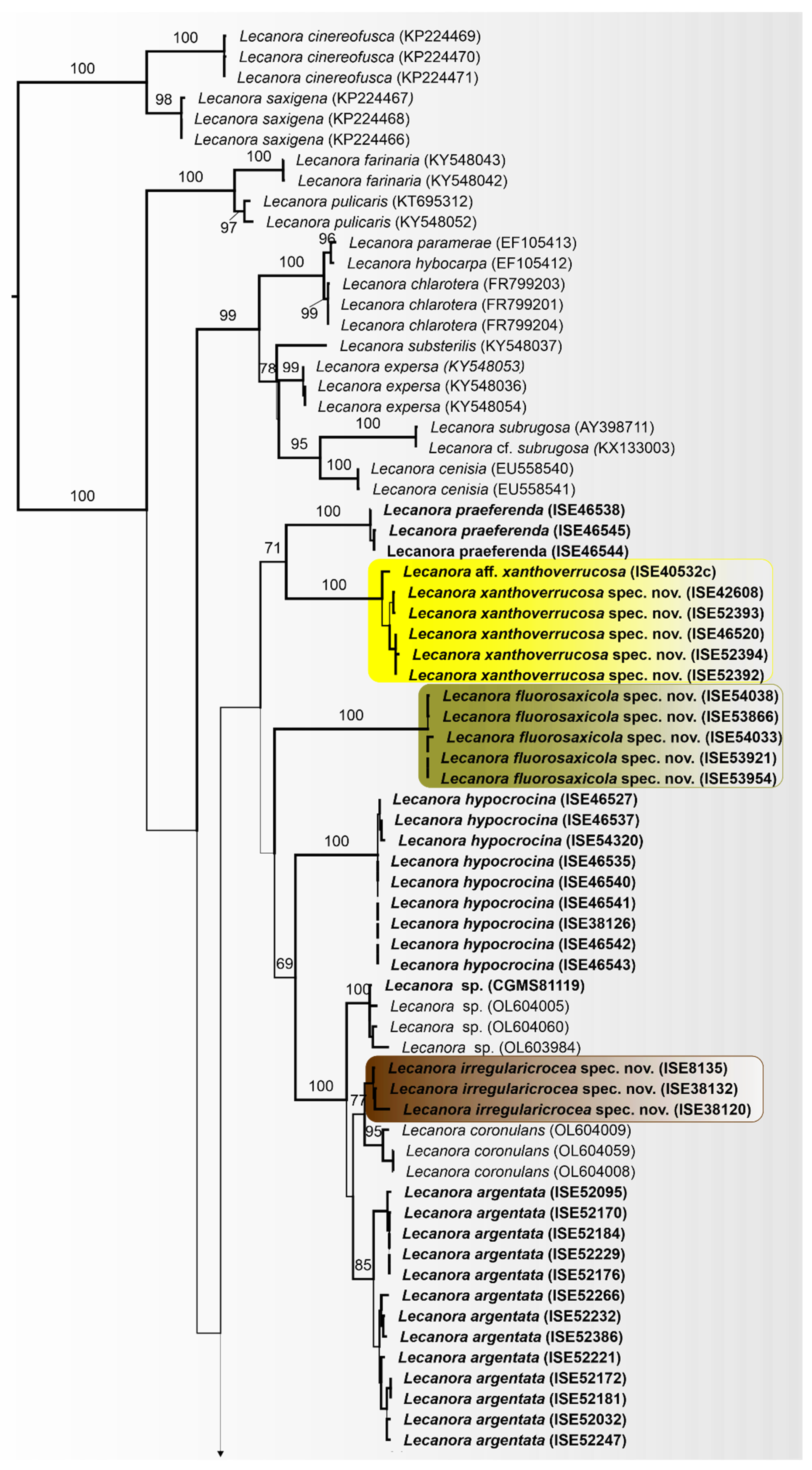
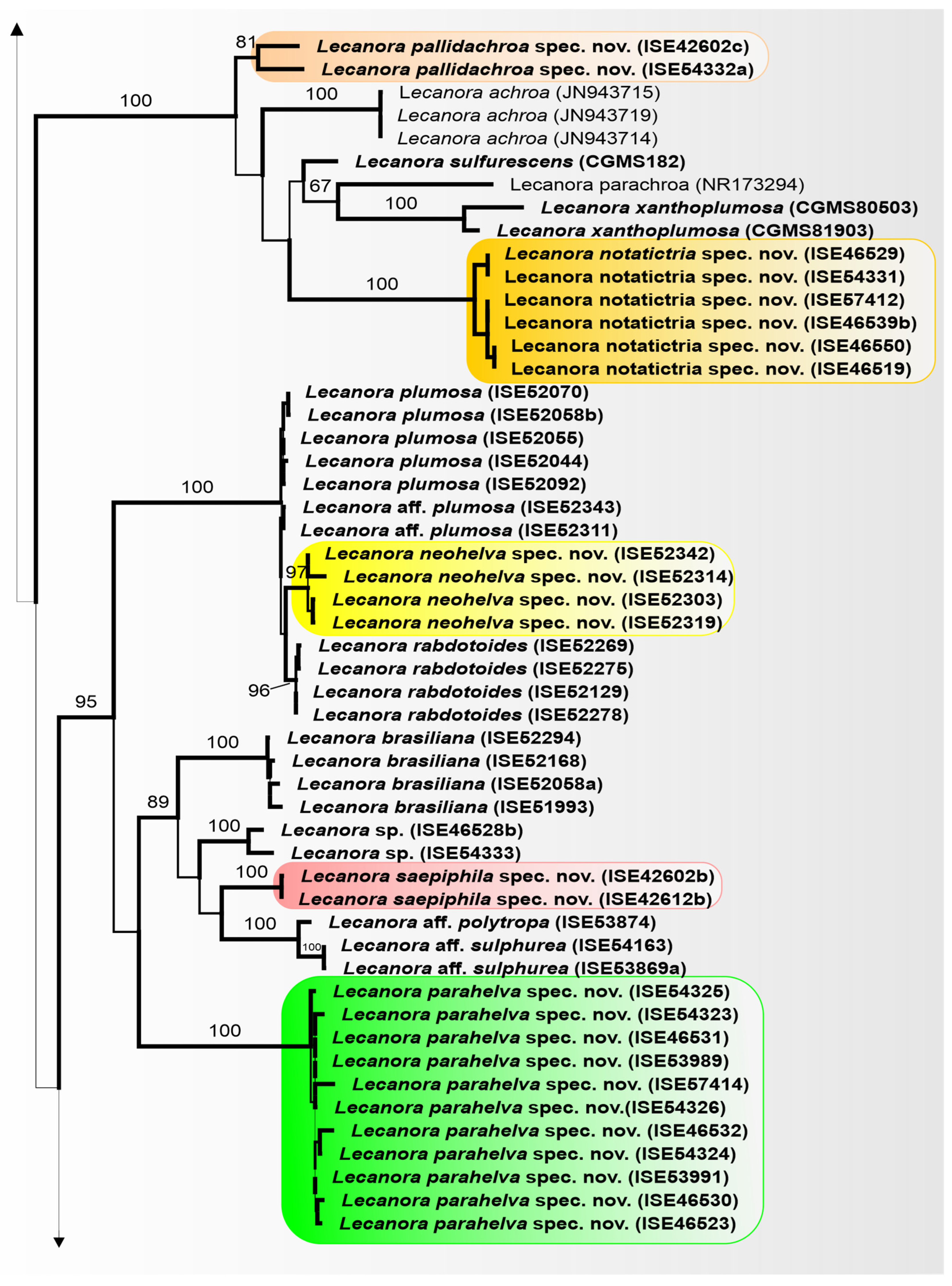
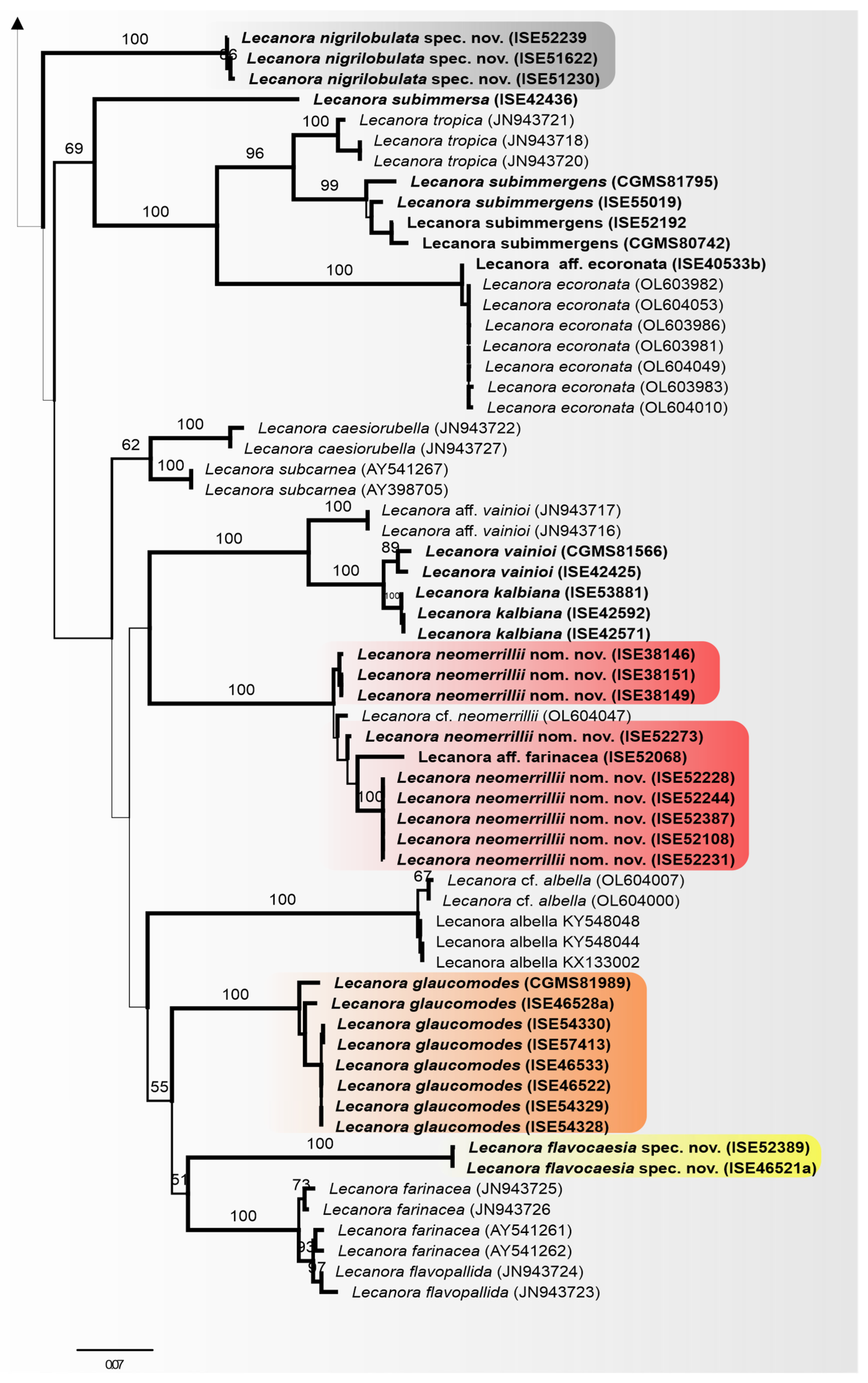
3.3. Taxonomy
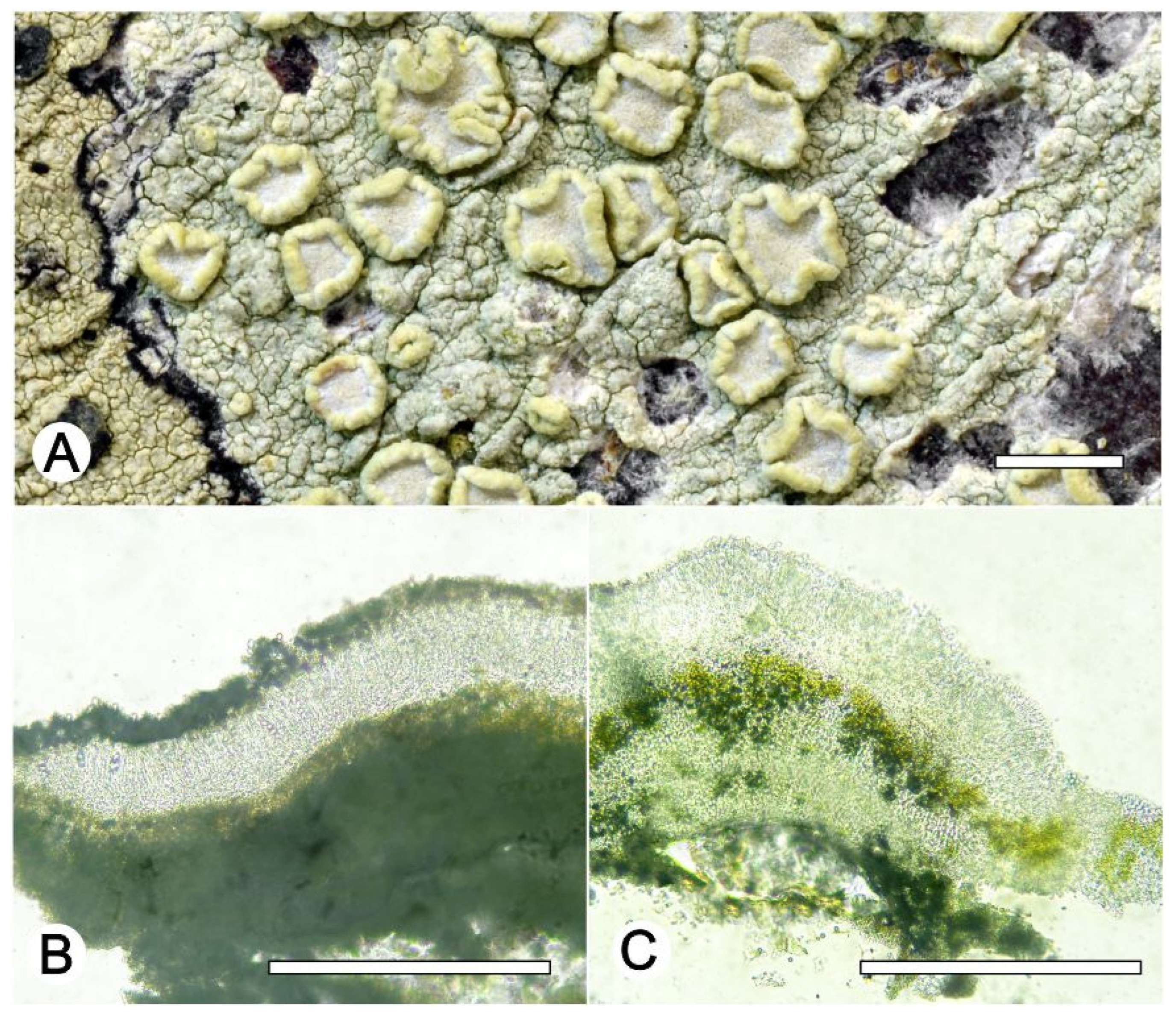
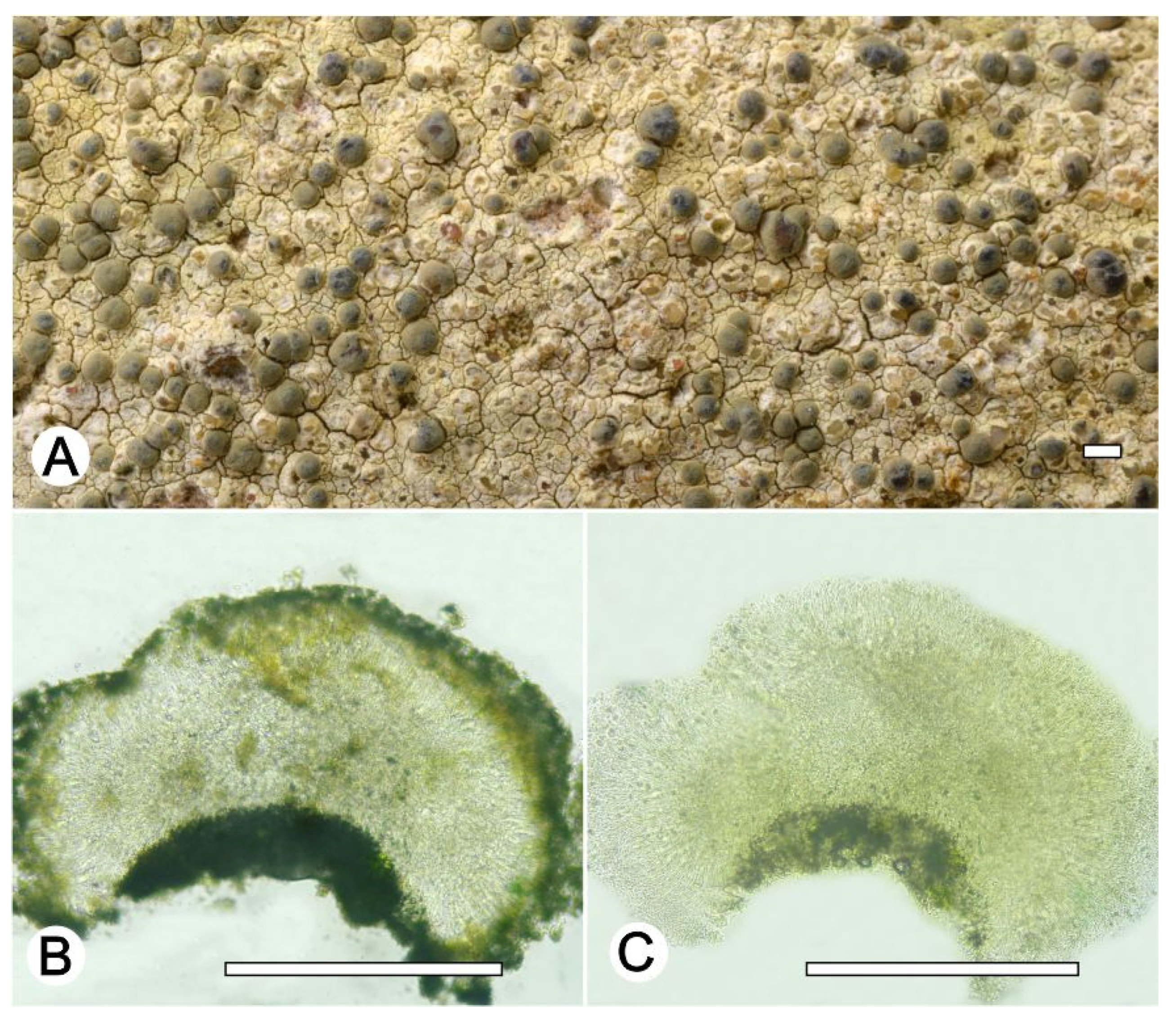
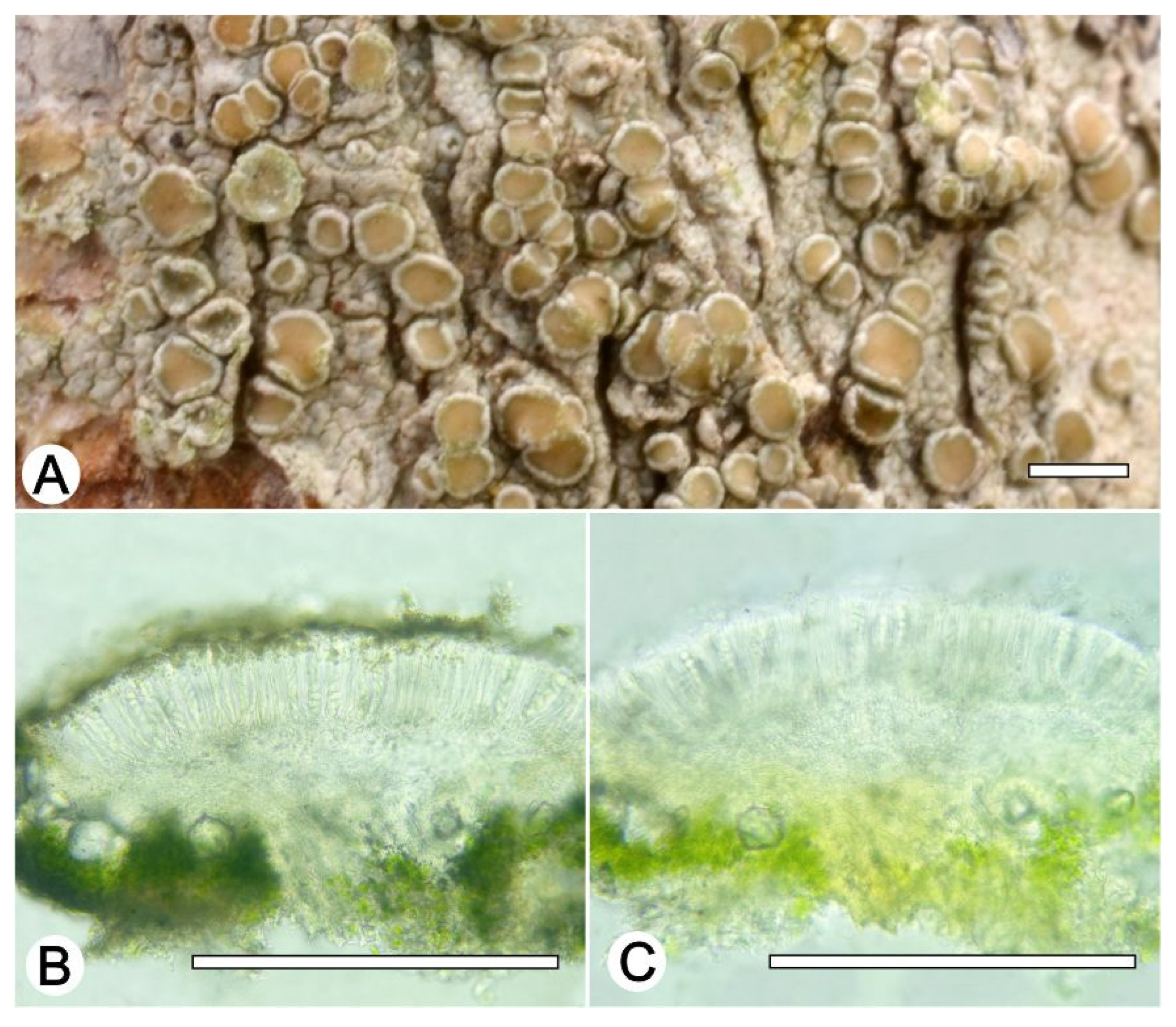
Lecanora flavocaesia L.A. Santos, M. Cáceres, Aptroot, and Lücking, sp. nov. (
Figure 2).
MycoBank MB 847936.
Etymology: The epithet refers to the colour of apothecial margin.
Typus: BRAZIL, Pernambuco, Parque Nacional Vale do Catimbau, Buíque, 8°30′32.6″ S, 8°30′32.6″ S, on bark of unidentified tree, 13–15 August 2017, L.A. Santos s.n. (ISE-46521a, holotype; ITS Genbank No.OQ650143).
Diagnosis: Similar to Lecanora caesiorubella Ach. but differing by the distinctly yellowish, very irregular margins, and much smaller apothecia.
Description: Thallus crustose, corticolous, up to 2.5 cm wide, c. 0.1 mm thick, areolate, rimose to verruculose, whitish yellow, dull, epruinose. Soredia absent. Prothallus absent or whitish grey. Apothecia adnate, 0.8–1.5 mm diam.; disc pale orange to whitish brown, with a thin, whitish grey pruina, flat to convex; margin pale yellow, prominent, becoming excluded or persisting, strongly flexuose. Amphithecium with small crystals, small crystals rapidly dissolving in KOH (=allophana-type), parathecium with small crystals. Hymenium hyaline, 37.5–50 µm high; epihymenium brown to dark brown, granular with small crystals, rapidly dissolving in KOH (=chlarotera-type), c. 15 µm high. Hypothecium hyaline. Ascospores 8 per ascus, ellipsoid, (8–) 10–12(–13) × 5–7 μm. Pycnidia not observed.
Chemistry: Thallus and apothecia margin K+ yellow, C–, P+ orange, UV-. TLC: atranorin, protocentratic acid.
Habitat and distribution: On exposed trees in Caatinga forest; known only from Brazil.
Additional sequenced specimens examined: BRAZIL, Sergipe, Monumento Natural Grota do Angico, Poço Redondo; 9°39′39.7″ S, 37°41′09.7″ W, on bark of unidentified tree, 24 November 2016, L.A. Santos s.n. (ISE-52389)
Notes: This new species shares the chemistry of atranorin and protocetraric acid with Lecanora caesiorubella subsp. glaucomodes (Nyl.) Imshaug and Brodo but differs in the pale yellow and very irregular apothecial margins and the areolate thallus. We therefore consider the morphological differences in the apothecial margins and the thallus diagnostic.
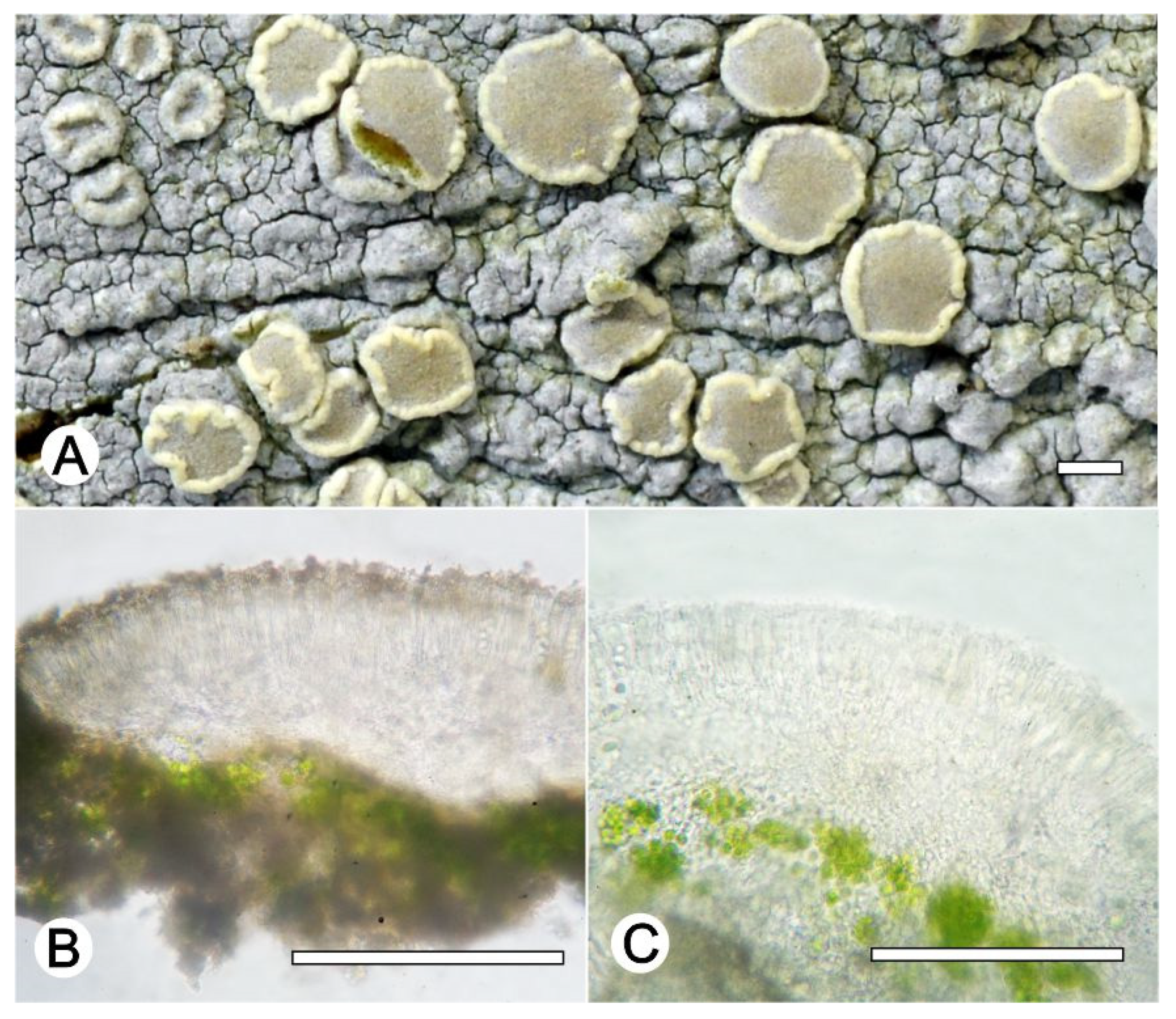
Lecanora fluorosaxicola L.A. Santos, M. Cáceres, and Aptroot, sp. nov. (
Figure 3).
MycoBank MB 847937.
Etymology: The epithet conveys the fluorescent yellow greenish colour of the thallus.
Typus: BRAZIL, Bahia, Estação Ecológica Raso da Catarina, Paulo Afonso, 9°39′58.8″ S, 38°28′05.2″ W, on rock, 10–12 October 2021, L.A. Santos s.n. (ISE-54033, holotype; ITS Genbank No.OQ650054).
Diagnosis: The new species can be distinguished from the other species of Lecanora by the fluorescent yellow greenish colour of the thallus.
Description: Thallus crustose, saxicolous, up to 7 cm wide, c. 0.1 mm thick, areolate, rimose to verruculose, yellowish green, dull, epruinose. Soredia absent. Prothallus absent or black. Apothecia adnate, 0.2–1 mm diam.; disc dark grey to greyish black, with a thick, greenish yellow pruina, flat when young to strongly concave; margin concolourous with the thallus, becoming excluded, smooth. Amphithecium with small crystals which are dissolving in KOH (=melacarpella-type), parathecium with small crystals. Hymenium hyaline, 42.5–62.5 µm high; epihymenium green to yellowish-green, the pigment and with small crystals soluble in KOH (= chlarotera-type), c. 12 µm high. Hypothecium hyaline. Ascospores 8 per ascus, ellipsoid, (9–) 10–12(–13) × 5–7.5 μm. Pycnidia not observed.
Chemistry: Thallus and apothecia margin K+ yellow, C+ yellow, P–, Thallus and apothecial disc UV+ orange. TLC: atranorin.
Habitat and distribution: On exposed rock in Caatinga forest; known only from Brazil.
Additional sequenced specimens examined: BRAZIL, Bahia, Estação Ecológica Raso da Catarina, Paulo Afonso, 9°39′58.8″ S, 38°28′05.2″ W, on rock, 10–12 October 2021, L.A. Santos s.n. (ISE-53866, ISE-53954, ISE-53921, ISE-54038).
Notes:
Lecanora fluorosaxicola is most likely to be confused with
L. vainioi Vänskä and
L. kalbiana Lumbsch in the more or less brown to grey-brown apothecial disc and the substrate, as both are found on rock. The two species were described from Brazil:
L. kalbiana was from Mato Grosso, currently only known from Brazil, and
L. vainioi from Rio de Janeiro, restricted to the Neotropics. The new species differs in the thallus being mostly yellow, the apothecial discs with a yellowish green pruina, the hyaline hypothecium, and in the absence of zeorin.
Lecanora ryanii T.H. Nash and Lumbsch is also superficially similar, but differs in the larger, sessile apothecia with epruinose disc; it is only known from Isla Margarita in México [
42].
The differences in chemistry and morphology are reflected in the molecular phylogeny (
Figure 1), where
L. fluorosaxicola represents a single taxon on a long branch in a subclade with absolute support, whereas
L. vainioi,
L. kalbiana and two sequences from GenBank (Thailand) labeled
L. vainioi, form another sub-clade. Both subclades fell in a clade with pruinose species (
Figure 1).
Synonym: Lecanora caesiorubella subsp. glaucomodes (Nyl.) Imshaug and Brodo, Nova Hedwigia 12(1+2): 15 (1966)
Typus: Cuba, C. Wright Lich. Cub. II. 60 (H-NYL 27145, holotype)
Description specimens examined: Thallus crustose, corticolous, up to 3.5 cm wide, c. 0.1 mm thick, areolate, rimose to verruculose, whitish grey to grey, dull, epruinose. Soredia absent. Prothallus absent or whitish grey. Apothecia adnate, 0.5–1.5 mm diam.; disc pale orange to brownish orange, carneous, whitish grey pruina, flat to convex; margin slightly paler than the thallus, prominent, becoming excluded or persisting, entire to flexuose, smooth. Amphithecium with small crystals which are dissolving in KOH (= allophana-type), parathecium with small crystals. Hymenium hyaline, 50–75 µm high; epihymenium brown to dark brown with pigment and small crystals dissolving in KOH (= chlarotera-type), c. 12 µm high. Hypothecium hyaline. Ascospores 8 per ascus, ellipsoid, (10–) 10–13(–14) × 6–8 μm. Pycnidia not observed.
Chemistry: Thallus and apothecia margin K+ yellow, C–, P+ orange to red, UV–. TLC: atranorin, protocetraric acid.
Sequenced specimens examined: Brazil, Pernambuco, Parque Nacional Vale do Catimbau, Buíque, 8°31′30.6″ S, 37°14′58.8″ W, on bark of unidentified tree, 13–15 August 2017, L.A. Santos s.n. (ISE–46528a, 54328, 54329); Sergipe, Monumento Natural Grota do Angico, Poço Redondo, 9°39′39.7″ S, 37°41′09.7″ W, on bark of unidentified tree, 24 November 2016, L.A. Santos s.n. (ISE–46522, 46533, 54330, 57413); Mato Grosso, Chapada dos Guimarães, Pousada do Parque private area, 15°26′50″ S, 55°49′50″ W, “on bark of unidentified tree, 12–18 September 2020, A. Aptroot and M.F. Souza 81,989 (CGMS).
Notes: This pruinose species is chemically characterized by the presence of protocetraric acid and atranorin. Given its phylogeny, it is here recognized at species level as
Lecanora glaucomodes. If we consider all chemical variation recently found (see Brodo et al. [
43]), this material agrees partially with
L. caesiorubella subsp.
glaucomodes. However, the chemical definition of
L. glaucomodes is complex. Imshaug and Brodo [
44] reported atranorin and protocetraric acid as a major compound, which was found in the isotype. Later, Lumbsch et al. [
45] reported that the holotype of this species produces virensic acid as a major substance. In a recent review of the
L. albella group, it was demonstrated that the definition of species or subspecies just observing the chemistry remains complex [
43]. However, the new sequences generated in the present work form a clade distinct from Genbank sequences identified as
L. caesiorubella s.str., from the USA and Australia (
Figure 1).
Lecanora irregularicrocea L.A. Santos, M. Cáceres, Aptroot, and Lücking sp. nov. (
Figure 5).
MycoBank MB 847938.
Etymology: The name refers to the strongly flexuose apothecial margins and to differ the new species of the coronulans group.
Typus: BRAZIL, Rio de Janeiro, Parque Nacional de Itatiaia, Itatiaia, 22°33′ S, 42°19′ W, on bark of unidentified tree, 11–14 October 2016, M.E.S. Cáceres s.n. (ISE-38135, holotype; ITS Genbank No.OQ650060).
Diagnosis: The new species can be distinguished from the somewhat similar species
Lecanora coronulans Nyl. by the strongly irregularly and flexuose apothecium margin and larger ascospores.
Description: Thallus crustose, corticolous, up to 3 cm wide, c. 0.1 mm thick, areolate, rimose to verruculose, white yellowish to whitish grey, dull, epruinose. Soredia absent. Prothallus absent or whitish grey. Apothecia adnate to sessile, 0.2–1.8 mm diam.; disc brown to dark brown, epruinose, flat to convex; margin slightly paler than the thallus, prominent, flexuose, smooth, entire, sometimes strongly flexuose, whitish to whitish grey. Amphithecium with small and large crystals (= melacarpella-type), parathecium with small crystals. Hymenium hyaline, 50–62.5 µm high; epihymenium dark red-brown to dark brown without crystals, pigmentation not altered by KOH (= glabrata-type), c. 12 µm high. Hypothecium reddish brown. Ascospores 8 per ascus, broad ellipsoid, (12–) 13–17(–18) × 7–10 μm. Pycnidia not observed.
Chemistry: Thallus and apothecia margin K+ yellow, C–, P–, UV–. TLC: atranorin, zeorin.
Habitat and distribution: on bark of tree in Itatiaia rain forest; known only from Brazil.
Additional sequenced specimens examined: BRAZIL, Rio de Janeiro, Parque Nacional de Itatiaia, Itatiaia, 22°33′ S, 42°19′ W, on bark of unidentified tree, 11–14 October 2016, M.E.S. Cáceres s.n. (ISE-38120, ISE-38132).
Notes:
Lecanora irregularicrocea represents a hitherto undescribed member of the
L. coronulans group. All species of this group have a pigmented hypothecium and occur exclusively in the tropics [
46]. This new species is similar to an assemblage of taxa that occurs in Brazil:
Lecanora concilianda Vain.,
L. concilians Nyl.,
L. coronulans Nyl.,
L. hypocrocina Nyl. All agree in the more or less brown to dark brown apothecial disc and the
melacarpella-type amphithecium.
Lecanora concilianda was described from Brazil;
L. concilians originates from Colombia, whereas
L. coronulans and
L. hypocrocina were described from Cuba.
The differences between the five species are mainly morphological and in part chemical: Lecanora concilianda and L. concilians differ in spore size and thallus morphology; L. coronulans can be distinguished by its smaller apothecia and the presence of zeorin, and L. hypocrocina is different from all other species by the KOH+ red to purple reaction of the hypothecium (boryquinone). The new species somewhat resembles L. coronulans in the presence of zeorin, but differs in the strongly irregularly and flexuose apothecium margin and larger ascospores (13–17 × 7–10 μm vs. 9.5–14 × 5–8 μm).
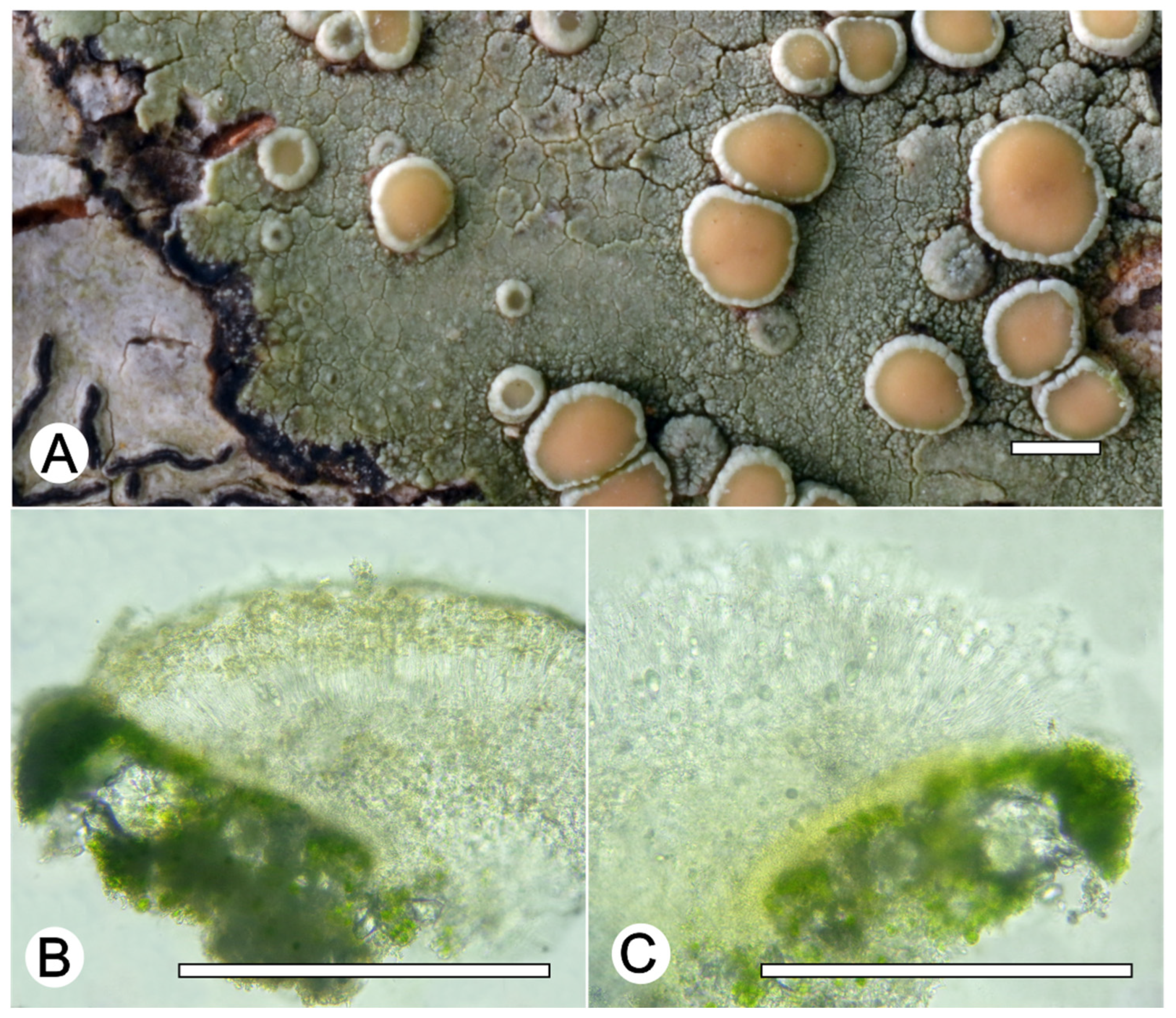
Lecanora neohelva L.A. Santos, M. Cáceres and Aptroot, sp. nov. (
Figure 6)
M MycoBank MB 847939.
Etymology: The epithet is taken from the Latin neo-, meaning no, and helva, referring similar species Lecanora helva.
Typus: BRAZIL, Minas Gerais, Reserva Particular do Patrimônio Natural Santuário do Caraça, Catas Altas, 7°2′1″ S, 36°3′1″ W, on bark of unidentified tree, 17–25 May 2021, L.A. Santos and A. Aptroot (ISE-52342, holotype; CGMS, isotype; ITS Genbank No. OQ650118)
Diagnosis: The new species can be distinguished by the very green thallus, pale orange apothecial discs, larger apothecia and larger ascospores than in the Lecanora helva. The phylogenetic tree also indicate it forms an independent clade.
Description: Thallus crustose, corticolous, up to 2.5 cm wide, c. 0.1 mm thick, continuous, rimose, green to whitish green, dull, epruinose. Soredia absent. Prothallus absent or greyish black. Apothecia adnate, 0.5–1.5 mm diam.; disc pale orange to yellowish orange, epruinose, flat to convex; margin slightly paler than the thallus, prominent, becoming excluded or persisting, smooth to verruculose. Amphithecium with large and small crystals (=melacarpella-type), parathecium with small crystals. Hymenium hyaline, 50–75 µm high; epihymenium pale yellow to yellowish brown, with small crystals and pigment soluble in KOH (=chlarotera-type), c. 10 µm high. Hypothecium hyaline to pale yellowish (intensifying in KOH). Ascospores 8 per ascus, ellipsoid, (10–) 12–15 × 5–7 μm. Pycnidia not observed.
Chemistry: Thallus and apothecia margin K+ yellow, C–, P–, thallus UV+ pink, apothecia disc UV+ green. TLC: atranorin, ‘2-O-methylperlatolic.
Habitat and distribution: On exposed trees in campo rupestre; known only from Brazil.
Additional sequenced specimens examined: BRAZIL: Minas Gerais, Reserva Particular do Patrimônio Natural Santuário do Caraça, Catas Altas, 7°2′1″ S, 36°3′1″ W, on bark of unidentified tree, 17–25 May 2021, L.A. Santos and A. Aptroot (ISE-52303, ISE-52314, ISE-52319, CGMS);
Notes: This species can be recognized by the pale orange apothecial discs, the chlarotera-type epihymenium, and the greenish thallus. Lecanora helva Stizenb. is morphologically similar but can be distinguished by the smaller apothecia (0.4–1.0 mm diam. vs. 0.5–1.5 mm diam.) and smaller ascospores (5–7 × 10–13.5 μm vs. 12–15 × 5–7 μm). Lecanora stramineoalbida Vain. is another similar species, which differs in having a reddish brown hypothecium.
Lecanora neomerrillii L.A. Santos, Aptroot, M. Cáceres and Lücking, nom. nov. (
Figure 7)
MycoBank MB 847940.
Synonym: Lecanora caesiorubella Ach. subsp. merrillii Imshaug and Brodo, Nova Hedwigia 12: 28. 1966. Type species—U.S.A. CALIFORNIA: Berkeley, 1893, M.A. Howe [=Cummings and Seymour, Decades of North American Lichens I, 133] (MSC, holotype).
Description specimens examined: Thallus crustose, corticolous, up to 7 cm wide, c. 0.1 mm thick, continuous, rimose to verruculose, whitish grey, dull, epruinose. Soredia absent. Prothallus absent or greyish black. Apothecia adnate, 0.5–2 mm diam.; disc pale yellow to brownish orange, carneous, whitish grey pruina, flat to convex; margin slightly paler than the thallus, prominent, becoming excluded or persisting, crenate. Amphithecium with small crystals which are dissolving in KOH (=allophana-type), parathecium with small crystals. Hymenium hyaline, 50–62.5 µm high; brown to dark brown with pigment and small crystals dissolving in KOH (=chlarotera-type), c. 12 µm high. Hypothecium hyaline. Ascospores 8 per ascus, ellipsoid, (14–) 15–18 × 8–12 μm. Pycnidia not observed.
Chemistry: Thallus K+ yellow, C–, P+ orange to red, UV–. TLC, Chemotype I: atranorin and norstictic acid. Chemotype II: atranorin and protocetraric acid.
Sequenced specimens examined: Chemotype I: BRAZIL, Minas Gerais, Reserva Particular do Patrimônio Natural Santuário do Caraça, Catas Altas, 7°2′1″ S, 36°3′1″ W, on bark of unidentified tree, 17–25 May 2021, L.A. Santos s.n. (ise–, cgms–, 52228, 52387, 52244, 52231, 52108); Chemotype II: Minas Gerais, Reserva Particular do Patrimônio Natural Santuário do Caraça, Catas Altas, 7°2′1″ S, 36°3′1″ W, on bark of unidentified tree, 17–25 May 2021, L.A. Santos s.n. (ise–, cgms– 52273); Rio de Janeiro, Parque Nacional de Itatiaia, Itatiaia, Est. Agulhas Negras, 22°22′31″ S, 44°39′44″ O, on bark of undertermined tree, 12–13 October 2016, Cáceres s.n. (ISE–38146, ISE–38149, ISE–38151).
Notes: This taxon of the
Lecanora cesiorubella morphotype with pruinose apothecia can be separated into two chemotypes. Chemotype I, characterized by the presence of norstictic acid and atranorin; and Chemotype II, with atranorin, protocetraric acid, and undefined fatty acids, but lacking norstictic acid. Additionally, phylogenetically (
Figure 1), the sequences generated for this material clustered into two distinct subclades, one with specimens that produce norstictic acid and atranorin, which agree with subsp.
merrillii [
43,
44,
45]. The other produces atranorin, protocetraric acid, undefined fatty acids (except the specimen ISE-52273) and lacks norstictic acid. Based on the chemistry, part of our material agrees with the
L. albella (Chemotype I). However,
Lecanora albella has smaller apothecia and ascospores [
45], additionally, the new sequences generated in the present work form a clade distinct of Genbank sequences identified as
L. albella (
Figure 1). It is therefore recognized at species level. Therefore, we proposed the new name
Lecanora neomerrilli, because there is already
Lecanora merrillii Nyl, as, e.g., mentioned by Brodo et al. [
43].
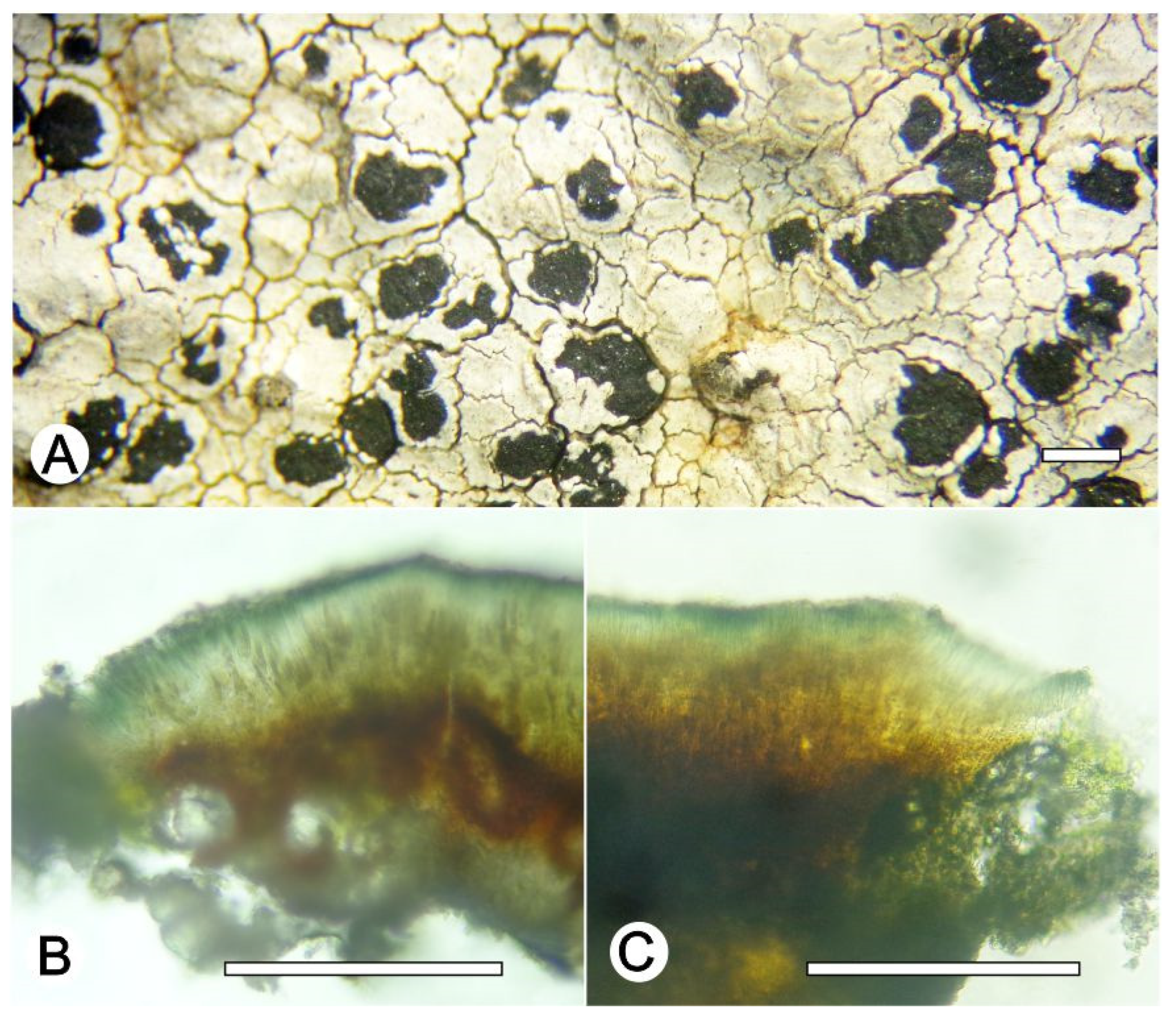
Lecanora nigrilobulata L.A. Santos, M. Cáceres and Aptroot, sp. nov. (
Figure 8)
MycoBank MB 847941.
Etymology: named after the immersed, very irregular apothecia that somewhat resembles black paint spots.
Typus: BRAZIL, Minas Gerais, Reserva Particular do Patrimônio Natural Santuário do Caraça, Catas Altas, 7°2′1″ S, 36°3′1″ W, on sandstone, 17–25 May 2021, L.A. Santos and A. Aptroot (ISE-52239, holotype; CGMS, isotype; ITS Genbank No.OQ650091).
Diagnosis: Lecanora nigrilobulata is similar to L. subimmersa (Fée) Vain. and L. oreinoides (Körb.) Hertel and Rambold in the immersed apothecia, areolated thallus, but can be distinguished by the hypothecium colour.
Description: Thallus crustose, saxicolous, up to 3 cm wide, c. 0.1 mm thick, areolate, puzzlle shaped areoles, whitish grey, dull, epruinose. Soredia absent. Prothallus absent or black. Apothecia immersed to adnate, 0.8–1.8 mm diam.; disc black, epruinose, flat; margin concolourous with the thallus, prominent, becoming excluded or persisting, areolate. Amphithecium with large crystals which are not dissolving in KOH (= pulicaris-type), parathecium with small crystals. Hymenium dark brown, 50–75 µm high; epihymenium black to greenish black, the pigment turning green in KOH, without crystals (gangaleoides-type), c. 15 µm high. Hypothecium dark brown. Ascospores 8 per ascus, ellipsoid, (8–) 10–13(–15) × 5–7.5 μm. Pycnidia not observed.
Chemistry: Thallus and apothecia margin K+ yellow, C–, P–, UV–. TLC: atranorin.
Habitat and distribution: On exposed sandstone in campo rupestre; known only from Brazil.
Additional sequenced specimens examined: BRAZIL, Minas Gerais, Reserva Particular do Patrimônio Natural Santuário do Caraça, Catas Altas, 7°2′1″ S, 36°3′1″ W, on sandstone, 17–25 May 2021, A. Aptroot and L.A. Santos (ise–, cgms–,51230, 51622).
Notes: The new species is characterized by an areolate thallus and large, immersed apothecia with black disc and dark brown hypothecium. In the field, L. nigrilobulata can be confused with L. oreinoides (Körb.) Hertel and Rambold, but the latter differs by the shape of the areoles, which are becoming almost rounded, and the hyaline hypothecium. Lecanora nigrilobulata is also similar to L. subimmersa (Fée) Vain., but the latter has a non-areolate thallus surface, brown apothecial disc, and hyaline hypothecium.
Lecanora notatictria L.A. Santos, M. Cáceres, Aptroot and Lücking, sp. nov. (
Figure 9)
MycoBank MB 847942.
Etymology: The name refers to the presence of notatic, isonotatic, subnotatic acids.
Typus: BRAZIL, Pernambuco, Parque Nacional Vale do Catimbau, Buíque, 8°30′20.7″ S, 37°16′34.3″ W, on bark of unidentified tree, 13–15 August 2017, L.A. Santos s.n. (ISE-46529, holotype; ITS Genbank No. OQ650041).
Diagnosis: The new species differs from the similar species Lecanora leprosa Fée and L. notatica Guderley by the chemistry and morphology.
Description: Thallus crustose, corticolous, up to 2 cm wide, c. 0.1 mm thick, areolate, rimose to verruculose, whitish grey, dull, epruinose. Soredia absent. Prothallus absent or whitish grey. Apothecia adnate, 0.3–0.8 (–1) mm diam.; disc pale yellow to brownish orange, epruinose, flat to convex; margin persistent, white, thick, entire, somewhat crenulate. Amphithecium with large crystals which are not dissolving in KOH (= pulicaris-type), parathecium with small crystals. Hymenium hyaline, (–37.5) 45–62 µm high; epihymenium brown to yellowish-brown with the pigment and crystals soluble in KOH (= chlarotera-type), c. 12 µm high. Hypothecium hyaline. Ascospores 8 per ascus, ellipsoid, (8–) 10–13 (–14) × 5–8 μm. Pycnidia not observed.
Chemistry: Thallus and apothecia margin K+ yellow, C–, PD–, UV–. TLC: atranorin, usnic acid, notatic, isonotatic, subnotatic.
Habitat and distribution: On exposed trees in Caatinga forest; known only from Brazil.
Additional sequenced specimens examined: BRAZIL, Paraíba, Reserva Particular do Patrimônio Natural Fazenda das Almas, São José dos Cordeiros, 7°28′15″ S, 36°53′51″ W, on bark of unidentified tree, 22 August 2017, L.A. Santos s.n. (ISE-46519, ISE-57412, ISE-54331, ISE-46539b, ISE-46550).
Notes:
Lecanora notatictria can readily be identified by the pale yellow to brownish orange apothecial discs, the
pulicaris-type amphithecium, a
charotera-type epihymenium and the presence of atranorin, usnic acid, and notatic, isonotatic and subnotatic acids. Morphologically this species resembles
Lecanora achroa Nyl.,
L. helva,
L. leprosa Fée and
L. parachroa L.A. Santos, J.G. Cavalcante and M. Cáceres, but is readily distinguished by its different chemistry. Notatic and subnotatic also occur in
L. notatica Guderley, but this species has larger apothecia and in addition contains confluentic acid [
47].
Lecanora pallidachroa L.A. Santos, Aptroot and M. Cáceres, sp. nov. (
Figure 10)
MycoBank MB 847943.Etymology: The epithet is taken from the Latin pallid-, meaning pale, and achroa, referring similar species Lecanora achroa.
Typus: BRAZIL, Pernambuco, Parque Nacional Vale do Catimbau, Buíque; 8°30′26.7″ S, 37°18′50.9″ W, on bark of unidentified tree, 13–15 August 2017, L.A. Santos s.n. (ISE 54332a, holotype; ITS Genbank No.OQ650046).
Diagnosis: The new species can be distinguished from the somewhat similar species Lecanora achroa by the paler orange disc.
Description: Thallus crustose, corticolous, up to 1.5 cm wide, c. 0.1 mm thick, continuous, rimose to verruculose, whitish grey, dull, epruinose. Soredia absent. Prothallus absent or black. Apothecia adnate, 0.2–0.7 mm diam.; disc pale yellow to yellowish orange, epruinose, flat to slightly concave; margin concolourous the thallus, becoming excluded or persisting, smooth to crenate. Amphithecium with large crystals which are not dissolving in KOH (= pulicaris-type), parathecium with small crystals. Hymenium hyaline, 50–62.5 µm high; epihymenium brown to yellowish-brown with the pigment and crystals soluble in KOH (= chlarotera-type), c. 12 µm high. Hypothecium hyaline. Ascospores 8 per ascus, ellipsoid, (7.5–) 10–12 (–15) × 5–7.5 μm. Pycnidia not observed.
Chemistry: Thallus and apothecia margin K+ yellow, C–, PD–, UV–. TLC: atranorin.
Habitat and distribution: On exposed trees in Caatinga forest; known only from Brazil.
Additional sequenced specimens examined: BRAZIL. Alagoas, Pedra Talhada private area, 9°15′ S, 36°25′35″ W, ca. 500 m; on wooden pole, 21–23 October 2017, M.E.S. Cáceres and A. Aptroot (ise–, ABL–, 42602c)
Notes: Lecanora pallidachroa is most likely to be confused with L. achroa, but differs in the paler orange disc, the smaller ascospores (7.5–) 10–12 (–15) × 5–7.5 vs. 11–16 × 5–8), and the absence of usnic acid.
Lecanora parahelva L.A. Santos, M. Cáceres, Aptroot and Lücking, sp. nov. (
Figure 11)
MycoBank MB 847944.
Etymology: The epithet refers to the superficial morphological similarity of the new species with L. helva.
Typus: BRAZIL, Sergipe, Monumento Natural Grota do Angico, Poço Redondo, 9°39′39.7″ S, 37°41′09.7″ W, on bark of unidentified tree, 24 November 2016, L.A. Santos s.n (ISE-54323, holotype; ITS Genbank No.OQ650113).
Diagnosis: The new species differs from the similar species Lecanora helva by the presence of the alectoronic acid.
Description: Thallus crustose, corticolous, up to 3 cm wide, c. 0.1 mm thick, areolate, rimose to verruculose, yellow to pale orange, dull, epruinose. Soredia absent. Prothallus absent or black to greyish black. Apothecia adnate, 0.2–1.5 mm diam.; disc pale yellow to yellowish orange, epruinose, flat to convex; margin concolourous slightly paler than the thallus, prominent, persisting, thick, entire to wavy, smooth. Amphithecium with small and large crystals (=melacarpella-type), parathecium with small crystals. Hymenium hyaline, 40–62.5 µm high, inspersed; epihymenium brown to yellowish-brown, the pigment and small crystals soluble in KOH (=chlarotera-type), c. 10 µm high. Hypothecium hyaline to pale yellowish (intensifying in KOH). Ascospores 8 per ascus, ellipsoid, (8–) 10–12 × 5–7 μm. Pycnidia not observed.
Chemistry: K+ yellow, C–, P–, medulla apothecial UV+ greenish-white, thallus UV+ orange. TLC: atranorin, 2-chloro-6-O-methyl-norlichexanthone, 4,5 dichloronorlichexanthone and alectoronic acid.
Habitat and distribution: On exposed trees in Caatinga forest; known only from Brazil.
Additional sequenced specimens examined: Sergipe, Monumento Natural Grota do Angico, Poço Redondo, 9°39′39.7″ S, 37°41′09.7″ W, on bark of unidentified tree, 24 November 2016, L.A. Santos s.n (ISE-57414); Pernambuco, Parque Nacional Vale do Catimbau, Buíque, 8°33′29.1″ S, 37°13′58.3″ W, on bark of unidentified tree, 13–15 August 2017, L.A. Santos s.n. (ISE-46531, ISE-46532, ISE-54324, ISE-54326, ISE-54325, ISE-46523, ISE-46530); Bahia, Estação Ecológica Raso da Catarina, Paulo Afonso, 9°39′58.8″ S, 38°28′05.2″ W, on bark of unidentified tree, 10–12 October 2021, L.A. Santos s.n. (ISE-53991, ISE-53989).
Notes: Lecanora parahelva is chemically characterized by the presence of alectoronic acid in addition to atranorin, 2-chloro-6-O-methyl-norlichexanthone and 4,5 dichloronorlichexanthone. Anatomically it can be recognized by the presence of yellow thallus, large apothecia with margins slightly paler than the thallus.
Lecanora saepiphila L.A. Santos, M. Cáceres, Aptroot and Lücking, sp. nov. (
Figure 12)
MycoBank MB 847945.
Etymology: The epithet is taken from the Latin saepi phila, meaning fence, referring to the substrate which the new species was collected.
Typus: BRAZIL, Alagoas, Pedra Talhada private area, 9°15′ S, 36°25′35″ W, ca. 500 m; on wooden pole, 21–23 October 2017, M.E.S. Cáceres and A. Aptroot (ISE-42612b, holotype; ABL, isotype; ITS Genbank No.OQ650100).
Diagnosis: The new species differs from the similar species Lecanora achroa by the absence of usnic acid.
Description: Thallus crustose, corticolous, up to 1.8 cm wide, c. 0.1 mm thick, continuous, thin, white to whitish grey, dull, epruinose. Soredia absent. Prothallus absent or greyish black. Apothecia adnate, 0.3–0.8 (–1) mm diam.; disc pale orange to orange brownish yellow, epruinose, flat to convex; margin concolourous with the thallus, prominent, persisting, entire to wavy, smooth. Amphithecium with large crystals which are not dissolving in KOH (pulicaris-type), parathecium with small crystals. Hymenium hyaline, 42–62.5 µm high; epihymenium brown to yellowish brown, the pigment and small crystals soluble in KOH (=chlarotera-type), c. 12 µm high. Hypothecium hyaline. Ascospores 8 per ascus, ellipsoid, 0.8–10 × 5–6 μm. Pycnidia not observed.
Chemistry: Thallus and apothecia margin K+ yellow, C–, P–, apothecia disc UV+ green. TLC: atranorin, ‘2-O-methylperlatolic.
Habitat and distribution: On wooden pole in Pedra Talhada rain forest remant; known only from Brazil.
Additional sequenced specimens examined: BRAZIL, Alagoas, Pedra Talhada private area, 9°15′ S, 36°25′35″ W, ca. 500 m; on wooden pole, 21–23 October 2017, M.E.S. Cáceres and A. Aptroot (ise–, ABL–,42602b).
Notes:
Lecanora saepiphila is characterized by the small apothecia with pale orange to orange brownish apothecial discs, which are UV+ green. Morphologically it is similar to
L. achroa and
L. parachroa. However, both species have slightly larger ascospores. Moreover, those two species can be readily distinguished from
L. saepiphila by their different chemistry; in addition to atranorin,
L. achroa produces usnic acid and
L. parachroa an unidentified fatty acid [
22].
Lecanora xanthoverrucosa L.A. Santos, M. Cáceres, Aptroot, and Lücking sp. nov. (
Figure 13).
MycoBank MB 847946.
Etymology: The name refers to strongly verrucose thallus and apothecial margins and to the presence of lichexanthone.
Typus: BRAZIL, Sergipe, Monumento Natural Grota do Angico, Poço Redondo; 9°39′39.7″ S, 37°41′09.7″ W, on bark of unidentified tree, 24 November 2016, L.A. Santos s.n. (ISE-52394, holotype; ITS Genbank No. OQ650078).
Diagnosis: The new species can be distinguished from the somewhat similar species Lecanora coronulans Nyl. by the verrucose thallus and apothecial margins and smaller ascospores.
Description: Thallus crustose, corticolous, up to 3 cm wide, c. 0.1 mm thick, areolate to densely verrucose, whitish grey, dull, epruinose. Soredia absent. Prothallus absent or blackish grey. Apothecia adnate, 0.3–0.9 mm diam.; disc dark brown to black, epruinose, flat to convex; margin concolourous with thallus, prominent, crenate or verruculose to densely verrucose, whitish to whitish grey. Amphithecium with small and large crystals (=melacarpella-type), parathecium with crystals. Upper part of hymenium brownish, lower part hyaline, 50–62.5 µm high; epihymenium brownish with crystals, pigmentation not altered by KOH (=pulicaris-type), c. 12 µm high. Hypothecium reddish brown. Ascospores 8 per ascus, broad ellipsoid, (7.5–) 8–11(–12) × 5–7 μm. Pycnidia not observed.
Chemistry: Thallus and apothecia margin K+ yellow, C–, P–, UV+ yellow. TLC: atranorin and a xanthone, possibly lichexanthone.
Habitat and distribution: On exposed trees in Caatinga forest; known only from Brazil.
Additional sequenced specimens examined: BRAZIL, Sergipe, Monumento Natural Grota do Angico, Poço Redondo; 9°39′39.7″ S, 37°41′09.7″ W, on bark of unidentified tree, 24 November 2016, L.A. Santos s.n. (ISE-46520); Povoado Rio dos Negros, Carira; 9°29′ S, 37°48′ W, on bark of unidentified tree, 14 November 2019, L.A. Santos s.n. (ISE-52392, ISE-52393); Alagoas, Pedra Talhada private area, 9°15′ S, 36°25′35″ W, ca. 500 m; on wooden pole, 21–23 October 2017, M.E.S. Cáceres and A. Aptroot (ISE-42608c; ABL); Bahia, Lençois, Chapada Diamantina; 12˚33′33″ S, 41˚23′20″ W, on bark of unidentified tree, 21 July 2017, M.E.S. Cáceres and A. Aptroot (ise–, ABL–, 40532c)
Notes: Lecanora xanthoverrucosa represents another undescribed member of the L. coronulans group. It resembles L. coronulans, L. hypocrocina and L. egranulosa Nyl., but can be readily distinguished by the pigmentation of the epihymenium, the verruculose thallus, and the presence of a xanthone.


 and
and 




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}