
Assessing the Biocontrol Potential of Clonostachys Species Isolated as Endophytes from Coffea Species and as Mycoparasites of Hemileia Rusts of Coffee in Africa
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Survey and Isolation of Purported Antagonists of Hemileia vastatrix
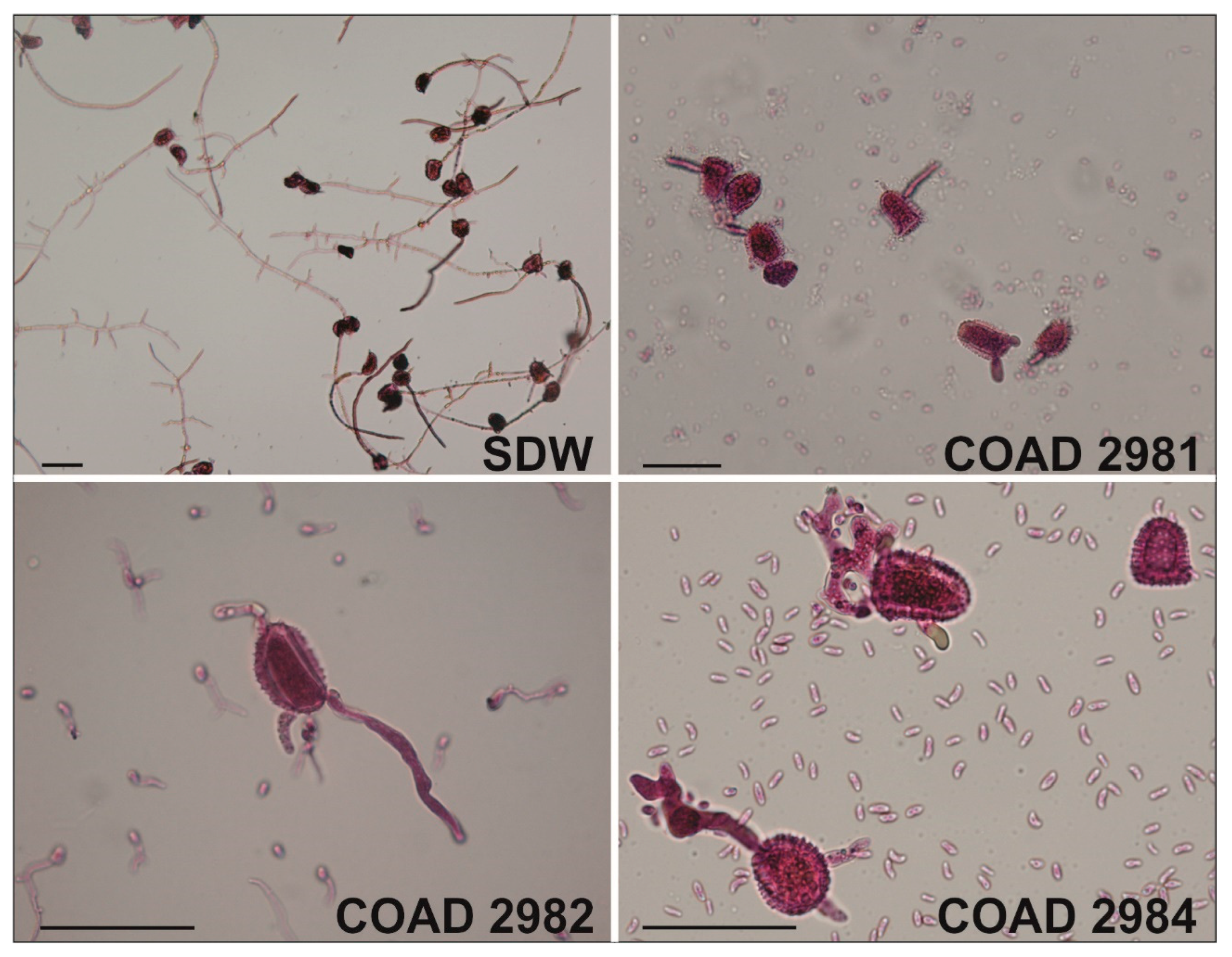
2.2. Morphology and Cultural Studies
2.3. DNA Extraction, Polymerase Chain Reactions (PCRs) and Sequencing
2.4. Sequences—Phylogenetic Analyses
2.5. In Planta and In Vitro Antagonism of Isolates of Clonostachys against Hemileia vastatrix
2.5.1. Assay Settings
2.5.2. Inoculum Preparation and Application
2.5.3. Dual Purpose Assay: In Planta Antagonism of Clonostachys against Hemileia vastatrix and Evaluation of Endophytic Establishment in Coffea arabica
2.5.4. Inhibition of Hemileia vastatrix Urediniospore Germination and Mycoparasitism
2.6. Statistical Analysis
3. Results
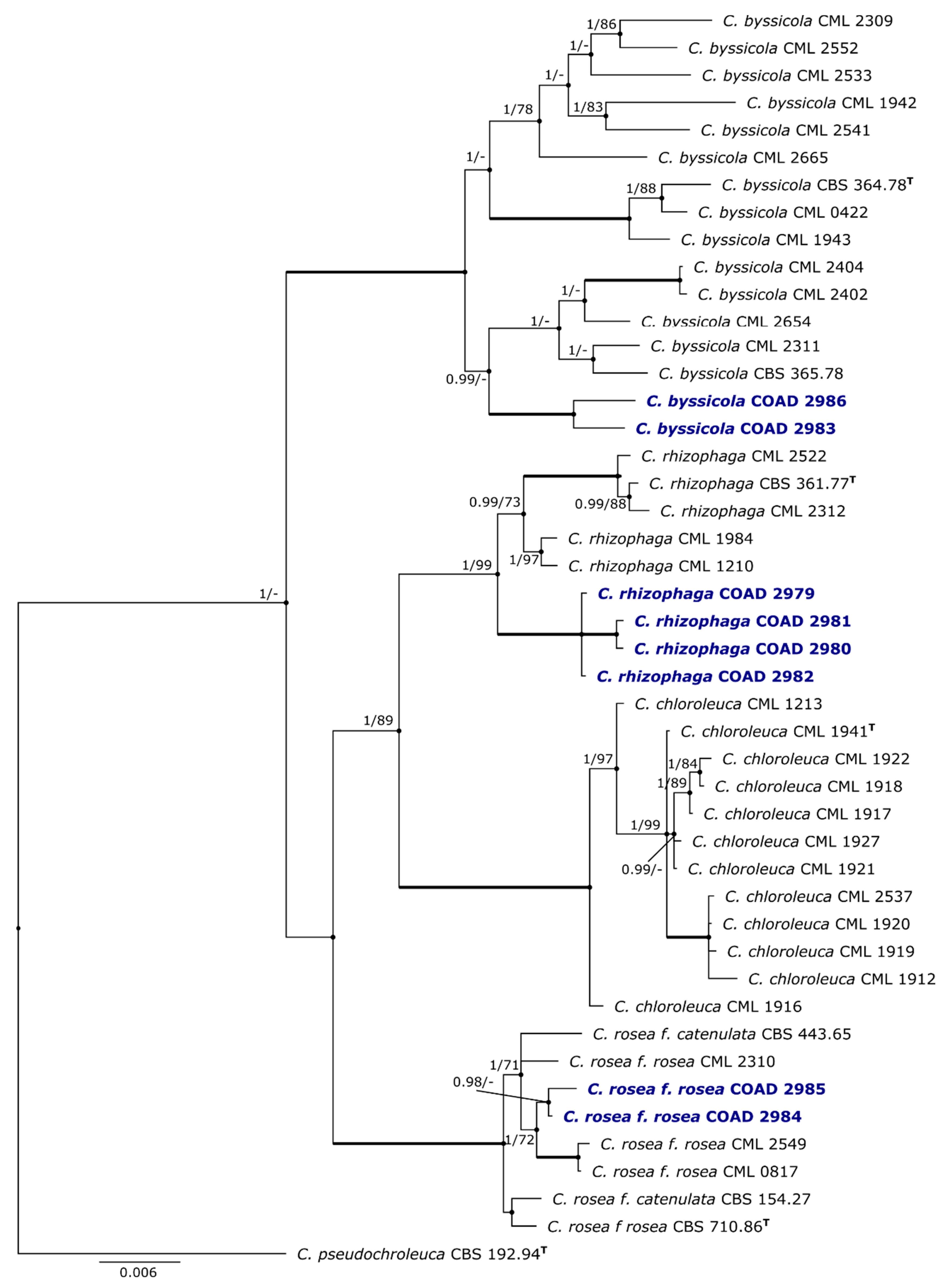
3.1. Phylogeny
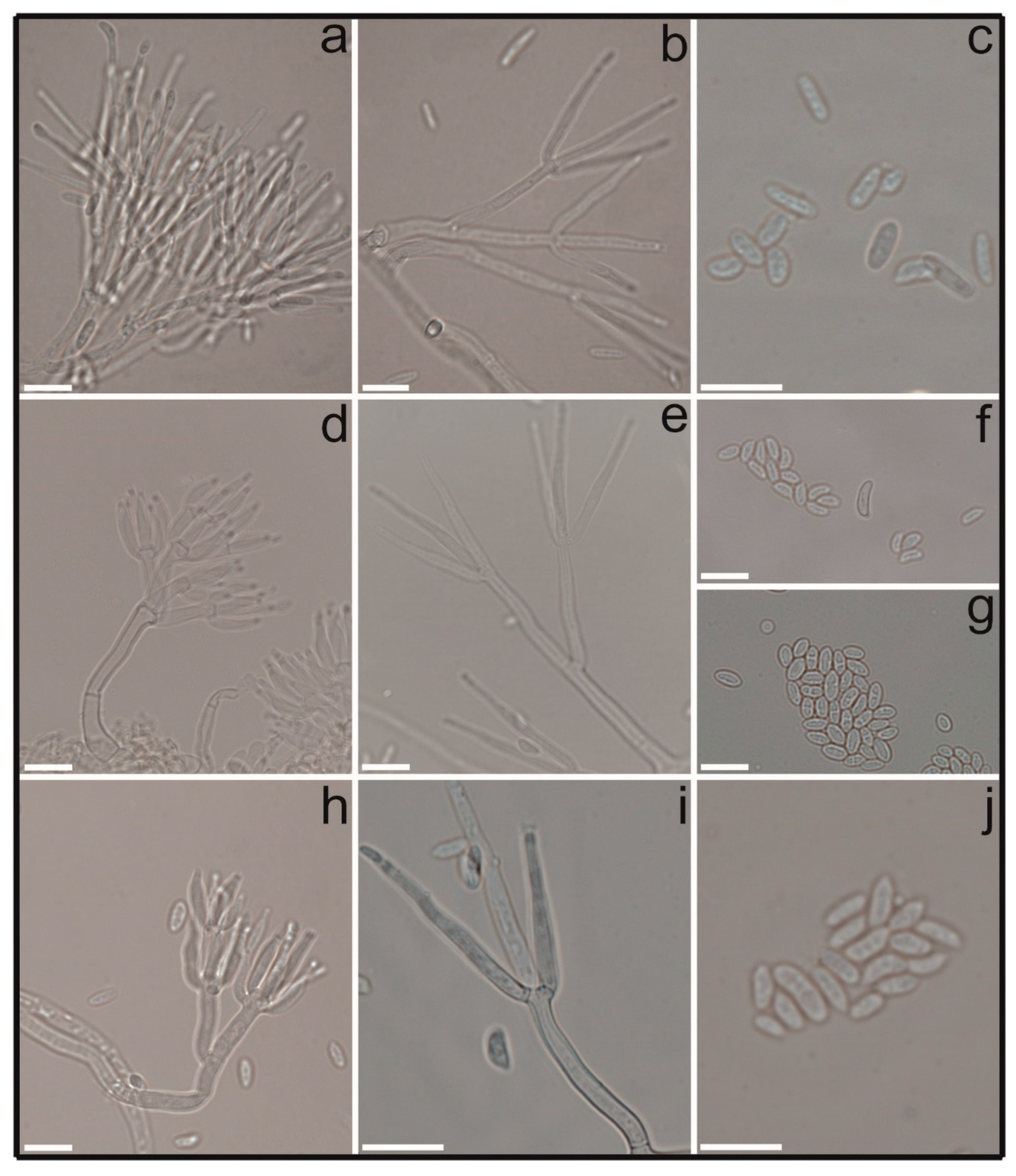
3.2. Taxonomy
3.3. Dual-Purpose Assay: In Planta Antagonism of Clonostachys against Hemileia vastatrix and Evaluation of Endophytic Establishment
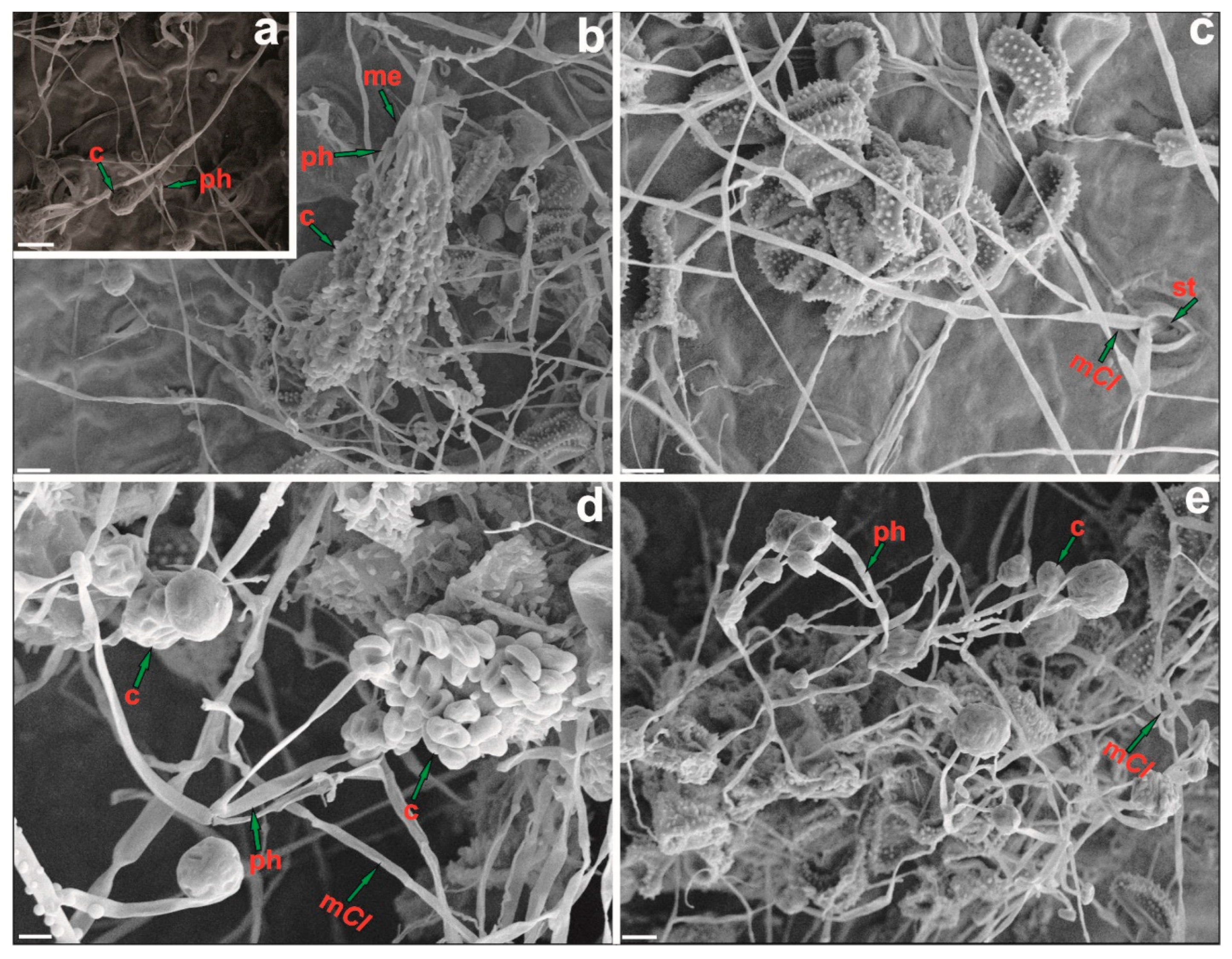
3.3.1. Endophytic Establishment of Clonostachys in Coffea arabica
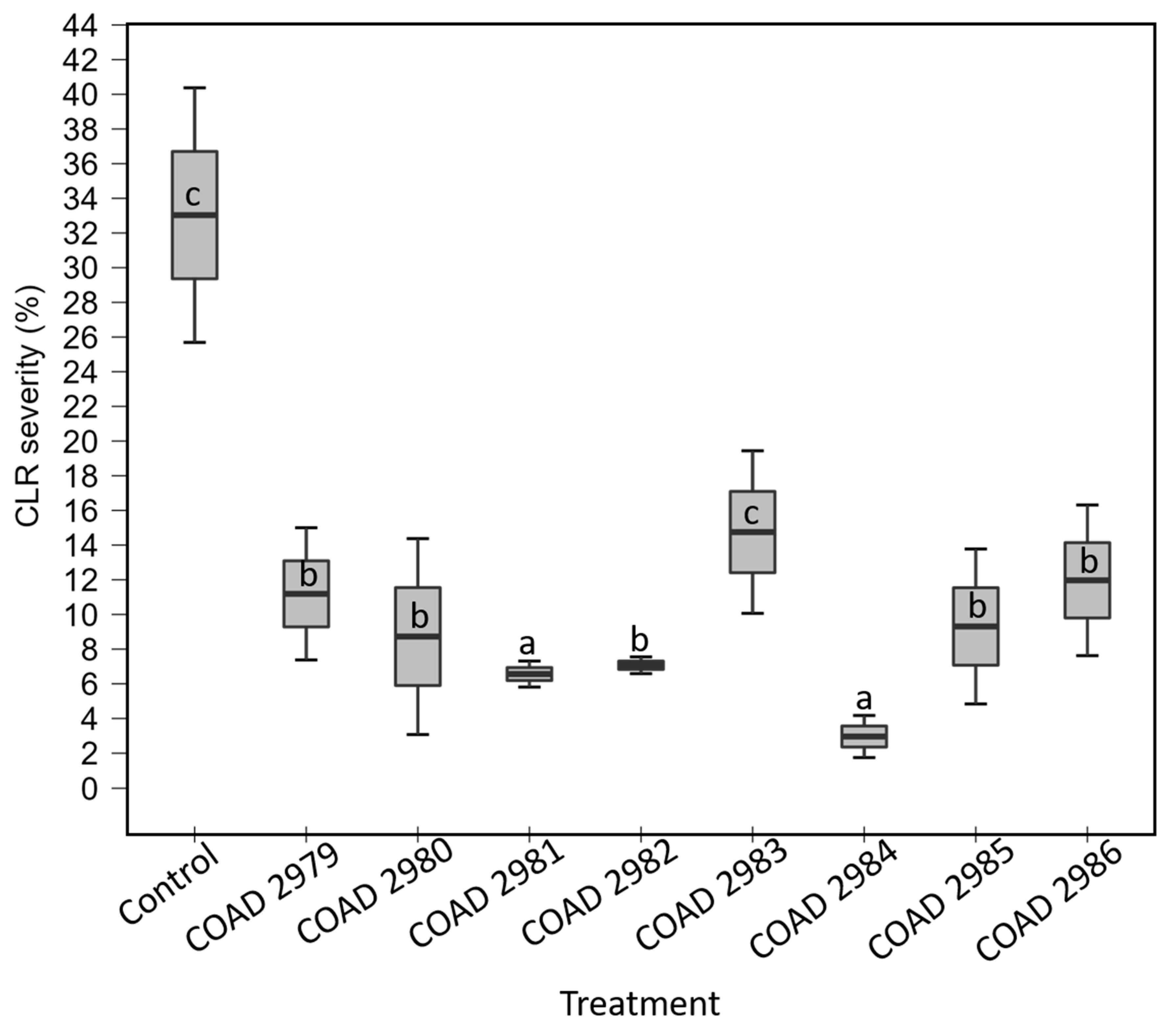
3.3.2. In Planta Antagonism of Clonostachys against Hemileia vastatrix
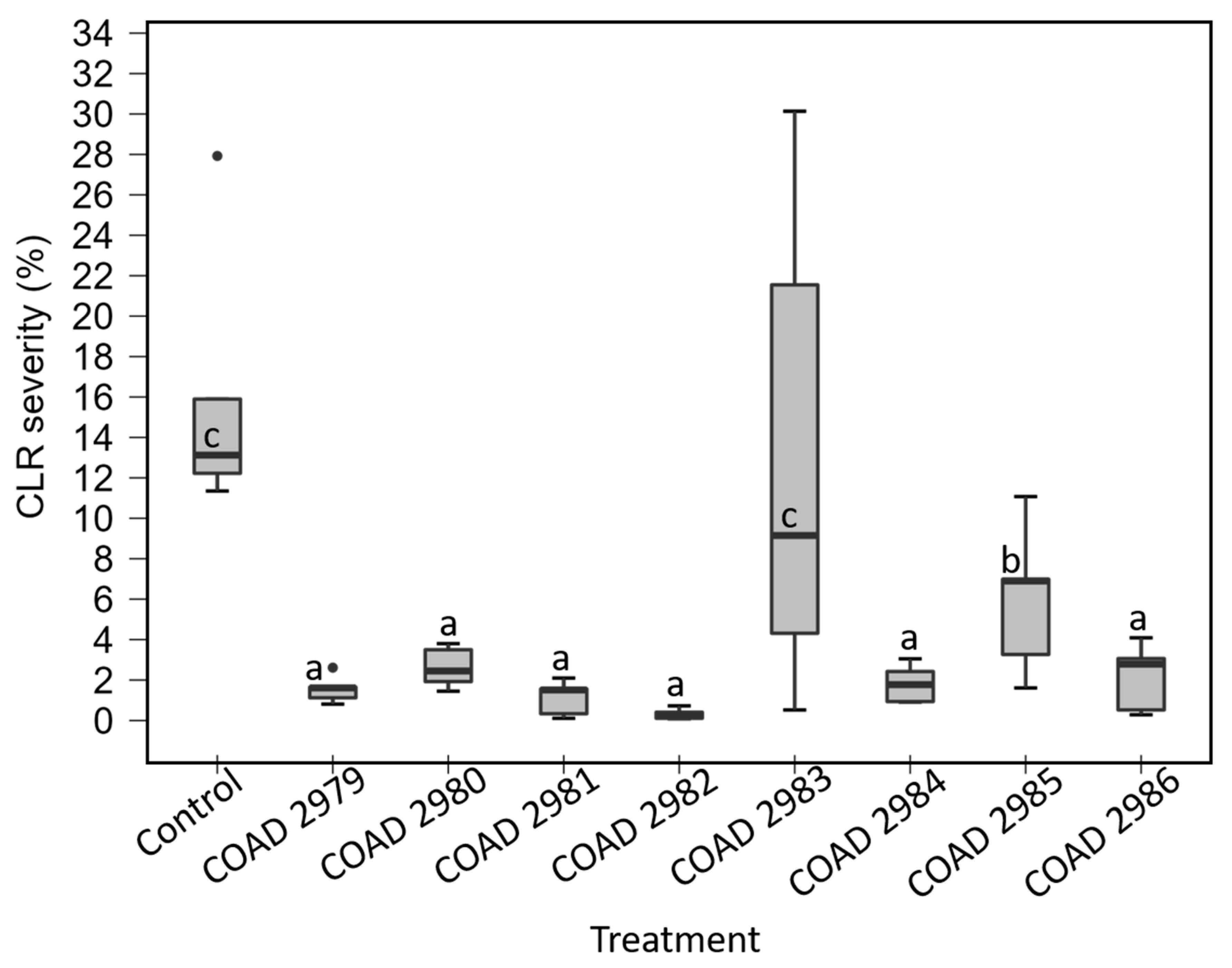
3.4. Inhibition of Hemileia vastatrix Urediniospore Germination and Mycoparasitism
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ICE (Intercontinental Exchange). Via Investopedia: Trading the Soft Commodity Markets. Available online: https://www.investopedia.com/articles/optioninvestor/07/soft_market.asp (accessed on 5 January 2022).
- ICO (International Coffee Organization). Sustainability of the Coffee Sector in Africa. 2015, pp. 1–29. Available online: https://www.ico.org/documents/cy2014-15/icc-114-5e-overview-coffee-sector-africa (accessed on 5 January 2022).
- Avelino, J.; Cristancho, M.; Georgiou, S.; Imbach, P.; Aguilar, L.; Bornemann, G.; Läderach, P.; Anzueto, F.; Hruska, A.J.; Morales, C. The coffee rust crises in Colombia and Central America (2008–2013): Impacts, plausible causes and proposed solutions. Food Sec. 2015, 7, 303–321. [Google Scholar] [CrossRef]
- Heeb, L.; Jenner, E.; Cock, M.J.W. Climate-smart pest management: Building resilience of farms and landscapes to changing pest threats. J. Pest Sci. 2019, 92, 951–969. [Google Scholar] [CrossRef]
- Zambolim, L. Current status and management of coffee leaf rust in Brazil. Trop. Plant Pathol. 2016, 41, 1–8. [Google Scholar] [CrossRef]
- Keith, L.M.; Sugiyama, L.S.; Brill, E.; Adams, B.L.; Fukada, M.; Hoffman, K.M.; Ocenar, J.; Kawabata, A.; Kong, A.T.; McKemy, J.M.; et al. First report of coffee leaf rust caused by Hemileia vastatrix on coffee (Coffea arabica) in Hawaii. Plant Dis. 2021, 106, 761. [Google Scholar] [CrossRef]
- Talhinhas, P.; Batista, D.; Diniz, I.; Vieira, A.; Silva, D.N.; Loureiro, A.; Tavares, S.; Pereira, A.P.; Azinheira, H.G.; Guerra-Guimarães, L.; et al. The coffee leaf rust pathogen Hemileia vastatrix: One and a half centuries around the tropics. Mol. Plant Pathol. 2017, 18, 1039–1051. [Google Scholar] [CrossRef]
- Belachew, B.K.; Senbeta, G.A.; Garedew, W.; Barreto, R.W.; Del Ponte, E.M. Altitude is the main driver of coffee leaf rust epidemics: A large-scale survey in Ethiopia. Trop. Plant Pathol. 2020, 45, 511–521. [Google Scholar] [CrossRef]
- Vega, F.E.; Posada, F.; Peterson, S.W.; Gianfagna, T.; Chaves, F. Penicillium species endophytic in coffee plants and ochratoxin A production. Mycologia 2006, 98, 37–48. [Google Scholar] [CrossRef]
- Vega, F.E.; Posada, F.; Aime, M.C.; Peterson, S.W.; Rehner, S.A. Fungal endophytes in green coffee seeds. Mycosystema 2008, 27, 75–84. [Google Scholar]
- Vega, F.E.; Simpkins, A.; Aime, M.C.; Posada, F.; Peterson, S.W.; Rehner, S.A.; Infante, F.; Castillo, A.; Arnold, A.E. Fungal endophyte diversity in coffee plants from Colombia, Hawai’i, Mexico and Puerto Rico. Fungal Ecol. 2010, 3, 122–138. [Google Scholar] [CrossRef]
- Bongiorno, V.A.; Rhoden, S.A.; Garcia, A.; Polonio, J.C.; Azevedo, J.L.; Pereira, J.O.; Pamphile, J.A. Genetic diversity of endophytic fungi from Coffea arabica cv. IAPAR-59 in organic crops. Ann. Microbiol. 2016, 66, 855–865. [Google Scholar] [CrossRef]
- Fulthorpe, R.; Martin, A.R.; Isaac, M.E. Root endophytes of coffee (Coffea arabica): Variation across climatic gradients and relationships with functional traits. Phytobiomes J. 2020, 4, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Meza, C.P.; Leguizamón, J. Evaluación de formulaciones de Bacillus thuringiensis Berliner y Verticillium lecanii Zimm (Viegas) en el control de la roya del cafeto Hemileia vastatrix Berk. y Br. Fitopatol. Colomb. 1995, 19, 50–56. [Google Scholar]
- González, E.; Martinez, B. Control biologico de la roya del cafeto (Hemileia vastatrix Berk y Br.) con Verticillium lecanii (Zimm) Viegas. Rev. Prot. Veg. 1998, 13, 81–84. [Google Scholar]
- Canjura-Saraiva, E.M.; Sánchez-Garita, V.; Krauss, U.; Somarriba, E. Reproducción masiva de Verticillium sp., hiperparásito de la roya del café, Hemileia vastatrix. Manejo Integr. Plagas Agroecol. 2002, 66, 13–19. [Google Scholar]
- Carrión, G.; Rico-Gray, V. Mycoparasites on the coffee rust in Mexico. Fungal Divers 2002, 11, 49–60. [Google Scholar]
- Vandermeer, J.; Perfecto, I.; Liere, H. Evidence for hyperparasitism of coffee rust (Hemileia vastatrix) by the entomogenous fungus, Lecanicillium lecanii, through a complex ecological web. Plant Pathol. 2009, 58, 636–641. [Google Scholar] [CrossRef]
- Mulaw, T.B.; Druzhinina, I.S.; Kubicek, C.P.; Atanasova, L. Novel endophytic Trichoderma spp. isolated from healthy Coffea arabica roots are capable of controlling coffee tracheomycosis. Diversity 2013, 5, 750–766. [Google Scholar] [CrossRef]
- Colmán, A.A.; Evans, H.C.; Salcedo-Sarmiento, S.S.; Braun, U.; Belachew-Bekele, K.; Barreto, R.W. A fungus-eat-fungus world: Digitopodium, with particular reference to mycoparasites of the coffee leaf rust, Hemileia vastatrix. IMA Fungus 2021, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Nóbrega, T.F. Taxonomy of Fusarium spp. Associated with Uredinia of Hemileia vastatrix in Brazil, Cameroon, Ethiopia and Paraguay and Their Biocontrol Potential. Ph.D. Thesis, Universidade Federal de Viçosa, Viçosa, MG, Brazil, 2021. [Google Scholar]
- Rodríguez, M.C.H.; Evans, H.C.; Abreu, L.M.; Macedo, D.M.; Kapeua-Ndacnou, M.; Bekele, K.B.; Barreto, R.W. New species and reports of Trichoderma isolated as mycoparasites and endophytes from cultivated and wild coffee in Africa. Sci. Rep. 2021, 11, 5671. [Google Scholar] [CrossRef]
- Pereira, C.M. Taxonomy of Fungi of Cladosporium and Cordyceps Genera with Potential for Biocontrol of Hemileia vastatrix. Master’s Thesis, Universidade Federal de Viçosa, Viçosa, MG, Brazil, 2021. [Google Scholar]
- Kapeua-Ndacnou, M.; Nóbrega, F.T.; Batista, R.L.; Evans, H.C.; Begoudé, A.B.D.; Barreto, R.W. Aspergillus flavus: A non-aflatoxigenic endophytic isolate from coffee in Cameroon antagonistic to coffee leaf rust (Hemileia vastatrix). J. Appl. Microbiol. 2022. submitted. [Google Scholar]
- Rodríguez, M.C.H. Trichoderma spp. Associated with Coffee in Africa: Taxonomy and Potential Uses for the Biological Control of Coffee Leaf Rust and Plant Growth Promotion. Ph.D. Thesis, Universidade Federal de Viçosa, Viçosa, MG, Brazil, 2019. [Google Scholar]
- Colmán, A.A. Hidden Diversity behind the White Colony Forming Fungi on H. vastatrix (Coffee Leaf Rust). Ph.D. Thesis, Universidade Federal de Viçosa, Viçosa, MG, Brazil, 2018. [Google Scholar]
- Salcedo-Sarmiento, S. New Mycoparasitic Fungi with Potential for the Biocontrol of Hemileia vastatrix (Coffee Leaf Rust). Ph.D. Thesis, Universidade Federal de Viçosa, Viçosa, MG, Brazil, 2018. [Google Scholar]
- Gonçalves, M.R.; Alfenas, C.A. Preparações e observações microscópicas de espécimes fúngicos. In Métodos em Fitopatologia, 2nd ed.; Gonçalves, M.R., Alfenas, C.A., Eds.; Universidade Federal de Viçosa: Viçosa, MG, Brazil, 2016; pp. 216–217. [Google Scholar]
- Moreira, M.G.; Abreu, M.L.; Carvalho, G.V.; Schroers, H.-J.; Pfenning, H.L. Multilocus phylogeny of Clonostachys subgenus Bionectria from Brazil and description of Clonostachys chloroleuca sp. nov. Mycol. Progress 2016, 15, 1031–1039. [Google Scholar] [CrossRef]
- Crous, P.W.; Verkley, J.Z.; Groenewald, S.R.A. Fungal Biodiversity; CBS KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2009. [Google Scholar]
- Rayner, R.W. A Mycological Colour Chart; CMI and British Mycological Society: Kew, UK, 1970. [Google Scholar]
- Dhingra, O.B.; Sinclair, J.B. Basic Plant Pathology Methods, 2nd ed.; CRC Press: Boca Raton, FL, USA, 1995. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed]
- Gräfenhan, T.; Schroers, H.-J.; Nirenberg, H.I.; Seifert, K.A. An overview of the taxonomy, phylogeny, and typification of nectriaceous fungi in Cosmospora, Acremonium, Fusarium, Stilbella, and Volutella. Stud. Mycol. 2011, 68, 79–113. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Sutton, D.A.; Rinaldi, M.G.; Sarver, B.A.J.; Balajee, S.A.; Schroers, H.-J.; Summerbell, R.C.; Robert, V.A.R.G.; Crous, P.W.; Zhang, N.; et al. Internet-accessible DNA sequence database for identifying fusaria from human and animal infections. J. Clin. Microbiol. 2010, 48, 3708–3718. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Hepperle, D.H. DNA Dragon 1.4.1—DNA Sequence Contig Assembler Software. 2011. Available online: www.dna-dragon.com (accessed on 19 March 2020).
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large data sets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Salcedo-Sarmiento, S.; Aucique-Pérez, C.E.; Silveira, P.R.; Colmán, A.A.; Silva, A.L.; Mansur, P.L.C.; Rodrigues, F.A.; Evans, H.C.; Barreto, R.W. Elucidating the interactions between the rust Hemileia vastatrix and a Calonectria mycoparasite and the coffee plant. iScience 2021, 24, 102352. [Google Scholar] [CrossRef]
- Belan, L.L.; Belan, L.L.; Matta Rafael, A.; Gonçalves Gomes, A.C.; Alves, R.F.; Cintra de Jesus Junior, W.; Moraes, B.W. Standard area diagram with color photograhs to estimate de severity of coffee leaf rust in Coffea canephora. Crop. Prot. 2020, 130, 105077. [Google Scholar] [CrossRef]
- Silva-Castro, I.; Barreto, R.W.; Rodríguez, M.C.H.; Matei, P.M.; Martín-Gil, J. Control of coffee leaf rust by chitosan oligomers and propolis. Sciendo 2018, 1, 311–315. [Google Scholar] [CrossRef]
- Schroers, H.-J. A monograph of Bionectria (Ascomycota, Hypocreales, Bionectriaceae) and its Clonostachys anamorphs. Stud. Mycol. 2001, 46, 1–214. [Google Scholar]
- Torcato, C.; Gonçalves, F.M.M.; Rodríguez-Gálvez, E.; Artur, A. Clonostachys viticola sp. nov., a novel species isolated from Vitis vinifera. Int. J. Syst. Evol. Microbiol. 2020, 70, 4321–4328. [Google Scholar] [CrossRef] [PubMed]
- Schroers, H.-J.; Samuels, G.S.; Seifert, K.A.; Gams, W. Classification of the mycoparasite Gliocladium roseum in Clonostachys as C. rosea, its relationship to Bionectria ochroleuca, and notes on other Gliocladium-like fungi. Mycologia 1999, 91, 365–385. [Google Scholar] [CrossRef]
- Tehon, J. Clonostachys rhizophaga (Tehon & Jacobs) Schroers, sp. nov ≡ Verticillium rhizophagum; Bull Davey Tree Expert Company: Kent, OH, USA, 1936; Volume 6, p. 16. [Google Scholar]
- Sun, Z.-B.; Li, S.-D.; Ren, Q.; Xu, J.-L.; Lu, X.; Sun, M.-H. Biology and applications of Clonostachys rosea. J. Appl. Microbiol. 2020, 129, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Abreu, M.L.; Moreira, M.G.; Ferreira, D.; Rodrigues-Filho, E.; Pfenning, H.L. Diversity of Clonostachys species assessed by molecular phylogenetics and MALDI-TOF mass spectrometry. Fungal Biol. 2014, 118, 1004–1012. [Google Scholar] [CrossRef] [PubMed]
- Nobre, S.; Maffia, L.; Mizubuti, E.; Cota, L.; Dias, A. Selection of Clonostachys rosea isolates from Brazilian ecosystems effective in controlling Botrytis cinerea. Biol. Control 2005, 34, 132–143. [Google Scholar] [CrossRef]
- Cota, L.; Maffia, L.; Mizubuti, E.; Macedo, P.; Antunes, R. Biological control of strawberry gray mold by Clonostachys rosea under field conditions. Biol. Control 2008, 46, 515–522. [Google Scholar] [CrossRef]
- Zaldúa, S.; Sanfuentes, E. Control of Botrytis cinerea in Eucalyptus globulus mini-cuttings using Clonostachys and Trichoderma strains. Chil. J. Agric. Res. 2010, 70, 576–582. [Google Scholar] [CrossRef]
- Ten Hoopen, G.M.; George, A.; Martinez, A.; Stirrup, T.; Flood, J.; Krauss, U. Compatibility between Clonostachys isolates with a view to mixed inocula for biocontrol. Mycologia 2010, 102, 1204–1215. [Google Scholar] [CrossRef]
- Krauss, U.; Ten Hoopen, M.; Rees, R.; Stirrup, T.; Argyle, T.; George, A. Mycoparasitism by Clonostachys byssicola and Clonostachys rosea on Trichoderma spp. from cocoa (Theobroma cacao) and implication for the design of mixed biocontrol agents. Biol. Control 2013, 67, 317–327. [Google Scholar] [CrossRef]
- Alvindia, D.G.; Natsuaki, K.T. Evaluation of fungal epiphytes isolated from banana fruit surfaces for biocontrol of banana crown rot disease. Crop Prot. 2008, 27, 1200–1207. [Google Scholar] [CrossRef]
- Mendoça Garciá, R.A.; Ten Hoopen, G.M.; Kass, D.C.J.; Sánchez Garita, V.A.; Krauss, U. Evaluation of mycoparasites as biocontrol agents of Rosellinia root rot in cocoa. Biol. Control 2003, 27, 210–227. [Google Scholar] [CrossRef]
- Silva, H.A.O.; Teixeira, W.D.; Borges, Á.V.; Silva Junior, A.L.; Alves, K.S.; Rodrigues Junior, O.M.; Abreu, L.M. Biocontrol of potato early blight and suppression of Alternaria grandis sporulation by Clonostachys spp. Plant Pathol. 2021, 70, 1677–1685. [Google Scholar] [CrossRef]
- Dugan, F.M.; Lupien, S.L.; Hernandez-Bello, M.; Peever, T.L.; Chen, W. Fungi resident in chickpea debris and their suppression of growth and reproduction of Didymella rabiei under laboratory conditions. J. Phytopathol. 2005, 153, 431–439. [Google Scholar] [CrossRef]
- Corrêa, E.B.; Bettiol, W.; Morandi, M.A.B. Controle biológico da podridão de raiz causada por Pythium aphanidermatum e promoção de crescimento de alface hidropônica com Clonostachys rosea. Trop. Plant Pathol. 2010, 35, 248–252. [Google Scholar] [CrossRef]
- Nygren, K.; Dubey, M.; Zapparata, A. The mycoparasitic fungus Clonostachys rosea responds with both common and specific gene expression during interspecific interactions with fungal prey. Evol. Appl. 2018, 11, 931–949. [Google Scholar] [CrossRef] [Green Version]
- AGROFIT. 2019. Available online: http://agrofit.agricultura.gov.br/agrofit_cons/!ap_ing_ativo_detalhe_cons?p_id_ingrediente_ativo=794 (accessed on 5 October 2021).
- Xue, A.G. Gliocladium roseum Strains Useful for the Control of Fungal Pathogens in Plants. U.S. Patent 6,495,133 B1, 17 December 2002. [Google Scholar]
- Bettiol, W.; Morandi, M.A.B.; Pinto, Z.V.; Júnior, T.J.P.; Corrêa, E.B.; Moura, A.B.; Lucon, C.M.M.; Costa, J.C.B.; Bezerra, J.L. Produtos Comerciais à Base de Agentes Biocontrole de Doenças de Plantas; Embrapa Meio Ambiente: Jaguariúna, SP, Brazil, 2012; 155p, ISSN 1517-5111. [Google Scholar]
- Nemec, S.; Datnoff, L.; Strandberg, J. Efficacy of biocontrol agents in planting mixes to colonize plant roots and control root diseases of vegetables and citrus. Crop Prot. 1996, 15, 735–742. [Google Scholar] [CrossRef]
- Mejía, L.C.; Rojas, E.I.; Maynard, Z.; Bael, S.V.; Arnold, A.E.; Hebbar, P.; Samuels, G.J.; Robbins, N.; Herre, E.A. Endophytic fungi as biocontrol of Theobroma cacao pathogens. Biol. Control 2008, 46, 4–14. [Google Scholar] [CrossRef]
- Evans, H.C.; Holmes, K.A.; Thomas, S.E. Endophytes and mycoparasites associated with an indigenous forest tree, Theobroma gileri, in Ecuador and a preliminary assessment of their potential as biocontrol agents of cocoa diseases. Mycol. Progress 2003, 2, 149–160. [Google Scholar] [CrossRef]
- Abang, M.M.; Kabbabeh, S.; Ahmed, S.; Murad, S.; Chilvers, M.I.; Peever, T.L.; Schroers, H.-J. First report of chickpea wilt caused by Clonostachys rhizophaga in Syria. Plant Dis. 2009, 93, 666. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, A.-F.; Gu, C.-Y.; Zang, H.-Y.; Chen, Y. First report of Clonostachys rhizophaga as a pathogen of water chestnut (Eleocharis dulcis) in Anhui Province of China. Plant Dis. 2019, 103, 151. [Google Scholar] [CrossRef]
- Bienapfl, J.C.; Floyd, C.M.; Percich, J.A.; Malvick, D.K. First report of Clonostachys rosea causing root rot of soybean in the United States. Plant Dis. 2012, 96, 1700. [Google Scholar] [CrossRef] [PubMed]
- Afshari, N.; Hemmati, R. First report of the occurrence and pathogenecity of Clonostachys rosea on faba bean. Australasian Plant Pathol. 2017, 46, 231–234. [Google Scholar] [CrossRef]
- Guzzo, S.D.; Martins, E.M.F. Local and systemic induction of β-1,3-glucanase and chitinase in coffee leaves protected against Hemileia vastatrix by Bacillus thuringiensis. J. Phytopathol. 1996, 144, 449–454. [Google Scholar] [CrossRef]
- Howell, C.R. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef]
- Nga, N.T.T.; Giau, N.T.; Long, N.T.; Lübeck, M.; Shetty, N.P.; De Neergaard, E.; Thuy, T.T.T.; Kim, P.V.; Jørgensen, H.J.L. Rhizobacterially induced protection of watermelon against Didymella bryoniae. J. Appl. Microbiol. 2010, 109, 567–582. [Google Scholar] [CrossRef] [PubMed]
- Mejía, L.C.; Herre, E.A.; Sparks, J.P.; Winter, K.; García, M.N.; Van Bael, S.N.; Stitt, J.; Shi, Z.; Zhang, Y.; Guiltinan, M.J.; et al. Pervasive effects of a dominant foliar endophytic fungus on host genetic and phenotypic expression in a tropical tree. Font. Microbiol. 2014, 5, 479. [Google Scholar] [CrossRef]
- Harman, G.E.; Uphoff, N. Symbiotic root-endophytic soil microbes improve crop productivity and provide environmental benefits. Scientifica 2019, 2019, 9106395. [Google Scholar] [CrossRef]
- Sudha, V.; Govindaraj, R.; Baskar, K.; Al-Dhabi, N.A.; Duraipandiyan, V. Biological properties of endophytic fungi. Braz. Arch. Biol. Technol. 2016, 59, e16150436. [Google Scholar] [CrossRef]
- Saraiva, R.M.; Borges, Á.V.; Borel, F.C.; Maffia, L.A. Compounds produced by Clonostachys rosea deleterious to Botrytis cinerea. Braz. J. Agric. 2020, 95, 34–47. [Google Scholar] [CrossRef]
- Han, P.; Zhang, X.; Xu, D.; Zhang, B.; Lai, D.; Zhou, L. Metabolites from Clonostachys fungi and their biological activities. J. Fungi 2020, 6, 229. [Google Scholar] [CrossRef] [PubMed]
- Muvea, A.M.; Meyhöfer, R.; Subramanian, S.; Poehling, H.M.; Ekesi, S.; Maniania, N.K. Colonization of onions by endophytic fungi and their impacts on the biology of Thrips tabaci. PLoS ONE 2014, 9, e108242. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain No. | Host/Substrate | Locality/Country | GenBank Accession No. | |||
|---|---|---|---|---|---|---|---|
| TEF1-α | RPB1 | ACL1 | TUB | ||||
| Clonostachys byssicola | CML 0422 | Soil from secondary forest | Benjamin Constant, AM, Brazil | KX184964 | KX184899 | KX184833 | KF871150 |
| CML1942 | Soil from Amazon forest | Benjamin Constant, AM, Brazil | KX184968 | KX184903 | KX184837 | KF871148 | |
| CML1943 | Soil from Amazon forest | Benjamin Constant, AM, Brazil | KX184965 | KX184900 | KX184834 | KF871151 | |
| CML2309 | Fragaria ananassa | Bento Gonçalves, RS, Brazil | KX184966 | KX184901 | KX184835 | KF871149 | |
| CML2311 | Parasitizing colony of Sordaria sp. | Lavras, MG, Brazil | KX184969 | KX184904 | KX184838 | KF871152 | |
| CML2402 | Fruit of Annona squamosa | Januária, MG, Brazil | KX184970 | KX184905 | KX184839 | KX185030 | |
| CML2404 | Fruit of Annona x atemoya | Jaíba, MG, Brazil | KX184971 | KX184906 | KX184840 | KF871153 | |
| CML 2510/CBS 364.78T | Bark | Venezuela | KX184967 | KX184902 | KX184836 | AF358153 | |
| CML 2511/CBS 365.78 | Wood | Venezuela | KX184972 | KX184907 | KX184841 | AF358154 | |
| CML2533 | Bryophyte | Itumirim, MG, Brazil | KX184973 | KX184908 | KX184842 | KX185031 | |
| CML2541 | Litter | Itumirim, MG, Brazil | KX184974 | KX184909 | KX184843 | KX185032 | |
| CML2552 | Piper nigrum | Montes Claros, MG, Brazil | KX184975 | KX184910 | KX184844 | KX185033 | |
| CML2654 | Litter | Barroso, MG, Brazil | KX184976 | KX184911 | KX184845 | KX185034 | |
| CML2665 | Litter | Lavras, MG, Brazil | KX184977 | KX184912 | KX184846 | KX185035 | |
| COAD 2983 | Hemileia sp., Coffea arabica | Bonga, Ethiopia | OM038397 | OM038404 | OM038391 | ||
| COAD 2986 | Endophyte/stems, wild Coffea arabica | Bonga, Ethiopia | OM038397 | OM038401 | OM038390 | ||
| C. chloroleuca | CML 2537 | Bryophyte | Itumirim, MG, Brazil | KX184989 | KX184924 | KX184858 | KX185038 |
| CML 1941T | Native soil from Cerrado | Montividiu, GO, Brazil | KX184988 | KX184923 | KX184857 | KF871172 | |
| CML 1927 | Soil under soybean field | Montividiu, GO, Brazil | KX184987 | KX184922 | KX184856 | KF871171 | |
| CML 1922 | Native soil from Cerrado | Montividiu, GO, Brazil | KX184986 | KX184921 | KX184855 | KF871170 | |
| CML 1921 | Native soil from Cerrado | Montividiu, GO, Brazil | KX184985 | KX184920 | KX184854 | KF871166 | |
| CML 1920 | Native soil from Cerrado | Montividiu, GO, Brazil | KX184984 | KX184919 | KX184853 | KX185037 | |
| CML 1919 | Native soil from Cerrado | Montividiu, GO, Brazil | KX184983 | KX184918 | KX184852 | KF871167 | |
| CML 1918 | Native soil from Cerrado | Montividiu, GO, Brazil | KX184982 | KX184917 | KX184851 | KX185036 | |
| CML 1917 | Native soil from Cerrado | Montividiu, GO, Brazil | KX184981 | KX184916 | KX184850 | KF871169 | |
| CML 1916 | Soil under cotton field | Montividiu, GO, Brazil | KX184980 | KX184915 | KX184849 | KF871174 | |
| CML 1912 | Native soil from Cerrado | Montividiu, GO, Brazil | KX184979 | KX184914 | KX184848 | KF871168 | |
| CML 1213 | Native soil from Cerrado | Montividiu, GO, Brazil | KX184978 | KX184913 | KX184847 | KF871173 | |
| C. rhizophaga | CML 2522 | Soil | Lavras, MG, Brazil | KX184994 | KX184929 | KX184863 | KX185039 |
| CML2514 /CBS 361.77T | Culture contaminant | Switzerland | KX184993 | KX184928 | KX184862 | AF358158 | |
| CML 2312 | Parasitizing colony of Fusarium oxysporum | Lavras, MG, Brazil | KX184992 | KX184927 | KX184861 | KF871157 | |
| CML 1984 | Native soil from Cerrado | Montividiu, GO, Brazil | KX184991 | KX184926 | KX184860 | KF871155 | |
| CML 1210 | Soil under soybean field | Montividiu, GO, Brazil | KX184990 | KX184925 | KX184859 | KF871156 | |
| COAD 2979 | H. vastatrix/coffeicola, Coffea canephora | Somalomo, Cameroon | OM038395 | ||||
| COAD 2980 | H. vastatrix/coffeicola, Coffea canephora | Somalomo, Cameroon | OM038394 | OM038402 | |||
| COAD 2981 | H. vastatrix/coffeicola, Coffea canephora | Somalomo, Cameroon | OM038393 | ||||
| COAD 2982 | H. vastatrix/coffeicola, Coffea canephora | Somalomo, Cameroon | OM038396 | OM038403 | |||
| C. pseudochroleuca | CML 2562/CBS 192.94T | Bark | French Guiana | KX185016 | KX184950 | KX184885 | AF358171 |
| C. rosea f. catenulata | CML 2517/CBS 443.65 | Soil | USA | KX184996 | KX184931 | KX184865 | AF358166 |
| CML 2516/CBS 154.27T | Soil | USA | KX184995 | KX184930 | KX184864 | AF358160 | |
| C. rosea f. rosea | CML 2549 | Litter | Itumirim, MG, Brazil | KX185001 | KX184935 | KX184870 | KX185040 |
| CML 2518/CBS 710.86T | Soil, on sclerotia of Sclerotinia minor | Netherlands | KX184999 | KX184934 | KX184868 | AF358161 | |
| CML 2310 | Fragaria ananassa | Caxias do Sul, RS, Brazil | KX184998 | KX184933 | KX184867 | KF871146 | |
| CML 0817 | Endophyte, Lychnophora pinaster | Ingaí, MG, Brazil | KX184997 | KX184932 | KX184866 | KF871147 | |
| COAD 2984 | Endophyte/stems, Coffea arabica | Bonga, Ethiopia | OM038392 | OM038399 | OM038389 | ||
| COAD 2985 | Endophyte/stems, Coffea arabica | Bonga, Ethiopia | OM038400 | OM038388 | |||
| Character | COAD 2983/2986 | C. byssicola * | COAD 2979/2980/ 2981/2982 | C. rhizophaga * | COAD 2984/2985 | C. rosea f. rosea * |
|---|---|---|---|---|---|---|
| Stipe Length (µm) | 10–63 | 10–100 | 20–92 | 10–100 | 20–122 | 25–200 |
| Penicillus (µm) | 19–55.5 | 20–100 | 19–87 | 30–100 | 25.5–73.5 | 30–120 |
| Phialide Shape | divergent | divergent | divergent | divergent | divergent | divergent |
| Phialides in Whorls | 2–4 | 2–4 | 2–5 | 2–5 | 2–5 | 2–5 |
| Phialide Size (µm) | 19–54 × 2–3 | 12.4–48 × 1.4–2.8 | (9.5)11–35 × 1–3 | 15.6–48.2 × 2.2–3.2 | 20.5–40.5 × 1.5–3 | 25–45 × 1.6–3 |
| Secondary Conidiophore | adpressed | adpressed to divergent | penicillate | penicillate | adpressed | adpressed or divergent |
| Phialide Size (µm) | 10–32.5 × 1–2.5 | 7.6–27.8 × 1.4–2.8 | (5–)14.5–17(–28) × 1–2.5(–7) | 5.8–25.2 × 2.2–3.2 | 10–19 × 1–3 | 8–18 × 2–3 |
| Phialide Shape | adpressed | ±adpressed | adpressed | adpressed or divergent | adpressed | adpressed |
| Phialides in Whorls | 4–7 | 3–5 | 3–6 | 3–5 | 5–7 | – |
| No. of verticillia | 2–3 | 2–5 | 1–4 | 3–4 | 2–4 | 2–4 |
| Conidia Size (µm) | 2.5–10(–14) × 1.5–4 | 3.2–10.8 × 1.8–4 | 3.5–9(11) × 2–5.5(–7.5) | 4.8–9 × 2.4–4.2 | 4–10 × 1.5–3.5 | 5.6–10 × 2–3.6 |
| Assay I a | Assay II b | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Month of Isolation (After First Inoculation) | |||||||||||||||
| 1st | 2nd | 3rd | 5th | 5th | |||||||||||
| Isolate c | Leaf | Stem | Root | Leaf | Stem | Root | Leaf | Stem | Root | Leaf | Stem | Root | Leaf | Stem | Root |
| Control | − * | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| COAD 2979 | + * | + | + | + | + | + | + | + | + | + | + | + | + | + | − |
| COAD 2980 | + | + | − | + | + | + | + | − | + | + | + | + | + | + | + |
| COAD 2981 | + | + | − | + | + | + | − | + | + | + | + | − | + | − | − |
| COAD 2982 | + | + | + | + | + | + | − | − | + | + | + | + | + | + | − |
| COAD 2983 | + | + | + | + | + | − | − | + | + | + | + | + | + | + | + |
| COAD 2984 | + | + | − | + | + | + | + | − | + | + | + | + | + | − | + |
| COAD 2985 | + | + | − | + | + | + | − | − | + | + | + | + | + | + | + |
| COAD 2986 | + | − | + | + | + | + | − | + | + | + | + | + | + | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapeua-Ndacnou, M.; de Abreu, L.M.; de Macedo, D.M.; da Nóbrega, T.F.; Pereira, C.M.; Evans, H.C.; Barreto, R.W. Assessing the Biocontrol Potential of Clonostachys Species Isolated as Endophytes from Coffea Species and as Mycoparasites of Hemileia Rusts of Coffee in Africa. J. Fungi 2023, 9, 248. https://doi.org/10.3390/jof9020248
Kapeua-Ndacnou M, de Abreu LM, de Macedo DM, da Nóbrega TF, Pereira CM, Evans HC, Barreto RW. Assessing the Biocontrol Potential of Clonostachys Species Isolated as Endophytes from Coffea Species and as Mycoparasites of Hemileia Rusts of Coffee in Africa. Journal of Fungi. 2023; 9(2):248. https://doi.org/10.3390/jof9020248
Chicago/Turabian StyleKapeua-Ndacnou, Miraine, Lucas Magalhães de Abreu, Davi Mesquita de Macedo, Thaisa Ferreira da Nóbrega, Caio Mattos Pereira, Harry Charles Evans, and Robert Weingart Barreto. 2023. "Assessing the Biocontrol Potential of Clonostachys Species Isolated as Endophytes from Coffea Species and as Mycoparasites of Hemileia Rusts of Coffee in Africa" Journal of Fungi 9, no. 2: 248. https://doi.org/10.3390/jof9020248