
AMF Inoculation Alleviates Molybdenum Toxicity to Maize by Protecting Leaf Performance


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Materials
2.2. Sample Analysis
2.3. Statistical Analysis
3. Results
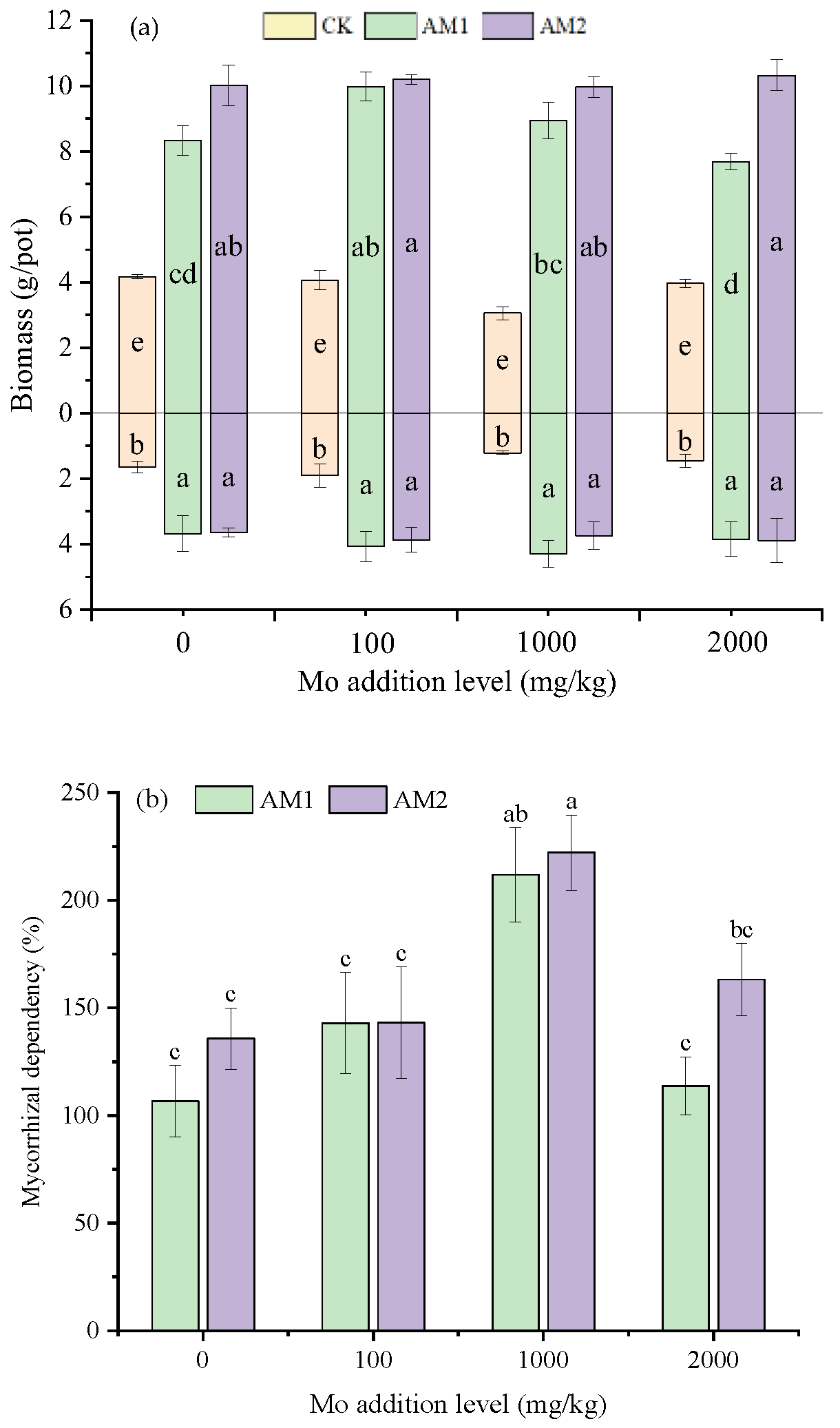
3.1. Effects of AMF Inoculation on Plant Biomass under Mo Stress
3.2. Mo Concentration in Maize under Different Levels of Mo Addition
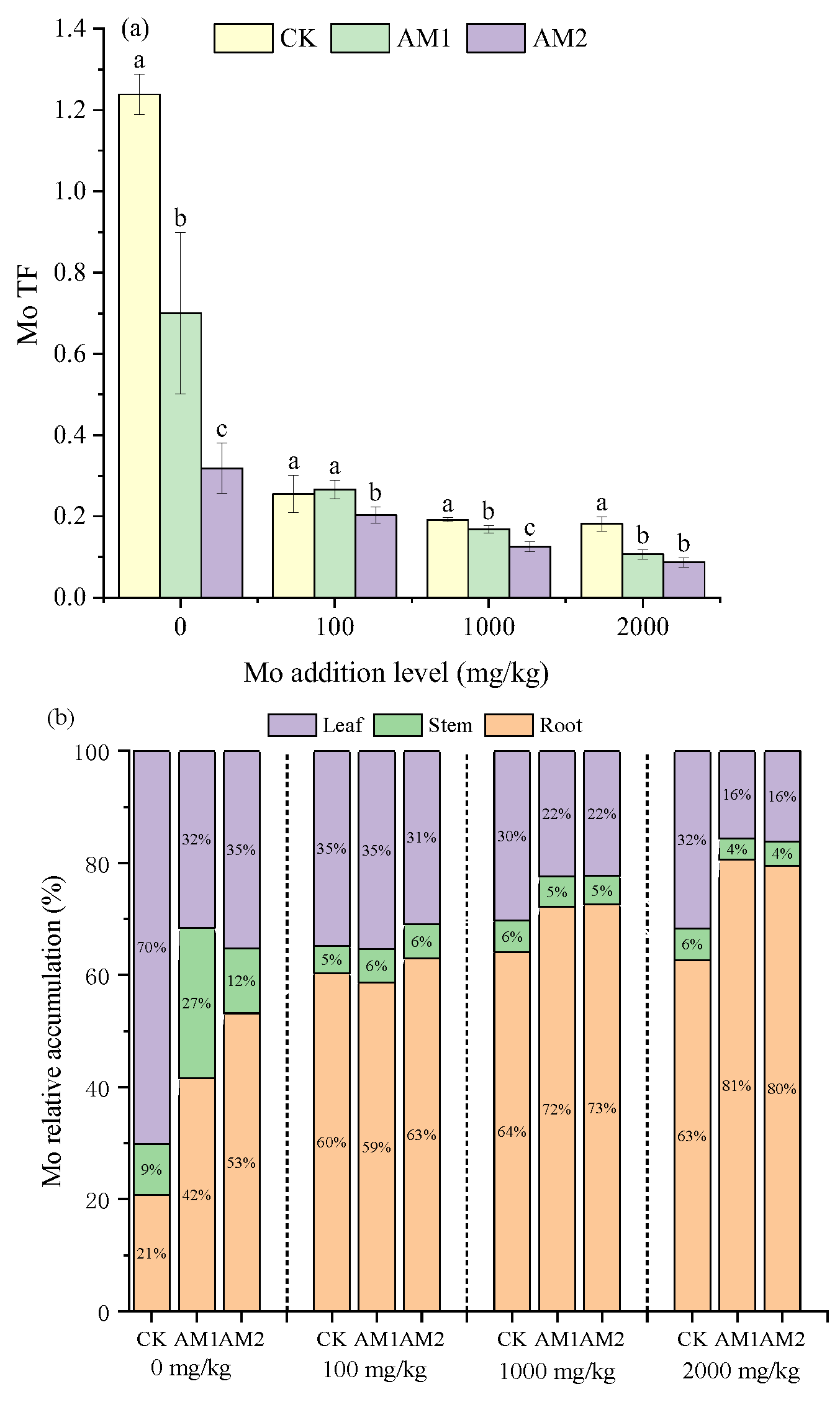
3.3. Effects of AMF on Mo Distribution in Different Organs
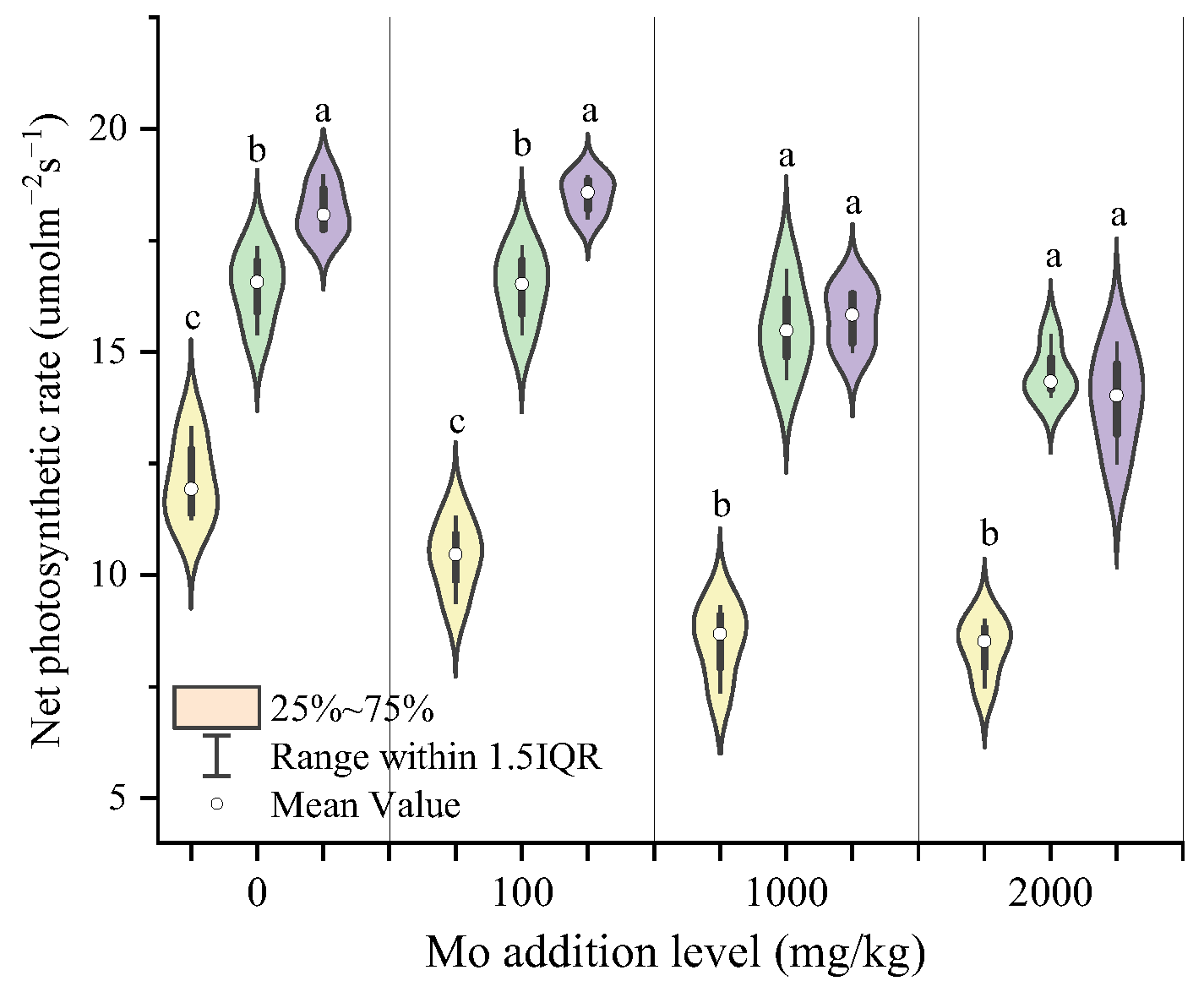
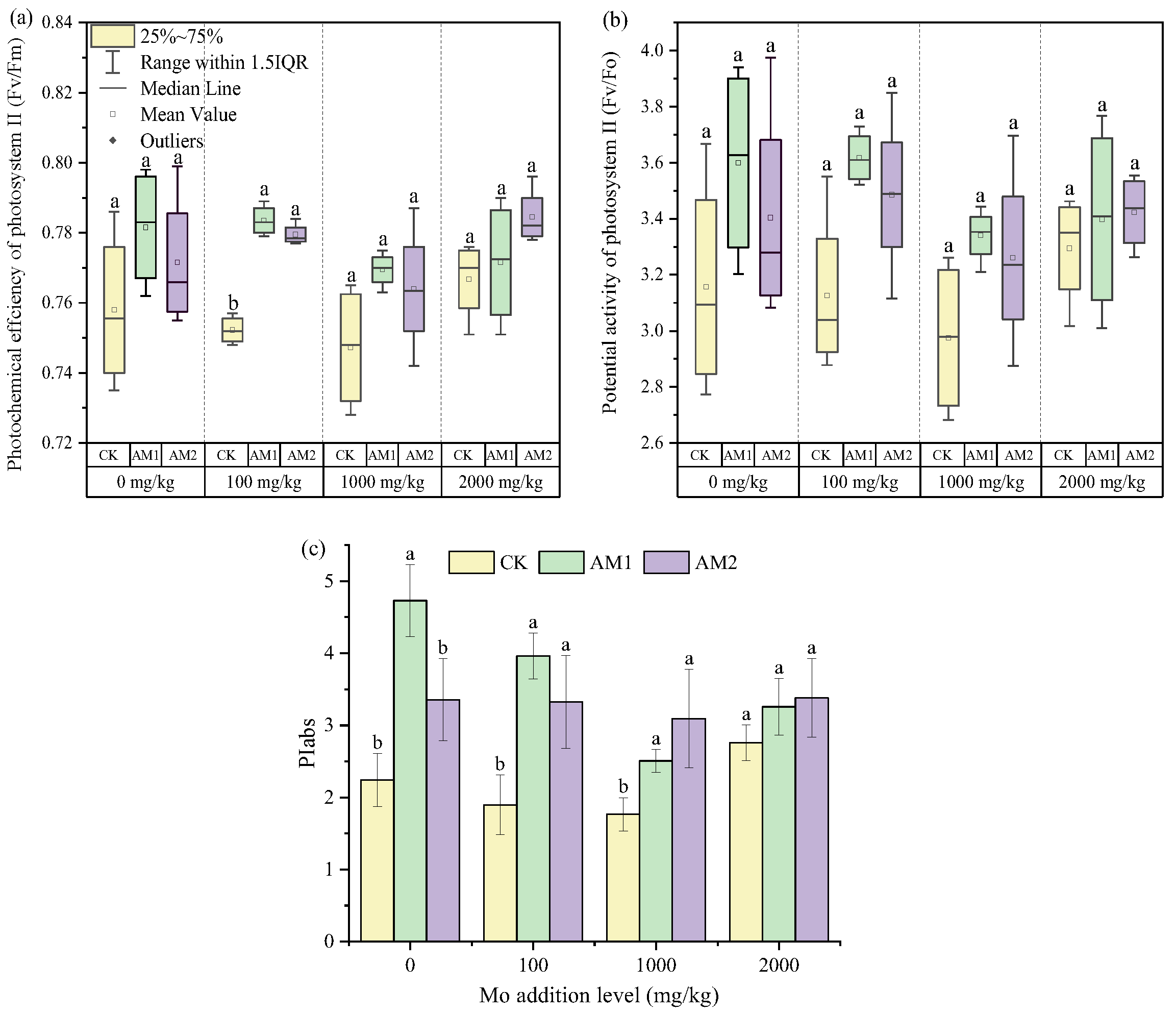
3.4. Effects of AMF on Photosynthetic Parameters of Maize under Different Degrees of Mo Stress
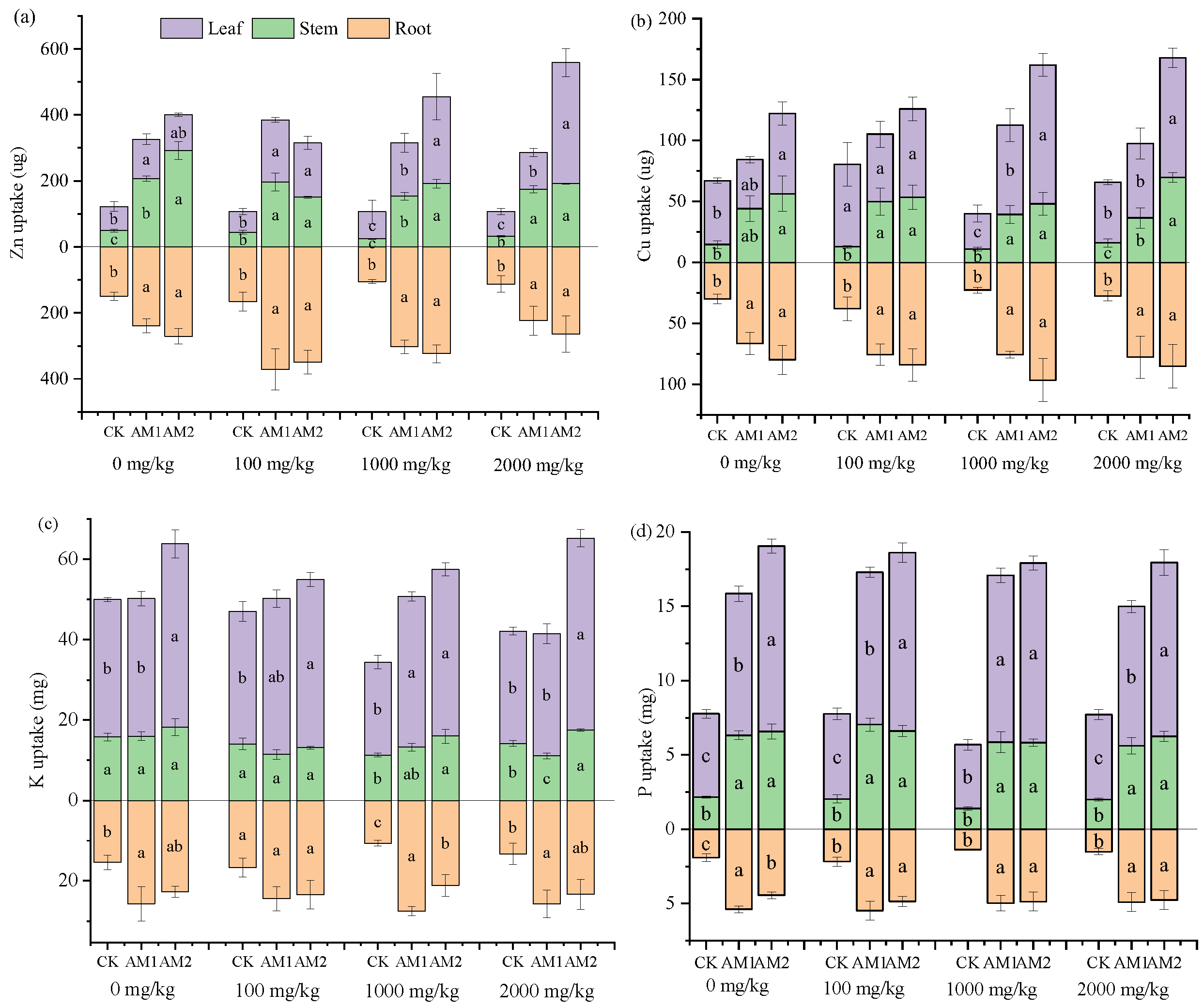
3.5. Uptake of Nutrient in Plants Organs
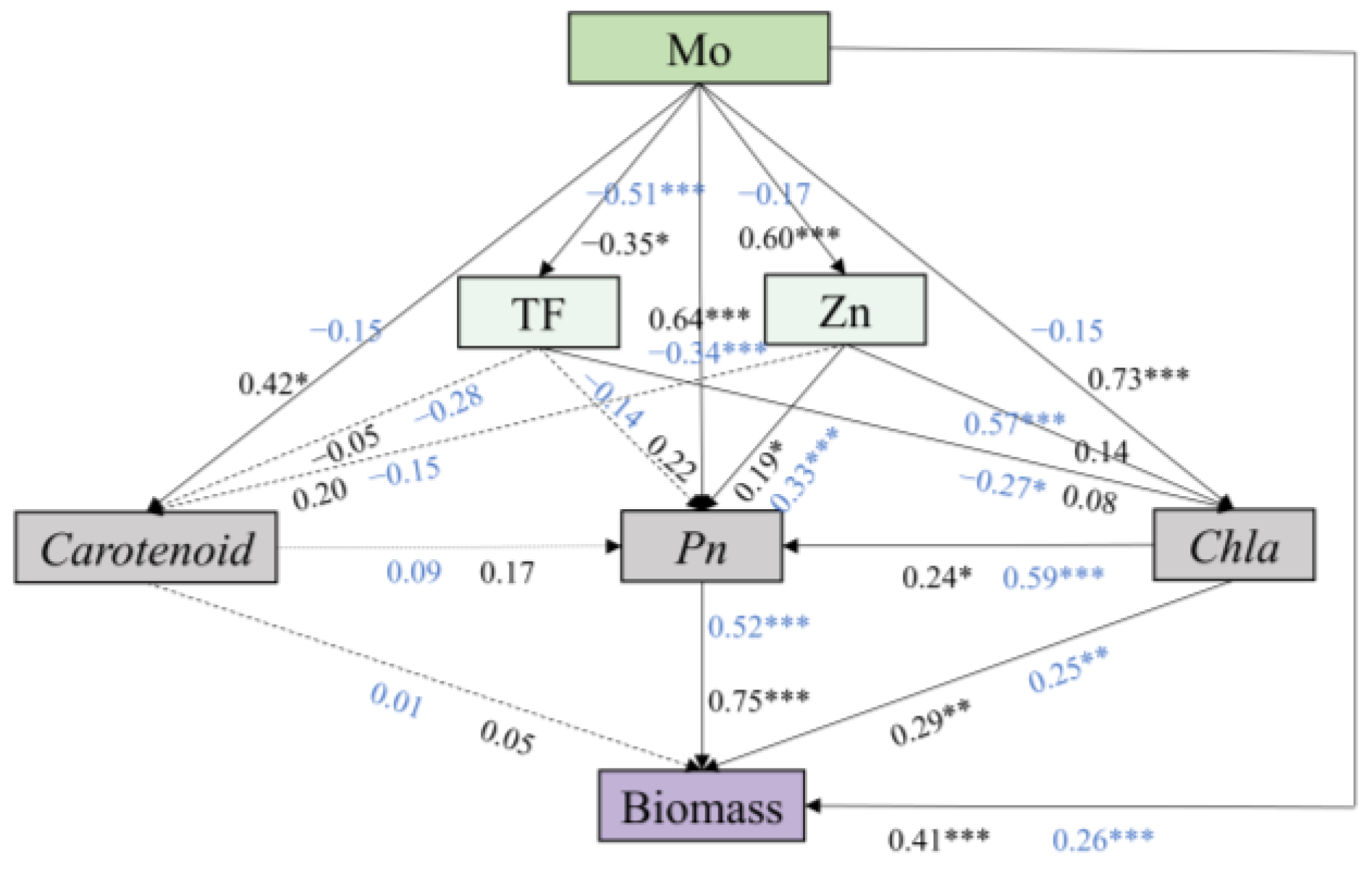
3.6. The Effect of Physiological Characteristics on Maize Biomass under AMF and Mo Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaiser, B.N.; Gridley, K.L.; Brady, J.N.; Phillips, T.; Tyerman, S.D. The role of molybdenum in agricultural plant production. Ann. Bot. 2005, 96, 745–754. [Google Scholar] [CrossRef] [PubMed]
- van Gestel, C.A.M.; McGrath, S.P.; Smolders, E.; Ortiz, M.D.; Borgman, E.; Verweij, R.A.; Buekers, J.; Oorts, K. Effect of long-term equilibration on the toxicity of molybdenum to soil organisms. Environ. Pollut. 2012, 162, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mcgrath, S.P.; Mico, C.; Curdy, R.; Zhao, F.J. Predicting molybdenum toxicity to higher plants: Influence of soil properties. Environ. Pollut. 2010, 158, 3095–3102. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, W.S.; Lewis, A.H.; Watson, S.J. The teart pastures of Somerset: I. The cause and cure of teartness. J. Agric. Sci. 1943, 33, 44–51. [Google Scholar] [CrossRef]
- Henckens, M.L.C.M.; Driessen, P.P.J.; Worrell, E. Molybdenum resources: Their depletion and safeguarding for future generations. Resour. Conserv. Recycl. 2018, 134, 61–69. [Google Scholar] [CrossRef]
- Xu, S.J.; Hu, C.X.; Tan, Q.L.; Qin, S.Y.; Sun, X.C. Subcellular distribution of molybdenum, ultrastructural and antioxidative responses in soybean seedlings under excess molybdenum stress. Plant Physiol. Bioch. 2018, 123, 75–80. [Google Scholar] [CrossRef]
- Kluge, R.; Anke, M. Molybdenum toxicity in plants. In Proceedings Mengen-und Spurenelemente Arbeitstagung, Leipzig; Institut für Pflanzenernährung: Jena, Germany, 1983; pp. 10–17. [Google Scholar]
- Zeng, Q.D.; Liu, J.M.; Qin, K.Z.; Fan, H.R.; Chu, S.X.; Wang, Y.B.; Zhou, L.L. Types, characteristics, and time-space distribution of molybdenum deposits in China. Int. Geol. Rev. 2013, 55, 1311–1358. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.; Li, J.K.; Zhang, H.X.; Xia, Y.; Chen, C.; Shen, Z.G.; Chen, Y.H. Several newly discovered Mo-enriched plants with a focus on Macleaya cordata. Environ. Sci. Pollut. Res. 2018, 25, 26493–26503. [Google Scholar] [CrossRef]
- Zhang, Q.C.; Wang, C.C. Natural and human factors affect the distribution of soil heavy metal pollution: A review. Water Air Soil Pollut. 2020, 231, 7. [Google Scholar] [CrossRef]
- Han, Z.X.; Wan, D.J.; Tian, H.X.; He, W.X.; Wang, Z.Q.; Liu, Q. Pollution assessment of heavy metals in soils and plants around a molybdenum mine in central China. Pol. J. Environ. Stud. 2019, 28, 123–133. [Google Scholar] [CrossRef]
- Wang, X.Q.; Brunetti, G.; Tian, W.J.; Owens, G.; Qu, Y.; Jin, C.X.; Lombi, E. Effect of soil amendments on molybdenum availability in mine affected agricultural soils. Environ. Pollut. 2020, 269, 116132. [Google Scholar] [CrossRef] [PubMed]
- Jia, T.; Jia, Y.Y.; Yu, S.J.; Qu, Y.M.; Jiang, Z.L.; Chen, Y.H.; Wang, G. Pollution of molybdenum and heavy metals of the soils and rice near a molybdenum mining site in eastern Fujian. Environ. Monito. China 2015, 31, 45–49. [Google Scholar]
- Matnur, S.; Jajoo, A. Arbuscular mycorrhizal fungi protect maize plants from high temperature stress by regulating photosystem II heterogeneity. Ind. Crops Prod. 2020, 143, 111934. [Google Scholar] [CrossRef]
- Di, L.; Zheng, K.Y.; Wang, Y.; Zhang, Y.; Lao, R.M.; Qin, Z.Y.; Li, T.; Zhao, Z.W. Harnessing an arbuscular mycorrhizal fungus to improve the adaptability of a facultative metallophytic poplar (Populus yunnanensis) to cadmium stress: Physiological and molecular responses. J. Hazard. Mater. 2021, 42, 127430. [Google Scholar] [CrossRef]
- Liu, R.C.; Ding, Y.E.; Wu, Q.S.; Zou, Y.N. Mycorrhizae enhance drought tolerance of trifoliate orange by regulating circadian clock response patterns. Sci. Hortic. 2022, 305, 111426. [Google Scholar] [CrossRef]
- Kumar, A.; Dames, J.F.; Gupta, A.; Sharma, S.; Gilbert, J.A.; Ahmad, P. Current developments in arbuscular mycorrhizal fungi research and its role in salinity stress alleviation: A biotechnological perspective. Crit. Rev. Biotechnol. 2015, 35, 461–474. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D. Mycorrhizal Symbiosis, 3rd ed.; Elsevier: New York, NY, USA, 2008; pp. 1–800. [Google Scholar]
- Tiwari, J.; Ma, Y.; Bauddh, K. Arbuscular mycorrhizal fungi: An ecological accelerator of phytoremediation of metal contaminated soils. Arch. Agron. Soil Sci. 2020, 68, 283–296. [Google Scholar] [CrossRef]
- Wipf, D.; Krajinski, F.; van Tuinen, D.; Recorbet, G.; Courty, P.E. Trading on the arbuscular mycorrhiza market: From arbuscules to common mycorrhizal networks. New Phytol. 2019, 223, 1127–1142. [Google Scholar] [CrossRef]
- Sun, S.; Feng, Y.H.; Huang, G.D.; Zhao, X.; Song, F.Q. Rhizophagus irregularis enhances tolerance to cadmium stress by altering host plant hemp (Cannabis sativa L.) photosynthetic properties. Environ. Pollut. 2022, 314, 120309. [Google Scholar] [CrossRef]
- Wang, H.R.; Zhao, X.Y.; Zhang, J.M.; Lu, C.; Feng, F.J. Arbuscular mycorrhizal fungus regulates cadmium accumulation, migration, transport, and tolerance in Medicago sativa. J. Hazard. Mater. 2022, 435, 129077. [Google Scholar] [CrossRef]
- Wang, F. Occurrence of arbuscular mycorrhizal fungi in mining-impacted sites and their contribution to ecological restoration: Mechanisms and applications. Crit. Rev. Environ. Sci. Technol. 2017, 47, 1901–1957. [Google Scholar] [CrossRef]
- Li, H.; Chen, X.W.; Wu, L.; Luo, N.; Huang, W.X.; Mo, C.H.; Li, Y.W.; Xiang, L.; Zhao, H.M.; Cai, Q.Y. Effects of arbuscular mycorrhizal fungi on redox homeostasis of rice under Cd stress. Plant Soil 2020, 1, 121–138. [Google Scholar] [CrossRef]
- Hu, Z.H.; Zhuo, F.; Jing, S.H.; Li, X.; Yan, T.X.; Lei, L.L.; Lu, R.R.; Zhang, X.F.; Jing, Y.X. Combined application of arbuscular mycorrhizal fungi and steel slag improves plant growth and reduces Cd, Pb accumulation in Zea may. Int. J. Phytoremediat. 2019, 21, 857–865. [Google Scholar] [CrossRef] [PubMed]
- He, Y.M.; Fan, X.M.; Zhang, G.Q.; Li, B.; Li, T.G.; Zu, Y.Q.; Zhan, F.D. Effects of arbuscular mycorrhizal fungi and dark septate endophytes on maize performance and root traits under a high cadmium stress. S. Afr. J. Bot. 2021, 134, 415–423. [Google Scholar] [CrossRef]
- Tan, Q.Y.; Guo, Q.J.; Wei, R.F.; Zhu, G.X.; Du, C.J.; Hu, H.Y. Influence of arbuscular mycorrhizal fungi on bioaccumulation and bioavailability of As and Cd: A meta-analysis. Environ. Pollut. 2023, 36, 120619. [Google Scholar] [CrossRef]
- Wang, F.Y.; Cheng, P.; Zhang, S.Q.; Zhang, S.W.; Sun, Y.H. Contribution of arbuscular mycorrhizal fungi and soil amendments to remediation of a heavy metal-contaminated soil using sweet sorghum. Pedosphere 2022, 32, 844–855. [Google Scholar] [CrossRef]
- Shi, Z.Y.; Zhang, J.C.; Wang, F.Y.; Li, K.; Yuan, W.K.; Liu, J.B. Arbuscular mycorrhizal inoculation increases molybdenum accumulation but decreases molybdenum toxicity in maize plants grown in polluted soil. J. Fungi 2018, 8, 37069–37076. [Google Scholar] [CrossRef]
- Yin, K.J.; Shi, Z.Y.; Zhang, M.G.; Li, Y.J. Effects of mining on the molybdenum absorption and translocation of plants in the Luanchuan molybdenum mine. PeerJ 2021, 8, e9183. [Google Scholar] [CrossRef]
- Dawood, M.; Cao, F.; Jahangir, M.M.; Zhang, G.; Wu, F. Alleviation of aluminum toxicity by hydrogen sulfide is related to elevated ATPase, and suppressed aluminum uptake and oxidative stress in barley. J. Hazard. Mater. 2012, 209, 121–128. [Google Scholar] [CrossRef]
- Chaves, E.S.; dos Santos, E.J.; Araujo, R.G.O.; Oliveira, J.V.; Frescura, V.L.A.; Curtius, A.J. Metals and phosphorus determination in vegetable seeds used in the production of biodiesel by ICP OES and ICP-MS. Microchem. J. 2010, 96, 71–76. [Google Scholar] [CrossRef]
- Wang, G.; Wang, L.; Ma, F.; You, Y.; Wang, Y.; Yang, D. Integration of earthworms and arbuscular mycorrhizal fungi into phytoremediation of cadmium contaminated soil by Solanum nigrum L. J. Hazard. Mater. 2020, 389, 121873. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Jing, X.; Adams, C.A.; Shi, Z.; Sun, Y. Decreased ZnO nanoparticle phytotoxicity to maize by arbuscular mycorrhizal fungus and organic phosphorus. Environ. Sci. Pollut. Res. 2018, 25, 23736–23747. [Google Scholar] [CrossRef]
- You, Y.Q.; Ju, C.; Wang, L.; Wang, X.; Ma, F.; Wang, G.; Wang, Y.J. The mechanism of arbuscular mycorrhizal enhancing cadmium uptake in Phragmites australis depends on the phosphorus concentration. J. Hazard. Mater. 2022, 440, 129800. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.Y.; Adams, C.A.; Shi, Z.Y.; Sun, Y.H. Combined effects of ZnO NPs and Cd on sweet sorghum as influenced by an arbuscular mycorrhizal fungus. Chemosphere 2018, 209, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhu, S.X.; Yang, X.Q.; Xia, G.D.; Wang, B.C.; Gu, B.J. Arbuscular mycorrhizal fungi alter rhizosphere bacterial community characteristics to improve Cr tolerance of Acorus calamus. Ecotoxicol. Environ. Saf. 2023, 253, 114652. [Google Scholar] [CrossRef]
- Kuang, Y.X.; Li, X.; Wang, Z.H.; Wang, X.Y.; Wei, H.J.; Chen, H.; Hu, W.T.; Tang, M. Effects of arbuscular mycorrhizal fungi on the growth and root cell ultrastructure of Eucalyptus grandis under cadmium stress. J. Fungi 2023, 9, 140. [Google Scholar] [CrossRef]
- Li, H.; Gao, M.Y.; Mo, C.H.; Wong, M.H.; Chen, X.W.; Wang, J.J. Potential use of arbuscular mycorrhizal fungi for simultaneous mitigation of arsenic and cadmium accumulation in rice. J. Exp. Bot. 2022, 73, 50–67. [Google Scholar] [CrossRef] [PubMed]
- Khalid, M.; Ur-Rahman, S.; Hassani, D.; Hayat, K.; Zhou, P.; Hui, N. Advances in fungal-assisted phytoremediation of heavy metals: A review. Pedosphere 2021, 31, 475–495. [Google Scholar] [CrossRef]
- Kovacs, B.; Puskas-Preszner, A.; Huzsvai, L.; Levai, L.; Bodi, E. Effect of molybdenum treatment on molybdenum concentration and nitrate reduction in maize seedlings. Plant Physiol. Bioch. 2015, 96, 38–44. [Google Scholar] [CrossRef]
- Guo, W.; Zhao, R.X.; Zhao, W.J.; Fu, R.Y.; Guo, J.Y.; Bi, N.; Zhang, J. Effects of arbuscular mycorrhizal fungi on maize (Zea mays L.) and sorghum (Sorghum bicolor L. Moench) grown in rare earth elements of mine tailings. Appl. Soil Ecol. 2013, 72, 85–92. [Google Scholar] [CrossRef]
- Adriano, D.C. Trace Elements in the Terrestrial Environment; Springer: New York, NY, USA, 1986; p. 329. [Google Scholar]
- Ferguson, W.S.; Lewis, A.H.; Watson, S.J. Action of molybdenum in nutrition of milking cattle. Nature 1938, 141, 553. [Google Scholar] [CrossRef]
- Liu, H.; Yuan, M.; Tan, S.Y.; Yang, X.P.; Lan, Z.; Jiang, Q.Y.; Ye, Z.H.; Jing, Y.X. Enhancement of arbuscular mycorrhizal fungus (Glomus versiforme) on the growth and Cd uptake by Cd-hyperaccumulator Solanum nigrum. Appl. Soil Ecol. 2015, 89, 44–49. [Google Scholar] [CrossRef]
- Zhang, X.F.; Hu, Z.H.; Yan, T.X.; Lu, R.R.; Peng, C.L.; Li, S.S.; Jing, Y.X. Arbuscular mycorrhizal fungi alleviate Cd phytotoxicity by altering Cd subcellular distribution and chemical forms in Zea mays. Ecotoxicol. Environ. Saf. 2019, 171, 352–360. [Google Scholar] [CrossRef]
- Janeeshma, E.; Puthur, J.T. Direct and indirect influence of arbuscular mycorrhizae on enhancing metal tolerance of plants. Arch. Microbiol. 2020, 202, 1–16. [Google Scholar] [CrossRef]
- Zhan, F.; Li, B.; Jiang, M.; Yue, X.; He, Y.; Xia, Y.; Wang, Y. Arbuscular mycorrhizal fungi enhance antioxidant defense in the leaves and the retention of heavy metals in the roots of maize. Environ. Sci. Pollut. Res. 2018, 25, 24338–24347. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, X.; Huang, L.; Chen, B. Arbuscular mycorrhiza and plant chromium tolerance. Soil Ecol. Lett. 2019, 1, 94–104. [Google Scholar] [CrossRef]
- You, Y.Q.; Wang, L.; Ju, C.; Wang, G.; Ma, F.; Wang, Y.J.; Yang, D.G. Effects of arbuscular mycorrhizal fungi on the growth and toxic element uptake of Phragmites australis (Cav.) Trin. ex Steud under zinc/cadmium stress. Ecotoxicol. Environ. Saf. 2021, 213, 112023. [Google Scholar]
- Yu, Z.H.; Zhao, X.L.; Liang, X.R.; Li, Z.R.; Wang, L.; He, Y.M.; Zhan, F.D. Arbuscular mycorrhizal fungi reduce cadmium leaching from sand columns by reducing availability and enhancing uptake by maize roots. J. Fungi 2022, 8, 866. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.Z.; McGee, R.; Hoque, M.A.; Ahammed, G.J.; Carpenter-Boggs, L. Effect of arbuscular mycorrhizal Fungi, Selenium and biochar on photosynthetic pigments and antioxidant enzyme activity under arsenic stress in Mung Bean (Vigna radiata). Front. Physiol. 2019, 10, 193. [Google Scholar] [CrossRef] [PubMed]
- Shabani, L.; Sabzalian, M.R.; Mostafavi pour, S. Arbuscular mycorrhiza affects nickel translocation and expression of ABC transporter and metallothionein genes in Festuca arundinacea. Mycorrhiza 2016, 26, 67–76. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, X.; Sun, Y.; Wu, Z.; Li, T.; Hu, Y.; Lv, J.; Li, G.; Zhang, Z.; Zhang, J.; et al. Chromium immobilization by extra- and intraradical fungal structures of arbuscular mycorrhizal symbioses. J. Hazard. Mater. 2016, 316, 34–42. [Google Scholar]
- Liu, L.; Li, J.; Yue, F.; Yan, X.; Wang, F.; Bloszies, S.; Wang, Y. Effects of arbuscular mycorrhizal inoculation and biochar amendment on maize growth, cadmium uptake and soil cadmium speciation in Cd-contaminated soil. Chemosphere 2018, 194, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Debeljak, M.; van Elteren, J.T.; Spruk, A.; Izmer, A.; Vanhaecke, F.; Vogel-Mikus, K. The role of arbuscular mycorrhiza in mercury and mineral nutrient uptake in maize. Chemosphere 2018, 212, 1076–1084. [Google Scholar] [CrossRef] [PubMed]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L.X. Role of Arbuscular mycorrhizal fungi in plant growth regulation: Implications in abiotic stress tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef]
- Yuan, J.; Shi, K.; Zhou, X.Y.; Wang, L.; Xu, C.; Zhang, H.; Zhu, G.P.; Si, C.C.; Wang, J.D.; Zhang, Y.C. Interactive impact of potassium and arbuscular mycorrhizal fungi on the root morphology and nutrient uptake of sweet potato (Ipomoea batatas L.). Front. Microbiol. 2023, 13, 1075957. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Smith, F.A.; Jakobsen, I. Functional diversity in arbuscular mycorrhizal (AM) symbioses: The contribution of the mycorrhizal P uptake pathway is not correlated with mycorrhizal responses in growth or total P uptake. New Phytol. 2004, 162, 511–524. [Google Scholar] [CrossRef]
- Shi, J.C.; Zhao, B.Y.; Zheng, S.; Zhang, X.W.; Wang, X.L.; Dong, W.T.; Xie, Q.J.; Wang, G.; Xiao, Y.P.; Chen, F.; et al. Phosphate starvation response-centered network regulates mycorrhizal symbiosis. Cell 2021, 184, 5527. [Google Scholar] [CrossRef]
- Hao, L.J.; Zhang, Z.C.; Hao, B.H.; Diao, F.W.; Zhang, J.X.; Bao, Z.H.; Guo, W. Arbuscular mycorrhizal fungi alter microbiome structure of rhizosphere soil to enhance maize tolerance to La. Ecotoxicol. Environ. Saf. 2021, 212, 111996. [Google Scholar] [CrossRef] [PubMed]
- Tondey, M.; Kalia, A.; Singh, A.; Abd-Elsalam, K.; Hassan, M.M.; Dheri, G.S. A comparative evaluation of the effects of seed invigoration treatments with precursor zinc salt and nano-sized zinc oxide (ZnO) particles on vegetative growth, grain yield, and quality characteristics of Zea mays. J. Anal. Sci. Technol. 2022, 13, 40. [Google Scholar] [CrossRef]
- Shreya, D.; Amaresan, N.; Supriya, N.R. Zinc solubilizing Bacillus sp (SS9) and Enterobacter sp (SS7) promote mung bean (Vigna radiata L.) growth, nutrient uptake and physiological profiles. Lett. Appl. Microbiol. 2023, 76, 2. [Google Scholar] [CrossRef]
- Lycka, M.; Bartak, M.; Helia, O.; Kopriva, S.; Moravcova, D.; Hajek, J.; Fojt, L.; Cmelik, R.; Fajkus, J.; Fojtova, M. Sulfate supplementation affects nutrient and photosynthetic status of Arabidopsis thaliana and Nicotiana tabacum differently under prolonged exposure to cadmium. J. Hazard. Mater. 2023, 445, 130527. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Z.; Song, L.L.; Zhang, H.B.; Wang, J.C.; Wang, Y.; Zhang, H.H. Cu and Zn Stress affect the photosynthetic and antioxidative systems of alfalfa (Medicago sativa). J. Plant Interact. 2022, 17, 695–704. [Google Scholar] [CrossRef]
- Cheng, X.N.; Chen, C.; Hu, Y.M.; Guo, X.L.; Wang, J.L. Photosynthesis and growth of Amaranthus tricolor under strontium stress. Chemosphere 2022, 308, 136234. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Yue, X.; Chen, W.; Jiang, H.; Han, Y.; Li, X. Detection of cadmium risk to the photosynthetic performance of hybrid Pennisetum. Front. Plant Sci. 2019, 10, 798. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.H.; Zhong, H.X.; Wang, J.F.; Sui, X.; Xu, N. Adaptive changes in chlorophyll content and photosynthetic features to low light in Physocarpus amurensis maxim and Physocarpus opulifolius “diabolo”. PeerJ 2016, 4, e2125. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.H.; Li, X.; Xu, Z.S.; Wang, Y.; Teng, Z.Y.; An, M.J.; Zhang, Y.H.; Zhu, W.X.; Xu, N.; Sun, G.Y. Toxic effects of heavy metals Pb and Cd on mulberry (Morus alba L.) seedling leaves: Photosynthetic function and reactive oxygen species (ROS) metabolism responses. Ecotoxicol. Environ. Saf. 2020, 195, 110469. [Google Scholar] [CrossRef]
- Santana, N.A.; Avelar Ferreira, P.A.; Tarouco, C.P.; Schardong, I.S.; Antoniolli, Z.I.; Nicoloso, F.T.; Seminoti, J.R.J. Earthworms and mycorrhization increase copper phytoextraction by Canavalia ensiformis in sandy soil. Ecotoxicol. Environ. Saf. 2019, 182, 109383. [Google Scholar] [CrossRef]
- Chaturvedi, R.; Favas, P.; Pratas, J.; Varun, M.; Paul, M.S. Assessment of edibility and effect of arbuscular mycorrhizal fungi on Solanum melongena L. grown under heavy metal (loid) contaminated soil. Ecotoxicol. Environ. Saf. 2018, 148, 318–326. [Google Scholar] [CrossRef]
- Wu, J.T.; Wang, L.; Zhao, L.; Huang, X.C.; Ma, F. Arbuscular mycorrhizal fungi effect growth and photosynthesis of Phragmites australis (Cav.) Trin ex. Steudel under copper stress. Plant Biol. 2019, 22, 62–69. [Google Scholar] [CrossRef]








{kind=link}
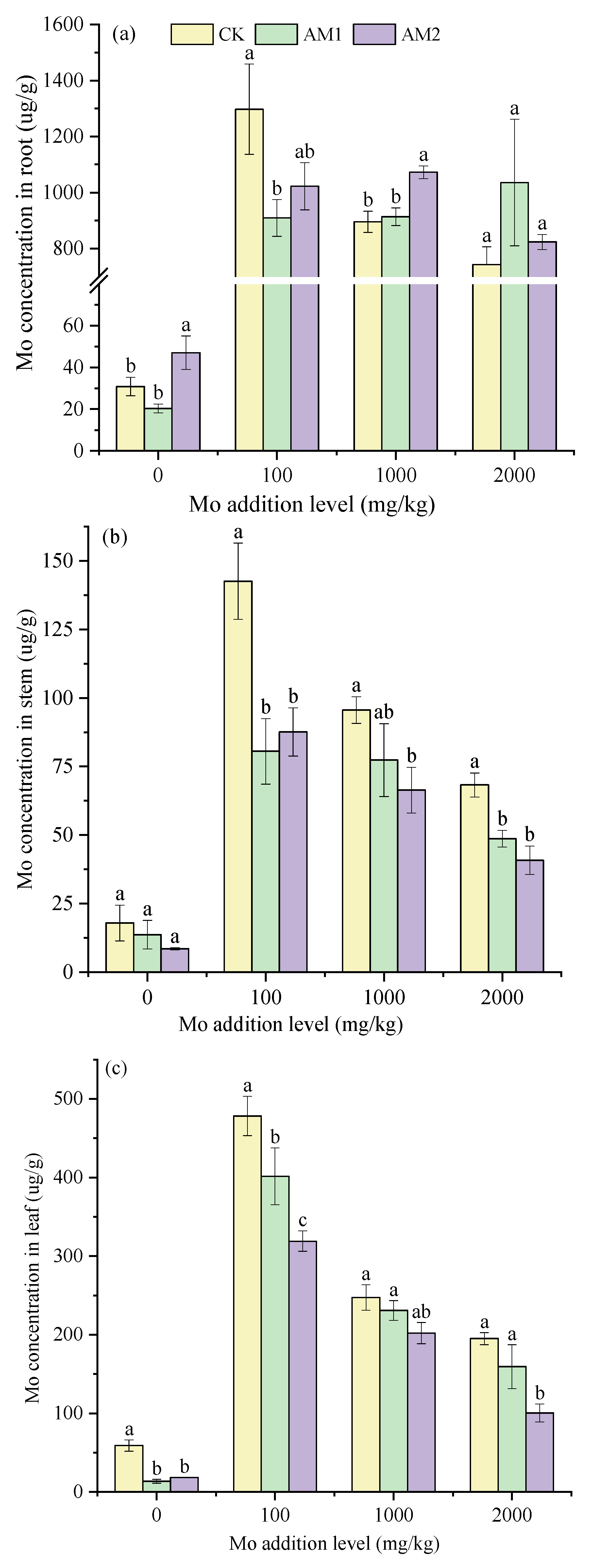{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Photosynthetic Pigment | 0 mg/kg | 100 mg/kg | 1000 mg/kg | 2000 mg/kg | Mo | AMF | Mo × AMF | |
|---|---|---|---|---|---|---|---|---|
| Chl a | CK | 1.88 ± 0.08 b | 1.67 ± 0.1 b | 1.67 ± 0.16 b | 1.63 ± 0.05 c | |||
| AM1 | 2.23 ± 0.1 a | 2.64 ± 0.12 a | 2.30 ± 0.11 a | 2.14 ± 0.03 b | 0.82 ns | 64.27 ** | 2.56 * | |
| AM2 | 2.50 ± 0.08 a | 2.45 ± 0.16 a | 2.48 ± 0.12 a | 2.61 ± 0.1 a | ||||
| Chl b | CK | 0.23 ± 0.01 a | 0.32 ± 0.01 a | 0.28 ± 0.02 a | 0.28 ± 0.02 a | |||
| AM1 | 0.20 ± 0.02 a | 0.18 ± 0.01 b | 0.23 ± 0.03 a | 0.24 ± 0.01 b | 3.17 * | 19.65 ** | 3.32 * | |
| AM2 | 0.21 ± 0.01 a | 0.20 ± 0.02 b | 0.25 ± 0.01 a | 0.20 ± 0.02 b | ||||
| Carotenoid | CK | 0.41 ± 0.03 b | 0.40 ± 0.06 a | 0.35 ± 0.04 b | 0.34 ± 0.01 c | |||
| AM1 | 0.46 ± 0.04 b | 0.60 ± 0.06 a | 0.53 ± 0.05 a | 0.43 ± 0 b | 1.38 ns | 23.78 ** | 1.33 ns | |
| AM2 | 0.59 ± 0.02 a | 0.56 ± 0.03 a | 0.57 ± 0.04 a | 0.58 ± 0.04 a |
| AMF | Mo | Mo × AMF | |
|---|---|---|---|
| Root Mo conc. | 0.08 ns | 84.61 *** | 2.98 ** |
| Stem Mo conc. | 15.97 *** | 66.76 *** | 2.57 * |
| Leaf Mo conc. | 23.19 *** | 230.12 *** | 2.74 * |
| TF of Mo | 19.44 *** | 61.86 *** | 11.13 *** |
| Root Zn uptake | 29.66 *** | 4.36 * | 0.86 ns |
| Stem Zn uptake | 196.23 *** | 12.99 *** | 6.19 *** |
| Leaf Zn uptake | 9.71 *** | 1.72 ns | 2.13 ns |
| Root P uptake | 71.30 *** | 0.66 ns | 0.33 ns |
| Stem P uptake | 173.39 *** | 3.10 * | 0.63 ns |
| Leaf P uptake | 199.22 *** | 0.35 ns | 2.55 * |
| Root Cu uptake | 27.65 *** | 0.23 ns | 0.34 ns |
| Stem Cu uptake | 29.03 *** | 0.52 ns | 0.67 ns |
| Leaf Cu uptake | 16.25 *** | 2.22 ns | 3.67 ** |
| Root K uptake | 19.45 *** | 0.23 ns | 0.52 ns |
| Stem K uptake | 8.12 ** | 6.23 ** | 2.07 ns |
| Leaf K uptake | 53.55 *** | 2.80 ns | 4.59 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Shi, Z.; Lu, S.; Wang, F. AMF Inoculation Alleviates Molybdenum Toxicity to Maize by Protecting Leaf Performance. J. Fungi 2023, 9, 479. https://doi.org/10.3390/jof9040479
Zhang M, Shi Z, Lu S, Wang F. AMF Inoculation Alleviates Molybdenum Toxicity to Maize by Protecting Leaf Performance. Journal of Fungi. 2023; 9(4):479. https://doi.org/10.3390/jof9040479
Chicago/Turabian StyleZhang, Mengge, Zhaoyong Shi, Shichuan Lu, and Fayuan Wang. 2023. "AMF Inoculation Alleviates Molybdenum Toxicity to Maize by Protecting Leaf Performance" Journal of Fungi 9, no. 4: 479. https://doi.org/10.3390/jof9040479