
Engineering Flocculation for Improved Tolerance and Production of d-Lactic Acid in Pichia pastoris

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Media, Strains, and Transformation
2.2. Plasmid Construction
2.3. Quantifying Production of d-Lactic Acid and Other Metabolites
2.4. RNA Isolation and Transcriptomics Analysis
2.5. Real-Time PCR
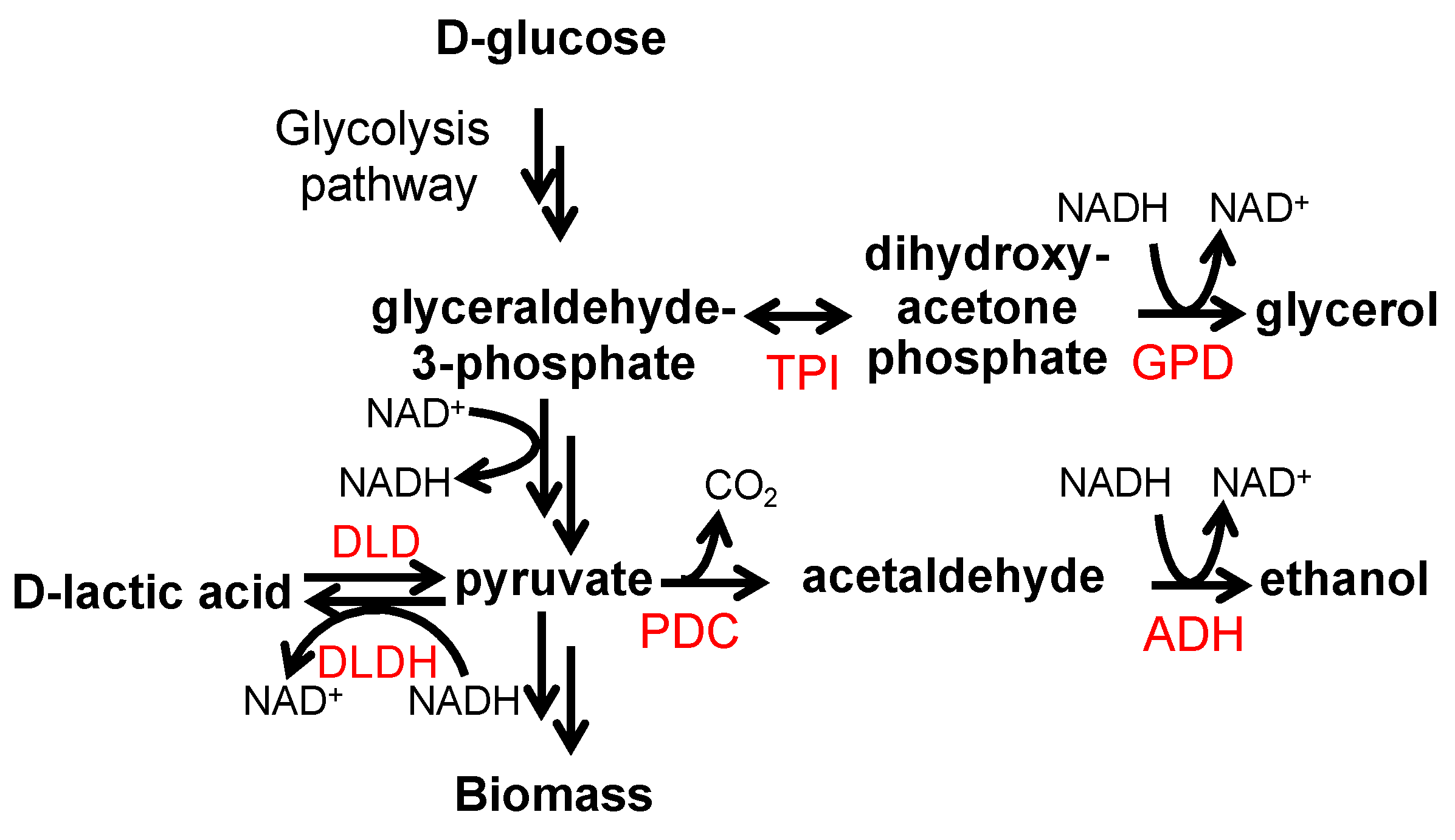
3. Results and Discussion
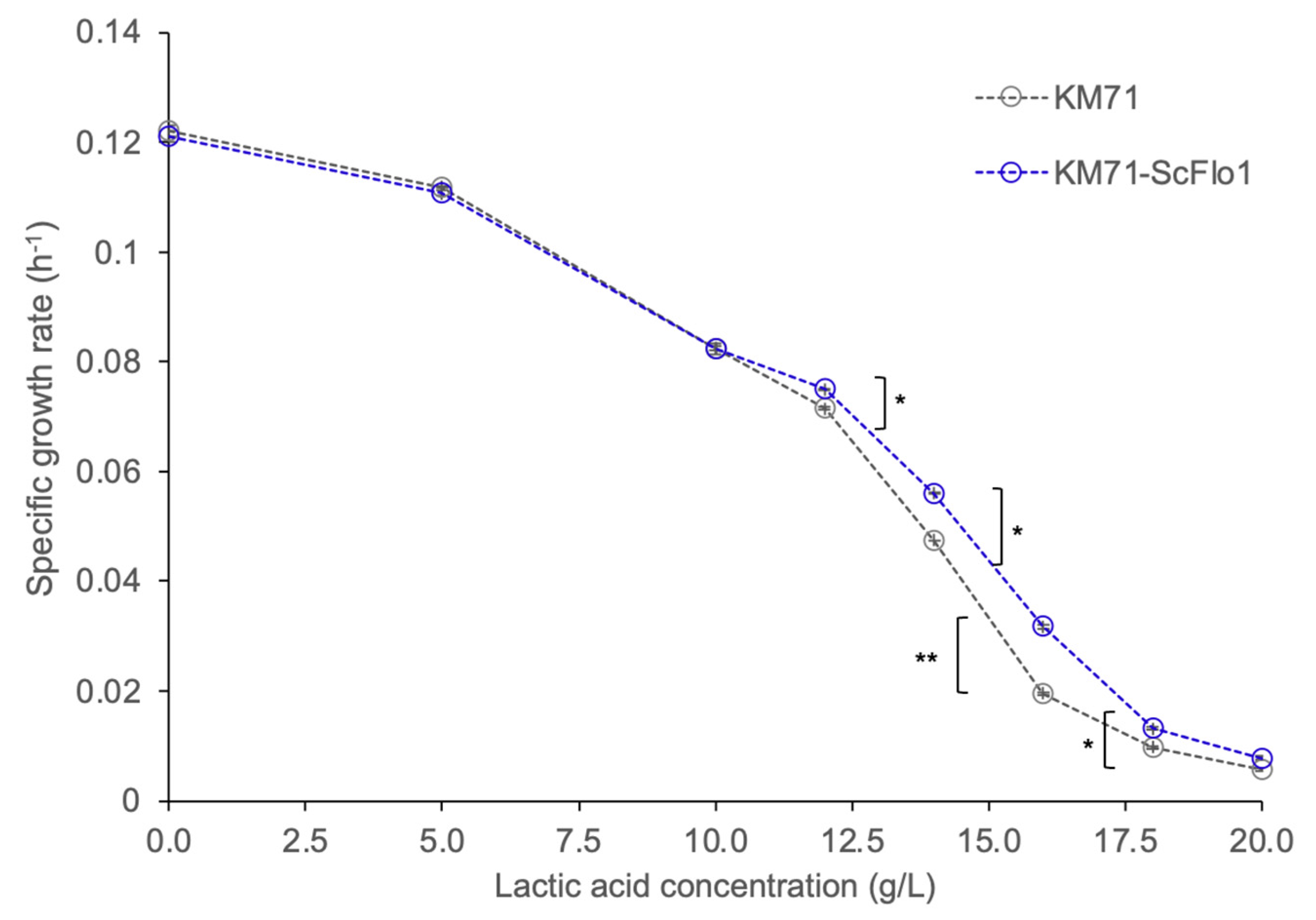
3.1. Expression of the Flocculation Protein Flo1 in P. pastoris Improves d-Lactic Acid Tolerance
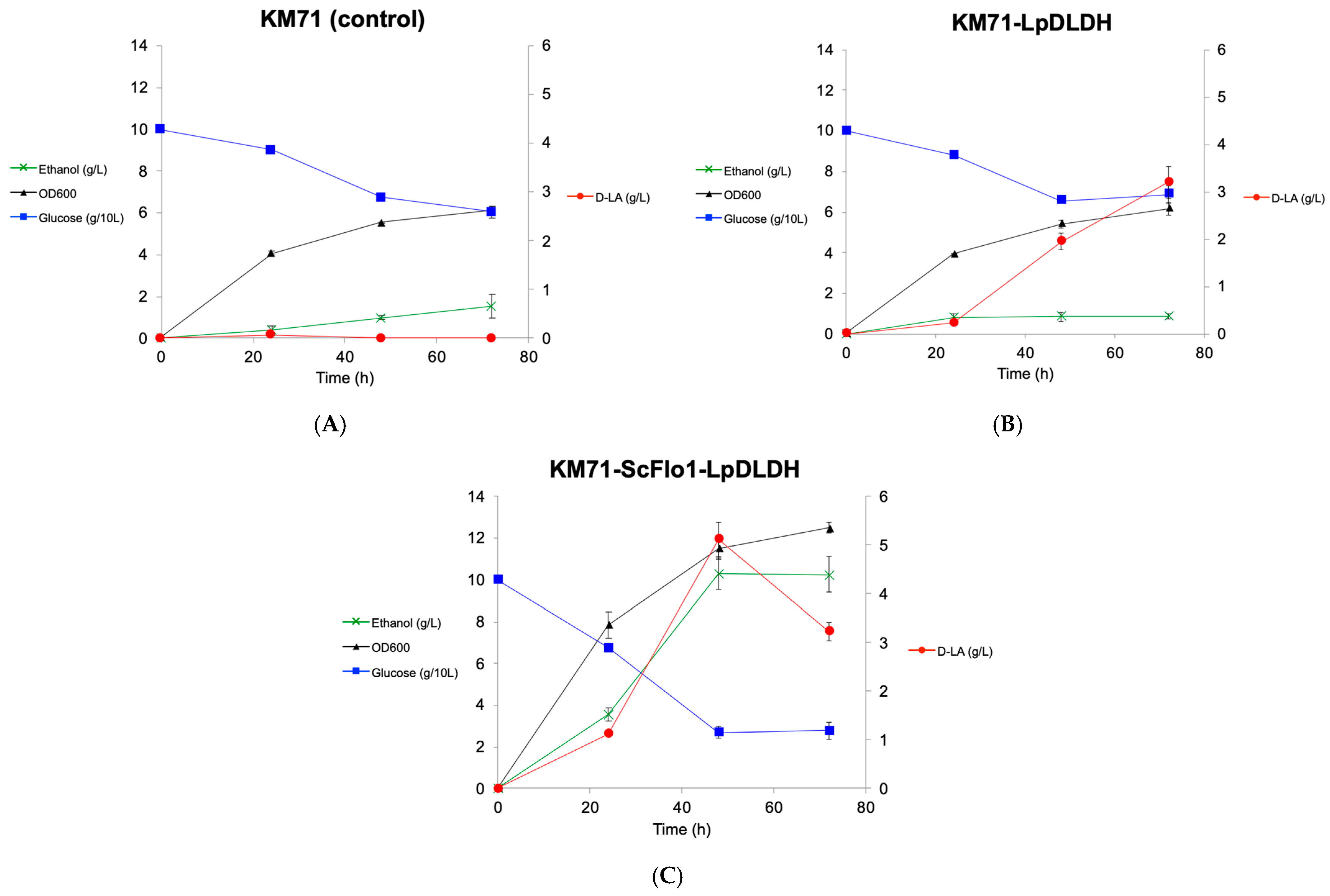
3.2. Expression of S. cerevisiae Flocculation Protein Flo1 in P. pastoris Improves d-Lactic Acid Production
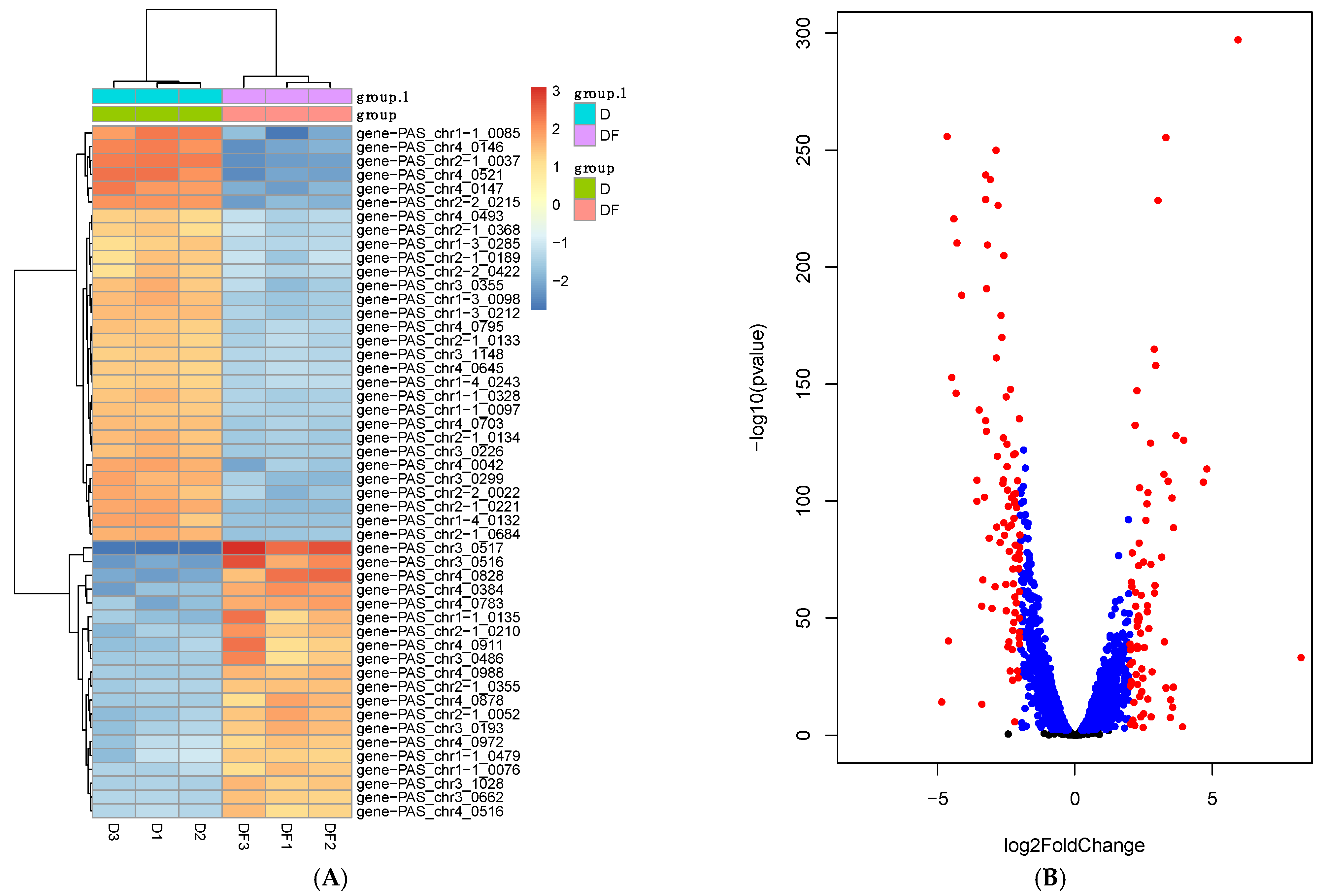
3.3. Transcriptomic Analysis of Engineered Strains
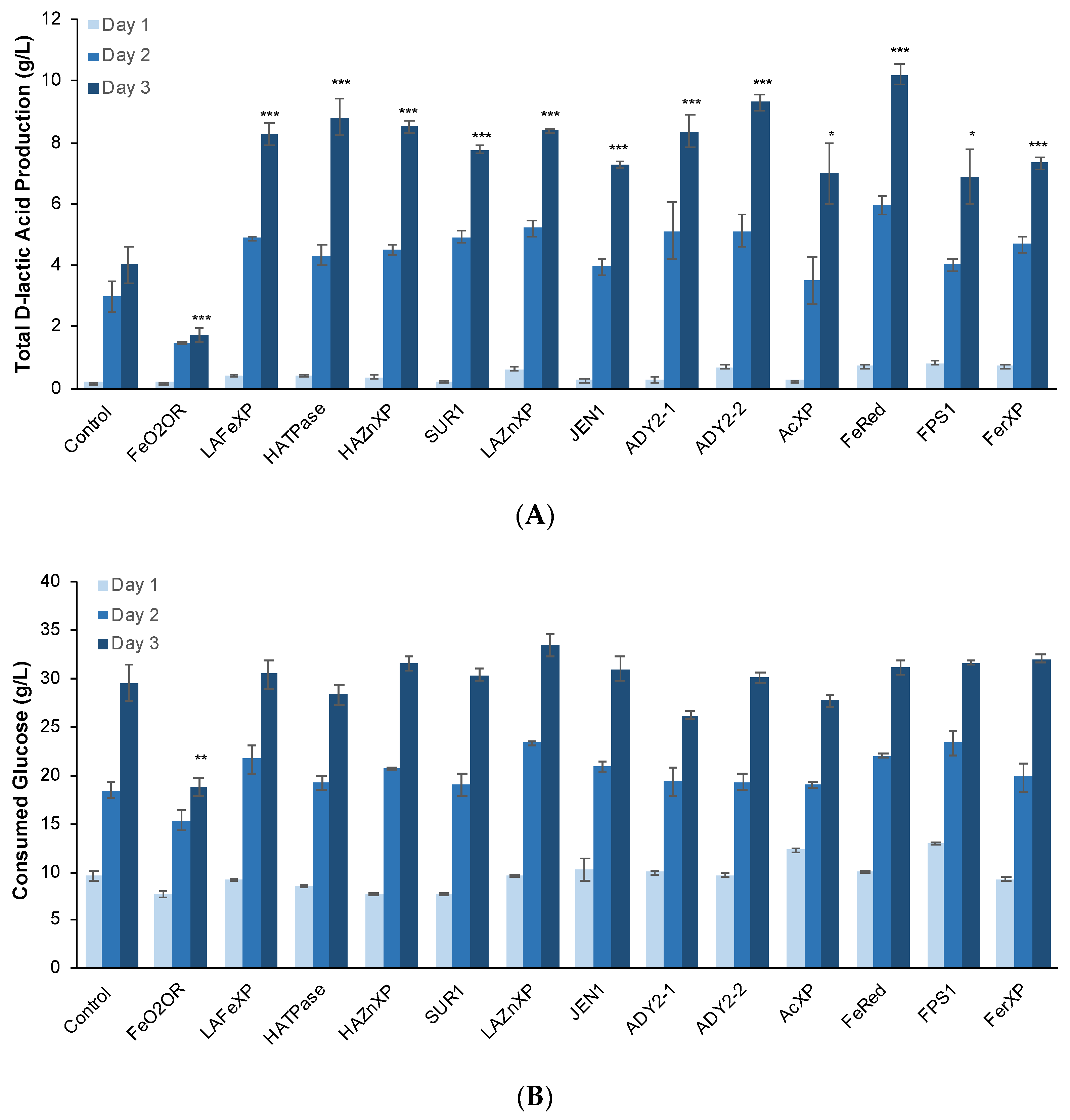
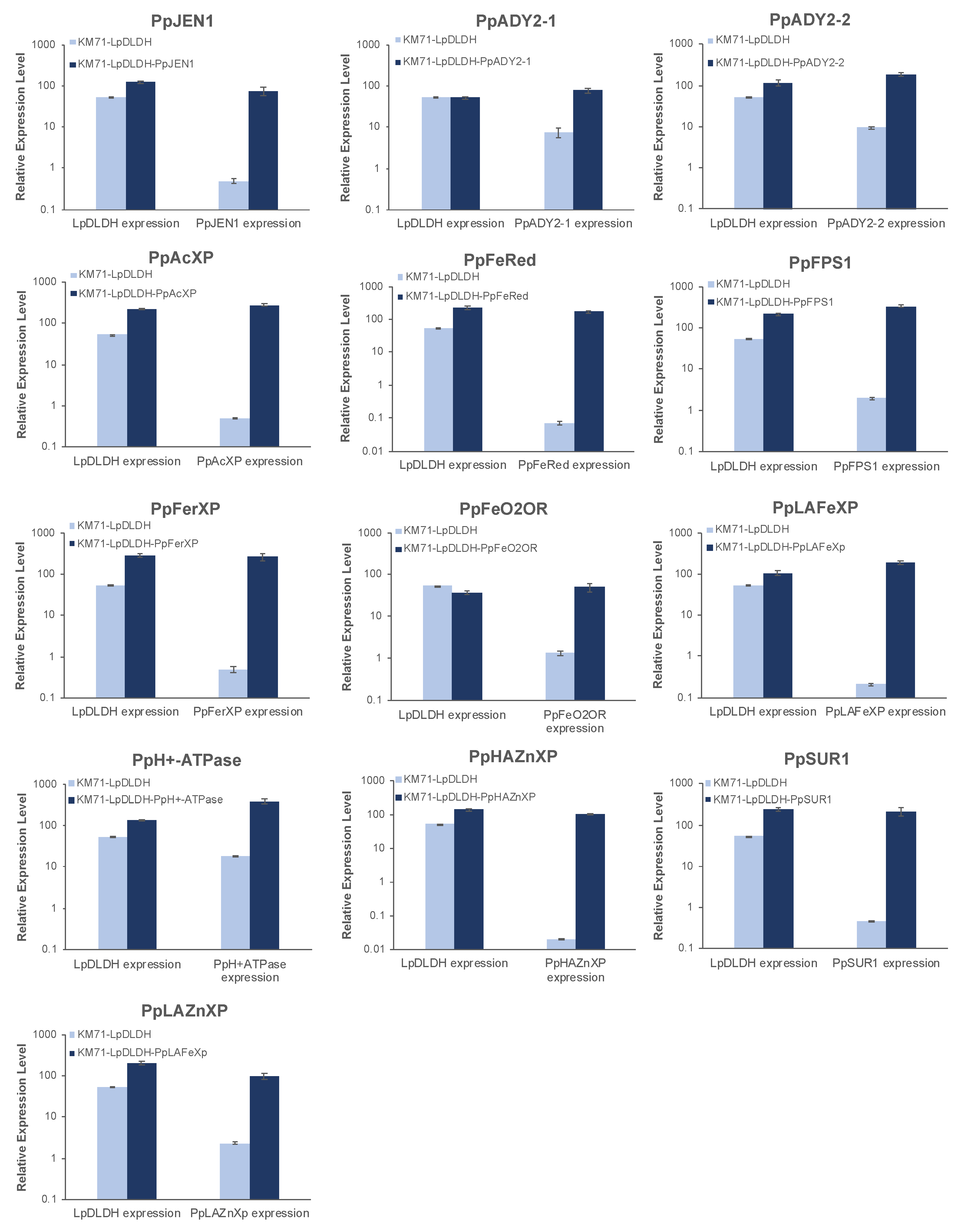
3.4. Overexpression of Several DE Genes Improved d-Lactic Acid Production
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liao, J.C.; Mi, L.; Pontrelli, S.; Luo, S. Fuelling the Future: Microbial Engineering for the Production of Sustainable Biofuels. Nat. Rev. Microbiol. 2016, 14, 288–304. [Google Scholar] [CrossRef] [PubMed]
- Watcharawipas, A.; Sae-Tang, K.; Sansatchanon, K.; Sudying, P.; Boonchoo, K.; Tanapongpipat, S.; Kocharin, K.; Runguphan, W. Systematic Engineering of Saccharomyces Cerevisiae for D-Lactic Acid Production with near Theoretical Yield. FEMS Yeast Res. 2021, 21, foab024. [Google Scholar] [CrossRef] [PubMed]
- Atsumi, S.; Hanai, T.; Liao, J.C. Non-Fermentative Pathways for Synthesis of Branched-Chain Higher Alcohols as Biofuels. Nature 2008, 451, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Kruyer, N.S.; Peralta-Yahya, P. Metabolic Engineering Strategies to Bio-Adipic Acid Production. Curr. Opin. Biotechnol. 2017, 45, 136–143. [Google Scholar] [CrossRef]
- Moon, T.S.; Yoon, S.H.; Lanza, A.M.; Roy-Mayhew, J.D.; Jones Prather, K.L. Production of Glucaric Acid from a Synthetic Pathway in Recombinant Escherichia Coli. Appl. Environ. Microbiol. 2009, 75, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.; Wang, J.; Zhao, Y.; Deng, Y. Metabolic Engineering of Saccharomyces Cerevisiae for Efficient Production of Glucaric Acid at High Titer. Microb. Cell Fact. 2018, 17, 67. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.J.; Lee, S.Y. Microbial Production of Short-Chain Alkanes. Nature 2013, 502, 571–574. [Google Scholar] [CrossRef]
- Schirmer, A.; Rude, M.A.; Li, X.; Popova, E.; Del Cardayre, S.B. Microbial Biosynthesis of Alkanes. Science 2010, 329, 559–562. [Google Scholar] [CrossRef]
- Castro-Aguirre, E.; Iñiguez-Franco, F.; Samsudin, H.; Fang, X.; Auras, R. Poly(Lactic Acid)—Mass Production, Processing, Industrial Applications, and End of Life. Adv. Drug Deliv. Rev. 2016, 107, 333–366. [Google Scholar] [CrossRef] [Green Version]
- Lahtinen, S.; Ouwehand, A.C.; Salminen, S.; von Wright, A. Lactic Acid Bacteria: Microbiological and Functional Aspects, 4th ed.; CRC Press: Boca Raton, FL, USA, 2011; ISBN 978-1-43-983678-1. [Google Scholar]
- Bai, H.; Deng, S.; Bai, D.; Zhang, Q.; Fu, Q. Recent Advances in Processing of Stereocomplex-Type Polylactide. Macromol. Rapid Commun. 2017, 38, 1700454. [Google Scholar] [CrossRef] [Green Version]
- Ikada, Y.; Jamshidi, K.; Tsuji, H.; Hyon, S.H. Stereocomplex Formation between Enantiomeric Poly(Lactides). Macromolecules 2002, 20, 904–906. [Google Scholar] [CrossRef]
- Luo, F.; Fortenberry, A.; Ren, J.; Qiang, Z. Recent Progress in Enhancing Poly(Lactic Acid) Stereocomplex Formation for Material Property Improvement. Front. Chem. 2020, 8, 688. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rahman, M.A.; Sonomoto, K. Opportunities to Overcome the Current Limitations and Challenges for Efficient Microbial Production of Optically Pure Lactic Acid. J. Biotechnol. 2016, 236, 176–192. [Google Scholar] [CrossRef]
- Bai, Z.; Gao, Z.; Sun, J.; Wu, B.; He, B. D-Lactic Acid Production by Sporolactobacillus Inulinus YBS1-5 with Simultaneous Utilization of Cottonseed Meal and Corncob Residue. Bioresour. Technol. 2016, 207, 346–352. [Google Scholar] [CrossRef]
- Coelho, L.F.; De Lima, C.J.B.; Bernardo, M.P.; Contiero, J. D(-)-Lactic Acid Production by Leuconostoc Mesenteroides B512 Using Different Carbon and Nitrogen Sources. Appl. Biochem. Biotechnol. 2011, 164, 1160–1171. [Google Scholar] [CrossRef]
- Zhu, T.; Sun, H.; Wang, M.; Li, Y. Pichia Pastoris as a Versatile Cell Factory for the Production of Industrial Enzymes and Chemicals: Current Status and Future Perspectives. Biotechnol. J. 2019, 14, 1800694. [Google Scholar] [CrossRef] [PubMed]
- Siripong, W.; Wolf, P.; Kusumoputri, T.P.; Downes, J.J.; Kocharin, K.; Tanapongpipat, S.; Runguphan, W. Metabolic Engineering of Pichia Pastoris for Production of Isobutanol and Isobutyl Acetate. Biotechnol. Biofuels 2018, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- Siripong, W.; Angela, C.; Tanapongpipat, S.; Runguphan, W. Metabolic Engineering of Pichia Pastoris for Production of Isopentanol (3-Methyl-1-Butanol). Enzym. Microb. Technol. 2020, 138, 109557. [Google Scholar] [CrossRef]
- Wriessnegger, T.; Augustin, P.; Engleder, M.; Leitner, E.; Müller, M.; Kaluzna, I.; Schürmann, M.; Mink, D.; Zellnig, G.; Schwab, H.; et al. Production of the Sesquiterpenoid (+)-Nootkatone by Metabolic Engineering of Pichia Pastoris. Metab. Eng. 2014, 24, 18–29. [Google Scholar] [CrossRef]
- Bhataya, A.; Schmidt-Dannert, C.; Lee, P.C. Metabolic Engineering of Pichia Pastoris X-33 for Lycopene Production. Process Biochem. 2009, 44, 1095–1102. [Google Scholar] [CrossRef]
- Peña, D.A.; Gasser, B.; Zanghellini, J.; Steiger, M.G.; Mattanovich, D. Metabolic Engineering of Pichia Pastoris. Metab. Eng. 2018, 50, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Haon, M.; Grisel, S.; Navarro, D.; Gruet, A.; BERRIN, J.-G.; Bignon, C. Recombinant Protein Production Facility for Fungal Biomass-Degrading Enzymes Using the Yeast Pichia Pastoris. Front. Microbiol. 2015, 6, 1002. [Google Scholar] [CrossRef] [Green Version]
- Lima, P.B.A.; Mulder, K.C.L.; Melo, N.T.M.; Carvalho, L.S.; Menino, G.S.; Mulinari, E.; Castro, V.H.; Reis, T.F.; Goldman, G.H.; Magalhães, B.S.; et al. Novel Homologous Lactate Transporter Improves L-Lactic Acid Production from Glycerol in Recombinant Strains of Pichia Pastoris. Microb. Cell Fact. 2016, 15, 1–9. [Google Scholar] [CrossRef]
- Yamada, R.; Ogura, K.; Kimoto, Y.; Ogino, H. Toward the Construction of a Technology Platform for Chemicals Production from Methanol: D-Lactic Acid Production from Methanol by an Engineered Yeast Pichia Pastoris. World J. Microbiol. Biotechnol. 2019, 35, 37. [Google Scholar] [CrossRef] [PubMed]
- Peetermans, A.; Foulquié-Moreno, M.R.; Thevelein, J.M. Mechanisms Underlying Lactic Acid Tolerance and Its Influence on Lactic Acid Production in Saccharomyces Cerevisiae. Microb. Cell 2021, 8, 111–130. [Google Scholar] [CrossRef]
- Ye, P.L.; Wang, X.Q.; Yuan, B.; Liu, C.G.; Zhao, X.Q. Manipulating Cell Flocculation-Associated Protein Kinases in Saccharomyces Cerevisiae Enables Improved Stress Tolerance and Efficient Cellulosic Ethanol Production. Bioresour. Technol. 2022, 348, 126758. [Google Scholar] [CrossRef]
- Westman, J.O.; Mapelli, V.; Taherzadeh, M.J.; Franzén, C.J. Flocculation Causes Inhibitor Tolerance in Saccharomyces Cerevisiae for Second-Generation Bioethanol Production. Appl. Environ. Microbiol. 2014, 80, 6908–6918. [Google Scholar] [CrossRef] [Green Version]
- Bumrungtham, P.; Promdonkoy, P.; Prabmark, K.; Bunterngsook, B.; Boonyapakron, K.; Tanapongpipat, S.; Champreda, V.; Runguphan, W. Engineered Production of Isobutanol from Sugarcane Trash Hydrolysates in Pichia pastoris. J. Fungi 2022, 8, 767. [Google Scholar] [CrossRef]
- Sornlek, W.; Sae-Tang, K.; Watcharawipas, A.; Wongwisansri, S.; Tanapongpipat, S.; Eurwilaichtr, L.; Champreda, V.; Runguphan, W.; Schaap, P.J.; Martins dos Santos, V.A.P. D-Lactic Acid Production from Sugarcane Bagasse by Genetically Engineered Saccharomyces Cerevisiae. J. Fungi 2022, 8, 816. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python Framework to Work with High-Throughput Sequencing Data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, E.V. Flocculation in Saccharomyces Cerevisiae: A Review. J. Appl. Microbiol. 2011, 110, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishida-Fujii, K.; Goto, S.; Sugiyama, H.; Takagi, Y.; Saiki, T.; Takagi, M. Breeding of Flocculent Industrial Alcohol Yeast Strains by Self-Cloning of the Flocculation Gene FLO1 and Repeated-Batch Fermentation by Transformants. J. Gen. Appl. Microbiol. 1998, 44, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.Z.; Shen, W.; Rao, Z.M.; Fang, H.Y.; Zhan, X.B.; Zhuge, J. Construction of a Flocculating Yeast for Fuel Ethanol Production. Biotechnol. Lett. 2008, 30, 97–102. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Derdelinckx, G.; Verachtert, H.; Delvaux, F.R. Yeast Flocculation: What Brewers Should Know. Appl. Microbiol. Biotechnol. 2003, 61, 197–205. [Google Scholar] [CrossRef]
- Nonklang, S.; Ano, A.; Babiker, M.A.A.B.; Saito, Y.; Hoshida, H.; Akada, R. Construction of Flocculent Kluyveromyces Marxianus Strains Suitable for High-Temperature Ethanol Fermentation. Biosci. Biotechnol. Biochem. 2009, 73, 1090–1095. [Google Scholar] [CrossRef]
- Almeida, C.; Queirós, O.; Wheals, A.; Teixeira, J.; Moradas-Ferreira, P. Acquisition of Flocculation Phenotype by Kluyveromyces Marxianus When Overexpressing GAP1 Gene Encoding an Isoform of Glyceraldehyde-3-Phosphate Dehydrogenase. J. Microbiol. Methods 2003, 55, 433–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellal, M.; Boudrant, J.; Elfoul, L.; Bonaly, R. Flocculation Dispersion in Kluyveromyces Lactis. Process Biochem. 1995, 30, 641–648. [Google Scholar] [CrossRef]
- Claro, F.B.; Rijsbrack, K.; Soares, E.V. Flocculation Onset in Saccharomyces Cerevisiae: Effect of Ethanol, Heat and Osmotic Stress. J. Appl. Microbiol. 2007, 102, 693–700. [Google Scholar] [CrossRef] [Green Version]
- Cunha, A.F.; Missawa, S.K.; Gomes, L.H.; Reis, S.F.; Pereira, G.A.G. Control by Sugar of Saccharomyces Cerevisiae Flocculation for Industrial Ethanol Production. FEMS Yeast Res. 2006, 6, 280–287. [Google Scholar] [CrossRef] [Green Version]
- Miki, B.L.A.; Poon, N.H.; James, A.P.; Seligy, V.L. Possible Mechanism for Flocculation Interactions Governed by Gene FLO1 in Saccharomyces Cerevisiae. J. Bacteriol. 1982, 150, 878–889. [Google Scholar] [CrossRef] [Green Version]
- Teunissen, A.W.R.H.; Steensma, H.Y. Review: The Dominant Flocculation Genes of Saccharomyces Cerevisiae Constitute a New Subtelomeric Gene Family. Yeast 1995, 11, 1001–1013. [Google Scholar] [CrossRef]
- Stewart, G.G. Yeast Flocculation—Sedimentation and Flotation. Ferment 2018, 4, 28. [Google Scholar] [CrossRef] [Green Version]
- El-Behhari, M.; Ngondi Ekomé, J.; Coulon, J.; Pucci, B.; Bonaly, R. Comparative Extraction Procedures for a Galactose-Specific Lectin Involved in Flocculation of Kluyveromyces Lactis Strains. Appl. Microbiol. Biotechnol. 1998, 49, 16–23. [Google Scholar] [CrossRef]
- Al-Mahmood, S.; Colin, S.; Bonaly, R. Kluyveromyces Bulgaricus Yeast Lectins. Isolation of Two Galactose-Specific Lectin Forms from the Yeast Cell Wall. J. Biol. Chem. 1991, 266, 20882–20887. [Google Scholar] [CrossRef] [PubMed]
- Van Mulders, S.E.; Ghequire, M.; Daenen, L.; Verbelen, P.J.; Verstrepen, K.J.; Delvaux, F.R. Flocculation Gene Variability in Industrial Brewer’s Yeast Strains. Appl. Microbiol. Biotechnol. 2010, 88, 1321–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, J.H.; Kim, H.J.; Kim, M.J.; Sung, B.H.; Jeon, J.H.; Kim, H.S.; Jin, Y.S.; Kweon, D.H.; Sohn, J.H. Direct Fermentation of Jerusalem Artichoke Tuber Powder for Production of L-Lactic Acid and d-Lactic Acid by Metabolically Engineered Kluyveromyces Marxianus. J. Biotechnol. 2018, 266, 27–33. [Google Scholar] [CrossRef] [PubMed]
- De Deken, R.H. The Crabtree Effect: A Regulatory System in Yeast. J. Gen. Microbiol. 1966, 44, 149–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumann, K.; Dato, L.; Graf, A.B.; Frascotti, G.; Dragosits, M.; Porro, D.; Mattanovich, D.; Ferrer, P.; Branduardi, P. The Impact of Oxygen on the Transcriptome of Recombinant, S. Cerevisiae and P. Pastoris—A Comparative Analysis. BMC Genom. 2011, 12, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannattasio, S.; Guaragnella, N.; Corte-Real, M.; Passarella, S.; Marra, E. Acid Stress Adaptation Protects Saccharomyces Cerevisiae from Acetic Acid-Induced Programmed Cell Death. Gene 2005, 354, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Bauer, B.E.; Rossington, D.; Mollapour, M.; Mamnun, Y.; Kuchler, K.; Piper, P.W. Weak Organic Acid Stress Inhibits Aromatic Amino Acid Uptake by Yeast, Causing a Strong Influence of Amino Acid Auxotrophies on the Phenotypes of Membrane Transporter Mutants. Eur. J. Biochem. 2003, 270, 3189–3195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.S.; Cho, K.H.; Park, K.H.; Jang, J.; Hahn, J.S. Activation of Haa1 and War1 Transcription Factors by Differential Binding of Weak Acid Anions in Saccharomyces Cerevisiae. Nucleic Acids Res. 2019, 47, 1211–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.M.; Xiong, L.; Tang, Y.J.; Mehmood, M.A.; Zhao, Z.K.; Bai, F.W.; Zhao, X.Q. Enhanced Acetic Acid Stress Tolerance and Ethanol Production in Saccharomyces Cerevisiae by Modulating Expression of the de Novo Purine Biosynthesis Genes. Biotechnol. Biofuels 2019, 12, 116. [Google Scholar] [CrossRef] [PubMed]
- Mira, N.P.; Teixeira, M.C.; Sá-Correia, I. Adaptive Response and Tolerance to Weak Acids in Saccharomyces Cerevisiae: A Genome-Wide View. Omi. A J. Integr. Biol. 2010, 14, 525–540. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.Q.; Bai, F.W. Zinc and Yeast Stress Tolerance: Micronutrient Plays a Big Role. J. Biotechnol. 2012, 158, 176–183. [Google Scholar] [CrossRef]
- Fernandes, A.R.; Mira, N.P.; Vargas, R.C.; Canelhas, I.; Sá-Correia, I. Saccharomyces Cerevisiae Adaptation to Weak Acids Involves the Transcription Factor Haa1p and Haa1p-Regulated Genes. Biochem. Biophys. Res. Commun. 2005, 337, 95–103. [Google Scholar] [CrossRef]
- Sugiyama, M.; Akase, S.P.; Nakanishi, R.; Horie, H.; Kaneko, Y.; Harashima, S. Nuclear Localization of Haa1, Which Is Linked to Its Phosphorylation Status, Mediates Lactic Acid Tolerance in Saccharomyces Cerevisiae. Appl. Environ. Microbiol. 2014, 80, 3488–3495. [Google Scholar] [CrossRef] [Green Version]
- Kawahata, M.; Masaki, K.; Fujii, T.; Iefuji, H. Yeast Genes Involved in Response to Lactic Acid and Acetic Acid: Acidic Conditions Caused by the Organic Acids in Saccharomyces Cerevisiae Cultures Induce Expression of Intracellular Metal Metabolism Genes Regulated by Aft1p. FEMS Yeast Res. 2006, 6, 924–936. [Google Scholar] [CrossRef] [Green Version]
- Maris, A.J.A.V.; Konings, W.N.; Dijken, J.P.V.; Pronk, J.T. Microbial Export of Lactic and 3-Hydroxypropanoic Acid: Implications for Industrial Fermentation Processes. Metab. Eng. 2004, 6, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, A.; Talaia, G.; Sá-Pessoa, J.; Bessa, D.; Gonçalves, M.J.; Moreira, R.; Paiva, S.; Casal, M.; Queirós, O. Lactic Acid Production in Saccharomyces Cerevisiae Is Modulated by Expression of the Monocarboxylate Transporters Jen1 and Ady2. FEMS Yeast Res. 2012, 12, 375–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDermott, J.R.; Rosen, B.P.; Liu, Z. Jen1p: A High Affinity Selenite Transporter in Yeast. Mol. Biol. Cell 2010, 21, 3934–3941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casal, M.; Paiva, S.; Andrade, R.P.; Gancedo, C.; Leão, C. The Lactate-Proton Symport of Saccharomyces Cerevisiae Is Encoded by JEN1. J. Bacteriol. 1999, 181, 2620–2623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branduardi, P.; Sauer, M.; De Gioia, L.; Zampella, G.; Valli, M.; Mattanovich, D.; Porro, D. Lactate Production Yield from Engineered Yeasts Is Dependent from the Host Background, the Lactate Dehydrogenase Source and the Lactate Export. Microb. Cell Fact. 2006, 5, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakamatsu, M.; Tomitaka, M.; Tani, T.; Taguchi, H.; Kida, K.; Akamatsu, T. Improvement of Ethanol Production from D-Lactic Acid by Constitutive Expression of Lactate Transporter Jen1p in Saccharomyces Cerevisiae. Biosci. Biotechnol. Biochem. 2013, 77, 1114–1116. [Google Scholar] [CrossRef] [Green Version]
- Paiva, S.; Devaux, F.; Barbosa, S.; Jacq, C.; Casal, M. Ady2p Is Essential for the Acetate Permease Activity in the Yeast Saccharomyces Cerevisiae. Yeast 2004, 21, 201–210. [Google Scholar] [CrossRef] [Green Version]
- Turner, T.L.; Lane, S.; Jayakody, L.N.; Zhang, G.C.; Kim, H.; Cho, W.; Jin, Y.S. Deletion of JEN1 and ADY2 Reduces Lactic Acid Yield from an Engineered Saccharomyces Cerevisiae, in Xylose Medium, Expressing a Heterologous Lactate Dehydrogenase. FEMS Yeast Res. 2019, 19, foz050. [Google Scholar] [CrossRef]
- Cassio, F.; Leao, C.; Van Uden, N. Transport of Lactate and Other Short-Chain Monocarboxylates in the Yeast Saccharomyces Cerevisiae. Appl. Environ. Microbiol. 1987, 53, 509–513. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Zhang, K.; Mehmood, M.A.; Zhao, Z.K.; Bai, F.; Zhao, X. Deletion of Acetate Transporter Gene ADY2 Improved Tolerance of Saccharomyces Cerevisiae against Multiple Stresses and Enhanced Ethanol Production in the Presence of Acetic Acid. Bioresour. Technol. 2017, 245, 1461–1468. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name | Overexpressed Genes | Description | References |
|---|---|---|---|
| KM71 | None | Laboratory strain | Invitrogen |
| KM71-ScFlo1 | ScFLO1 | KM71 overexpressing ScFLO1 from S. cerevisiae | This study |
| KM71-ScFlo1-LpDLDH | ScFLO1, LpDLDH | KM71 overexpressing ScFLO1 from S. cerevisiae and LpDLDH from Leuconostoc pseudomesenteroides | This study |
| KM71-LpDLDH | LpDLDH | KM71 overexpressing LpDLDH from L. pseudomesenteroides | This study |
| KM71-LpDLDH-PpJEN1 | LpDLDH, PpJEN1 | KM71-LpDLDH overexpressing endogenous JEN1 | This study |
| KM71-LpDLDH-PpADY2-1 | LpDLDH, PpADY2-1 | KM71-LpDLDH overexpressing endogenous ADY2-1 | This study |
| KM71-LpDLDH-PpADY2-2 | LpDLDH, PpADY2-2 | KM71-LpDLDH overexpressing endogenous ADY2-2 | This study |
| KM71-LpDLDH-PpFeRed | LpDLDH, PpFeRed | KM71-LpDLDH overexpressing endogenous FeRed | This study |
| KM71-LpDLDH-PpAcXP | LpDLDH, PpAcXP | KM71-LpDLDH overexpressing endogenous AcXP | This study |
| KM71-LpDLDH-PpLAZnXP | LpDLDH, PpLAZnXP | KM71-LpDLDH overexpressing endogenous LAZnXP | This study |
| KM71-LpDLDH-PpHAZnXP | LpDLDH, PpHAZnXP | KM71-LpDLDH overexpressing endogenous HAZnXP | This study |
| KM71-LpDLDH-PpFPS1 | LpDLDH, PpFPS1 | KM71-LpDLDH overexpressing endogenous FPS1 | This study |
| KM71-LpDLDH-PpFerXP | LpDLDH, PpFerXP | KM71-LpDLDH overexpressing endogenous FerXP | This study |
| KM71-LpDLDH-PpSUR1 | LpDLDH, PpSUR1 | KM71-LpDLDH overexpressing endogenous SUR1 | This study |
| KM71-LpDLDH-PpH+-ATPase | LpDLDH, PpH+-ATPase | KM71-LpDLDH overexpressing endogenous H+-ATPase | This study |
| KM71-LpDLDH-PpFeO2OR | LpDLDH, PpFeO2OR | KM71-LpDLDH overexpressing endogenous FeO2OR | This study |
| KM71-LpDLDH-PpLAFeXp | LpDLDH, PpLAFeXP | KM71-LpDLDH overexpressing endogenous LAFeXP | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sae-Tang, K.; Bumrungtham, P.; Mhuantong, W.; Champreda, V.; Tanapongpipat, S.; Zhao, X.-Q.; Liu, C.-G.; Runguphan, W. Engineering Flocculation for Improved Tolerance and Production of d-Lactic Acid in Pichia pastoris. J. Fungi 2023, 9, 409. https://doi.org/10.3390/jof9040409
Sae-Tang K, Bumrungtham P, Mhuantong W, Champreda V, Tanapongpipat S, Zhao X-Q, Liu C-G, Runguphan W. Engineering Flocculation for Improved Tolerance and Production of d-Lactic Acid in Pichia pastoris. Journal of Fungi. 2023; 9(4):409. https://doi.org/10.3390/jof9040409
Chicago/Turabian StyleSae-Tang, Kittapong, Pornsiri Bumrungtham, Wuttichai Mhuantong, Verawat Champreda, Sutipa Tanapongpipat, Xin-Qing Zhao, Chen-Guang Liu, and Weerawat Runguphan. 2023. "Engineering Flocculation for Improved Tolerance and Production of d-Lactic Acid in Pichia pastoris" Journal of Fungi 9, no. 4: 409. https://doi.org/10.3390/jof9040409