt ggt cat tta gag gaa gta a tcc tcc gct tat tga tat gc | 52 | Standard 35 | ~600 | All Fungi | [49] |
| Translation elongation factor 1α (Tef1) | EF1-1018F EF1-1620R | gay ttc atc aag aac atg at gac gtt gaa dcc rac rtt gtc | 56 | Touchdown 36 | ~600 | All Fungi * | [50] |
| β-tubulin (BenA) | Bt2a Bt2b | ggt aac caa atc ggt gct gct ttc acc ctc agt gta gtg acc ctt ggc | 55 | Standard 35 | ~500 | Aspergillus, Penicillium and Talaromyces | [51,52] |
| Calmodulin (CaM) | CMD5 CMD6 | ccg agt aca agg agg cct tc ccg ata gag gtc ata acg tgg | 55 | Standard 35 | ~580 | Aspergillus and Talaromyces | [52,53] |
| Partial Actin (Act) | Act-512F Act-783R | atg tgc aag gcc ggt ttc gc tac gag tcc ttc tgg ccc at | 55 | Standard 35 | ~370 | Cladosporium | [54,55] |
| Sample ID | Species Richness (S) | Shannon (H) | Evenness (e^H/S) | Dominance (D) |
|---|---|---|---|---|
| L1 | 11 | 1.77 | 0.53 | 0.22 |
| L2 | 9 | 1.39 | 0.44 | 0.29 |
| L3 | 10 | 1.27 | 0.35 | 0.37 |
| L4 | 3 | 0.53 | 0.56 | 0.73 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paiva, D.S.; Fernandes, L.; Pereira, E.; Trovão, J.; Mesquita, N.; Tiago, I.; Portugal, A. Exploring Differences in Culturable Fungal Diversity Using Standard Freezing Incubation—A Case Study in the Limestones of Lemos Pantheon (Portugal). J. Fungi 2023, 9, 501. https://doi.org/10.3390/jof9040501
Paiva DS, Fernandes L, Pereira E, Trovão J, Mesquita N, Tiago I, Portugal A. Exploring Differences in Culturable Fungal Diversity Using Standard Freezing Incubation—A Case Study in the Limestones of Lemos Pantheon (Portugal). Journal of Fungi. 2023; 9(4):501. https://doi.org/10.3390/jof9040501
Chicago/Turabian StylePaiva, Diana S., Luís Fernandes, Emília Pereira, João Trovão, Nuno Mesquita, Igor Tiago, and António Portugal. 2023. "Exploring Differences in Culturable Fungal Diversity Using Standard Freezing Incubation—A Case Study in the Limestones of Lemos Pantheon (Portugal)" Journal of Fungi 9, no. 4: 501. https://doi.org/10.3390/jof9040501