
Biological Crust Diversity Related to Elevation and Soil Properties at Local Scale in a Montane Scrub of Ecuador

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
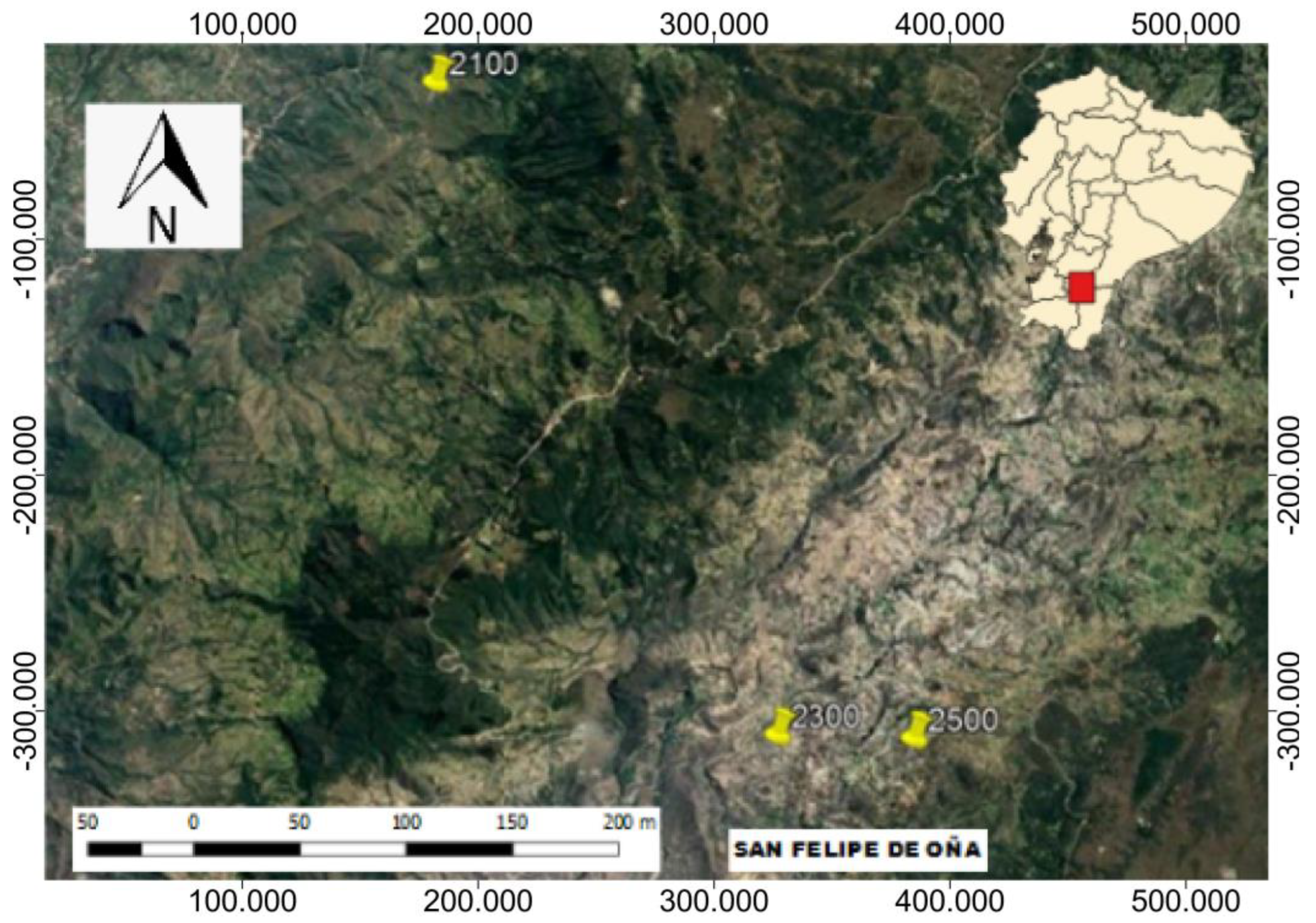
2.1. Study Area


2.2. Design and Data Collection
2.3. Soil Sampling
2.4. Data Analysis
3. Results
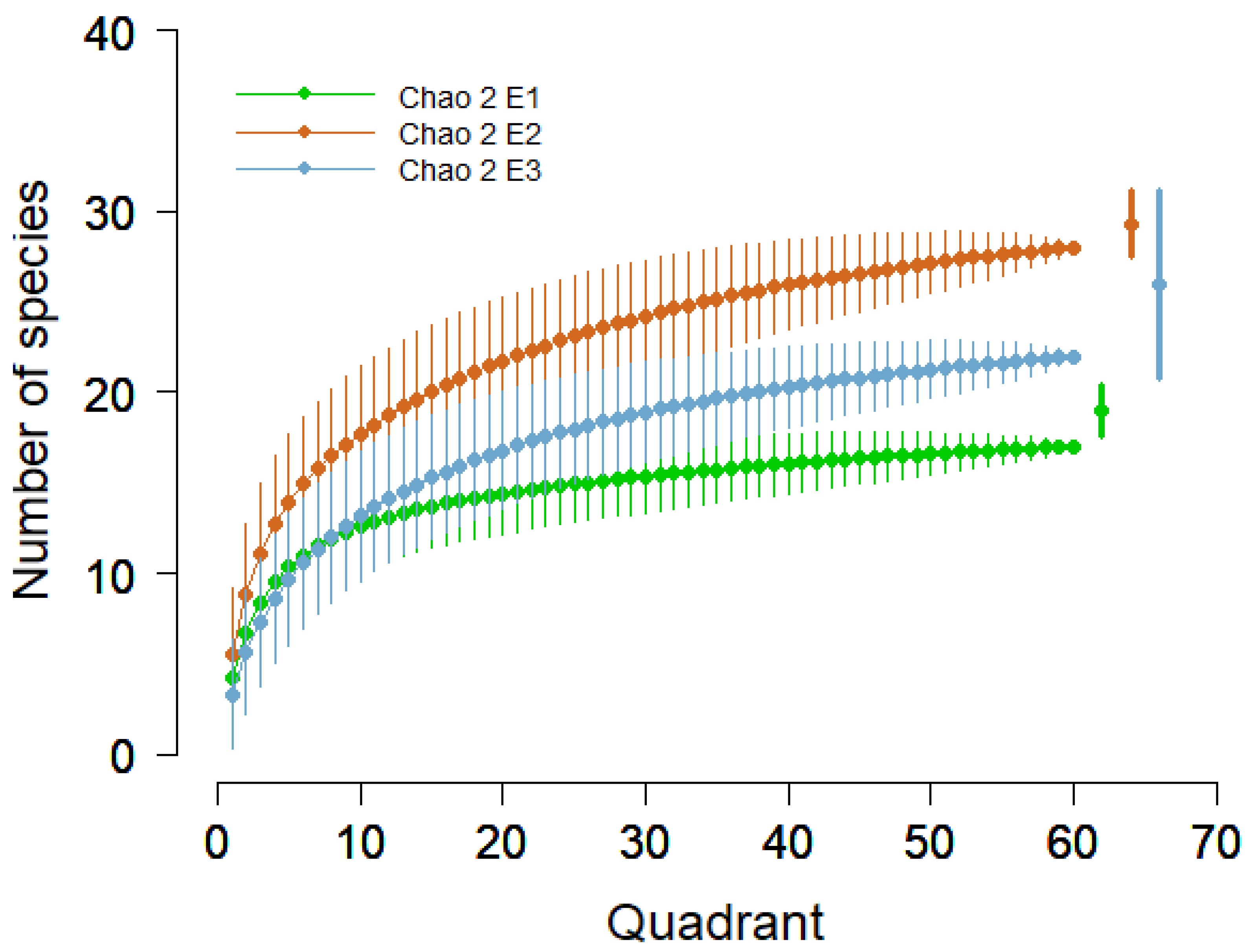
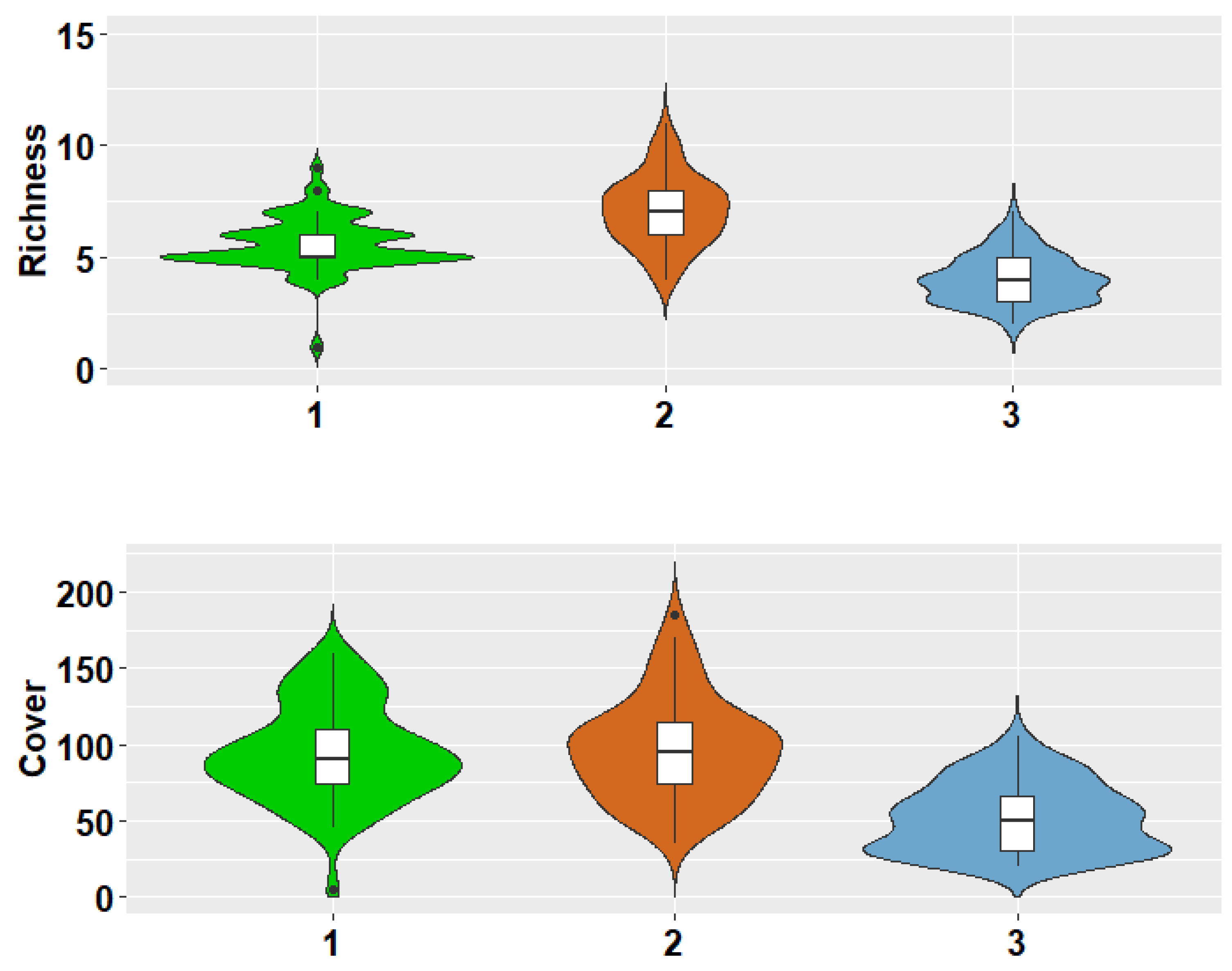
3.1. Alpha Diversity
3.2. Characteristics of the Physical-Chemical Properties of the Soil in Three Elevations
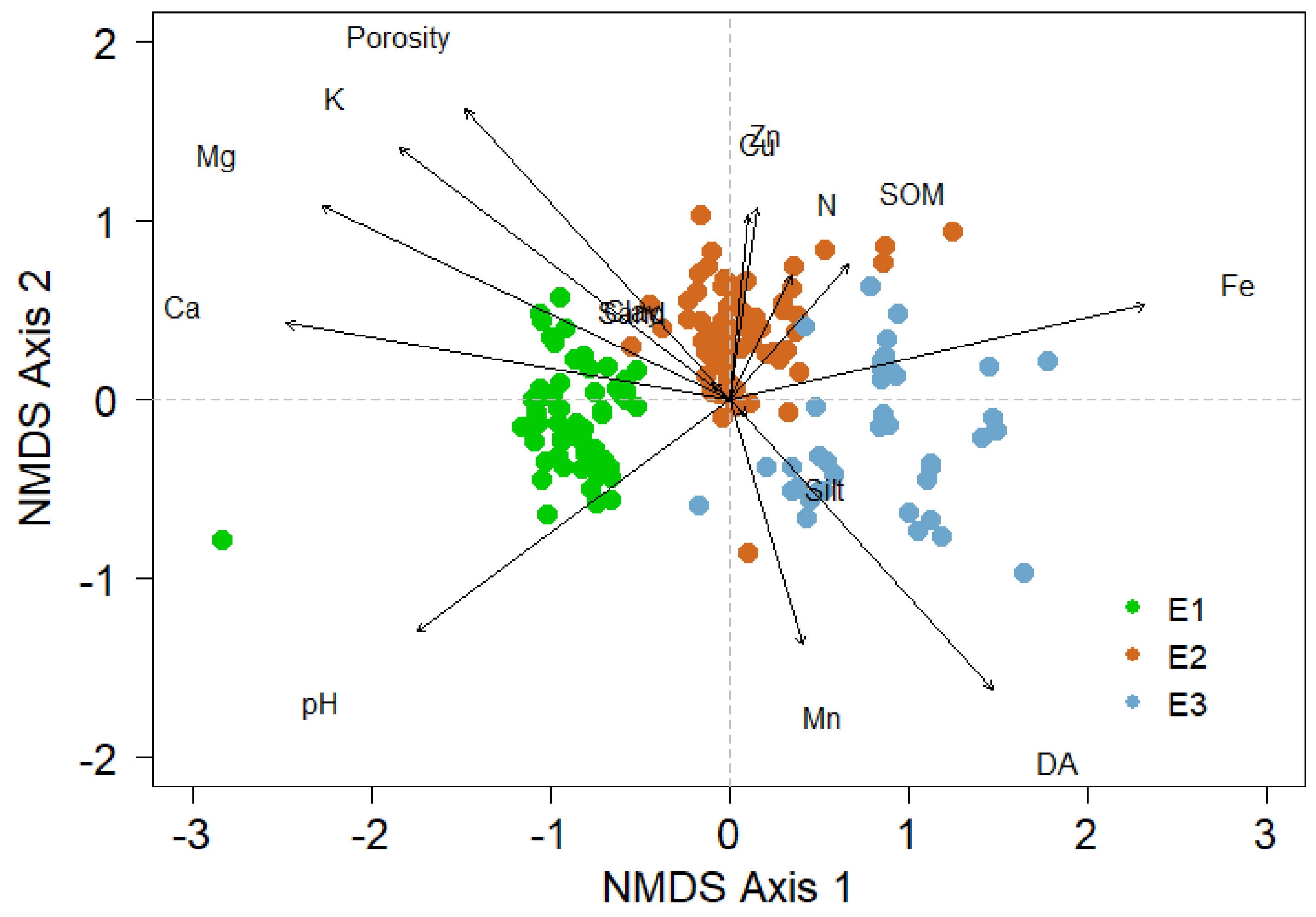
3.3. Beta-Diversity
4. Discussion
4.1. Alpha and Beta Diversity of BSC at Three Elevations
4.2. Relationships between Diversity of BSC and Soil Properties
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Cisneros, O.; Tinitana, F.; Cumbicus, N.; Prina, A. Vascular flora of dry scrub in Southern Ecuador. Cedamaz 2022, 12, 94–110. [Google Scholar] [CrossRef]
- Abad, K.; Gusmán-Montalván, E.; Ramón, P.; Burneo, J.I.; Quichimbo, P.; Jiménez, L. Edaphic properties under Vachellia macracantha in an elevation gradient of dry scrub in southern Ecuador. J. Arid. Environ. 2023, 210, 104878. [Google Scholar] [CrossRef]
- Tapia-Armijos, M.F.; Homeier, J.; Espinosa, C.I.; Leuschner, C.; de la Cruz, M. Deforestation and Forest Fragmentation in South Ecuador since the 1970s—Losing a Hotspot of Biodiversity. PLoS ONE 2015, 10, e0133701. [Google Scholar] [CrossRef] [Green Version]
- Sierra, R. Propuesta Preliminar de un Sistema de Clasificación de Vegetación para el Ecuador Continental; Proyecto INEFAN/GEF-BIRG y EcoCiencia: Quito, Ecuador, 1999; p. 194. [Google Scholar] [CrossRef]
- De la Torre, L.; Navarrete, H.; Muriel, M.P.; Macía, M.J.; Balslev, H. Diversidad de ecosistemas en Ecuador. In Enciclopedia de las Plantas Útiles del Ecuador, 1st ed.; De la Torre, L., Navarrete, H., Muriel, M.P., Macía, M.J., Balslev, H., Eds.; Herbario QC, Herbario AAU: Quito, Ecuador; Aarhus, Denmark, 2008; pp. 28–38. Available online: https://www.puce.edu.ec/portal/wr-resource/blobs/1/PUB-QCA-PUCE-2008-Enciclopedia.pdf (accessed on 1 January 2022).
- Carrión-Paladines, V.; Hinojosa, M.B.; Álvarez, L.J.; Reyes-Bueno, F.; Quezada, L.C.; García-Ruiz, R. Effects of the Severity of Wildfires on Some Physical-Chemical Soil Properties in a Humid Montane Scrublands Ecosystem in Southern Ecuador. Fire 2022, 5, 66. [Google Scholar] [CrossRef]
- Fries, A.; Silva, K.; Pucha-Cofrep, F.; Oñate-Valdivieso, F.; Ochoa-Cueva, P. Water Balance and Soil Moisture Deficit of Different Vegetation Units under Semiarid Conditions in the Andes of Southern Ecuador. Climate 2020, 8, 30. [Google Scholar] [CrossRef] [Green Version]
- Chamizo, S.; Cantón, Y.; Lázaro, R.; Solé-Benet, A.; Domingo, F. Crust Composition and Disturbance Drive Infiltration through Biological Soil Crusts in Semiarid Ecosystems. Ecosystems 2011, 15, 148–161. [Google Scholar] [CrossRef]
- Belnap, J.; Gillette, D.A. Vulnerability of desert biological soil crusts to wind erosion: The influences of crust development, soil texture, and disturbance. J. Arid. Environ. 1998, 39, 133–142. [Google Scholar] [CrossRef]
- Romero, A.N.; Moratta, M.H.; Vento, B.; Rodriguez, R.; Carretero, E.M. Variations in the coverage of biological soil crusts along an aridity gradient in the central-west Argentina. Acta Oecol. 2020, 109, 103671. [Google Scholar] [CrossRef]
- Castillo-Monroy, A.P.; Maestre, F.T. La costra biológica del suelo: Avances recientes en el conocimiento de su estructura y función ecológica. Rev. Chil. Hist. Nat. 2011, 84, 1–21. [Google Scholar] [CrossRef]
- Elbert, W.; Weber, B.; Burrows, S.; Steinkamp, J.; Büdel, B.; Andreae, M.O.; Pöschl, U. Contribution of cryptogamic covers to the global cycles of carbon and nitrogen. Nat. Geosci. 2012, 5, 459–462. [Google Scholar] [CrossRef]
- Belnap, J.; Phillips, S.L.; Miller, M.E. Response of desert biological soil crusts to alterations in precipitation frequency. Oecologia 2003, 141, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Monroy, A.P.; Bowker, M.A.; Maestre, F.T.; Rodríguez-Echeverría, S.; Martinez, I.; Barraza-Zepeda, C.E.; Escolar, C. Relationships between biological soil crusts, bacterial diversity and abundance, and ecosystem functioning: Insights from a semi-arid Mediterranean environment. J. Veg. Sci. 2011, 22, 165–174. [Google Scholar] [CrossRef]
- Toledo, V.; de Andreu, A.F. Evaluación de las propiedades biológicas y bioquímicas de la costra microbiótica de un suelo bajo vegetación natural en la región árida de Quíbor, Venezuela. Rev. Investig. 2012, 36, 143–162. [Google Scholar]
- Castillo-Monroy, A.P.; Benítez, Á.; Reyes-Bueno, F.; Donoso, D.A.; Cueva, A. Biocrust structure responds to soil variables along a tropical scrubland elevation gradient. J. Arid. Environ. 2016, 124, 31–38. [Google Scholar] [CrossRef]
- Bowker, M.A.; Belnap, J.; Davidson, D.W.; Goldstein, H. Correlates of biological soil crust abundance across a continuum of spatial scales: Support for a hierarchical conceptual model. J. Appl. Ecol. 2006, 43, 152–163. [Google Scholar] [CrossRef]
- Maestre, F.T.; Escudero, A.; Martinez, I.; Guerrero, C.; Rubio, A. Does spatial pattern matter to ecosystem functioning? Insights from biological soil crusts. Funct. Ecol. 2005, 19, 566–573. [Google Scholar] [CrossRef]
- Castillo-Monroy, A.P.; Benítez, Á. Patrones de abundancia y riqueza de componentes de la costra biológica del suelo en un matorral seco del sur de Ecuador. ACI 2015, 1, 88–97. [Google Scholar] [CrossRef] [Green Version]
- Novakovskaya, I.V.; Patova, E.N.; Dubrovskiy, Y.A.; Novakovskiy, A.B.; Kulyugina, E.E. Distribution of algae and cyanobacteria of biological soil crusts along the elevation gradient in mountain plant communities at the Northern Urals (Russian European Northeast). J. Mt. Sci. 2022, 19, 637–646. [Google Scholar] [CrossRef]
- Bruun, H.H.; Moen, J.; Virtanen, R.; Grytnes, J.; Oksanen, L.; Angerbjörn, A. Effects of altitude and topography on species richness of vascular plants, bryophytes and lichens in alpine communities. J. Veg. Sci. 2006, 17, 37–46. [Google Scholar] [CrossRef]
- Grytnes, J.A.; Heegaard, E.; Ihlen, P.G. Species richness of vascular plants, bryophytes, and lichens along an altitudinal gradient in western Norway. Acta Oecol. 2006, 29, 241–246. [Google Scholar] [CrossRef]
- Belnap, J.; Eldridge, D. Disturbance and Recovery of Biological Soil Crusts. In Biological Soil Crusts: Structure, Function, and Management; Belnap, J., Lange, O.L., Eds.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2001; Volume 150, pp. 363–364. [Google Scholar]
- Fischer, T.; Subbotina, M. Climatic and soil texture threshold values for cryptogamic cover development: A meta analysis. Biologia 2014, 69, 1520–1530. [Google Scholar] [CrossRef]
- Guzmán, P.; Benítez, Á.; Carrión-Paladines, V.; Salinas, P.; Cumbicus, N. Elevation and Soil Properties Determine Community Composition, but Not Vascular Plant Richness in Tropical Andean Roadside. Forests 2022, 13, 685. [Google Scholar] [CrossRef]
- Zhang, B.; Zhou, X.; Zhang, Y. Responses of microbial activities and soil physical-chemical properties to the successional process of biological soil crusts in the Gurbantunggut Desert, Xinjiang. J. Arid. Land 2014, 7, 101–109. [Google Scholar] [CrossRef]
- Choudhary, K.; Singh, R. Cyanobacterial diversity along altitudinal gradient in Eastern Himalayas of India. J. Algal Biomass Util. 2013, 4, 53–58. [Google Scholar]
- Sun, H.; Wu, Y.; Yu, D.; Zhou, J. Altitudinal Gradient of Microbial Biomass Phosphorus and Its Relationship with Microbial Biomass Carbon, Nitrogen, and Rhizosphere Soil Phosphorus on the Eastern Slope of Gongga Mountain, SW China. PLoS ONE 2013, 8, e72952. [Google Scholar] [CrossRef]
- Belnap, J.; Prasse, R.; Harper, K.T. Influence of Biological Soil Crusts on Soil Environments and Vascular Plants. In Biological Soil Crusts: Structure, Function, and Management; Belnap, J., Lange, O.L., Eds.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2001; Volume 150, pp. 281–300. [Google Scholar] [CrossRef]
- Belnap, J.; Lange, O.L. Biological Soil Crusts: Structure, Function, and Management, 2nd ed.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2003; pp. 3–471. [Google Scholar]
- Eldridge, D.J.; Bowker, M.A.; Maestre, F.T.; Alonso, P.; Mau, R.L.; Papadopoulos, J.; Escudero, A. Interactive Effects of Three Ecosystem Engineers on Infiltration in a Semi-Arid Mediterranean Grassland. Ecosystems 2010, 13, 499–510. [Google Scholar] [CrossRef]
- Maestre, F.; Bowker, M.; Cantón, Y.; Castillo-Monroy, A.; Cortina, J.; Escolar, C.; Escudero, A.; Lázaro, R.; Martínez, I. Ecology and functional roles of biological soil crusts in semi-arid ecosystems of Spain. J. Arid. Environ. 2011, 75, 1282–1291. [Google Scholar] [CrossRef] [Green Version]
- Pointing, S.B.; Belnap, J. Microbial colonization and controls in dryland systems. Nat. Rev. Microbiol. 2012, 10, 551–562. [Google Scholar] [CrossRef]
- Büdel, B.; Darienko, T.; Deutschewitz, K.; Dojani, S.; Friedl, T.; Mohr, K.I.; Salisch, M.; Reisser, W.; Weber, B. Southern African Biological Soil Crusts are Ubiquitous and Highly Diverse in Drylands, Being Restricted by Rainfall Frequency. Microb. Ecol. 2008, 57, 229–247. [Google Scholar] [CrossRef]
- Rivera-Aguilar, V.; Montejano, G.; Rodríguez-Zaragoza, S.; Durán-Díaz, A. Distribution and composition of cyanobacteria, mosses and lichens of the biological soil crusts of the Tehuacán Valley, Puebla, México. J. Arid. Environ. 2006, 67, 208–225. [Google Scholar] [CrossRef]
- Baumann, K.; Jung, P.; Samolov, E.; Lehnert, L.W.; Büdel, B.; Karsten, U.; Bendix, J.; Achilles, S.; Schermer, M.; Matus, F.; et al. Biological soil crusts along a climatic gradient in Chile: Richness and imprints of phototrophic microorganisms in phosphorus biogeochemical cycling. Soil Biol. Biochem. 2018, 127, 286–300. [Google Scholar] [CrossRef]
- Machado-De-Lima, N.M.; Fernandes, V.M.C.; Roush, D.; Ayuso, S.V.; Rigonato, J.; Garcia-Pichel, F.; Branco, L.H.Z. The Compositionally Distinct Cyanobacterial Biocrusts from Brazilian Savanna and Their Environmental Drivers of Community Diversity. Front. Microbiol. 2019, 10, 2798. [Google Scholar] [CrossRef]
- Armijos, J.M. Oña y Su Parroquia Susudel: Patrimonio Cultural de la Nación. Pregrade Thesis, Universidad de Cuenca, Cuenca, Ecuador, 2015. [Google Scholar]
- Ullauri, G. Plan de Desarrollo y Ordenamiento Territorial. Available online: http://app.sni.gob.ec/sni-link/sni/PORTAL_SNI/data_sigad_plus/sigadplusdocumentofinal/0160001830001_PDYOT%20San%20Felipe%20de%20O%C3%B1a_15-03-2015_22-45-23.pdf (accessed on 28 November 2022).
- Munkholm, L.J.; Schjønning, P.; Kay, B.D. Tensile strength of soil cores in relation to aggregate strength, soil fragmentation and pore characteristics. Soil Tillage Res. 2000, 64, 125–135. [Google Scholar] [CrossRef]
- Sandoval, M.; Fernández, J.; Seguel, O.; Becerra, J.; Salazar, D. Métodos de Análisis Físicos de Suelos. Ph.D. Thesis, Universidad de Concepción, Concepción, Chile, 2011. [Google Scholar]
- Guo, M.; Wang, W.; Kang, H.; Yang, B. Changes in soil properties and erodibility of gully heads induced by vegetation restoration on the Loess Plateau, China. J. Arid. Land 2018, 10, 712–725. [Google Scholar] [CrossRef] [Green Version]
- Black, C.A.; Evans, D.D.; White, J.L.; Ensminger, L.E.; Clark, F.E. Methods of Soil Analysis. In Part 2. Chemical and Microbiological Properties; Agronomy 9; ASA: New York, NY, USA, 1965; p. 1572. [Google Scholar]
- Page, A.L.; Miller, R.H.; Keeney, D.R. Methods of Soil Analysis; ASA: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- Bremner, J.M. Nitrogen-Total. In Methods of Soil Analysis. In Part 3 Chemical Methods; John Wiley & Sons: Hoboken, NJ, USA, 1996; Volume 5, pp. 1085–1121. [Google Scholar]
- Tan, D.; Jin, J.; Jiang, L.; Huang, S.; Liu, Z. Potassium assessment of grain producing soils in North China. Agric. Ecosyst. Environ. 2012, 148, 65–71. [Google Scholar] [CrossRef]
- Magurran, A.E. Species abundance distributions: Pattern or process? Funct. Ecol. 2005, 19, 177–181. [Google Scholar] [CrossRef]
- McCullagh, P.; Nelder, J.A. Generalized Linear Models, 2nd ed.; CRC Press, Chapman and Hall: Boca Raton, FL, USA, 1989; p. 506. [Google Scholar]
- Tikhonov, G.; Opedal, Ø.H.; Abrego, N.; Lehikoinen, A.; de Jonge, M.M.; Oksanen, J.; Ovaskainen, O. Joint species distribution modelling with the r -package H msc. Methods Ecol. Evol. 2019, 11, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Baniya, C.B.; Solhøy, T.; Gauslaa, Y.; Palmer, M.W. Richness and Composition of Vascular Plants and Cryptogams along a High Elevational Gradient on Buddha Mountain, Central Tibet. Folia Geobot. 2012, 47, 135–151. [Google Scholar] [CrossRef]
- Rashmi, S.; Rajkumar, H.G. Diversity of Lichens along Elevational Gradients in Forest Ranges of Chamarajanagar District, Karnataka State. Int. J. Sci. Res. Biol. Sci. 2019, 6, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Grau, O.; Grytnes, J.-A.; Birks, H.J.B. A comparison of altitudinal species richness patterns of bryophytes with other plant groups in Nepal, Central Himalaya. J. Biogeogr. 2007, 34, 1907–1915. [Google Scholar] [CrossRef]
- You, Y.; Aho, K.; Lohse, K.A.; Schwabedissen, S.G.; Ledbetter, R.N.; Magnuson, T.S. Biological Soil Crust Bacterial Communities Vary along Climatic and Shrub Cover Gradients within a Sagebrush Steppe Ecosystem. Front. Microbiol. 2021, 12, 569791. [Google Scholar] [CrossRef] [PubMed]
- González, Y.; Aragón, G.; Benítez, A.; Prieto, M. Changes in soil cryptogamic communities in tropical Ecuadorean páramos. Community Ecol. 2017, 18, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Concostrina-Zubiri, L.; Martinez, I.; Huber-Sannwald, E.; Escudero, A. Efectos y respuestas de la Costra Biológica del Suelo en ecosistemas áridos: Avances recientes a nivel de especie. Ecosistemas 2013, 22, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Blay, E.S.; Schwabedissen, S.G.; Magnuson, T.S.; Aho, K.A.; Sheridan, P.P.; Lohse, K.A. Variation in Biological Soil Crust Bacterial Abundance and Diversity as a Function of Climate in Cold Steppe Ecosystems in the Intermountain West, USA. Microb. Ecol. 2017, 74, 691–700. [Google Scholar] [CrossRef]
- Videla, M.E.C.; Taboada, M.D.L.; Aranibar, J.N. Diversidad de cianobacterias en costras biológicas de suelo de la ecorregión del Monte Central (Mendoza, Argentina). Lilloa 2018, 55, 30–46. [Google Scholar] [CrossRef] [Green Version]
- Williams, L.; Borchhardt, N.; Colesie, C.; Baum, C.; Komsic-Buchmann, K.; Rippin, M.; Becker, B.; Karsten, U.; Büdel, B. Biological soil crusts of Arctic Svalbard and of Livingston Island, Antarctica. Polar Biol. 2016, 40, 399–411. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, D.; Schaefer, C.E.R.; Putzke, J.; Francelino, M.R.; Ferrari, F.R.; Corrêa, G.R.; Villa, P.M. How does the pedoenvironmental gradient shape non-vascular species assemblages and community structures in Maritime Antarctica? Ecol. Indic. 2019, 108, 105726. [Google Scholar] [CrossRef]
- Kalníková, V.; Palpurina, S.; Peterka, T.; Kubešová, S.; Plesková, Z.; Sabovljević, M. Bryophytes on River Gravel Bars in the Balkan Mountains: New Records and Insights into Ecology. Herzogia 2017, 30, 370–386. [Google Scholar] [CrossRef]
- Hansen, C.J.; Goertzen, L.R. Psora icterica (Lecanorales, Psoraceae), una nueva e interesante disyunción de Alabama. Opusc. Philolichenum 2012, 11, 49–51. [Google Scholar]
- Kakeh, J.; Gorji, M.; Sohrabi, M.; Tavili, A.; Pourbabaee, A.A. Effects of biological soil crusts on some physicochemical characteristics of rangeland soils of Alagol, Turkmen Sahra, NE Iran. Soil Tillage Res. 2018, 181, 152–159. [Google Scholar] [CrossRef]
- Verrecchia, E.; Yair, A.; Kidron, G.J.; Verrecchia, K. Physical properties of the psammophile cryptogamic crust and their consequences to the water regime of sandy soils, north-western Negev Desert, Israel. J. Arid. Environ. 1995, 29, 427–437. [Google Scholar] [CrossRef]
- Bowker, M.A.; Johnson, N.C.; Belnap, J.; Koch, G.W. Short-term monitoring of aridland lichen cover and biomass using photography and fatty acids. J. Arid. Environ. 2008, 72, 869–878. [Google Scholar] [CrossRef]
- Murphy, B.W. Impact of soil organic matter on soil properties—A review with emphasis on Australian soils. Soil Res. 2015, 53, 605–635. [Google Scholar] [CrossRef]
- Aishah, A.W.; Zauyah, S.; Anuar, A.R.; Fauziah, C.I. Spatial variability of selected chemical characteristics of paddy soils in Sawah Sempadan, Selangor, Malaysia. Malaysian. J. Soil Sci. 2010, 14, 27–39. [Google Scholar]
- Musinguzi, P.; Ebanyat, P.; Tenywa, J.S.; Basamba, T.A.; Tenywa, M.M.; Mubiru, D.N. Critical soil organic carbon range for optimal crop response to mineral fertiliser nitrogen on a ferralsol. Exp. Agric. 2016, 52, 635–653. [Google Scholar] [CrossRef]
- Bowker, M.A.; Belnap, J.; Davidson, D.W.; Phillips, S.L. Evidence for micronutrient limitation of biological soil crusts: Importance to arid-lands restoration. Ecol. Appl. 2005, 15, 1941–1951. [Google Scholar] [CrossRef] [Green Version]
- Bowker, M.A.; Belnap, J.; Büdel, B.; Sannier, C.; Pietrasiak, N.; Eldridge, D.J.; Rivera-Aguilar, V. Controls on distribution patterns of biological soil crusts at micro- to global scales. In Biological Soil Crusts: An Organizing Principle in Drylands; Weber, B., Büdel, B., Belnap, J., Eds.; Ecological Studies; Springer: Cham, Switzerland, 2016; Volume 150, pp. 173–197. [Google Scholar] [CrossRef]
- Ochoa-Hueso, R.; Hernandez, R.R.; Pueyo, J.J.; Manrique, E. Spatial distribution and physiology of biological soil crusts from semi-arid central Spain are related to soil chemistry and shrub cover. Soil Biol. Biochem. 2011, 43, 1894–1901. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | E1 (2100 m.a.s.l.) | E2 (2300 m.a.s.l.) | E3 (2500 m.a.s.l.) |
|---|---|---|---|
| Mosses | |||
| Bryum argenteum Hedw. | 30 | 4 | 7 |
| Bryum sp. | 0 | 3 | 0 |
| Campylopus richardii Brid. | 0 | 0 | 26 |
| Campylopus sp. | 0 | 3 | 7 |
| Leptodontium viticulosoides (P.Beauv.) Wijk & Margad. | 0 | 4 | 35 |
| Syntrichia andicola (Mont.) Ochyra | 1 | 0 | 0 |
| Syntrichia sp. | 55 | 53 | 25 |
| Weissia sp. | 21 | 55 | 20 |
| Liverworts | |||
| Fossombronia peruviana (Gottsche & Hampe) | 0 | 1 | 4 |
| Gymnomitrion aff. bolivianum (Steph.) Váňa | 0 | 0 | 1 |
| Cyanobacteria | |||
| Nostoc commune Vaucher ex Bornet & Flahault | 42 | 0 | 0 |
| Scytonema sp. | 58 | 36 | 0 |
| Lichens | |||
| Bacidia sp. | 1 | 0 | 0 |
| Buellia sp. | 2 | 1 | 0 |
| Caloplaca sp. | 0 | 3 | 0 |
| Cladia aggregata (Sw.) Nyl. | 0 | 9 | 24 |
| Cladonia sp. | 0 | 6 | 10 |
| Cladonia sp1 | 0 | 25 | 17 |
| Cladonia rappii A.Evans | 0 | 17 | 7 |
| Coccocarpia palmicola (Spreng.) Arv. & D.J.Galloway | 0 | 26 | 8 |
| Cora glabrata (Spreng.) Fr. | 0 | 8 | 2 |
| Diploschistes diacapsis (Ach.) Lumbsch | 0 | 47 | 4 |
| Flakea sp. | 0 | 46 | 58 |
| Lepraria aff. diffusa (J.R. Laundon) Kukwa | 2 | 45 | 11 |
| Lepraria sp. | 14 | 9 | 1 |
| Leptogium sp. | 15 | 7 | 2 |
| Peltula obscurans var obscurans Wetm. | 1 | 0 | 0 |
| Placidium squamulosum (Ach.) Breuss | 28 | 15 | 0 |
| Psora aff. pruinosa Timdal | 0 | 29 | 5 |
| Psora icterica (Mont.) Müll.Arg. | 9 | 3 | 4 |
| Toninia sp. | 3 | 11 | 0 |
| Toninia aff. submexicana B.de Lesd. | 17 | 1 | 1 |
| Toninia wetmorei Timdal | 16 | 0 | 4 |
| Xanthoparmelia subplittii Hale | 14 | 2 | 0 |
| Xanthoparmelia mougeotii (Schaer.) Hale | 52 | 19 | 3 |
| Properties | E1 (m.a.s.l. 2100) | E2 (m.a.s.l. 2300) | E3 (m.a.s.l. 2500) | p-Value |
|---|---|---|---|---|
| Bulk density (g cm−3) | 0.87 ± 0.02 | 0.91 ± 0.03 | 1.05 ± 0.13 | 0.061 |
| Porosity (%) | 67.2 ± 0.75 | 65.7 ± 1.00 | 60.4 ± 4.86 | 0.062 |
| Sand (%) | 59.9 ± 5.77 | 62.1 ± 1.91 | 62.6 ± 0.42 | 0.632 |
| Silt (%) | 14.0 ± 3.46 | 12.7 ± 1.15 | 12.4 ± 0.26 | 0.633 |
| Clay (%) | 26.1 ± 2.31 | 25.2 ± 0.75 | 25.0 ± 0.15 | 0.632 |
| Textural class | Sandy clay loam | Sandy loam | Sandy clay loam | -- |
| pH | 7.3 ± 0.2 | 5.7 ± 0.2 | 6.1 ± 0.2 | 0.000 |
| SOM (%) | 1.6 ± 0.1 | 1.8 ± 0.1 | 1.9 ± 0.6 | 0.592 |
| N (%) | 0.09 ± 0.01 | 0.09 ± 0.01 | 0.10 ± 0.03 | 0.798 |
| P (mg/kg) | 3.5 ± 0.00 | 3.5 ± 0.00 | 3.5 ± 0.00 | 0.100 |
| K (mg/kg) | 0.49 ± 0.08 | 0.42 ± 0.08 | 0.24 ± 0.09 | 0.025 |
| SOC (%) | 0.94 ± 0.05 | 1.04 ± 0.06 | 1.11 ± 0.34 | 0.602 |
| C/N ratio | 10.8 ± 0.4 | 11.1 ± 0.00 | 11.5 ± 0.4 | 0.091 |
| Mn (mg/kg) | 4.4 ± 0.37 | 2.9 ± 1.27 | 5.5 ± 1.13 | 0.053 |
| Mg (cmol/kg) | 2.7 ± 0.12 | 1.8 ± 0.39 | 0.9 ± 0.29 | 0.000 |
| Ca (cmol/kg) | 10.6 ± 0.94 | 6.3 ± 0.42 | 4.2 ± 1.66 | 0.001 |
| Fe (mg/kg) | 24.8 ± 5.30 | 81.9 ± 11.25 | 87.3 ± 8.00 | 0.000 |
| Cu (mg/kg) | 1.8 ± 0.07 | 3.4 ± 3.57 | 1.7 ± 0.79 | 0.571 |
| Zn (mg/kg) | 1.6 ± 0.00 | 2.0 ± 0.66 | 1.6 ± 0.00 | 0.421 |
| Richness | ||||
|---|---|---|---|---|
| Estimate | Coefficient | Std. Error | T | p-Value |
| E1 | −21.1277 | 5.6888 | −3.7140 | 0.0003 |
| E2 | 3.4233 | 0.8211 | 4.1690 | 0.0000 |
| E3 | 2.4856 | 0.7452 | 3.3350 | 0.0010 |
| pH | 3.6763 | 0.9122 | 4.0300 | 0.0001 |
| DA | −5.0010 | 1.3484 | −3.7090 | 0.0003 |
| K | 2.8071 | 0.7409 | 3.7890 | 0.0002 |
| Fe | 0.0624 | 0.0157 | 3.9650 | 0.0001 |
| SOM | −1.6713 | 0.4026 | −4.1510 | 0.0001 |
| Cover | Coefficient | Std. Error | T | p-Value |
| E1 | −23.9494 | 9.1420 | −2.6200 | 0.0096 |
| E2 | 3.4135 | 1.3150 | 2.5960 | 0.0103 |
| E3 | 2.3121 | 1.2073 | 1.9150 | 0.0571 |
| pH | 4.6722 | 1.4855 | 3.1450 | 0.0020 |
| DA | −7.2094 | 2.3819 | −3.0270 | 0.0029 |
| K | 3.2673 | 1.2429 | 2.6290 | 0.0093 |
| Fe | 0.0907 | 0.0262 | 3.4660 | 0.0007 |
| SOM | −2.0850 | 0.6731 | −3.0980 | 0.0023 |
| Factors | df | SS | MS | F | R2 | p-Value |
|---|---|---|---|---|---|---|
| Elevation | 2 | 21.677 | 10.8385 | 65.305 | 0.38918 | 0.001 |
| Bulk density | 1 | 1.724 | 1.7244 | 10.39 | 0.03096 | 0.001 |
| K | 1 | 1.236 | 1.2359 | 7.446 | 0.02219 | 0.001 |
| Fe | 1 | 1.233 | 1.2335 | 7.432 | 0.02215 | 0.001 |
| pH | 1 | 0.574 | 0.574 | 3.459 | 0.01031 | 0.006 |
| SOM | 1 | 0.397 | 0.3966 | 2.39 | 0.00712 | 0.025 |
| Residuals | 171 | 28.38 | 0.166 | 0.50953 | ||
| Total | 179 | 55.699 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz, L.; Carrión-Paladines, V.; Vega, M.; López, F.; Benítez, Á. Biological Crust Diversity Related to Elevation and Soil Properties at Local Scale in a Montane Scrub of Ecuador. J. Fungi 2023, 9, 386. https://doi.org/10.3390/jof9030386
Ruiz L, Carrión-Paladines V, Vega M, López F, Benítez Á. Biological Crust Diversity Related to Elevation and Soil Properties at Local Scale in a Montane Scrub of Ecuador. Journal of Fungi. 2023; 9(3):386. https://doi.org/10.3390/jof9030386
Chicago/Turabian StyleRuiz, Leslye, Vinicio Carrión-Paladines, Marlon Vega, Fausto López, and Ángel Benítez. 2023. "Biological Crust Diversity Related to Elevation and Soil Properties at Local Scale in a Montane Scrub of Ecuador" Journal of Fungi 9, no. 3: 386. https://doi.org/10.3390/jof9030386