
Mebendazole Inhibits Histoplasma capsulatum In Vitro Growth and Decreases Mitochondrion and Cytoskeleton Protein Levels
 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Growth Conditions
2.2. Screening of the NIH Clinical Collection
2.3. Minimal Inhibitory Concentration (MIC) Determinations
2.4. Fungicidal Activity
2.5. Preparation of Protein Extracts
2.6. Sample Processing for Shotgun Proteomics
2.7. Mass Spectrometry Analysis
2.8. Protein Identification and Quantitation following Mass Spectrometry
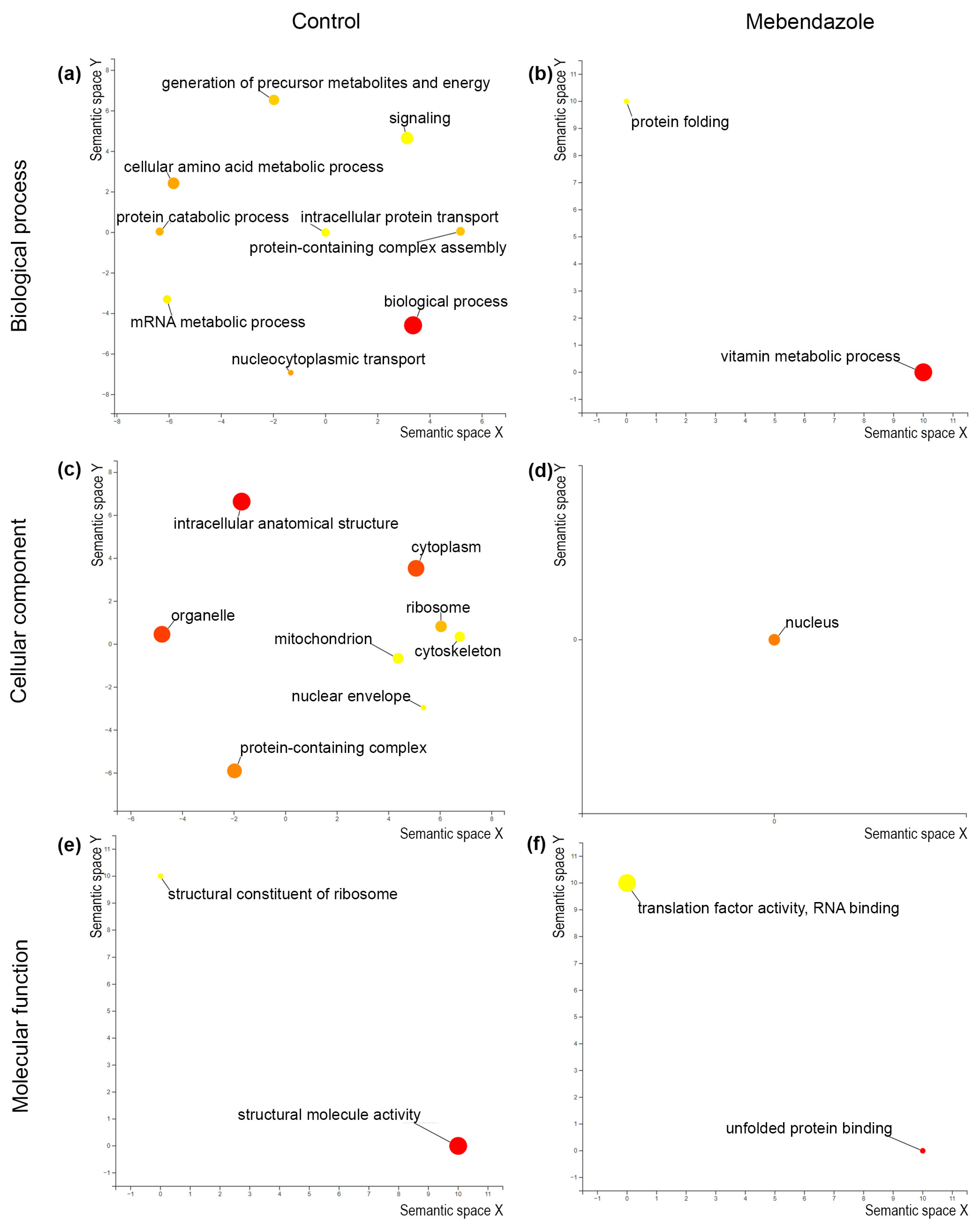
2.9. Bioinformatic Analyses
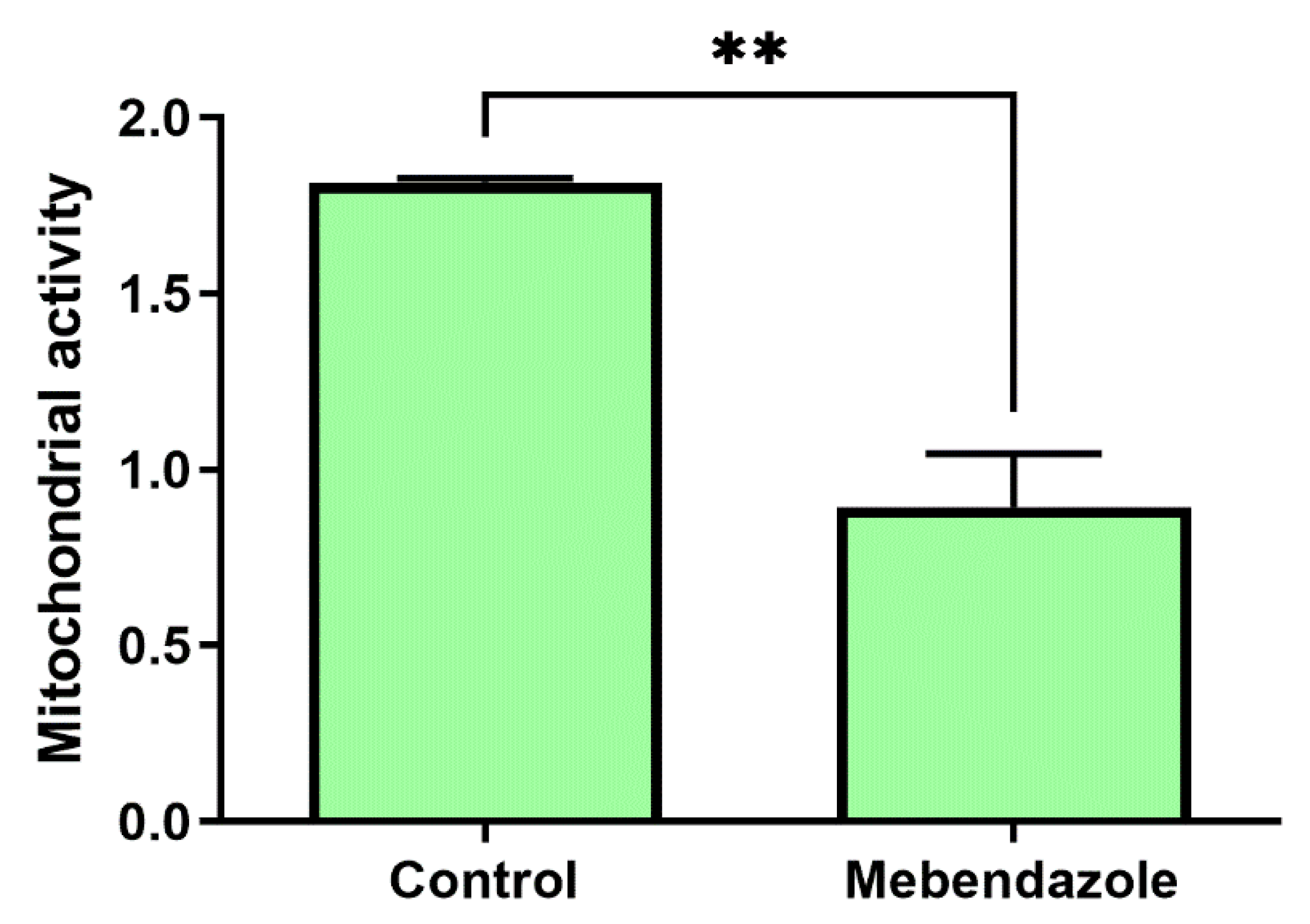
2.10. Measurement of Mitochondrial Metabolic Activity
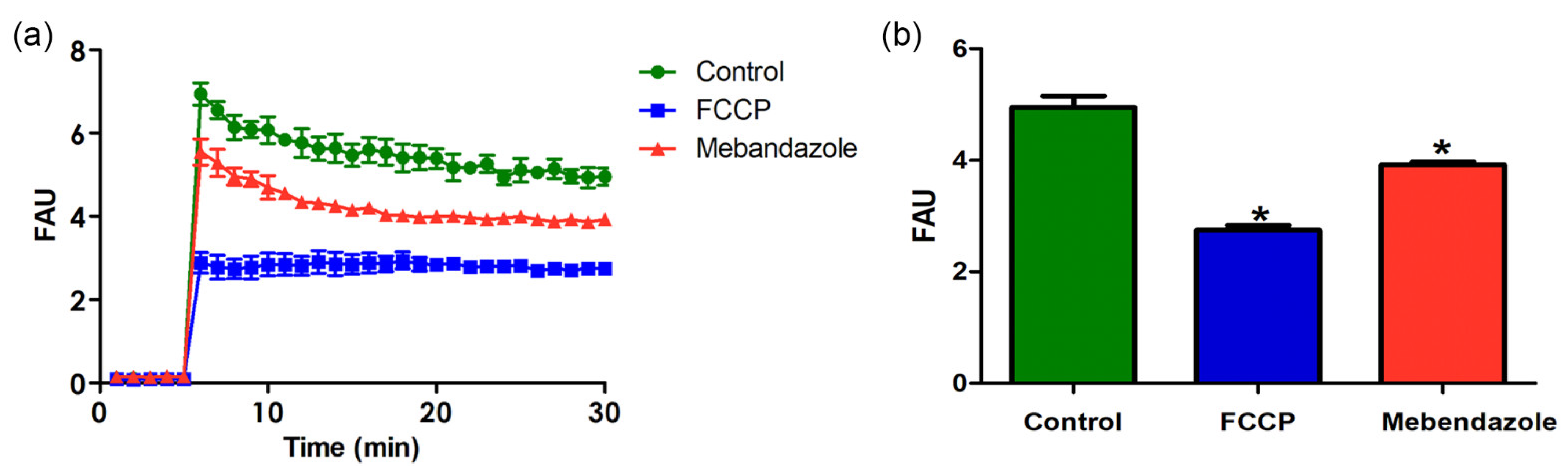
2.11. JC-1 Assay
2.12. Size Measurements
2.13. Western Blot Analysis of Cytoskeleton Proteins
2.14. Statistical Analyzes
3. Results
3.1. Drugs with Anti-H. capsulatum Activity in the NIH Clinical Collection
3.2. Mebendazole Antifungal Activity against H. capsulatum
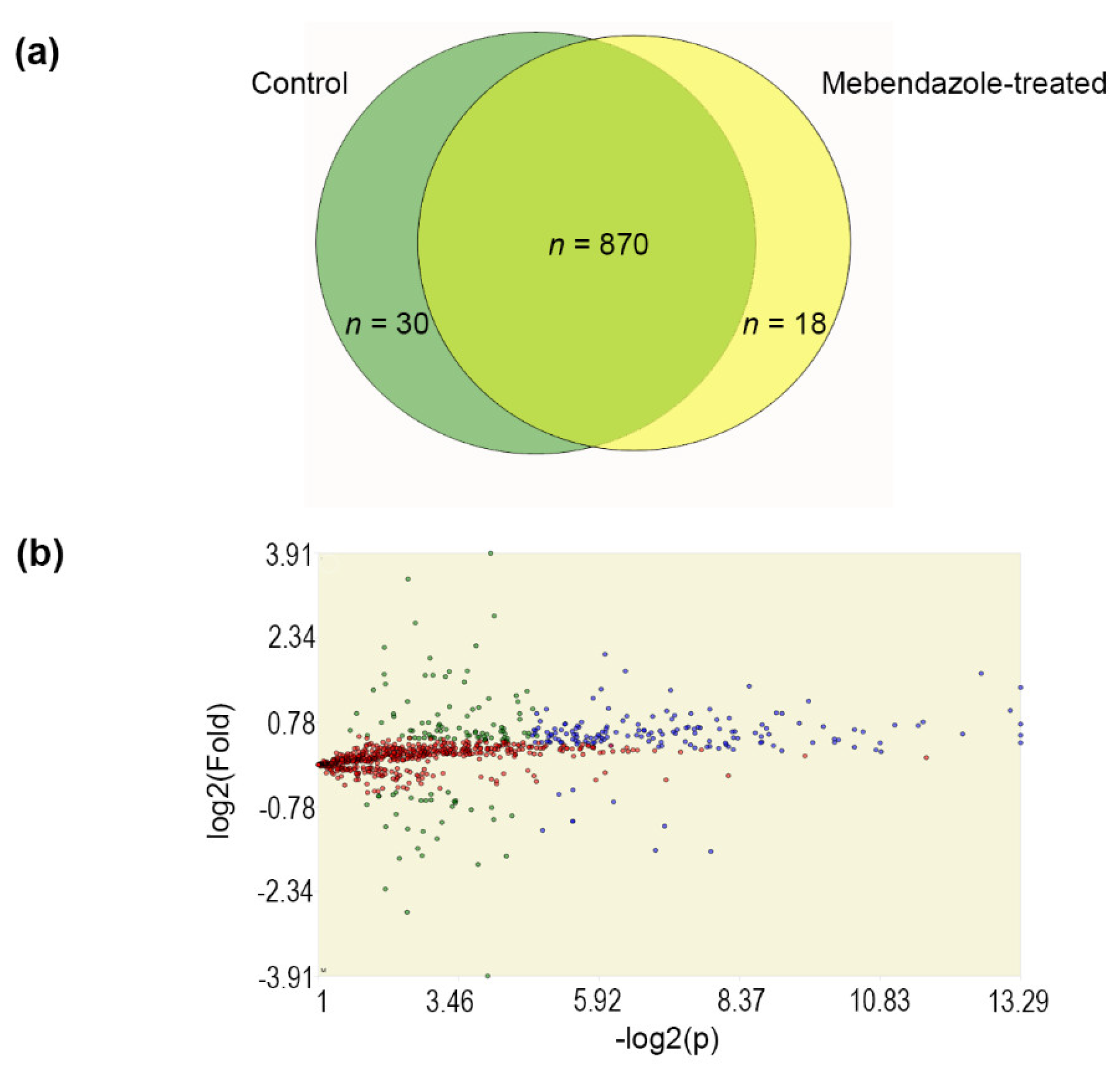
3.3. Proteomics of Mebendazole-Treated H. capsulatum
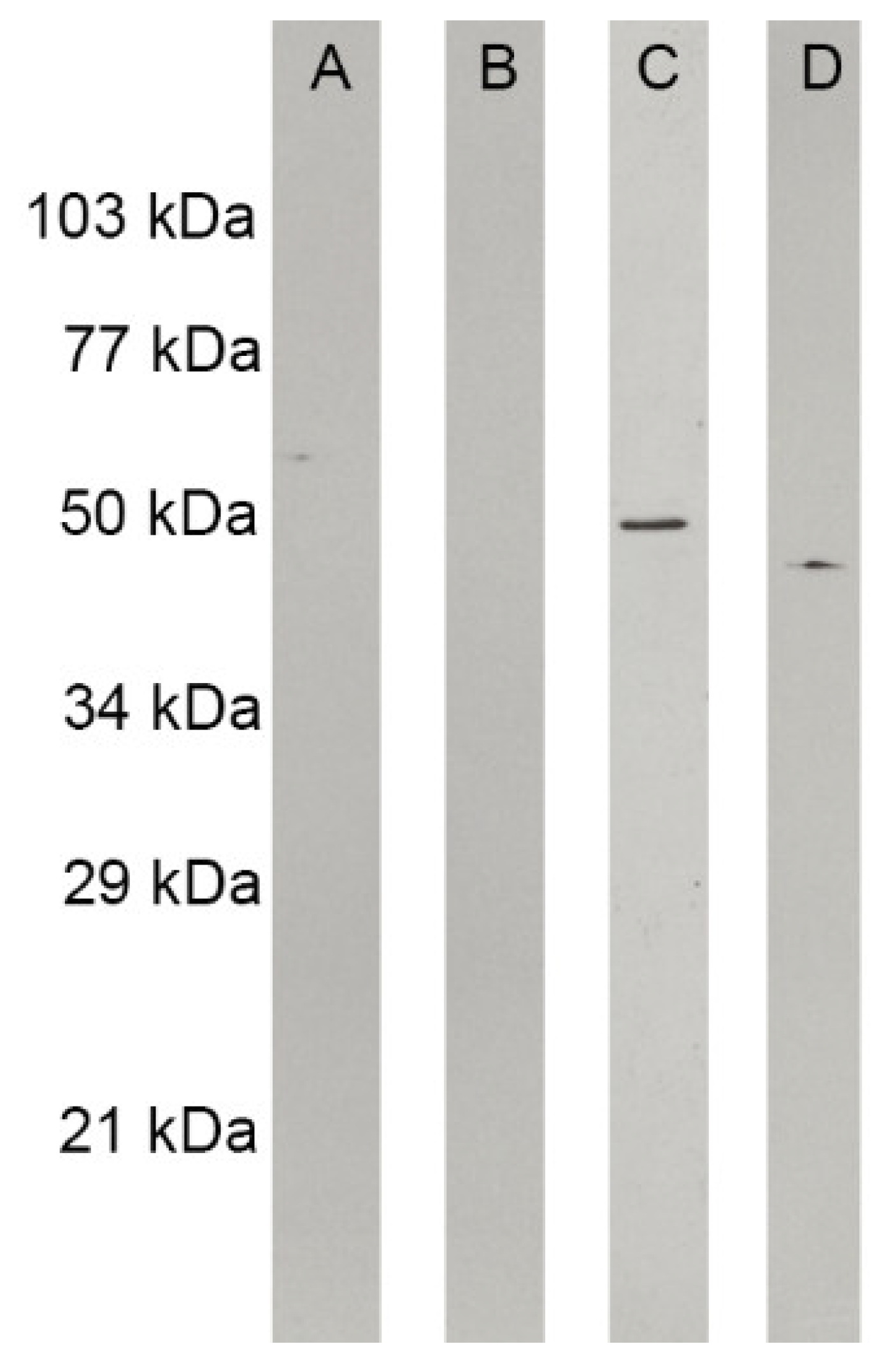
3.4. Mebendazole Affects Mitochondrion Activity and H. capsulatum Cytoskeleton Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Falci, D.R.; Monteiro, A.A.; Braz Caurio, C.F.; Magalhães, T.C.O.; Xavier, M.O.; Basso, R.P.; Melo, M.; Schwarzbold, A.V.; Ferreira, P.R.A.; Vidal, J.E.; et al. Histoplasmosis, An Underdiagnosed Disease Affecting People Living With HIV/AIDS in Brazil: Results of a Multicenter Prospective Cohort Study Using Both Classical Mycology Tests and Histoplasma Urine Antigen Detection. Open Forum Infect. Dis. 2019, 6, ofz073. [Google Scholar] [CrossRef]
- Bongomin, F.; Kwizera, R.; Denning, D.W. Getting Histoplasmosis on the Map of International Recommendations for Patients with Advanced HIV Disease. J. Fungi 2019, 5, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheat, L.J.; Connolly, P.; Smedema, M.; Durkin, M.; Brizendine, E.; Mann, P.; Patel, R.; McNicholas, P.M.; Goldman, M. Activity of Newer Triazoles against Histoplasma capsulatum from Patients with AIDS Who Failed Fluconazole. J. Antimicrob. Chemother. 2006, 57, 1235–1239. [Google Scholar] [CrossRef]
- Thompson, G.R.; Le, T.; Chindamporn, A.; Kauffman, C.A.; Alastruey-Izquierdo, A.; Ampel, N.M.; Andes, D.R.; Armstrong-James, D.; Ayanlowo, O.; Baddley, J.W.; et al. Global Guideline for the Diagnosis and Management of the Endemic Mycoses: An Initiative of the European Confederation of Medical Mycology in Cooperation with the International Society for Human and Animal Mycology. Lancet Infect. Dis. 2021, 21, e364–e374. [Google Scholar] [CrossRef]
- Perez, F.; Caceres, D.H.; Ford, N.; Ravasi, G.; Gomez, B.L.; Pasqualotto, A.C.; Hine, P.; Adenis, A.A.; Nacher, M.; Chiller, T.; et al. Summary of Guidelines for Managing Histoplasmosis among People Living with HIV. J. Fungi 2021, 7, 134. [Google Scholar] [CrossRef]
- Noor, A.; Preuss, C.V. Amphotericin B. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Schiave, L.A.; Nascimento, E.; Gaspar, G.G.; Vilar, F.C.; Martinez, E.Z.; de Gaitani, C.M.; Martinez, R. Minimum Concentration of Amphotericin B in Serum According to the Formulation, Dose, and Daily or Prolonged Intermittent Therapeutic Regimen. Rev. Soc. Bras. Med. Trop. 2020, 53, e20180463. [Google Scholar] [CrossRef] [Green Version]
- Ostermann, H.; Solano, C.; Jarque, I.; Garcia-Vidal, C.; Gao, X.; Barrueta, J.A.; De Salas-Cansado, M.; Stephens, J.; Xue, M.; Weber, B.; et al. Cost Analysis of Voriconazole versus Liposomal Amphotericin B for Primary Therapy of Invasive Aspergillosis among Patients with Haematological Disorders in Germany and Spain. BMC Pharmacol. Toxicol. 2014, 15, 52. [Google Scholar] [CrossRef] [Green Version]
- De Carvalho, J.P.; de Assis, T.M.; Simões, T.C.; Cota, G. Estimating Direct Costs of the Treatment for Mucosal Leishmaniasis in Brazil. Rev. Soc. Bras. Med. Trop. 2021, 54, e04542020. [Google Scholar] [CrossRef]
- Jaruratanasirikul, S.; Sriwiriyajan, S. Pharmacokinetic Study of the Interaction between Itraconazole and Nevirapine. Eur. J. Clin. Pharmacol. 2007, 63, 451–456. [Google Scholar] [CrossRef]
- Adenis, A.A.; Aznar, C.; Couppié, P. Histoplasmosis in HIV-Infected Patients: A Review of New Developments and Remaining Gaps. Curr. Trop. Med. Rep. 2014, 1, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Piérard, G.E.; Arrese, J.E.; Piérard-Franchimont, C. Itraconazole. Expert Opin. Pharmacother. 2000, 1, 287–304. [Google Scholar] [CrossRef]
- Wheat, L.J.; Freifeld, A.G.; Kleiman, M.B.; Baddley, J.W.; McKinsey, D.S.; Loyd, J.E.; Kauffman, C.A. Infectious Diseases Society of America Clinical Practice Guidelines for the Management of Patients with Histoplasmosis: 2007 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2007, 45, 807–825. [Google Scholar] [CrossRef] [Green Version]
- Spec, A.; Connolly, P.; Montejano, R.; Wheat, L.J. In Vitro Activity of Isavuconazole against Fluconazole-Resistant Isolates of Histoplasma capsulatum. Med. Mycol. 2018, 56, 834–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheat, L.J.; Kauffman, C.A. Histoplasmosis. Infect. Dis. Clin. N. Am. 2003, 17, 1–19, vii. [Google Scholar] [CrossRef]
- Martino, J.; Fisher, B.T.; Bosse, K.R.; Bagatell, R. Suspected Posaconazole Toxicity in a Pediatric Oncology Patient. Pediatr. Blood Cancer 2015, 62, 1682. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, M.; Abu Saleh, O.; Sohail, M.R. Hypokalemia and Hypertension Associated with Supratherapeutic Posaconazole Levels. Antimicrob. Agents Chemother. 2017, 61, e00019-17. [Google Scholar] [CrossRef] [Green Version]
- Nosengo, N. Can You Teach Old Drugs New Tricks? Nature 2016, 534, 314–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berkes, C.; Franco, J.; Lawson, M.; Brann, K.; Mermelstein, J.; Laverty, D.; Connors, A. Kinase Inhibitor Library Screening Identifies the Cancer Therapeutic Sorafenib and Structurally Similar Compounds as Strong Inhibitors of the Fungal Pathogen Histoplasma capsulatum. Antibiotics 2021, 10, 1223. [Google Scholar] [CrossRef] [PubMed]
- Vaso, C.O.; Bila, N.M.; Pandolfi, F.; De Vita, D.; Bortolami, M.; Bonatti, J.L.C.; De Moraes Silva, R.A.; Gonçalves, L.N.C.; Tudino, V.; Costi, R.; et al. Evaluation of the Anti-Histoplasma capsulatum Activity of Indole and Nitrofuran Derivatives and Their Pharmacological Safety in Three-Dimensional Cell Cultures. Pharmaceutics 2022, 14, 1043. [Google Scholar] [CrossRef]
- Krasavin, M.; Parchinsky, V.; Kantin, G.; Manicheva, O.; Dogonadze, M.; Vinogradova, T.; Karge, B.; Brönstrup, M. New Nitrofurans Amenable by Isocyanide Multicomponent Chemistry Are Active against Multidrug-Resistant and Poly-Resistant Mycobacterium Tuberculosis. Bioorg. Med. Chem. 2017, 25, 1867–1874. [Google Scholar] [CrossRef] [PubMed]
- Karimabad, M.N.; Mahmoodi, M.; Jafarzadeh, A.; Darekordi, A.; Hajizadeh, M.R.; Hassanshahi, G. Molecular Targets, Anti-Cancer Properties and Potency of Synthetic Indole-3-Carbinol Derivatives. Mini Rev. Med. Chem. 2019, 19, 540–554. [Google Scholar] [CrossRef] [PubMed]
- Nieto, M.J.; Lupton, H.K. Indole and Indoline Scaffolds in Antimicrobials: Overview, Synthesis and Recent Advances in Antimicrobial Research. Curr. Med. Chem. 2021, 28, 4828–4844. [Google Scholar] [CrossRef]
- Almeida-Silva, F.; de Melo Teixeira, M.; Matute, D.R.; de Faria Ferreira, M.; Barker, B.M.; Almeida-Paes, R.; Guimarães, A.J.; Zancopé-Oliveira, R.M. Genomic Diversity Analysis Reveals a Strong Population Structure in Histoplasma capsulatum LAmA (Histoplasma suramericanum). J. Fungi 2021, 7, 865. [Google Scholar] [CrossRef] [PubMed]
- Fressatti, R.; Dias-Siqueira, V.L.; Svidzinski, T.I.; Herrero, F.; Kemmelmeier, C. A Medium for Inducing Conversion of Histoplasma capsulatum var. Capsulatum into Its Yeast-like Form. Mem. Inst. Oswaldo Cruz 1992, 87, 53–58. [Google Scholar] [CrossRef]
- Joffe, L.S.; Schneider, R.; Lopes, W.; Azevedo, R.; Staats, C.C.; Kmetzsch, L.; Schrank, A.; Del Poeta, M.; Vainstein, M.H.; Rodrigues, M.L. The Anti-Helminthic Compound Mebendazole Has Multiple Antifungal Effects against Cryptococcus neoformans. Front. Microbiol. 2017, 8, 535. [Google Scholar] [CrossRef] [Green Version]
- Subcommittee on Antifungal Susceptibility Testing (AFST) of the ESCMID European Committee for Antimicrobial Susceptibility Testing (EUCAST). EUCAST Definitive Document EDef 7.1: Method for the determination of broth dilution MICs of antifungal agents for fermentative yeasts. Clin. Microbiol. Infect. 2008, 14, 398–405. [Google Scholar] [CrossRef]
- Damasceno, L.S.; de Teixeira, M.M.; Barker, B.M.; Almeida, M.A.; de Medeiros Muniz, M.; Pizzini, C.V.; Mesquita, J.R.L.; Rodríguez-Arellanes, G.; Ramírez, J.A.; Vite-Garín, T.; et al. Novel Clinical and Dual Infection by Histoplasma capsulatum Genotypes in HIV Patients from Northeastern, Brazil. Sci. Rep. 2019, 9, 11789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, R.A.; Joffe, L.S.; Alves, G.M.; Figueiredo-Carvalho, M.H.G.; Brito-Santos, F.; Amaral, A.C.F.; Rodrigues, M.L.; Almeida-Paes, R. A Screening of the MMV Pathogen Box® Reveals New Potential Antifungal Drugs against the Etiologic Agents of Chromoblastomycosis. PLoS ONE 2020, 15, e0229630. [Google Scholar] [CrossRef]
- Almeida, M.A.; Baeza, L.C.; Almeida-Paes, R.; Bailão, A.M.; Borges, C.L.; Guimarães, A.J.; Soares, C.M.A.; Zancopé-Oliveira, R.M. Comparative Proteomic Analysis of Histoplasma capsulatum Yeast and Mycelium Reveals Differential Metabolic Shifts and Cell Wall Remodeling Processes in the Different Morphotypes. Front. Microbiol. 2021, 12, 640931. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.D.M.; Lima, D.B.; Fischer, J.S.G.; Clasen, M.A.; Kurt, L.U.; Camillo-Andrade, A.C.; Monteiro, L.C.; de Aquino, P.F.; Neves-Ferreira, A.G.C.; Valente, R.H.; et al. Simple, Efficient and Thorough Shotgun Proteomic Analysis with PatternLab, V. Nat. Protoc. 2022, 17, 1553–1578. [Google Scholar] [CrossRef]
- Eng, J.K.; Jahan, T.A.; Hoopmann, M.R. Comet: An Open-Source MS/MS Sequence Database Search Tool. Proteomics 2013, 13, 22–24. [Google Scholar] [CrossRef]
- Carvalho, P.C.; Fischer, J.S.G.; Xu, T.; Cociorva, D.; Balbuena, T.S.; Valente, R.H.; Perales, J.; Yates, J.R.; Barbosa, V.C. Search Engine Processor: Filtering and Organizing Peptide Spectrum Matches. Proteomics 2012, 12, 944–949. [Google Scholar] [CrossRef]
- Carvalho, P.C.; Yates, J.R.; Barbosa, V.C. Improving the TFold Test for Differential Shotgun Proteomics. Bioinforma 2012, 28, 1652–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE Database Resources in 2022: A Hub for Mass Spectrometry-Based Proteomics Evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]
- Basenko, E.Y.; Pulman, J.A.; Shanmugasundram, A.; Harb, O.S.; Crouch, K.; Starns, D.; Warrenfeltz, S.; Aurrecoechea, C.; Stoeckert, C.J.; Kissinger, J.C.; et al. FungiDB: An Integrated Bioinformatic Resource for Fungi and Oomycetes. J. Fungi 2018, 4, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO Summarizes and Visualizes Long Lists of Gene Ontology Terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umscheid, C.A.; Margolis, D.J.; Grossman, C.E. Key Concepts of Clinical Trials: A Narrative Review. Postgrad. Med. 2011, 123, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug Repurposing: Progress, Challenges and Recommendations. Nat. Rev. Drug Discov. 2019, 18, 41–58. [Google Scholar] [CrossRef]
- Rhein, J.; Morawski, B.M.; Hullsiek, K.H.; Nabeta, H.W.; Kiggundu, R.; Tugume, L.; Musubire, A.; Akampurira, A.; Smith, K.D.; Alhadab, A.; et al. Efficacy of Adjunctive Sertraline for the Treatment of HIV-Associated Cryptococcal Meningitis: An Open-Label Dose-Ranging Study. Lancet Infect. Dis. 2016, 16, 809–818. [Google Scholar] [CrossRef] [Green Version]
- Capoci, I.R.G.; Faria, D.R.; Sakita, K.M.; Rodrigues-Vendramini, F.A.V.; de Bonfim-Mendonça, P.S.; Becker, T.C.A.; Kioshima, É.S.; Svidzinski, T.I.E.; Maigret, B. Repurposing Approach Identifies New Treatment Options for Invasive Fungal Disease. Bioorganic Chem. 2019, 84, 87–97. [Google Scholar] [CrossRef]
- Fothergill, A.W. Miconazole: A Historical Perspective. Expert Rev. Anti Infect. Ther. 2006, 4, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Dolan, K.; Montgomery, S.; Buchheit, B.; Didone, L.; Wellington, M.; Krysan, D.J. Antifungal Activity of Tamoxifen: In Vitro and in Vivo Activities and Mechanistic Characterization. Antimicrob. Agents Chemother. 2009, 53, 3337–3346. [Google Scholar] [CrossRef] [Green Version]
- Tonini, M.; De Ponti, F.; Di Nucci, A.; Crema, F. Review Article: Cardiac Adverse Effects of Gastrointestinal Prokinetics. Aliment. Pharmacol. Ther. 1999, 13, 1585–1591. [Google Scholar] [CrossRef]
- Moriyama, B.; Henning, S.A.; Leung, J.; Falade-Nwulia, O.; Jarosinski, P.; Penzak, S.R.; Walsh, T.J. Adverse Interactions between Antifungal Azoles and Vincristine: Review and Analysis of Cases. Mycoses 2012, 55, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Pozzi, E.; Fumagalli, G.; Chiorazzi, A.; Canta, A.; Cavaletti, G. Genetic Factors Influencing the Development of Vincristine-Induced Neurotoxicity. Expert Opin. Drug Metab. Toxicol. 2021, 17, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Jiang, S.; Tan, L.; Shi, H.; Yang, L.; Sun, Y.; Wang, X. Antifungal Activity of Minocycline and Azoles Against Fluconazole-Resistant Candida Species. Front. Microbiol. 2021, 12, 649026. [Google Scholar] [CrossRef]
- Jadhav, A.; Bansode, B.; Phule, D.; Shelar, A.; Patil, R.; Gade, W.; Kharat, K.; Karuppayil, S.M. The Antibacterial Agent, Moxifloxacin Inhibits Virulence Factors of Candida Albicans through Multitargeting. World J. Microbiol. Biotechnol. 2017, 33, 96. [Google Scholar] [CrossRef] [PubMed]
- Loreto, E.S.; Tondolo, J.S.M.; Santurio, J.M.; Alves, S.H. Screening of Antibacterial Drugs for Antimicrobial Activity against Pythium Insidiosum. Med. Mycol. 2019, 57, 523–525. [Google Scholar] [CrossRef]
- Guerini, A.E.; Triggiani, L.; Maddalo, M.; Bonù, M.L.; Frassine, F.; Baiguini, A.; Alghisi, A.; Tomasini, D.; Borghetti, P.; Pasinetti, N.; et al. Mebendazole as a Candidate for Drug Repurposing in Oncology: An Extensive Review of Current Literature. Cancers 2019, 11, 1284. [Google Scholar] [CrossRef] [Green Version]
- Almeida-Silva, F.; Damasceno, L.S.; Serna, M.J.B.; Valero, C.; Quintella, L.P.; Almeida-Paes, R.; de Muniz, M.M.; Zancope-Oliveira, R.M. Multiple Opportunistic Fungal Infections in an Individual with Severe HIV Disease: A Case Report. Rev. Iberoam. Micol. 2016, 33, 118–121. [Google Scholar] [CrossRef] [PubMed]
- De Nunes, J.O.; Pillon, K.R.A.P.; Bizerra, P.L.; Paniago, A.M.M.; Mendes, R.P.; Chang, M.R. The Simultaneous Occurrence of Histoplasmosis and Cryptococcal Fungemia: A Case Report and Review of the Literature. Mycopathologia 2016, 181, 891–897. [Google Scholar] [CrossRef]
- Aronis, M.L.; dos Santos, R.P.; Goldani, L.Z. Disseminated Histoplasma capsulatum and Cryptococcus neoformans Co-Infection in Patients with AIDS. Mycopathologia 2011, 172, 233–236. [Google Scholar] [CrossRef]
- Ghosh, A.; Tilak, R.; Bhushan, R.; Dhameja, N.; Chakravarty, J. Lymphnodal Co-Infection of Cryptococcus and Histoplasma in a HIV-Infected Patient and Review of Published Reports. Mycopathologia 2015, 180, 105–110. [Google Scholar] [CrossRef]
- Bai, R.-Y.; Staedtke, V.; Wanjiku, T.; Rudek, M.A.; Joshi, A.; Gallia, G.L.; Riggins, G.J. Brain Penetration and Efficacy of Different Mebendazole Polymorphs in a Mouse Brain Tumor Model. Clin. Cancer Res. 2015, 21, 3462–3470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riddell, J.; Wheat, L.J. Central Nervous System Infection with Histoplasma capsulatum. J. Fungi 2019, 5, 70. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.; Nizami, W.A. In Vitro Effects of Mebendazole on the Carbohydrate Metabolism of Avitellina Lahorea (Cestoda). J. Helminthol. 1987, 61, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, J.; Ramesh, R.; Chada, S.; Gomyo, Y.; Roth, J.A.; Mukhopadhyay, T. The anthelmintic drug mebendazole induces mitotic arrest and apoptosis by depolymerizing tubulin in non-small cell lung cancer cells. Mol Cancer Ther. 2002, 1, 1201–1209. [Google Scholar]
- Mukhopadhyay, T.; Sasaki, J.; Ramesh, R.; Roth, J.A. Mebendazole elicits a potent antitumor effect on human cancer cell lines both in vitro and in vivo. Clin Cancer Res. 2002, 8, 2963–2969. [Google Scholar] [PubMed]
- Daniel, J.P.; Mesquita, F.P.; Da Silva, E.L.; de Souza, P.F.N.; Lima, L.B.; de Oliveira, L.L.B.; de Moraes, M.E.A.; Moreira-Nunes, C.F.A.; Burbano, R.M.R.; Zanatta, G.; et al. Anticancer potential of mebendazole against chronic myeloid leukemia: In silico and in vitro studies revealed new insights about the mechanism of action. Front Pharmacol. 2022, 13, 952250. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.S.S.M.; Baird, S.K. Treatment of breast and colon cancer cell lines with anti-helmintic benzimidazoles mebendazole or albendazole results in selective apoptotic cell death. J. Cancer Res. Clin. Oncol. 2021, 147, 2945–2953. [Google Scholar] [CrossRef] [PubMed]
- Schmahl, G.; Benini, J. Treatment of Fish Parasites. 11. Effects of Different Benzimidazole Derivatives (Albendazole, Mebendazole, Fenbendazole) on Glugea Anomala, Moniez, 1887 (Microsporidia): Ultrastructural Aspects and Efficacy Studies. Parasitol. Res. 1998, 84, 41–49. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H. capsulatum Strain | Linezolid | Mebendazole | Moxifloxacin |
|---|---|---|---|
| G217B | 10 µM | 2.5 µM | 10 µM |
| IGS 4/5 | >20 µM | 1.25 µM | >20 µM |
| 2690603 | >20 µM | 0.08 µM | >20 µM |
| CE 17/13 | >20 µM | 0.08 µM | 20 µM |
| GO 11/15 | 20 µM | 2.5 µM | 20 µM |
| M477/08 | 10 µM | 2.5 µM | >20 µM |
| CAO4 | 2.5 µM | 0.6 µM | >20 µM |
| IPEC 23/11 | 0.3 µM | 2.5 µM | >20 µM |
| CE12/14 | >20 µM | 5.0 µM | >20 µM |
| 39942 | >20 µM | 1.25 µM | > 20 µM |
| 20258-1 | >20 µM | 5.0 µM | >20 µM |
| H. capsulatum Strain | MFC 1 | MFC/MIC 2 Ratio |
|---|---|---|
| G217B | >20 µM | >8 |
| IGS 4/5 | >20 µM | >16 |
| 2690603 | 1.25 µM | 16 |
| CE 17/13 | 2.5 µM | 32 |
| GO 11/15 | 2.5 µM | 1 |
| M477/08 | 2.5 µM | 1 |
| CAO4 | 2.5 µM | 4 |
| IPEC 23/11 | 2.5 µM | 1 |
| CE12/14 | >20 µM | >4 |
| 39942 | 1.25 µM | 1 |
| 20258-1 | >20 µM | >4 |
| Proteome Condition | KEGG Metabolic Pathway 1 | Odds Ratio 2 | p Value 2 |
|---|---|---|---|
| Without mebendazole | Aminoacyl-tRNA biosynthesis | 3.06 | 0.0215 |
| Without mebendazole>Citrate cycle | Without mebendazole>3.21 | Without mebendazole>0.0472 | |
| Treated with mebendazole | Pyrimidine metabolism | 9.49 | 0.00814 |
| Nitrogen metabolism | 12.47 | 0.0167 | |
| Sulfur metabolism | 15.17 | 0.0117 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almeida, M.A.; Bernardes-Engemann, A.R.; Coelho, R.A.; Lugones, C.J.G.; de Andrade, I.B.; Corrêa-Junior, D.; de Oliveira, S.S.C.; dos Santos, A.L.S.; Frases, S.; Rodrigues, M.L.; et al. Mebendazole Inhibits Histoplasma capsulatum In Vitro Growth and Decreases Mitochondrion and Cytoskeleton Protein Levels. J. Fungi 2023, 9, 385. https://doi.org/10.3390/jof9030385
Almeida MA, Bernardes-Engemann AR, Coelho RA, Lugones CJG, de Andrade IB, Corrêa-Junior D, de Oliveira SSC, dos Santos ALS, Frases S, Rodrigues ML, et al. Mebendazole Inhibits Histoplasma capsulatum In Vitro Growth and Decreases Mitochondrion and Cytoskeleton Protein Levels. Journal of Fungi. 2023; 9(3):385. https://doi.org/10.3390/jof9030385
Chicago/Turabian StyleAlmeida, Marcos Abreu, Andrea Reis Bernardes-Engemann, Rowena Alves Coelho, Camila Jantoro Guzman Lugones, Iara Bastos de Andrade, Dario Corrêa-Junior, Simone Santiago Carvalho de Oliveira, André Luis Souza dos Santos, Susana Frases, Márcio Lourenço Rodrigues, and et al. 2023. "Mebendazole Inhibits Histoplasma capsulatum In Vitro Growth and Decreases Mitochondrion and Cytoskeleton Protein Levels" Journal of Fungi 9, no. 3: 385. https://doi.org/10.3390/jof9030385