
Improving Methanol Utilization by Reducing Alcohol Oxidase Activity and Adding Co-Substrate of Sodium Citrate in Pichia pastoris

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media, and Cultivation
2.2. Plasmids and Strains Construction
2.3. Growth Tests in 48-Well Plates
2.4. Formaldehyde Quantification
2.5. Alcohol Oxidase Activity Determination
2.6. The Biomass and Methanol Conversion Rate Determination
2.7. Intracellular NADH and NAD+ Quantitation
2.8. Transcriptome Analysis
2.9. Statistical Analysis
3. Results
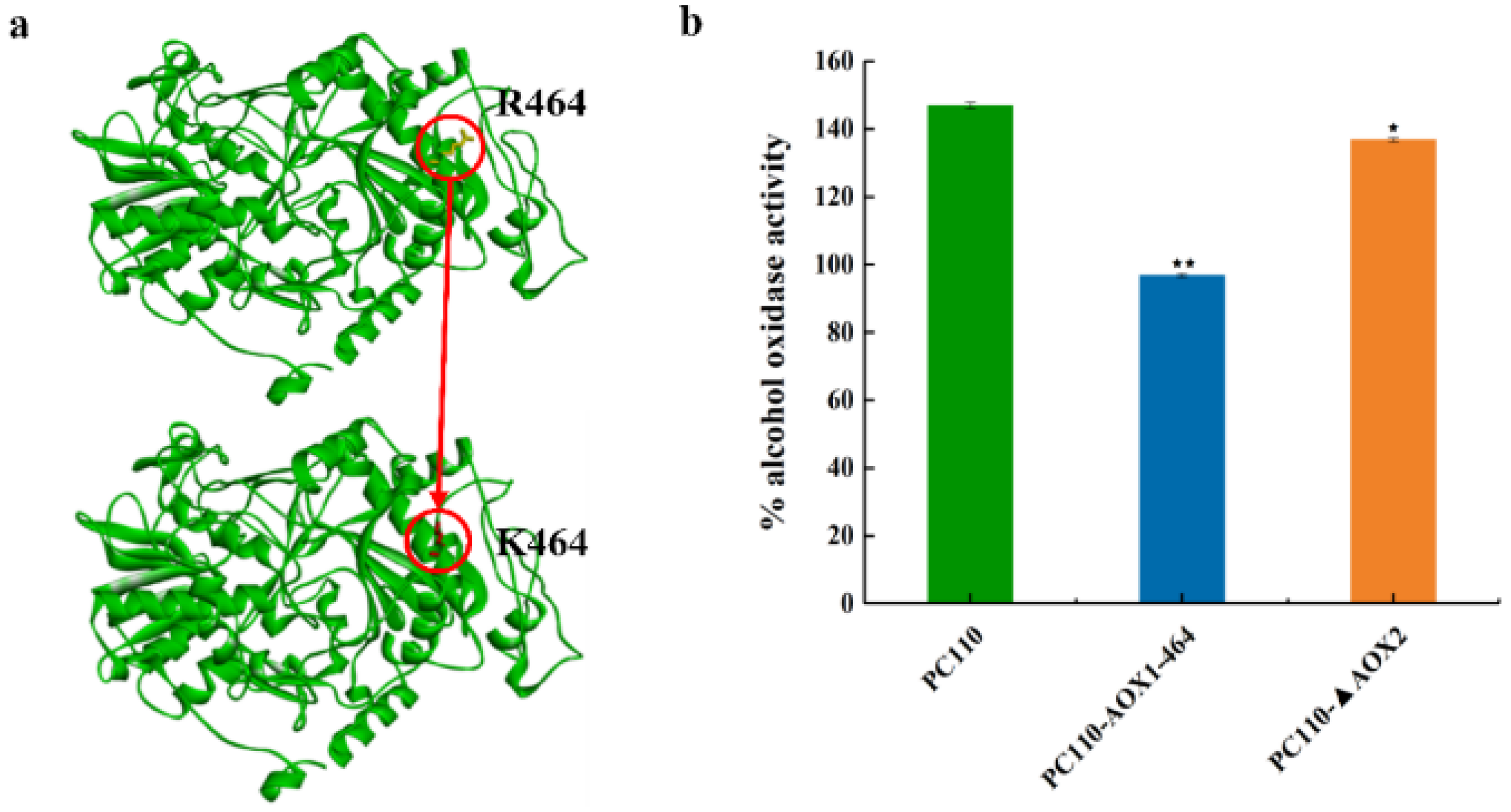
3.1. Construction of P. pastoris Strains with Reduced AOX Activity by Point Mutation of AOX1 or Deletion of AOX2
3.2. Data of CO2 Loss and Biomass Accumulation of P. pastoris with Reduced AOX Activity via Genome-Scale Metabolic Model (GSMM) Calculations
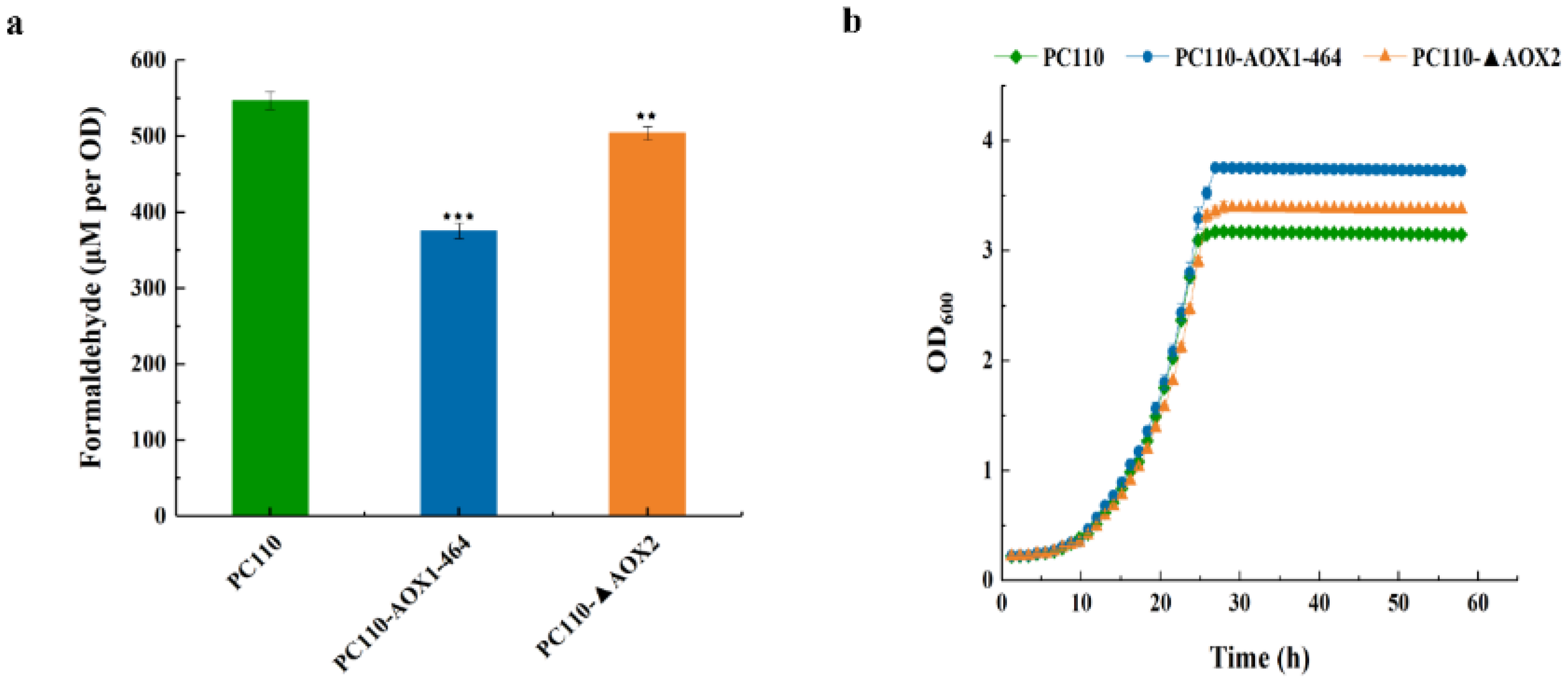
3.3. Reduced AOX Activity Decreased Intracellular Formaldehyde Accumulation and Increased Cell Growth on Methanol in P. pastoris
3.4. Reduced AOX Activity Promoted the Methanol Utilization Efficiency in P. pastoris
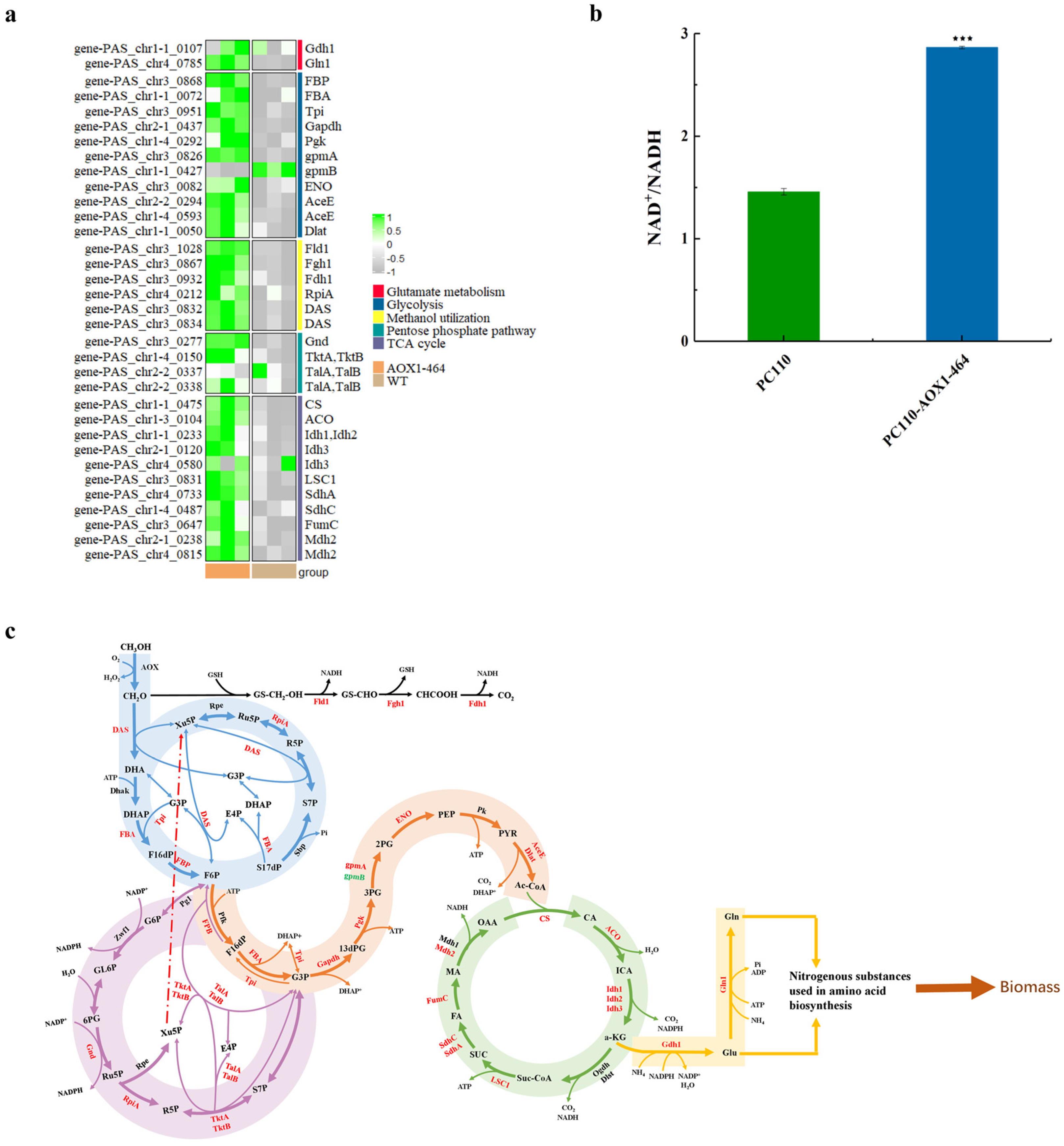
3.5. Transcriptomics Analysis Revealed the Mechanism of Enhanced Methanol Utilization Associated with Reduced AOX Activity in P. pastoris
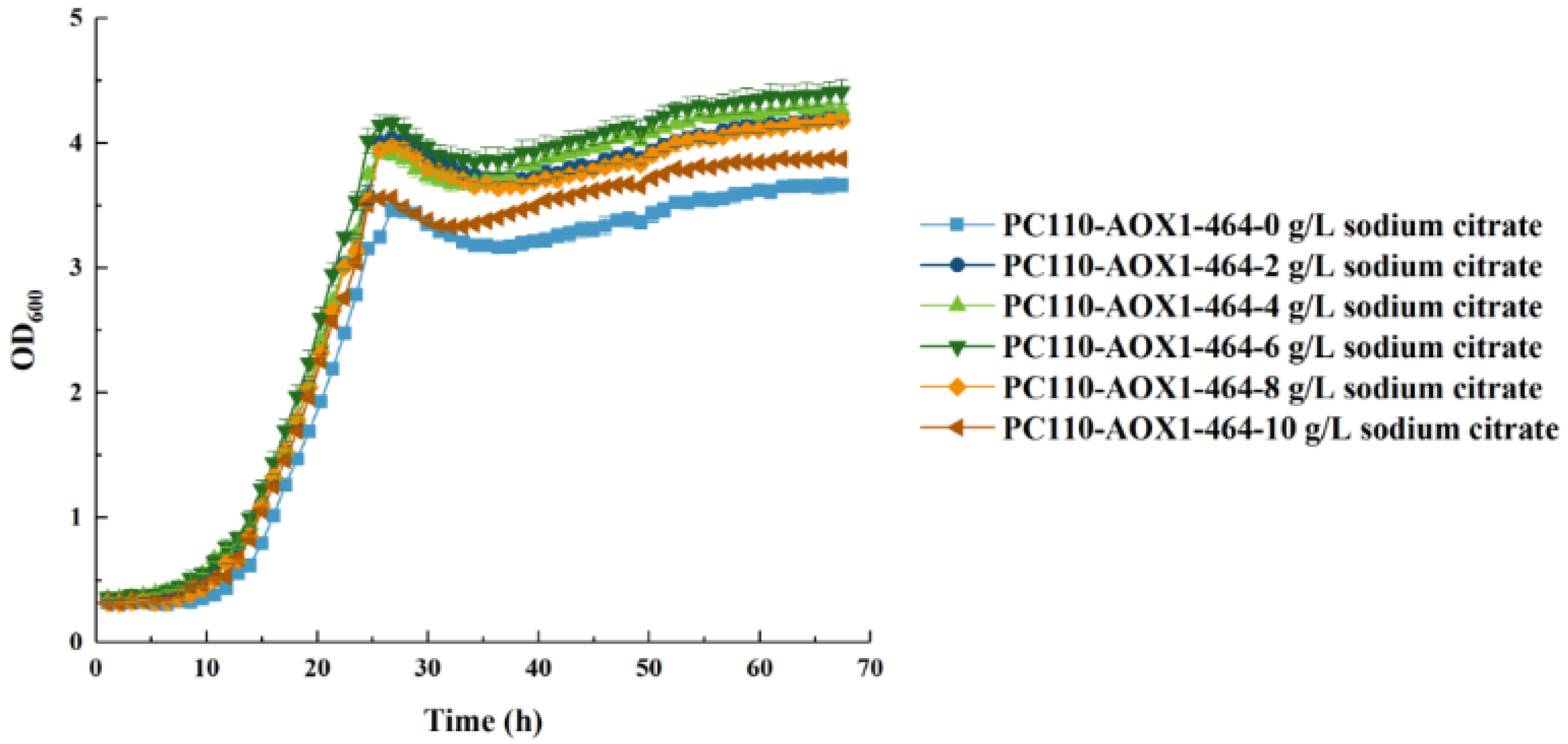
3.6. Addition of Co-Substrate of Sodium Citrate Further Promoted Cell Growth and Biomass Accumulation of PC110-AOX1-464 in Methanol-Minimal Medium
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zeng, A.P. New bioproduction systems for chemicals and fuels: Needs and new development. Biotechnol. Adv. 2019, 37, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Kircher, M. Bioeconomy-present status and future needs of industrial value chains. New Biotechnol. 2021, 60, 96–104. [Google Scholar] [CrossRef]
- Chen, C.T.; Chen, F.Y.; Bogorad, I.W.; Wu, T.Y.; Zhang, R.; Lee, A.S.; Liao, J.C. Synthetic methanol auxotrophy of Escherichia coli for methanol-dependent growth and production. Metab. Eng. 2018, 54, 292. [Google Scholar] [CrossRef] [PubMed]
- Gleizer, S.; Ben-Nissan, R.; Bar-On, Y.M.; Antonovsky, N.; Noor, E.; Zohar, Y.; Jona, G.; Krieger, E.; Shamshoum, M.; Bar-Even, A.; et al. Conversion of Escherichia coli to generate all biomass carbon from CO2. Cell 2019, 179, 1255–1263.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gassler, T.; Sauer, M.; Gasser, B.; Egermeier, M.; Troyer, C.; Causon, T.; Hann, S.; Mattanovich, D.; Steiger, M.G. The industrial yeast Pichia pastoris is converted from a heterotroph into an autotroph capable of growth on CO2. Nat. Biotechnol. 2020, 38, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Cotton, C.A.; Claassens, N.J.; Benito-Vaquerizo, S.; Bar-Even, A. Renewable methanol and formate as microbial feedstocks. Curr. Opin. Biotechnol. 2020, 62, 168–180. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Kerkhoven, E.J.; Nielsen, J. Barriers and opportunities in bio-based production of hydrocarbons. Nat. Energy 2018, 3, 925–935. [Google Scholar] [CrossRef]
- Schrader, J.; Schilling, M.; Holtmann, D.; Sell, D.; Filho, M.V.; Marx, A.; Vorholt, J.A. Methanol-based industrial biotechnology: Current status and future perspectives of methylotrophic bacteria. Trends Biotechnol. 2009, 27, 107–115. [Google Scholar] [CrossRef]
- Behr, A. Methanol: The basic chemical and energy feedstock of the future. Asinger’s Vision Today. Edited by Martin Bertau, Heribert Offermanns, Ludolf Plass, Friedrich Schmidt and Hans-Jürgen Wernicke. Angew. Chem. Int. Ed. 2014, 53, 12674. [Google Scholar] [CrossRef]
- Kuk, K.S.; Singh, R.K.; Nam, D.H.; Singh, R.; Lee, J.K.; Park, C.B. Photoelectrochemical reduction of carbon dioxide to methanol through a highly efficient enzyme cascade. Angew. Chem. Int. Ed. 2017, 56, 3827–3832. [Google Scholar] [CrossRef]
- Patel Sanjay, K.S.; Min, S.J.; Rahul, K.G.; Yale, J.; Vipin, C.K.; Sun, C.K.; Byung, K.C.; Dong, R.K.; Lee, J.K. Hierarchical macroporous particles for efficient whole-cell immobilization: Application in bioconversion of greenhouse gases to methanol. ACS Appl. Mater. Interfaces 2019, 11, 18968–18977. [Google Scholar] [CrossRef]
- Rafiee, A. Optimal design issues of a methanol synthesis reactor from CO2 hydrogenation. Chem. Eng. Technol. 2020, 43, 2092–2099. [Google Scholar] [CrossRef]
- Chen, F.Y.; Jung, H.W.; Tsuei, C.Y.; Liao, J.C. Converting Escherichia coli to a synthetic methylotroph growing solely on methanol. Cell 2020, 182, 933–946. [Google Scholar] [CrossRef]
- Vogl, T.; Sturmberger, L.; Kickenweiz, T.; Wasmayer, R.; Schmid, C.; Hatzl, A.M.; Gerstmann, M.A.; Pitzer, J.; Wagner, M.; Thallinger, G.G.; et al. A toolbox of diverse promoters related to methanol utilization: Functionally verified parts for heterologous pathway expression in Pichia pastoris. ACS Synth. Biol. 2016, 5, 172–186. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.Q.; Esposito, R.D.; Gowani, J.; Goodman, J.M. Alcohol oxidase and dihydroxyacetone synthase, the abundant peroxisomal proteins of methylotrophic yeasts, assemble in different cellular compartments. J. Cell Sci. 2001, 114, 2863–2868. [Google Scholar] [CrossRef] [PubMed]
- Kunze, G.; Kang, H.A.; Gellissen, G. Hansenula polymorpha (Pichia angusta): Biology and applications. In Yeast Biotechnology: Diversity and Applications; Satyanarayana, T., Kunze, G., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 47–64. [Google Scholar]
- Lee, B.; Yurimoto, H.; Sakai, Y.; Kato, N. Physiological role of the glutathione-dependent formaldehyde dehydrogenase in the methylotrophic yeast Candida boidinii. Microbiology 2002, 148, 2697–2704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, T.; Ito, T.; Fujimura, S.; Chikui, M.; Mizumura, T.; Miyaji, T.; Yurimoto, H.; Kato, N.; Sakai, Y.; Tomizuka, N. Molecular characterization of the glutathione-dependent formaldehyde dehydrogenase gene FLD1 from the methylotrophic yeast Pichia methanolica. Yeast 2004, 21, 445–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartner, F.S.; Glieder, A. Regulation of methanol utilization pathway genes in yeasts. Microb. Cell Factories 2006, 5, 39. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, T.; Mizumura, T.; Mukaiyama, H.; Miyaji, T.; Yurimoto, H.; Kato, N.; Sakai, Y.; Tomizuka, N. Physiological role of the second alcohol oxidase gene MOD2 in the methylotrophic growth of Pichia methanolica. Yeast 2003, 20, 1067–1073. [Google Scholar]
- Yurimoto, H.; Oku, M.; Sakai, Y. Yeast methylotrophy: Metabolism, gene regulation and peroxisome homeostasis. Int. J. Microbiol. 2011, 2011, 101298. [Google Scholar] [CrossRef] [Green Version]
- Yamada, R.; Ogura, K.; Kimoto, Y.; Ogino, H. Toward the construction of a technology platform for chemicals production from methanol: D-lactic acid production from methanol by an engineered yeast Pichia pastoris. World J. Microbiol. Biotechnol. 2019, 35, 37. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.R.; Zhang, W.J.; Wang, Y.; Sheng, J.Y.; Xu, R.R.; Li, J.H.; Du, G.C.; Kang, Z. Biosynthesis of non-animal chondroitin sulfate from methanol using genetically engineered Pichia pastoris. Green Chem. 2021, 23, 4365–4374. [Google Scholar] [CrossRef]
- Guo, F.; Dai, Z.; Peng, W.; Zhang, S.; Zhou, J.; Ma, J.; Dong, W.; Xin, F.; Zhang, W.; Jiang, M. Metabolic engineering of Pichia pastoris for malic acid production from methanol. Biotechnol. Bioeng. 2021, 118, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Li, Y.; Yu, W.; Zhou, Y.J. Rescuing yeast from cell death enables overproduction of fatty acids from sole methanol. Nat. Metab. 2022, 4, 932–943. [Google Scholar] [CrossRef]
- Zavec, D.; Troyer, C.; Maresch, D.; Altmann, F.; Hann, S.; Gasser, B.; Mattanovich, D. Beyond alcohol oxidase: The methylotrophic yeast Komagataella phaffii utilizes methanol also with its native alcohol dehydrogenase Adh2. FEMS Yeast Res. 2021, 21, foab009. [Google Scholar] [CrossRef]
- Moser, J.W.; Prielhofer, R.; Gerner, S.M.; Graf, A.B.; Wilson, I.B.; Mattanovich, D.; Dragosits, M. Implications of evolutionary engineering for growth and recombinant protein production in methanol-based growth media in the yeast Pichia pastoris. Microb. Cell Factories 2017, 16, 49. [Google Scholar] [CrossRef] [Green Version]
- Cai, P.; Duan, X.; Wu, X.; Gao, L.; Ye, M.; Zhou, Y.J. Recombination machinery engineering facilitates metabolic engineering of the industrial yeast Pichia pastoris. Nucleic Acids Res. 2021, 49, 7791–7805. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Buijs, N.A.; Zhu, Z.; Gómez, D.O.; Boonsombuti, A.; Siewers, V.; Nielsen, J. Harnessing yeast peroxisomes for biosynthesis of fatty-acid-derived biofuels and chemicals with relieved side-pathway competition. J. Am. Chem. Soc. 2016, 138, 15368–15377. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Buijs, N.A.; Zhu, Z.; Qin, J.; Siewers, V.; Nielsen, J. Production of fatty acid-derived oleochemicals and biofuels by synthetic yeast cell factories. Nat. Commun. 2016, 7, 11709. [Google Scholar] [CrossRef] [Green Version]
- Woolston, B.M.; King, J.R.; Reiter, M.; Hove, B.V.; Stephanopoulos, G. Improving formaldehyde consumption drives methanol assimilation in engineered E. coli. Nat. Commun. 2018, 9, 2387. [Google Scholar] [CrossRef] [Green Version]
- Cai, P.; Wu, X.; Deng, J.; Gao, L.; Shen, Y.; Yao, L.; Zhou, Y.J. Methanol biotransformation toward high-level production of fatty acid derivatives by engineering the industrial yeast Pichia pastoris. Proc. Natl. Acad. Sci. USA 2022, 119, e2201711119. [Google Scholar] [CrossRef]
- Cregg, J.M.; Madden, K.R.; Barringer, K.J.; Thill, G.P.; Stillman, C.A. Functional characterization of the two alcohol oxidase genes from the yeast Pichia pastoris. Mol. Cell. Biol. 1989, 9, 1316–1323. [Google Scholar]
- Potvin, G.; Ahmad, A.; Zhang, Z.S. Bioprocess engineering aspects of heterologous protein production in Pichia pastoris: A review. Biochem. Eng. J. 2012, 64, 91–105. [Google Scholar] [CrossRef]
- Kim, T.Y.; Sohn, S.B.; Kim, Y.B.; Kim, W.J.; Lee, S.Y. Recent advances in reconstruction and applications of genome-scale metabolic models. Curr. Opin. Biotechnol. 2012, 23, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Dikicioglu, D.; Kırdar, B.; Oliver, S.G. Biomass composition: The “elephant in the room” of metabolic modelling. Metabolomics 2015, 11, 1690–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vartiainen, E.; Blomberg, P.; Ilmen, M.; Andberg, M.; Toivari, M.; Penttila, M. Evaluation of synthetic formaldehyde and methanol assimilation pathways in Yarrowia lipolytica. Fungal Biol. Biotechnol. 2019, 6, 27. [Google Scholar] [CrossRef]
- Zepeda, A.B.; Pessoa, A.; Farias, J.G. Carbon metabolism influenced for promoters and temperature used in the heterologous protein production using Pichia pastoris yeast. Braz. J. Microbiol. 2018, 49, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Zhan, C.; Li, X.; Baidoo, E.E.K.; Yang, Y.; Sun, Y.; Wang, S.; Wang, Y.; Wang, G.; Nielsen, J.; Keasling, J.D.; et al. The Glyoxylate-serine pathway enables conversion of Saccharomyces cerevisiae to a synthetic methylotroph. Cell Press Sneak Peek 2021, 2021. [Google Scholar] [CrossRef]
- Rußmayer, H.; Buchetics, M.; Gruber, C.; Valli, M.; Grillitsch, K.; Modarres, G.; Guerrasio, R.; Klavins, K.; Neubauer, S.; Drexler, H.; et al. Systems-level organization of yeast methylotrophic lifestyle. BMC Biol. 2015, 13, 80. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.F.; Yang, J.; Zhao, F.; Lin, Y.; Han, S. Comparative transcriptome and metabolome analyses reveal the methanol dissimilation pathway of Pichia pastoris. BMC Genom. 2022, 23, 366. [Google Scholar] [CrossRef]
- Magasanik, B.; Kaiser, C.A. Nitrogen regulation in Saccharomyces cerevisiae. Gene 2002, 290, 1–18. [Google Scholar] [CrossRef]
- Grenson, M.; Dubois, E.; Piotrowska, M.; Drillien, R.; Aigle, M. Ammonia assimilation in Saccharomyces cerevisiae as mediated by the two glutamate dehydrogenases. Evidence for the gdhA locus being a structural gene for the NADP-dependent glutamate dehydrogenase. Mol. Genet. Genom. 1974, 128, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.P.; Magasanik, B. Purification and properties of glutamine synthetase from Saccharomyces cerevisiae. J. Biol. Chem. 1983, 258, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.P. The GLN1 locus of Saccharomyces cerevisiae encodes glutamine synthetase. Genetics 1985, 111, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Du, G.; Zhou, J.; Chen, J. Regulation of sensing. transportation, and catabolism of nitrogen sources in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2018, 82, e00040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Wang, Z.; Wang, Z.; Dou, J.; Zhou, C. Improving methionine and ATP availability by MET6 and SAM2 co-expression combined with sodium citrate feeding enhanced SAM accumulation in Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2016, 32, 56. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.H.; Liu, N.; Zhu, Y.X.; Yu, S.Y.; Liu, Q.L.; Shi, X.L.; Xu, J.G.; Xu, G.Q.; Zhang, X.M.; Shi, J.S.; et al. Improving glutathione production by engineered Pichia pastoris: Strain construction and optimal precursor feeding. Appl. Microbiol. Biotechnol. 2022, 106, 1905–1917. [Google Scholar] [CrossRef] [PubMed]
- Pfeifenschneider, J.; Brautaset, T.; Wendisch, V.F. Methanol as carbon substrate in the bio-economy: Metabolic engineering of aerobic methylotrophic bacteria for production of value-added chemicals. Biofuels Bioprod. Biorefining 2017, 11, 719–731. [Google Scholar] [CrossRef]
- Zhu, T.C.; Zhao, T.X.; Bankefa, O.E.; Li, Y. Engineering unnatural methylotrophic cell factories for methanol-based biomanufacturing: Challenges and opportunities. Biotechnol. Adv. 2020, 39, 107467. [Google Scholar] [CrossRef]
- Hartadi, Y.; Widmann, D.; Behm, R.J. Methanol synthesis via CO2 hydrogenation over a Au/ZnO catalyst: An isotope labelling study on the role of CO in the reaction process. Phys. Chem. Chem. Phys. 2016, 18, 10781–10791. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.K.; Liao, Y.; Wang, J.; Ma, C.; Qin, J.L.; Feng, J.; Li, Y.; Wang, X.; Chen, K.Q. Methylotrophy of Pichia pastoris: Current advances, applications, and future perspectives for methanol-based biomanufacturing. ACS Sustain. Chem. Eng. 2022, 10, 1741–1752. [Google Scholar] [CrossRef]
- Wang, X.L.; Wang, Q.; Wang, J.J.; Bai, P.; Shi, L.; Shen, W.; Zhou, M.; Zhou, X.S.; Zhang, Y.X.; Cai, M.H. Mit1 transcription factor mediates methanol signaling and regulates the alcohol oxidase 1 (AOX1) promoter in Pichia pastoris. J. Biol. Chem. 2016, 291, 6245–6261. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains and Plasmids | Relevant Characteristics | Sources |
|---|---|---|
| P. pastoris | ||
| PC110 | Derived from GS115, PGAP-PpRAD52-TAOX1 | [28] |
| PC110-AOX1-464 | Derived from PC110, AOX1∷R464 to K464 | This study |
| PC110-ΔAOX2 | Derived from PC110, ΔAOX2 | This study |
| E. coli | ||
| DH5a | F−, φ80lacZ ΔM15, Δ(lacZYA-argF)U169, deoR, recA1, endA1, hsdR17(rk−, mk+), phoA, supE44, λ−, thi-1, gyrA96, relA1 | Lab stock |
| Plasmids | ||
| pPICZ-Cas9-gGUT1 | ori, Amp, Zeocin, TDAS1-Cas9-PHTX1-GUT1-gRNA2-TAOX1 | [28] |
| pPICZ-Cas9-gAOX1 | ori, Amp, Zeocin, TDAS1-Cas9-PHTX1-AOX1-gRNA-TAOX1 | This study |
| pPICZ-Cas9-gAOX2 | ori, Amp, Zeocin, TDAS1-Cas9-PHTX1-AOX2-gRNA-TAOX1 | This study |
| Primer | Sequences (5′→3′) |
|---|---|
| gAOX1-1F | AACAGACTGATGAGTCCGTGAGGACGAAACGAGTAAGCTCGTCTCTGTTCCCATACTCATCCGGTTTTAGAGCTAGAAATAGCA |
| gAOX1-1R | ACGGGAAGTCTTTACAGTTT |
| gAOX1-2F | CTCCTAACTAAAACTGTAAAGACTTCCCGTTTAAACTTTTCTTTTCTTCT |
| gAOX1-2R | GTTTCGTCCTCACGGACTCATCAGTCTGTTTTTGATTTGTTTAGGTAACT |
| AOX1-464-Up-F | ACTTGCCAGGTGTCGGAAGA |
| AOX1-464-Up-R | TTTAGACTTCTTGTAAGCCC |
| AOX1-464-Down-F | CCTATGGTTTGGGCTTACAAGAAGTCTAAAGAAACCGCTAGAAGAATGGA |
| AOX1-464-Down-R | TTAGAATCTAGCAAGACCGGTC |
| AOX1-Up-F | GAGTCTTTCGATGACTTCGT |
| AOX1-Up-R | TTCGGATGAGTATGGGAACAGA |
| AOX1-Down-F | CACCACCCTCTGTTCCCATACTCATCCGAAGCCAGAGCCTTGGAAATGGA |
| AOX1-Down-R | CTTGAACTGAGGAACAGTCA |
| gAOX2-1F | CAGAGGCTGATGAGTCCGTGAGGACGAAACGAGTAAGCTCGTCCCTCTGGTTCTGCAAAGATCGTTTTAGAGCTAGAAATAGCA |
| gAOX2-1R | ACGGGAAGTCTTTACAGTTT |
| gAOX2-2F | CTCCTAACTAAAACTGTAAAGACTTCCCGTTTAAACTTTTCTTTTCTTCT |
| gAOX2-2R | GTTTCGTCCTCACGGACTCATCAGCCTCTGTTTGATTTGTTTAGGTAACT |
| AOX2-Up-F | TTAAGCGAAAGAGACAAGACAACGA |
| AOX2-Up-R | TTTTCTCAGTTGATTTGTTTGTGGG |
| AOX2-Down-F | AAATCCCCACAAACAAATCAACTGAGAAAATTTATGTTGTATCTATGAATATTTT |
| AOX2-Down-R | TTAGACTACTCTGAATCCGAGAAGA |
| Parameter | PC110 | PC110-AOX1-464 | PC110-ΔAOX2 | |
|---|---|---|---|---|
| Input | Methanol utilization rate (mmol/gDcw/h) | 10 | 6.6 | 9.3 |
| Output | CO2 (mmol CO2/mmol methanol) | 6.83 | 4.98 | 6.45 |
| Biomass (%) | 2.56 | 2.62 | 2.57 |
| Strains | DCW (g) | Methanol Utilization Rate (g DCW/g) |
|---|---|---|
| PC110 | 0.126 ± 0.002 | 0.318 ± 0.005 |
| PC110-AOX1-464 | 0.144 ± 0.001 | 0.364 ± 0.003 |
| PC110-ΔAOX2 | 0.128 ± 0.002 | 0.324 ± 0.006 |
| PC110-AOX1-464 Strain with Addition of Different Concentrations of Sodium Citrate (g/L) | DCW (g) | Methanol Utilization Rate (g DCW/g) |
|---|---|---|
| 0 | 0.146 ± 0.005 | 0.368 ± 0.007 |
| 2 | 0.167 ± 0.003 | 0.421 ± 0.007 |
| 4 | 0.170 ± 0.004 | 0.430 ± 0.007 |
| 6 | 0.175 ± 0.002 | 0.442 ± 0.006 |
| 8 | 0.167 ± 0.003 | 0.420 ± 0.008 |
| 10 | 0.155 ± 0.001 | 0.390 ± 0.003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Dong, H.; Hong, K.; Meng, J.; Lin, L.; Wu, X. Improving Methanol Utilization by Reducing Alcohol Oxidase Activity and Adding Co-Substrate of Sodium Citrate in Pichia pastoris. J. Fungi 2023, 9, 422. https://doi.org/10.3390/jof9040422
Liu S, Dong H, Hong K, Meng J, Lin L, Wu X. Improving Methanol Utilization by Reducing Alcohol Oxidase Activity and Adding Co-Substrate of Sodium Citrate in Pichia pastoris. Journal of Fungi. 2023; 9(4):422. https://doi.org/10.3390/jof9040422
Chicago/Turabian StyleLiu, Shufan, Haofan Dong, Kai Hong, Jiao Meng, Liangcai Lin, and Xin Wu. 2023. "Improving Methanol Utilization by Reducing Alcohol Oxidase Activity and Adding Co-Substrate of Sodium Citrate in Pichia pastoris" Journal of Fungi 9, no. 4: 422. https://doi.org/10.3390/jof9040422