
Biodiversity of Basidiomycetous Yeasts Associated with Cladonia rei Lichen in Japan, with a Description of Microsporomyces cladoniophilus sp. nov
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Isolation, Culturing, and Characterization of the Yeast Strains
2.3. Sequencing and Phylogenetic Analyses
2.4. Meta-Barcoding Sequencing and OTU Identity Analysis
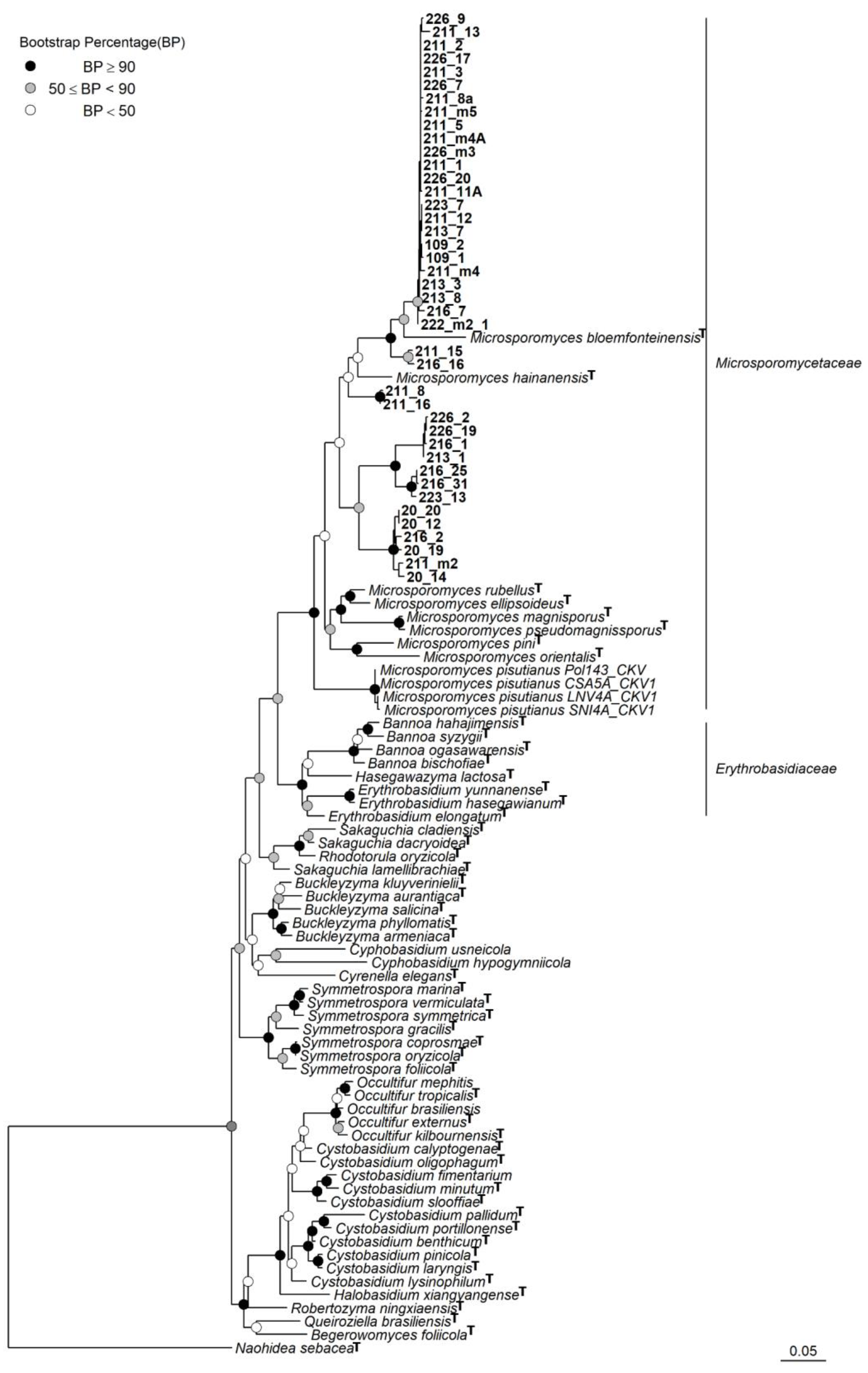
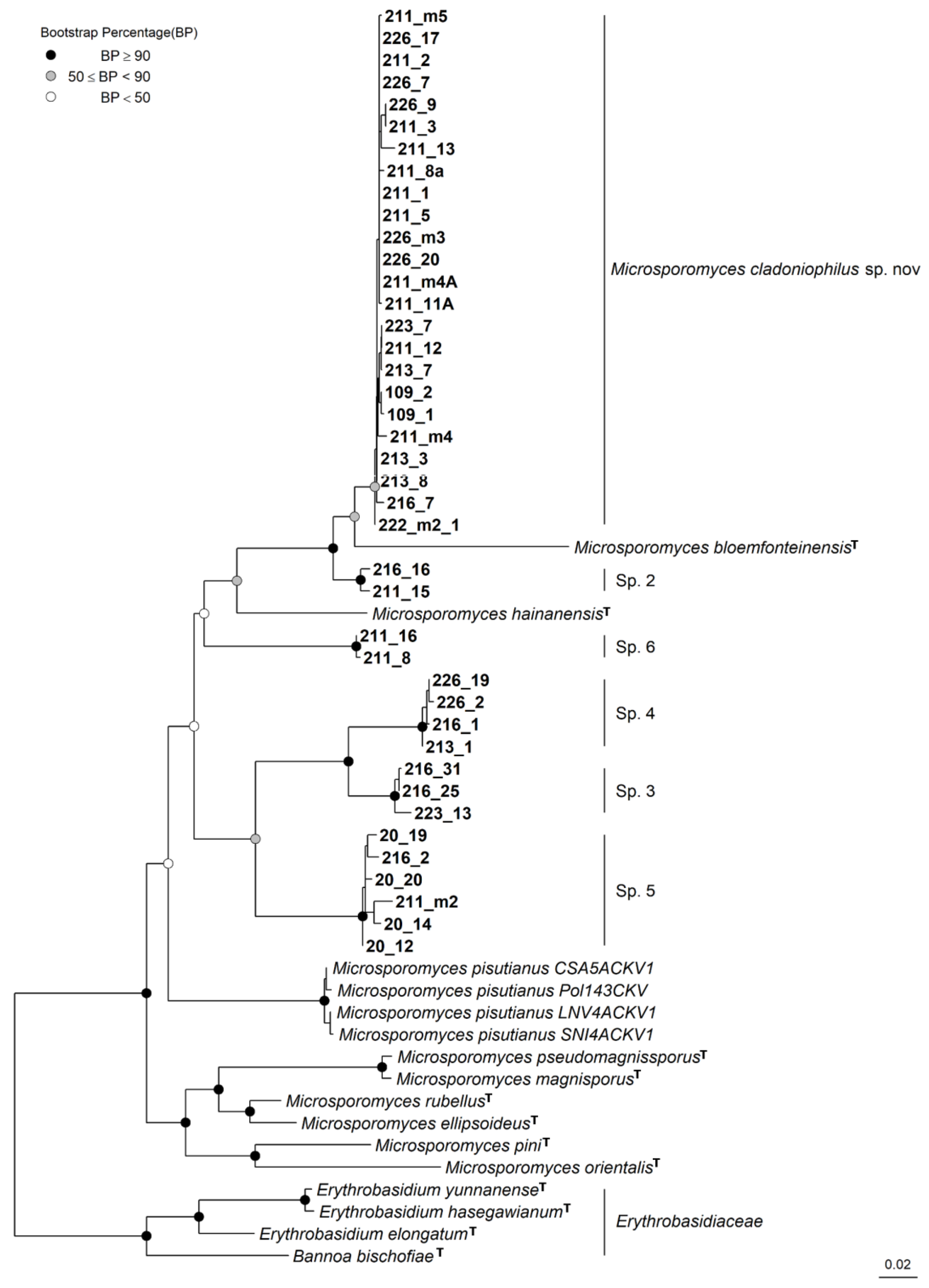
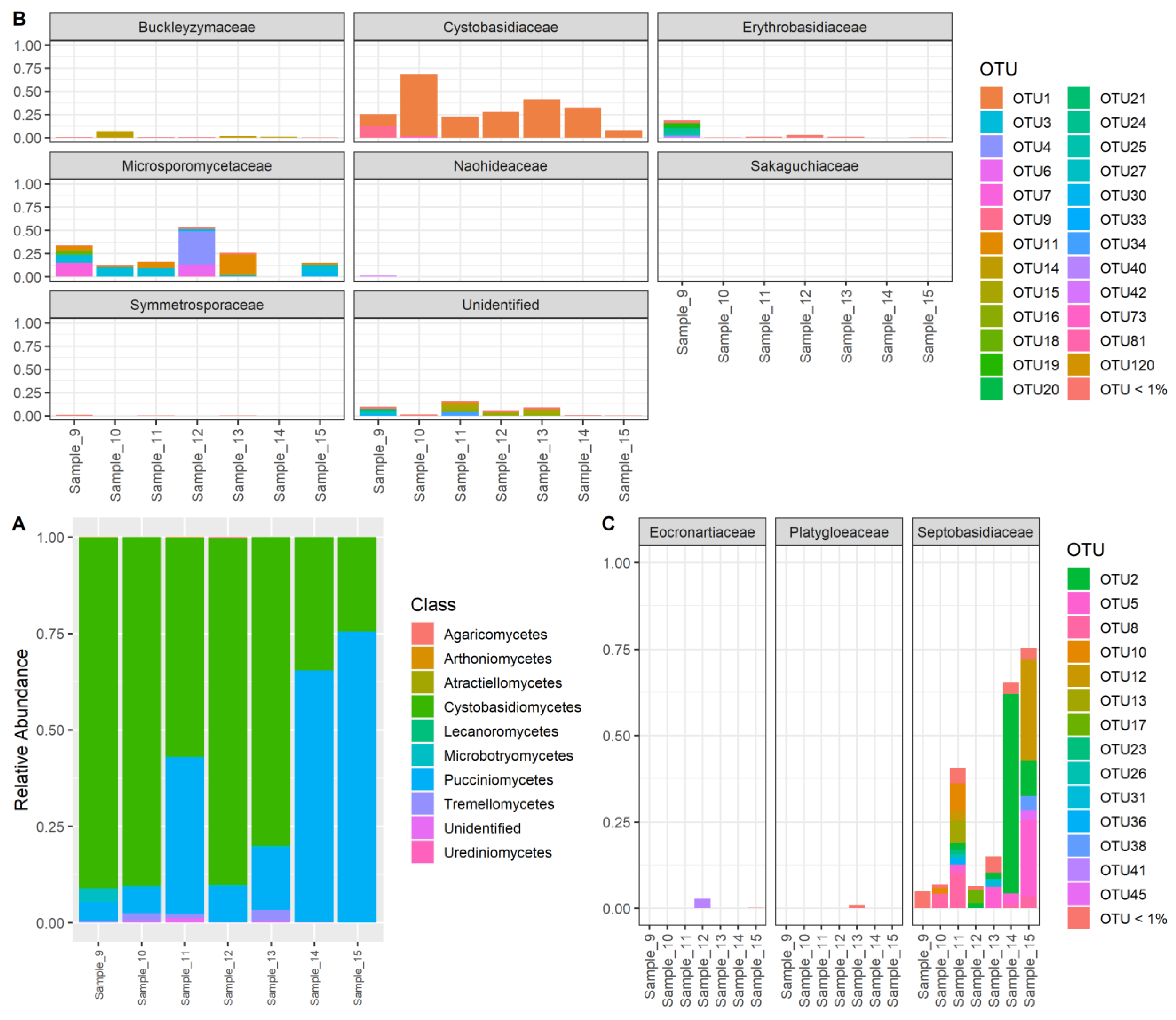
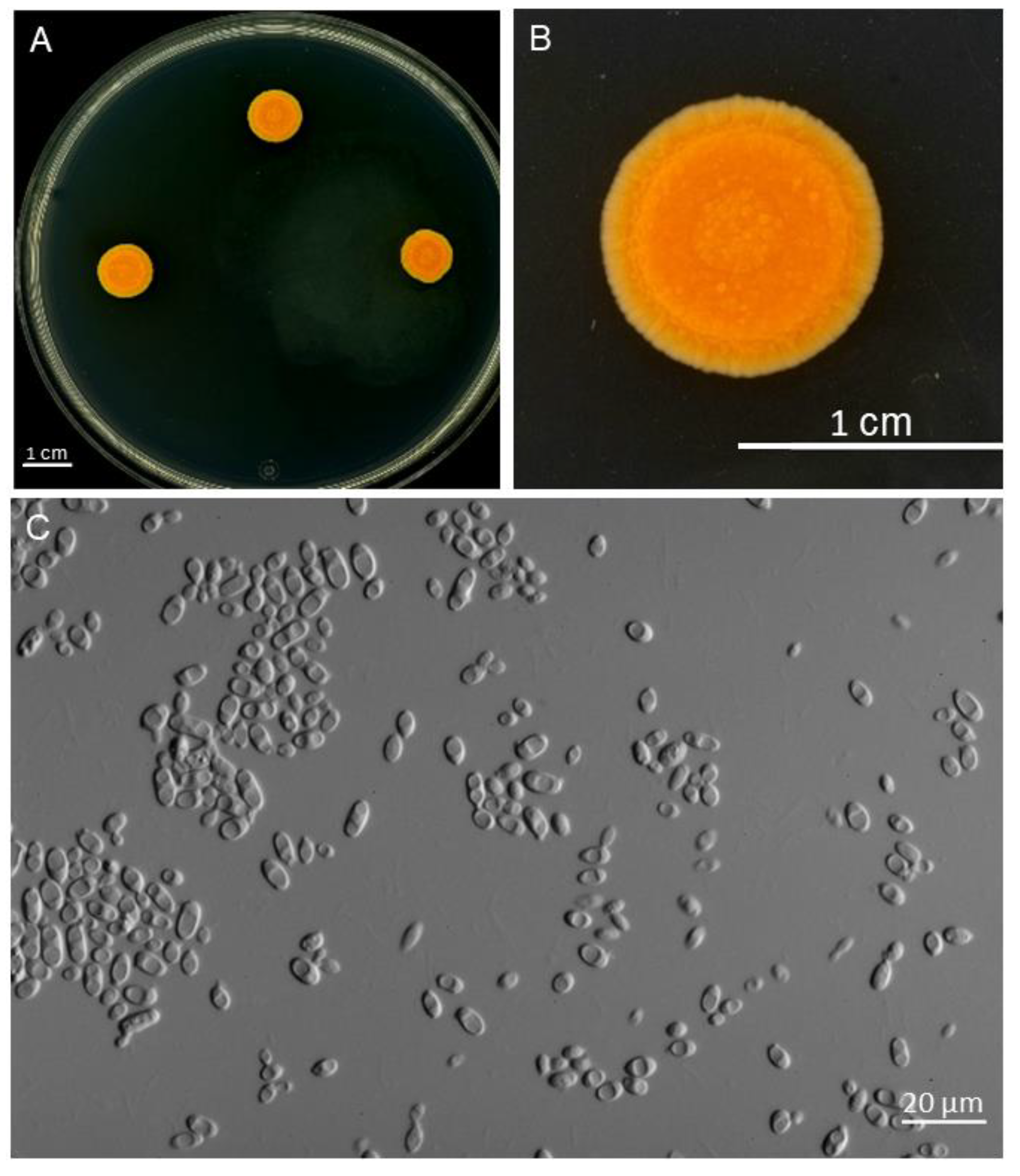
3. Results
3.1. Isolated Cultures
3.2. Meta-Barcoding Results



3.3. Taxonomy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asplund, J.; Wardle, D.A. How lichens impact on terrestrial community and ecosystem properties. Biol. Rev. 2017, 92, 1720–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nash, T.H. Lichen Biology; Cambridge University Press: Cambridge, UK, 1996; ISBN 0521459745. [Google Scholar]
- Nelsen, M.P.; Lücking, R.; Boyce, C.K.; Lumbsch, H.T.; Ree, R.H. The macroevolutionary dynamics of symbiotic and phenotypic diversification in lichens. Proc. Natl. Acad. Sci. USA 2020, 117, 202001913. [Google Scholar] [CrossRef] [PubMed]
- Stenroos, S.; Hyvönen, J.; Myllys, L.; Thell, A.; Ahti, T. Phylogeny of the genus Cladonia s.lat. (Cladoniaceae, Ascomycetes) inferred from molecular, morphological, and chemical data. Cladistics 2002, 18, 237–278. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Grube, M. Lichens redefined as complex ecosystems. New Phytol. 2020, 227, 1281–1283. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Xiao, S.; Taylor, T.N. Lichen-like symbiosis 600 million years ago. Science 2005, 308, 1017–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honegger, R.; Axe, L.; Edwards, D. Bacterial epibionts and endolichenic actinobacteria and fungi in the Lower Devonian lichen Chlorolichenomycites salopensis. Fungal Biol. 2013, 117, 512–518. [Google Scholar] [CrossRef] [PubMed]
- U’Ren, J.M.; Lutzoni, F.; Miadlikowska, J.; Laetsch, A.D.; Arnold, A.E. Host and geographic structure of endophytic and endolichenic fungi at a continental scale. Am. J. Bot. 2012, 99, 898–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muggia, L.; Fleischhacker, A.; Kopun, T.; Grube, M. Extremotolerant fungi from alpine rock lichens and their phylogenetic relationships. Fungal Divers. 2016, 76, 119–142. [Google Scholar] [CrossRef] [Green Version]
- Bates, S.T.; Berg-Lyons, D.; Lauber, C.L.; Walters, W.A.; Knight, R.; Fierer, N. A preliminary survey of lichen associated eukaryotes using pyrosequencing. Lichenol 2012, 44, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Zhurbenko, M.P.; Pino-Bodas, R. Lichenicolous fungi growing on Cladonia, mainly from the Northern Hemisphere, with a worldwide key to the known species. Opusc. Philolichenum 2017, 16, 188–266. [Google Scholar] [CrossRef]
- Suryanarayanan, T.S.; Thirunavukkarasu, N. Endolichenic fungi: The lesser known fungal associates of lichens. Mycology 2017, 8, 189–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spribille, T.; Tuovinen, V.; Resl, P.; Vanderpool, D.; Wolinski, H.; Aime, M.C.; Schneider, K.; Stabentheiner, E.; Toome-Heller, M.; Thor, G.; et al. Basidiomycete yeasts in the cortex of ascomycete macrolichens. Science 2016, 353, 488–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Černajová, I.; Škaloud, P. The first survey of Cystobasidiomycete yeasts in the lichen genus Cladonia; with the description of Lichenozyma pisutiana gen. nov., sp. nov. Fungal Biol. 2019, 123, 625–637. [Google Scholar] [CrossRef] [PubMed]
- Tuovinen, V.; Ekman, S.; Thor, G.; Vanderpool, D.; Spribille, T.; Johannesson, H. Two basidiomycete fungi in the cortex of wolf lichens. Curr. Biol. 2019, 29, 476–483.e5. [Google Scholar] [CrossRef] [Green Version]
- Mark, K.; Laanisto, L.; Bueno, C.G.; Niinemets, Ü.; Keller, C.; Scheidegger, C. Contrasting co-occurrence patterns of photobiont and cystobasidiomycete yeast associated with common epiphytic lichen species. New Phytol. 2020, 227, 1362–1375. [Google Scholar] [CrossRef]
- Lendemer, J.C.; Keepers, K.G.; Tripp, E.A.; Pogoda, C.S.; McCain, C.M.; Kane, N.C. A taxonomically broad metagenomic survey of 339 species spanning 57 families suggests cystobasidiomycete yeasts are not ubiquitous across all lichens. Am. J. Bot. 2019, 106, 1090–1095. [Google Scholar] [CrossRef]
- Steinová, J.; Škaloud, P.; Yahr, R.; Bestová, H.; Muggia, L. Reproductive and dispersal strategies shape the diversity of mycobiont-photobiont association in Cladonia lichens. Mol. Phylogenet. Evol. 2019, 134, 226–237. [Google Scholar] [CrossRef]
- Li, A.H.; Yuan, F.X.; Groenewald, M.; Bensch, K.; Yurkov, A.M.; Li, K.; Han, P.J.; Guo, L.D.; Aime, M.C.; Sampaio, J.P.; et al. Diversity and phylogeny of basidiomycetous yeasts from plant leaves and soil: Proposal of two new orders, three new families, eight new genera and one hundred and seven new species. Stud. Mycol. 2020, 96, 17–140. [Google Scholar] [CrossRef]
- Paul, F.; Otte, J.; Schmitt, I.; Dal Grande, F. Comparing Sanger sequencing and high-throughput metabarcoding for inferring photobiont diversity in lichens. Sci. Rep. 2018, 8, 8624. [Google Scholar] [CrossRef] [Green Version]
- Tabacchioni, S.; Chiarini, L.; Bevivino, A.; Cantale, C.; Dalmastri, C. Bias caused by using different isolation media for assessing the genetic diversity of a natural microbial population. Microb. Ecol. 2000, 40, 169–176. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kinoshita, Y.; Yoshimura, I. Culture of thallus fragments and redifferentiation of lichens. In Protocols in Lichenology: Culturing, Biochemistry, Ecophysiology and Use in Biomonitoring; Kranner, I.C., Beckett, R.P., Varma, A.K., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; pp. 34–46. ISBN 978-3-642-56359-1. [Google Scholar]
- Slatko, B.E.; Gardner, A.F.; Ausubel, F.M. Overview of next generation sequencing technologies. Curr. Protoc. Mol. Biol. 2019, 122, e59. [Google Scholar] [CrossRef] [PubMed]
- Op De Beeck, M.; Lievens, B.; Busschaert, P.; Declerck, S.; Vangronsveld, J.; Colpaert, J.V. Comparison and validation of some ITS primer pairs useful for fungal metabarcoding studies. PLoS ONE 2014, 9, e97629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makiola, A.; Dickie, I.A.; Holdaway, R.J.; Wood, J.R.; Orwin, K.H.; Lee, C.K.; Glare, T.R. Biases in the metabarcoding of plant pathogens using rust fungi as a model system. Microbiologyopen 2019, 8, e00780. [Google Scholar] [CrossRef] [Green Version]
- Castaño, C.; Berlin, A.; Durling, M.B.; Ihrmark, K.; Lindahl, B.D.; Stenlid, J.; Clemmensen, K.E.; Olson, Å. Optimized metabarcoding with Pacific biosciences enables semi-quantitative analysis of fungal communities. New Phytol. 2020, 228, 1149–1158. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Tooming-Klunderud, A.; Anslan, S. PacBio metabarcoding of Fungi and other eukaryotes: Errors, biases and perspectives. New Phytol. 2018, 217, 1370–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauvert, C.; Bu, M.; Lesur, I.; Vallance, J.; Fauchery, L.; Vacher, C. Bioinformatics matters: The accuracy of plant and soil fungal community data is highly dependent on the metabarcoding pipeline. Fungal Ecol. 2019, 41, 23–33. [Google Scholar] [CrossRef]
- Lena, E.; Sundy, F.E.; Luis, M.; Martin, P.M.; Inger, S.; Håvard, S. The influence of intraspecific sequence variation during DNA metabarcoding: A case study of eleven fungal species. Mol. Ecol. Resour. 2021, 21, 1141–1148. [Google Scholar] [CrossRef]
- Ahmadjian, V. Studies on Lichenized Fungi. Bryologist 1961, 64, 168–179. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T.; Robert, V. Chapter 7—Methods for isolation, phenotypic characterization and maintenance of yeasts. In The Yeasts, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: London, UK, 2011; pp. 87–110. ISBN 978-0-444-52149-1. [Google Scholar]
- do Carmo-Sousa, L.; Phaff, H.J. An improved method for the detection of spore discharge in the Sporobolomycetaceae. J. Bacteriol. 1962, 83, 434–435. [Google Scholar] [CrossRef] [Green Version]
- Nakada, M.; Tanaka, C.; Tsunewaki, K.; Tsuda, M. RFLP analysis for species separation in the genera Bipolaris and Curvularia. Mycoscience 1994, 35, 271–278. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. 1990, 315–322. [Google Scholar] [CrossRef]
- Hopple, J.S.; Vilgalys, R. Phylogenetic relationships among coprinoid taxa and allies based on data from restriction site mapping of nuclear rDNA. Mycologia 1994, 86, 96–107. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree—Tree Figure Drawing Tool; University of Edinburgh: Edinburgh, Scotland, UK, 2018. [Google Scholar]
- RStudio Team RStudio: Integrated Development Environment for R 2016. Available online: http://www.rstudio.com/ (accessed on 2 October 2016).
- R Core Team R: A Language and Environment for Statistical Computing 2018. Available online: https://www.R-project.org/ (accessed on 1 October 2018).
- Yu, G. Using ggtree to visualize data on tree-like structures. Curr. Protoc. Bioinforma 2020, 69, e96. [Google Scholar] [CrossRef]
- Wang, L.-G.; Lam, T.T.-Y.; Xu, S.; Dai, Z.; Zhou, L.; Feng, T.; Guo, P.; Dunn, C.W.; Jones, B.R.; Bradley, T.; et al. Treeio: An R Package for Phylogenetic Tree Input and Output with Richly Annotated and Associated Data. Mol. Biol. Evol. 2020, 37, 599–603. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Hannon, G.J. FASTX-Toolkit. 2010. Available online: http://hannonlab.cshl.edu/fastx_toolkit/ (accessed on 2 October 2018).
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, H.R.; Schigel, D.; Tedersoo, L.; Larsson, K.H.; May, T.W.; Taylor, A.F.S.; Jeppesen, T.S.; Frøslev, T.G.; Lindahl, B.D.; et al. The Taxon Hypothesis Paradigm—On the Unambiguous Detection and Communication of Taxa. Microorganisms 2020, 8, 1910. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Nakase, T.; Tsuzuki, S.; Lee, F.-L.; Sugita, T.; Jindamorakot, S.; Jan-ngam, H.; Potacharoen, W.; Tanticharoen, M.; Kudo, T.; Takashima, M. Sporobolomyces magnisporus sp. nov., a new yeast species in the Erythrobasidium cluster isolated from plants in Taiwan. J. Gen. Appl. Microbiol. 2003, 49, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Pohl, C.H.; Smit, M.S.; Albertyn, J. Rhodotorula bloemfonteinensis sp. nov., Rhodotorula eucalyptica sp. nov., Rhodotorula orientis sp. nov. and Rhodotorula pini sp. nov., yeasts isolated from monoterpene-rich environments. Int. J. Syst. Evol. Microbiol. 2011, 61, 2320–2327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, F.; Liu, Y.; Li, N.; Yao, S.; Li, N.; Wang, W.; Cheng, C. Microsporomyces hainanensis sp. nov., Isolated from Hybrid Rice (Oryza sativa L.) Seeds. Curr. Microbiol. 2016, 73, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Glushakova, A.M.; Chernov, I.Y. Seasonal dynamics of the structure of epiphytic yeast communities. Microbiology 2010, 79, 830–839. [Google Scholar] [CrossRef]
- Jumpponen, A.; Jones, K.L. Seasonally dynamic fungal communities in the Quercus macrocarpa phyllosphere differ between urban and nonurban environments. New Phytol. 2010, 186, 496–513. [Google Scholar] [CrossRef]
- Guo, Z.; Wang, Y.; Hou, Q.; Li, W.; Zhao, H.; Sun, Z.; Zhang, Z. Halobasidium xiangyangense gen. nov., sp. nov., a new xyloseutilizing yeast in the family cystobasidiaceae, isolated from the pickling sauce used to make datoucai, a high-salt fermented food. Int. J. Syst. Evol. Microbiol. 2019, 69, 139–145. [Google Scholar] [CrossRef]
- Yurkov, A.M.; Kachalkin, A.V.; Daniel, H.M.; Groenewald, M.; Libkind, D.; de Garcia, V.; Zalar, P.; Gouliamova, D.E.; Boekhout, T.; Begerow, D. Two yeast species Cystobasidium psychroaquaticum f.a. sp. nov. and Cystobasidium rietchieii f.a. sp. nov. isolated from natural environments, and the transfer of Rhodotorula minuta clade members to the genus Cystobasidium. Antonie van Leeuwenhoek 2015, 107, 173–185. [Google Scholar] [CrossRef]
- Takashima, M.; Sugita, T.; Van, B.H.; Nakamura, M.; Endoh, R.; Ohkuma, M. Taxonomic richness of yeasts in Japan within subtropical and cool temperate areas. PLoS ONE 2012, 7, e50784. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Wei, Z.; Pu, S.; Xiang, M.-; Yan, A.; Zhang, Y.; Wang, X. Diversity of endophytic fungi of Paeonia lactiflora Pallas and screening for fungal paeoniflorin producers. FEMS Microbiol. Lett. 2018, 365, fny263. [Google Scholar] [CrossRef]
- Zhang, W.; Wendel, J.F.; Clark, L.G. Bamboozled again! Inadvertent isolation of fungal rDNA sequences from bamboos (Poaceae: Bambusoideae). Mol. Phylogenet. Evol. 1997, 8, 205–217. [Google Scholar] [CrossRef]
- Egidi, E.; Delgado-Baquerizo, M.; Plett, J.M.; Wang, J.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K. A few Ascomycota taxa dominate soil fungal communities worldwide. Nat. Commun. 2019, 10, 2369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boraks, A.; Amend, A.S. Fungi in soil and understory have coupled distribution patterns. PeerJ 2021, 9, e11915. [Google Scholar] [CrossRef] [PubMed]
- Henk, D.A.; Vilgalys, R. Molecular phylogeny suggests a single origin of insect symbiosis in the Pucciniomycetes with support for some relationships within the genus Septobasidium. Am. J. Bot. 2007, 94, 1515–1526. [Google Scholar] [CrossRef]
- Yamamoto, W. Species of Septobasidium from Japan. Jpn. J. Phytopathol. 1956, 21, 9–12. [Google Scholar] [CrossRef]
- Wang, Q.M.; Yurkov, A.M.; Göker, M.; Lumbsch, H.T.; Leavitt, S.D.; Groenewald, M.; Theelen, B.; Liu, X.Z.; Boekhout, T.; Bai, F.Y. Phylogenetic classification of yeasts and related taxa within Pucciniomycotina. Stud. Mycol. 2015, 81, 149–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specimen ID | Lichen Species | Locality | Collecting Date | Lichen Thalli Isolating | Meta-Barcoding | Herbarium ID |
|---|---|---|---|---|---|---|
| 1 | Cladonia rei | Kimitsu, Chiba N35.18731, E140.11192 | Mar. 2017 | X | CBM-FL-203638 | |
| 2 | Cladonia rei | Odawara, Kanagawa N35.24410, E139.11775 | Oct. 2017 | X | CBM-FL-206689 | |
| 3 | Cladonia rei | Ichihara, Chiba N35.30606, E140.12924 | Jun. 2018 | X | CBM-FL-103149 | |
| 4 | Cladonia rei | Ichihara, Chiba N35.30591, E140.12884 | Jun. 2018 | X | CBM-FL-103150 | |
| 5 | Cladonia rei | Kimitsu, Chiba N35.19563, E140.07037 | Jun. 2018 | X | CBM-FL-103152 | |
| 6 | Cladonia rei | Kimitsu, Chiba N35.19563, E140.07037 | Jun. 2018 | X | CBM-FL-103153 | |
| 7 | Cladonia rei | Kimitsu, Chiba N35.20902, E140.09221 | Jun. 2018 | X | CBM-FL-103154 | |
| 8 | Cladonia rei | Kimitsu, Chiba N35.20820, E140.09119 | Jun. 2018 | X | CBM-FL-103155 | |
| 9 | Cladonia rei | Ichihara, Chiba N35.305886, E140.12929 | Nov. 2018 | X | X | CBM-FL-205144 |
| 10 | Cladonia rei | Ichihara, Chiba N35.305886, E140.12929 | Nov. 2018 | X | X | CBM-FL-205145 |
| 11 | Cladonia rei | Ichihara, Chiba N35.305886, E140.12929 | Nov. 2018 | X | X | CBM-FL-205146 |
| 12 | Cladonia rei | Ichihara, Chiba N35.305858, E140.128931 | Nov. 2018 | X | X | CBM-FL-205147 |
| 13 | Cladonia rei | Kimitsu, Chiba N35.195252, E140.07036 | Nov. 2018 | X | X | CBM-FL-205148 |
| 14 | Cladonia rei | Kimitsu, Chiba N35.19493, E140.071076 | Nov. 2018 | X | X | CBM-FL-205149 |
| 15 | Cladonia rei | Kimitsu, Chiba N35.19493, E140.071076 | Nov. 2018 | X | X | CBM-FL-205150 |
| Lineage | Strain/Voucher | ITS | LSU | Specimen ID |
|---|---|---|---|---|
| Microsporomyces cladoniophilus sp. nov | 109_1 | MZ505462 | MZ513978 | 2 |
| Microsporomyces cladoniophilus sp. nov | 109_2 | MZ505463 | MZ513979 | 2 |
| Microsporomyces cladoniophilus sp. nov | 211_1 | MZ505473 | MZ513980 | 1 |
| Microsporomyces cladoniophilus sp. nov | 211_11A | MZ505468 | MZ513983 | 1 |
| Microsporomyces cladoniophilus sp. nov | 211_12 | MZ505469 | MZ513984 | 1 |
| Microsporomyces cladoniophilus sp. nov | 211_13 | MZ505470 | MZ513985 | 1 |
| Microsporomyces cladoniophilus sp. nov | 211_2 | MZ505474 | – | 3 |
| Microsporomyces cladoniophilus sp. nov | 211_3 | MZ505475 | – | 3 |
| Microsporomyces cladoniophilus sp. nov | 211_5 | MZ505476 | MZ513981 | 3 |
| Microsporomyces cladoniophilus sp. nov | 211_m5 | MZ505482 | MZ513990 | 3 |
| Microsporomyces cladoniophilus sp. nov | 211_8a | MZ505477 | MZ513982 | 3 |
| Microsporomyces cladoniophilus sp. nov | 211_m4 | MZ505481 | MZ513988 | 3 |
| Microsporomyces cladoniophilus sp. nov | 211_m4A | MZ505480 | MZ513989 | 3 |
| Microsporomyces cladoniophilus sp. nov | 213_3 | MZ505484 | MZ513992 | 3 |
| Microsporomyces cladoniophilus sp. nov | 213_7 | MZ505485 | MZ513993 | 3 |
| Microsporomyces cladoniophilus sp. nov | 213_8 | MZ505486 | MZ513994 | 3 |
| Microsporomyces cladoniophilus sp. nov | 216_7 | MZ505492 | – | 3 |
| Microsporomyces cladoniophilus sp. nov | 222_m2_1 | MZ505493 | MZ514000 | 3 |
| Microsporomyces cladoniophilus sp. nov | 223_7 | MZ505495 | MZ514001 | 3 |
| Microsporomyces cladoniophilus sp. nov | 226_17 | MZ505496 | – | 3 |
| Microsporomyces cladoniophilus sp. nov | 226_20 | MZ505498 | MZ514006 | 3 |
| Microsporomyces cladoniophilus sp. nov | 226_7 | MZ505500 | – | 4 |
| Microsporomyces cladoniophilus sp. nov | 226_9 | MZ505501 | MZ514004 | 4 |
| Microsporomyces cladoniophilus sp. nov | 226_m3 | MZ505502 | MZ514007 | 4 |
| Microsporomyces Sp.2 | 211_15 | MZ505471 | MZ513986 | 6 |
| Microsporomyces Sp.2 | 216_16 | MZ505487 | MZ513997 | 13 |
| Microsporomyces Sp.6 | 211_16 | MZ505472 | – | 6 |
| Microsporomyces Sp.6 | 211_8 | MZ505478 | – | 6 |
| Microsporomyces Sp.5 | 20_12 | MZ505464 | MZ513974 | 4 |
| Microsporomyces Sp.5 | 20_14 | MZ505465 | MZ513975 | 6 |
| Microsporomyces Sp.5 | 20_19 | MZ505466 | MZ513976 | 6 |
| Microsporomyces Sp.5 | 20_20 | MZ505467 | MZ513977 | 6 |
| Microsporomyces Sp.5 | 211_m2 | MZ505479 | MZ513987 | 14 |
| Microsporomyces Sp.5 | 216_2 | MZ505490 | MZ513996 | 13 |
| Microsporomyces Sp.3 | 216_25 | MZ505489 | MZ513998 | 13 |
| Microsporomyces Sp.3 | 216_31 | MZ505491 | MZ513999 | 13 |
| Microsporomyces Sp.3 | 223_13 | MZ505494 | MZ514002 | 13 |
| Microsporomyces Sp.4 | 213_1 | MZ505483 | MZ513991 | 10 |
| Microsporomyces Sp.4 | 216_1 | MZ505488 | MZ513995 | 10 |
| Microsporomyces Sp.4 | 226_19 | MZ505497 | MZ514005 | 13 |
| Microsporomyces Sp.4 | 226_2 | MZ505499 | MZ514003 | 13 |
| Microsporomyces cladoniophilus sp. nov. | Microsporomyces magnisporus a | Microsporomyces pseudomagnisporus d | Microsporomyces rubellus d | Microsporomyces ellipsoideus d | Microsporomyces pini b | Microspomyces bloemfonteinensis b | Microsporomyces orientalisb | Microsporomyces hainanensis c | |
|---|---|---|---|---|---|---|---|---|---|
| D + Glucose | + | + | + | + | + | + | + | + | + |
| L-Sorbose | w/s | +(w) | + | + | + | − | − | − | n/a |
| Sucrose | + | +(−) | + | − | − | − | − | + | n/a |
| Maltose | + | +(w) | + | + | + | − | + | + | − |
| D + Cellobose | + | − | − | + | + | − | + | + | + |
| D + Trehalose | + | + | − | − | − | − | + | + | + |
| Lactose | +/w/l | − | w | + | + | + | + | + | − |
| Melibiose | − | + | − | − | − | − | + | + | n/a |
| Raffinose | w/l | +(w) | w | + | w | − | + | + | n/a |
| D-Melezitose | − | +(w) | w | + | + | − | + | w | − |
| Inulin | − | + | w | + | − | w | + | + | − |
| Starch soluble | − | +(w) | l | − | − | − | − | − | + |
| D + Xylose | + | +(s)/− | − | − | + | − | + | w | w |
| L + Arabinose | − | +(w)/− | − | − | − | − | − | + | − |
| D-Arabinose | − | − | − | − | − | − | − | + | n/a |
| D_Ribose | − | +(s)/− | w | − | − | − | + | + | + |
| α_L + Rhamnose | − | +(w)/− | − | − | − | − | w | + | n/a |
| D-Glucosamine | − | +(l/w)/− | − | − | − | − | − | − | n/a |
| Glycerol | + | +/− | − | − | − | − | − | − | n/a |
| meso_Erythritol | − | − | − | − | − | + | − | + | − |
| D-Mannitol | + | +(w) | w | lw | − | + | w | w | + |
| D_Glucitol | + | +(w) | w | w | + | − | w | + | − |
| α methyl-D + Glucoside | − | +(w)/− | w | − | + | − | − | + | n/a |
| Gluconic acid | + | + | n/a | n/a | n/a | + | + | + | n/a |
| DL Lactic acid | − | +(w) | − | v | − | + | + | + | − |
| Succinic acid salt | − | + | w | − | lw | + | + | + | − |
| Citric acid | − | +(l) | − | − | − | + | − | + | − |
| D-Glucuronic acid | + | + | n/a | n/a | n/a | n/a | n/a | n/a | n/a |
| Inositol | − | − | − | − | − | − | w | − | − |
| Salicin | +/w | +(w)/− | − | v | + | + | + | + | − |
| D + Galacturonic acid | + | +(s) | n/a | n/a | n/a | n/a | n/a | n/a | n/a |
| 2,3-Butanediol | − | +(l/w)/− | n/a | n/a | n/a | n/a | n/a | n/a | n/a |
| D + Glucono-1,5-lactone | + | + | n/a | n/a | n/a | n/a | n/a | n/a | n/a |
| Xylitol | + | +(w) | n/a | n/a | n/a | n/a | n/a | n/a | n/a |
| Ribitol | + | +(w) | w | + | − | − | + | − | n/a |
| L-Arabitol | − | − | n/a | n/a | n/a | n/a | n/a | n/a | n/a |
| Galactitol | − | − | − | + | − | − | − | − | n/a |
| 1,2-Propanediol | − | − | n/a | n/a | n/a | n/a | n/a | n/a | n/a |
| Hexadecane | − | n/a | − | − | − | − | − | − | − |
| D + Galactose | + | +(w) | + | + | + | − | − | − | n/a |
| Ethanol | w | − | + | − | − | − | w | + | − |
| Methanol | − | − | − | − | − | − | − | − | − |
| N-Acetyl-D(+)Glucosamine | +/w/− | + | − | n/a | n/a | + | − | + | n/a |
| 2-keto-D-gluconate (2-Oxoglutaric acid) | − | + | n/a | n/a | n/a | n/a | n/a | n/a | n/a |
| Cadaverine | w | − | + | − | − | + | n/a | + | n/a |
| Potassium nitrate | + | + | + | v | + | − | − | + | + |
| Sodium nitrite | w | + | − | − | − | − | + | + | + |
| Ethylamine | − | − | + | − | − | − | − | + | − |
| L-Lysine | + | − | + | v | w | + | − | + | − |
| Fermentation of glucose | − | − | − | − | − | − | − | − | − |
| Growth in vitamin-free medium | − | − | + | − | − | − | + | + | n/a |
| Growth at 22 °C | + | + | − | + | + | n/a | n/a | n/a | n/a |
| Growth at 25 °C | + | + | − | + | + | + | + | + | + |
| Growth at 30 °C | − | + | − | + | − | n/a | n/a | n/a | n/a |
| Growth at 32 °C | − | − | − | w | − | n/a | n/a | n/a | n/a |
| Growth at 35 °C | − | − | − | − | − | n/a | n/a | n/a | − |
| Growth with 50% glucose | − | − | − | − | − | − | − | − | n/a |
| Starch like compounds formation | − | − | − | − | − | − | − | − | − |
| Hydrolysis of Urea | + | + | + | + | + | + | − | − | n/a |
| Diazonium blue B reaction | + | + | + | + | + | n/a | n/a | n/a | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, N.-H.; Nguyen, P.-T.; Otake, H.; Nagata, A.; Hirano, N.; Imanishi-Shimizu, Y.; Shimizu, K. Biodiversity of Basidiomycetous Yeasts Associated with Cladonia rei Lichen in Japan, with a Description of Microsporomyces cladoniophilus sp. nov. J. Fungi 2023, 9, 473. https://doi.org/10.3390/jof9040473
Nguyen N-H, Nguyen P-T, Otake H, Nagata A, Hirano N, Imanishi-Shimizu Y, Shimizu K. Biodiversity of Basidiomycetous Yeasts Associated with Cladonia rei Lichen in Japan, with a Description of Microsporomyces cladoniophilus sp. nov. Journal of Fungi. 2023; 9(4):473. https://doi.org/10.3390/jof9040473
Chicago/Turabian StyleNguyen, Ngoc-Hung, Phuong-Thao Nguyen, Hitomi Otake, Ayana Nagata, Nobuharu Hirano, Yumi Imanishi-Shimizu, and Kiminori Shimizu. 2023. "Biodiversity of Basidiomycetous Yeasts Associated with Cladonia rei Lichen in Japan, with a Description of Microsporomyces cladoniophilus sp. nov" Journal of Fungi 9, no. 4: 473. https://doi.org/10.3390/jof9040473