Recent Advances in Search of Bioactive Secondary Metabolites from Fungi Triggered by Chemical Epigenetic Modifiers

 and
and
Abstract
:1. Introduction
2. Chemical Epigenetic Modifiers and Their Action Mechanisms
2.1. DNA Methyltransferase Modifiers
2.2. Histone Deacetylase Modifiers
2.3. Other Chemical Epigenetic Modifiers

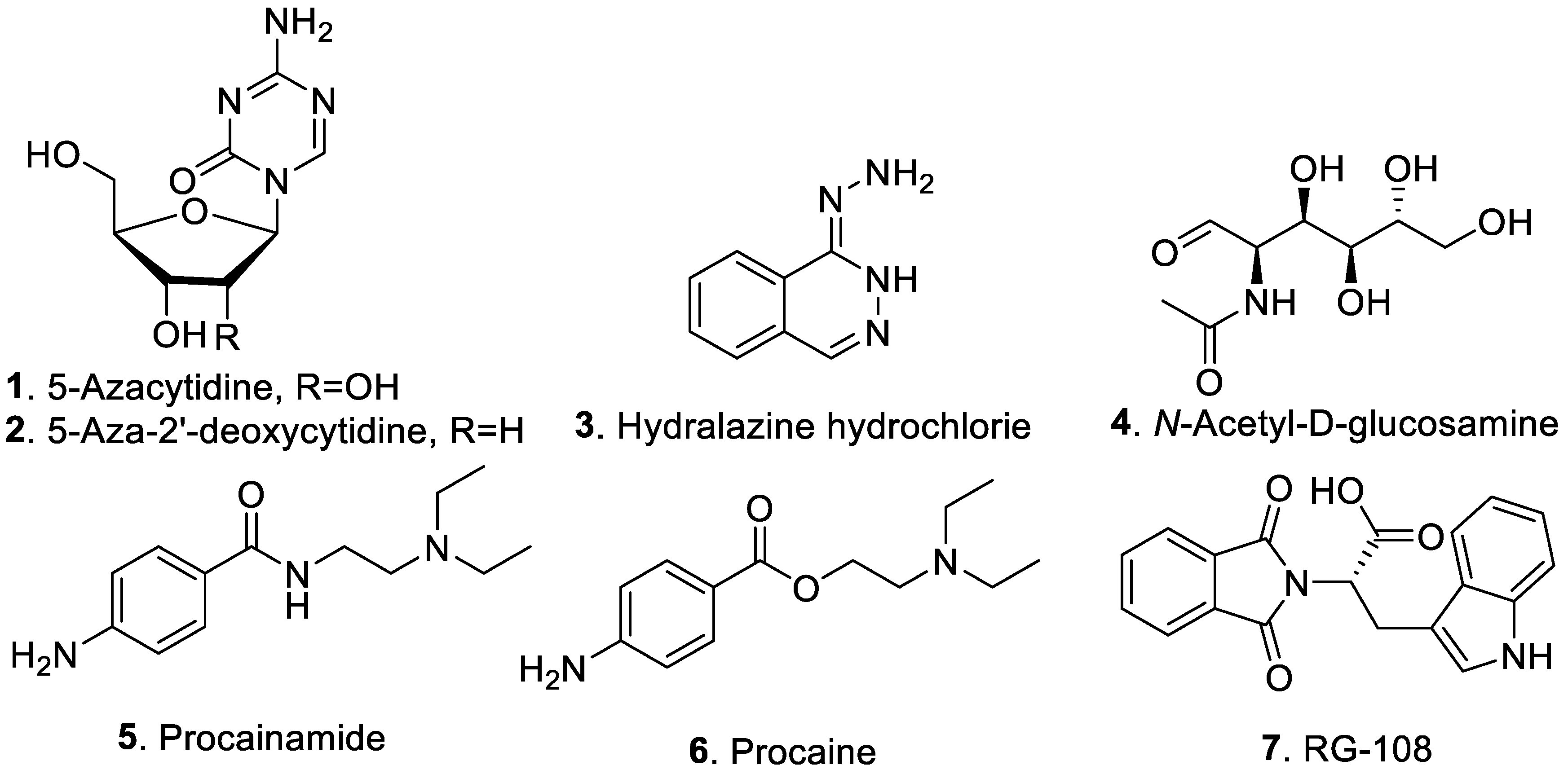{kind=link}
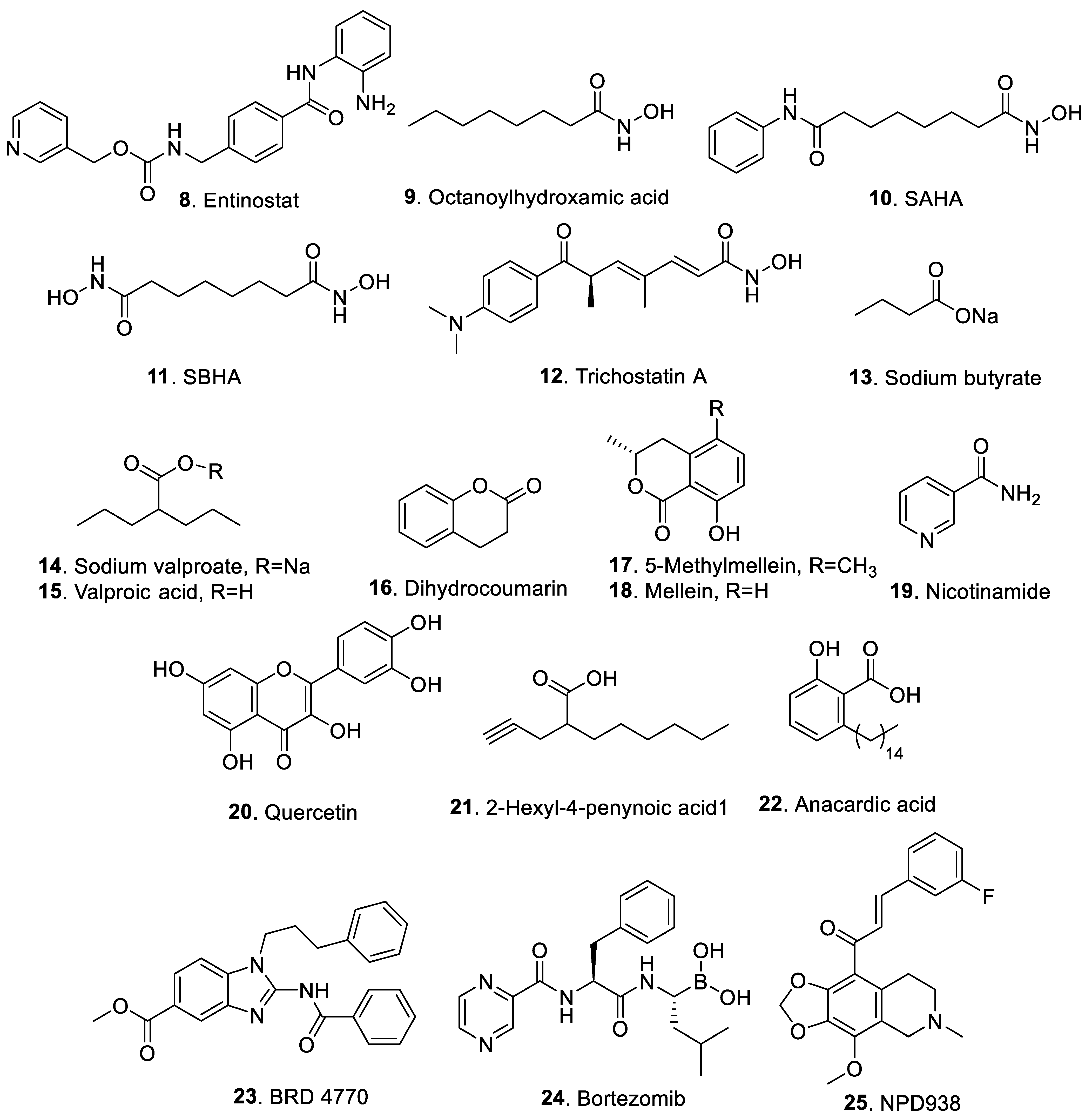{kind=link}
| Modifier | Mechanism of Action | Ref. |
|---|---|---|
| 5-Azacytidine (1) | Inhibition of DNA methyltransferase | [26,35] |
| 5-Aza-2′-deoxycytidine (2) | Inhibition of DNA methyltransferase | [36] |
| Hydralazine hydrochloride (3) | Inhibition of DNA methyltransferase | [13] |
| N-Acetyl-D-glucosamine (4) | Inhibition of DNA methyltransferase | [25] |
| Procainamide (5) | Inhibition of DNA methyltransferase | [32] |
| Procaine (6) | Inhibition of DNA methyltransferase | [37] |
| N-Phthalyl-L-tryptophan (7) | Inhibition of DNA methyltransferase | [38] |
| Entinostat (8) | Inhibition of HDAC of class I | [39] |
| Octanoylhydroxamic acid (9) | Inhibition of HDAC of classes I and II | [40] |
| Suberoylanilide hydroxamic acid (10) | Inhibition of HDAC of classes I and II | [41,42] |
| Suberoylbishydroxamic acid (11) | Inhibition of HDAC of classes I and II | [42] |
| Trichostatin A (12) | Inhibition of HDAC of classes I and II | [43] |
| Sodium butyrate (13) | Inhibition of HDAC of classes I and II | [25] |
| Sodium valproate (14) | Inhibition of HDAC of classes I and II | [44] |
| Valproic acid (15) | Inhibition of HDAC of classes I and II | [44] |
| Dihydrocoumarin (16) | Inhibition of NAD+-dependent HDAC of class III | [45] |
| 5-Methylmellein (17) | Inhibition of NAD+-dependent HDAC of class III | [46] |
| Mellein (18) | Inhibition of NAD+-dependent HDAC of class III | [46] |
| Nicotinamide (19) | Inhibition of NAD+-dependent HDAC of class III | [47] |
| Quercetin (20) | Inhibition of NAD+-dependent HDAC of class III; inhibition of protein kinases; inhibition of DNA topoisomerases; regulation of gene expression | [18] |
| 2-Hexyl-4-pentynoic acid (21) | Inhibition of histone deacetylase | [48] |
| Anacardic acid (22) | Inhibition of histone acetyltransferase | [31] |
| BRD4770 (23) | Inhibition of histone methytransferase | [32] |
| Bortezomib (24) | Inhibition of proteasome | [33] |
| NPD938 (25) | The action mechanism was not clear | [34,49] |
3. Effects of DNA Methyltransferase Modifiers
3.1. Effects of 5-Azacytidine
3.2. Effects of Other DNA Methyltransferase Modifiers
3.3. Effects of Combinational Treatment with Two DNA Methyltransferase Modifiers
4. Effects of Histone Deacetylase Modifiers
4.1. Effects of Suberoylanilide Hydroxamic Acid
4.2. Effects of Suberoylbishydroxamic Acid
4.3. Effects of Valproic Acid and Sodium Valproate
4.4. Effects of Sodium Butyrate
4.5. Effects of Nicotinamide
4.6. Effects of Trichostatin A
4.7. Effects of Other Histone Deacetylase Modifiers
4.8. Effects of Combinational Treatment with Two Histone Deacetylase Modifiers
5. Effects of Other Chemical Epigenetic Modifiers
5.1. Effects of Histone Acetyltransferase Modifier Anacardic Acid
5.2. Effects of Histone Methyltransferase Modifier BRD4770
5.3. Effects of Proteasome Modifier Bortezomib
5.4. Effects of the ModifierNPD938 with Unclear Mechanisms
6. Effects of Two Types of Chemical Epigenetic Modifiers
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; The International Natural Product Sciences Taskforce; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism–from biochemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Hautbergue, T.; Jamin, E.L.; Debrauwer, L.; Puel, O.; Oswald, I.P. From genomics to metabolomics, moving toward an integrated strategy for the discovery of fungal secondary metabolites. Nat. Prod. Rep. 2018, 35, 147–173. [Google Scholar] [CrossRef]
- Zhang, X.; Hindra; Elliot, M.A. Unlocking the trove of metabolic treasures: Activating silent biosynthetic gene clusters in bacteria and fungi. Curr. Opin. Microbiol. 2019, 51, 9–15. [Google Scholar] [CrossRef]
- Macheleidt, J.; Mattern, D.J.; Fischer, J.; Netzker, T.; Weber, J.; Schroeckh, V.; Valiante, V.; Brankhage, A.A. Regulation and role of fungal secondary metabolites. Annu. Rev. Genet. 2016, 50, 371–392. [Google Scholar] [CrossRef]
- Okada, B.K.; Seyedsayamdost, M.R. Antibiotic dialogues: Induction of silent biosynthetic gene clusters by exogenous small molecules. FEMS Microbiol. Rev. 2017, 41, 19–33. [Google Scholar] [CrossRef] [Green Version]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef]
- Lyu, H.-N.; Liu, H.-W.; Keller, N.P.; Yin, W.-B. Harnessing diverse transcriptional regulators for natural product discovery in fungi. Nat. Prod. Rep. 2020, 37, 6–16. [Google Scholar] [CrossRef]
- Pinto, A.A.; Barua, J.E.; Almeida, M.O.; Viaud, M.; Zorrilla, D.; Collado, I.G.; Macias-Sanchez, A.J.; Duran-Patron, R. Structural and biosynthetic studies of botrycinereic acid, a new cryptic metabolite from the fungus Botrytis cinerea. Bioorg. Chem. 2022, 127, 105979. [Google Scholar] [CrossRef]
- Reyes, F.; Bills, G.F.; Duran-Patron, R. Editorial: Strategies for the discovery of fungal natural products. Front. Microbiol. 2022, 13, 897756. [Google Scholar] [CrossRef] [PubMed]
- Berger, S.L.; Kouzarides, T.; Shiekhattar, R.; Shilatifard, A. An operational definition of epigenetics. Genes Dev. 2009, 23, 781–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cichewicz, R.H. Epigenome manipulation as a pathway to new natural product scaffolds and their congeners. Nat. Prod. Rep. 2010, 27, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Menendez, V.; Perez-Bonilla, M.; Perez-Victoria, I.; Mrtin, J.; Munoz, F.; Reyes, F.; Tormo, J.R.; Genilloud, O. Multicomponent analysis of the differential induction of secondary metabolite profiles in fungal endophytes. Molecules 2016, 21, 234. [Google Scholar] [CrossRef] [Green Version]
- Toghueo, K.R.M.; Dinkar, S.; Boyom, F.F. Stimulation of the production of new volatile and nonvolatile metabolites by endophytic Aspergillus niger using small organic chemicals. Curr. Res. Environ. Appl. Mycol. 2016, 6, 256–267. [Google Scholar] [CrossRef]
- Li, C.-Y.; Chung, Y.-M.; Wu, Y.-C.; Hunyadi, A.; Wang, C.C.C.; Chang, F.-R. Natural products development under epigenetic modulation in fungi. Phytochem. Rev. 2020, 19, 1323–1340. [Google Scholar] [CrossRef]
- Pocas-Fonseca, M.J.; Cabral, C.G.; Manfrao-Netto, J.H.C. Epigenetic manipulation of filamentous fungi for biotechnological applications: A systematic review. Biotechnol. Lett. 2020, 42, 885–904. [Google Scholar] [CrossRef]
- Toghueo, R.M.K.; Sahal, D.; Boyom, F.F. Recent advances in inducing endophytic fungal specialized metabolites using small molecule elicitors including epigenetic modifiers. Phytochemistry 2020, 174, 112338. [Google Scholar] [CrossRef]
- Pillay, L.C.; Nekati, L.; Makhwitine, P.J.; Ndlovu, S. Epigenetic activation of silent biosynthetic gene clusters in endophytic fungi using small molecular modifiers. Front. Microbiol. 2022, 13, 815008. [Google Scholar] [CrossRef]
- An, P.; Zhang, L.-J.; Peng, W.; Chen, Y.-Y.; Liu, Q.-P.; Luan, X.; Zhang, H. Natural products are an important source for proteasome regulating agents. Phytomedicine 2021, 3, 153799. [Google Scholar] [CrossRef]
- Bharatiya, P.; Rathod, P.; Hiray, A.; Kate, A.S. Multifarious elicitors: Invoking biosynthesis of various bioactive secondary metabolite in fungi. Appl. Biochem. Biotechnol. 2021, 193, 668–686. [Google Scholar] [CrossRef] [PubMed]
- Juarez-Mercado, K.E.; Prieto-Martinez, F.D.; Sanchez-Cruz, N.; Pena-Castillo, A.; Prada-Gracia, D.; Medina-Franco, J.L. Expanding the structural diversity of DNA methyltransferase inhibitors. Phrmaceuticals 2021, 14, 17. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Hui, Z.; Xu, L.; Bai, R.; Gao, Y.; Wang, Z.; Xie, T.; Ye, X.-Y. Medicinal chemistry updates of novel HDACs inhibitors (2020 to present). Eur. J. Med. Chem. 2022, 227, 113946. [Google Scholar] [CrossRef] [PubMed]
- Lyko, F.; Brown, R. DNA methyltransferase inhibitors and the development of epigenetic cancer therapies. J. Natl. Cancer Inst. 2005, 97, 1498–1506. [Google Scholar] [CrossRef] [Green Version]
- Zutz, C.; Gacek, A.; Sulyok, M.; Wagner, M.; Strauss, J.; Rychli, K. Small chemical chromatin effectors alter secondary metabolite production in Aspergillus clavatus. Toxins 2013, 5, 1723–1741. [Google Scholar] [CrossRef] [Green Version]
- Williams, R.B.; Henrikson, J.C.; Hoover, A.R.; Lee, A.E.; Cichewicz, R.H. Epigenetic remodeling of the fungal secondary metabolome. Org. Biomol. Chem. 2008, 6, 1895–1897. [Google Scholar] [CrossRef]
- Akiyama, D.Y.; Rocha, M.C.; Costa, J.H.; Malavazi, I.; Fill, T.P. The Penicillium brasilianum histone deacetylase Clr3 regulates secondary metabolite production and tolerance to oxidative stress. J. Fungi 2022, 8, 514. [Google Scholar] [CrossRef]
- Shwab, E.K.; Bok, J.W.; Tribus, M.; Galehr, J.; Graessle, S.; Keller, N.P. Histone deacetylase activity regulates chemical diversity in Aspergillus. Eukaryot. Cell 2007, 6, 1656–1664. [Google Scholar] [CrossRef] [Green Version]
- Qiu, X.; Zhu, L.; Wang, H.; Tan, Y.; Yang, Z.; Yang, L.; Wan, L. From natural products to HDAC inhibitors: An overview of drug discovery and design strategy. Bioorg. Med. Chem. 2021, 52, 116510. [Google Scholar] [CrossRef]
- Zutz, C.; Bacher, M.; Parich, A.; Kluger, B.; Gacek-Mattews, A.; Schuhmacher, R.; Wagner, M.; Rychli, K.; Strauss, J. Valproic acid induces antimicrobial compound production in Doratomyces microspores. Front. Microbiol. 2016, 7, 510. [Google Scholar] [CrossRef]
- Mafezoli, J.; Xu, Y.; Hilario, F.; Freidhof, B.; Espinosa-Artiles, P.; Dos Santos, L.C.; Gunatilaka, A.A.L. Modulation of polyketide biosynthetic pathway of the endophytic fungus, Anteaglonium sp. FL0768, by copper (II) and anacardic acid. Phytochem. Lett. 2018, 28, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Nishad, J.H.; Singh, A.; Bharti, R.; Prajapati, P.; Sharma, V.K.; Gupta, V.K.; Kharvar, R.N. Effect of the histone methyltransferase specific probe BRD4770 on metabolic profiling of the endophytic fungus Diaporthe longicolla. Front. Microbiol. 2021, 12, 725463. [Google Scholar] [CrossRef] [PubMed]
- Vander Moen, K.M.; Darveaux, B.A.; Chen, W.-L.; Swanson, S.M.; Pearce, C.J.; Oberlies, N.H. Epigenetic manipulation of a filamentous fungus by the proteasome-inhibitor bortezomib induces the production of an additional secondary metabolite. RSC Adv. 2014, 4, 18329–18335. [Google Scholar] [CrossRef]
- Motoyama, T.; Ishii, T.; Kamakura, T.; Osada, H. Screening of tenuazonic acid production-inducing compounds and identification of NPD938 as a regulator of fungal secondary metabolism. Biosci. Biotechnol. Biochem. 2021, 85, 2200–2208. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sena Fiho, J.G.; Hoover, A.R.; King, J.B.; Ellis, T.K.; Powell, D.R.; Cichewicz, R.H. Chemical epigenetics alters the secondary metabolite composition of guttate excreted by an atlantic-forest-soil-derived Penicillium citreonigrum. J. Nat. Prod. 2010, 73, 942–948. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.-Z.; Liang, B.-W.; Li, X.-F.; Liu, Q. Induced production of new diterpenoids in the fungus Penicillium funiculosum. Nat. Prod. Commun. 2014, 9, 607–608. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.-C.; Bao, H.-Y.; Liu, Y.-Y.; Nie, Y.-Y.; Yang, J.-M.; Hong, P.-Z.; Zhang, Y. Depsidone derivatives and a cyclopeptide produced by marine fungus Aspergillus unguis under chemical induction and by its plasma induced mutant. Molecules 2018, 23, 2245. [Google Scholar] [CrossRef] [Green Version]
- Asai, T.; Chung, Y.-M.; Sakurai, H.; Ozeki, T.; Chang, F.-R.; Wu, Y.-C.; Yamashita, K.; Oshima, Y. Highly oxidized ergosterols and isariotin analogs from an entomopathogenic fungus, Gibellula formosana, cultivated in the presence of epigenetic modifying agents. Tetrahedron 2012, 68, 5817–5823. [Google Scholar] [CrossRef]
- Bele, S.; Girada, S.B.; Ray, A.; Gupta, A.; Oruganti, S.; Babu, P.P.; Rayalla, R.S.; Kalivendi, S.V.; Ibrahim, A.; Puri, V.; et al. MS-275, a class 1 histone deacetylase inhibitor augments glucagon-like peptide- 1 receptor agonism to improve glycemic control and reduce obesity in diet-induced obese mice. eLife 2020, 9, e52212. [Google Scholar] [CrossRef]
- Siless, G.E.; Gallardo, G.L.; Rodriguez, M.A.; Rincon, Y.A.; Godeas, A.M.; Cabrera, G.M. Metabolites from the dark septate endophyte Drechslera sp. evaluation by LC/MS and principal component analysis of culture extracts with histone deacetylase inhibitors. Chem. Biodivers. 2018, 15, e1800133. [Google Scholar] [CrossRef]
- Henrikson, J.C.; Hoover, A.R.; Joyner, P.M.; Cichewicz, R.H. A chemical epigenetics approach for engineering the in situ biosynthesis of a cryptic natural product from Aspergillus niger. Org. Biomol. Chem. 2009, 7, 435–438. [Google Scholar] [CrossRef] [PubMed]
- Vervoort, H.C.; Draskovic, M.; Crews, P. Histone deacetylase inhibitors as a tool to up-regulate new fungal biosynthetic products: Isolation of EGM-556, a cyclodepsipeptide, from Microascus sp. Org. Lett. 2011, 13, 410–413. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Rodriguez, E.Y.; Uresti-Rvera, E.E.; Patron-Soberano, O.A.; Islas-Osuna, M.A.; Flores-Martinez, A.; Riego-Ruiz, L.; Rosales-Saavedra, M.T.; Casas-Flores, S. Histone acetyltransferase TGF-1 regulates Trichoderma atroviride secondary metabolism and mycoparasitism. PLoS ONE 2018, 13, e0193872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triastuti, A.; Vansteelandt, M.; Barakat, F.; Trinel, M.; Jargeat, P.; Fabre, N.; Guerra, C.A.A.; Mejia, K.; Valentin, A.; Valentin, A.; et al. How histone deacetylase inhibitors alter the secondary metabolites of Botryosphaeria mamane, an endophytic fungus isolated from Bixa orellana. Chem. Biodiversity 2019, 16, e1800485. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhou, Y.; Mao, Z.; Li, H.; Chen, F.; Shao, Y. NAD+-dependent HDAC inhibitor stimulates Monascus pigment production but inhibit citrinin. AMB Expr. 2017, 7, 166. [Google Scholar] [CrossRef] [Green Version]
- Shigemoto, R.; Matsumoto, T.; Masuomoto, T.; Takaya, N. 5-Methylmellein is a novel inhibitor of fungal sirtuin and modulates fungal secondary metabolite production. J. Gen. Appl. Microbiol. 2018, 64, 240–247. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.M.; Bradshaw, E.; Seipke, R.F.; Hutchings, M.I.; McArthur, M. Use and discovery of chemical elicitors that stimulate biosynthetic gene clusters in Streptomyces bacteria. Methods Enzymol. 2012, 517, 367–385. [Google Scholar]
- Zhu, J.; Wang, T.; Zhou, K.; Ding, L.; Wu, X.; He, S. Epigenetic manipulation regulates secondary metabolites from the marine-derived fungus Aspergillus versicolor DJ013. Chin. J. Mar. Drugs 2017, 36, 14–18. [Google Scholar]
- Abdelhakim, I.A.; Mahmud, F.B.; Motoyama, T.; Futamura, Y.; Takahashi, S.; Osada, H. Dihydrolucilactaene, a potent antimalarial compound from Fusarium sp. RK97-94. J. Nat. Prod. 2022, 85, 63–69. [Google Scholar] [CrossRef]
- Gupta, S.; Kulkarni, M.G.; White, J.F.; Van Staden, J. Epigenetic-based developments in the field of plant endophytic fungi. S. Afr. J. Bot. 2020, 134, 394–400. [Google Scholar] [CrossRef]
- Sun, J.; Awakawa, T.; Noguchi, H.; Abe, I. Induced production of mycotoxins in an endophytic fungus from the medicinal plant Datura stramonium L. Bioorg. Med. Chem. Lett. 2012, 22, 6397–6400. [Google Scholar] [CrossRef]
- Wang, C.-Y.; Liu, Y.-F.; Cao, F.; Wang, C.-Y. Bisabolane-type sesquiterpenoids from a gorgonian-derived Aspergillus sp. fungus induced by DNA methyltransferase inhibitor. Chem. Nat. Compd. 2016, 52, 1129–1132. [Google Scholar] [CrossRef]
- Lin, J.-Q.; Zhao, X.X.; Zhi, Q.Q.; Zhao, M.; He, Z.-M. Transcriptomic profiling of Aspergillus flavus in response to 5-azacytidine. Fungal Genet. Biol. 2013, 56, 78–86. [Google Scholar] [CrossRef]
- Lin, J.-Q.; Zhao, X.-X.; Wang, C.-C.; Xie, Y.; Li, G.-H.; He, Z.-M. 5-Azacytidine inhibits aflatoxin biosynthesis in Aspergillus flavus. Ann. Microbiol. 2013, 63, 763–769. [Google Scholar] [CrossRef]
- Yang, K.; Zhuang, Z.; Zhang, F.; Song, F.; Zhong, H.; Ran, F.; Yu, S.; Xu, G.; Lan, F.; Wang, W. Inhibition of aflatoxin metabolism and growth of Aspergillus flavus in liquid culture by a DNA methylation inhibitor. Food Addit. Comtam. B 2014, 32, 554–563. [Google Scholar] [CrossRef]
- Wilkinson, J.R.; Kale, S.P.; Bhatnagar, D.; Yu, J.; Ehrlich, K.C. Expression profiling of non-aflatoxigenic Aspergillus parasiticus mutants obtained by 5-azacytosine treatment or serial mycelial transfer. Toxins 2011, 3, 932–948. [Google Scholar] [CrossRef] [Green Version]
- Chung, Y.-M.; Wei, C.K.; Chuang, D.-W.; El-Shazly, M.; Hsieh, C.T.; Asai, T.; Oshima, Y.; Hsieh, T.J.; Hwang, T.L.; Wu, Y.C.; et al. An epigenetic modifier enhances the production of anti-diabetic and anti-inflammatory sesquiterpenoids from Aspergillus sydowii. Bioorg. Med. Chem. 2013, 21, 3866–3872. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, X.; Tang, Y.; Liu, Y.; Zhao, Z.; Cui, H. New butanolide derivatives from the marine derived fungus Aspergillus terreus GZU-31-1 by chemical epigenetic manipulation. Nat. Prod. Res. 2022. [Google Scholar] [CrossRef]
- Vasanthakumari, M.M.; Jadhav, S.S.; Sachin, N.; Vinod, G.; Shweta, S.; Manjunatha, B.L.; Mohana Kumara, P.; Ravikanth, G.; Nataraja, K.N.; Uma Shaanker, R. Restoration of camptothecine production in attenuated endophytic fungus on re-inoculation into host plant and treatment with DNA methyltransferase inhibitor. World J. Microbiol. Biotechnol. 2015, 31, 1629–1639. [Google Scholar] [CrossRef]
- Akone, S.H.; Mandi, A.; Kurtan, T.; Hartmann, R.; Lin, W.; Daletos, G.; Proksch, P. Inducing secondary metabolite production by the endophytic fungus Chaetomium sp. through fungal-bacterial co-culture and epigenetic modification. Tetrahedron 2016, 72, 6340–6347. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.-Y.; Li, N.; Li, Y.; Zhang, R.-J.; Lin, P.; Liu, L.; Ye, H.-Y.; Wu, W.-S.; Zhao, M. An epigenetic modifier enhances the generation of anti-phytopathogenic compounds from the endophytic fungus Chaetomium globosporum of Euphorbia humifusa. Phytochemistry 2022, 203, 113426. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, W.; Shao, C.-L.; Chi, Z.-M.; Wang, C.-Y. DNA methyltransferase inhibitor induced fungal biosynthetic products: Diethylene glycol phthalate ester oligomers from the marine-derived fungus Cochliobolus lunatus. Mar. Biotechnol. 2016, 18, 409–417. [Google Scholar] [CrossRef]
- Wu, J.-S.; Shi, X.-H.; Zhang, Y.-H.; Yu, J.-Y.; Fu, X.-M.; Li, X.; Chen, K.-X.; Guo, Y.-W.; Shao, C.-Y.; Wang, C.-Y. Co-cultivation with 5-azacytidine induced new metabolites from the zoanthid-derived fungus Cochliobolus lunatus. Front. Chem. 2019, 7, 763. [Google Scholar] [CrossRef] [Green Version]
- Pacheco-Tapia, R.; Vasquez-Ocmín, P.; Duthen, S.; Ortiz, S.; Jargeat, P.; Amasifuen, C.; Haddad, M.; Vansteelandt, M. Chemical modulation of the metabolism of an endophytic fungal strain of Cophinforma mamane using epigenetic modifiers and amino-acids. Fungal Biol. 2022, 126, 385–394. [Google Scholar] [CrossRef]
- Asai, T.; Yamamoto, T.; Chung, Y.-M.; Chang, F.-R.; Wu, Y.-C.; Yamashita, K.; Oshima, Y. Aromatic polyketide glycosides from an entomopathogenic fungus, Cordyceps indigotica. Tetrahedron Lett. 2012, 53, 277–280. [Google Scholar] [CrossRef]
- Deepika, V.B.; Vohra, M.; Mishra, S.; Dorai, K.; Rai, P.; Satyamoorthy, K.; Murali, T.S. DNA demethylation overcomes attenuation of colchicine biosynthesis in an endophytic fungus Diaporthe. J. Biotechnol. 2020, 323, 33–41. [Google Scholar] [CrossRef]
- Gonzalez-Menendez, V.; Crespo, G.; Toro, C.; Martin, J.; de Pedro, N.; Tormo, J.R.; Genilloud, O. Extending the metabolite diversity of the endophyte Dimorphosporicola tragani. Metabolites 2019, 9, 197. [Google Scholar] [CrossRef] [Green Version]
- Gakuubi, M.M.; Ching, K.C.; Munusamy, M.; Wibowo, M.; Liang, Z.-X.; Kanagasundaram, Y.; Ng, S.B. Enhancing the discovery of bioactive secondary metabolites from fungal endophytes using chemical elicitation and variation of fermentation media. Front. Microbiol. 2022, 13, 898976. [Google Scholar] [CrossRef]
- Qadri, M.; Nalli, Y.; Jain, S.K.; Chaubey, A.; Ali, A.; Strobel, G.A.; Vishwakarma, R.A.; Riyaz-Ul-Hassan, S. An insight into the secondary metabolism of Muscodor yucatanensis: Small molecule epigenetic modifiers induce expression of secondary metabolism-related genes and production of new metabolites in the endophyte. Microb. Ecol. 2017, 73, 954–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Wang, Q.; Liu, S.; Zhang, X.; Che, Q.; Zhang, G.; Zhu, T.; Gu, Q.; Li, D. Methylsulfonylated polyketides produced by Neosartorya udagawae HDN13-313 via exogenous addition of small molecules. J. Nat. Prod. 2019, 82, 998–1001. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.-Y.; Zhang, Q.; Gao, Y.-Q.; Zhang, A.-L.; Gao, J.-M. Miniolins A–C, novel isomeric furanones induced by epigenetic manipulation of Penicillium minioluteum. RSC Adv. 2015, 5, 2185–2190. [Google Scholar] [CrossRef]
- He, X.; Zhang, Z.; Chen, Y.; Che, Q.; Zhu, T.; Gu, Q.; Li, D. Varitatin A, a highly modified fatty acid amide from Penicillium variabile cultured with a DNA methyltransferase inhibitor. J. Nat. Prod. 2015, 78, 2841–2845. [Google Scholar] [CrossRef]
- Yang, X.L.; Huang, L.; Ruan, X.L. Epigenetic modifiers alter the secondary metabolite composition of a plant endophytic fungus, Pestalotiopsis crassiuscula obtained from the leaves of Fragaria chiloensis. J. Asian Nat. Prod. Res. 2014, 16, 412–417. [Google Scholar] [CrossRef]
- Yang, D.; Liu, F.; Yang, X. DNA methyltransferase inhibitor dramatically alters the secondary metabolism of Pestalotiopsis microspora. J. Chin. Pharm. Sci. 2017, 26, 355–359. [Google Scholar] [CrossRef]
- Dwibedi, V.; Kalia, S.; Saxena, S. Isolation and enhancement of resveratrol production in Xylaria psidii by exploring the phenomenon of epigenetics: Using DNA methyltransferases and histone deacetylase as epigenetic modifiers. Mol. Biol. Rep. 2019, 46, 4123–4137. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.A.; Bacha, N.; Ahmad, B.; Bakht, J.; Lutfullah, G.; Ali, J. Synthesis of secondary metabolites by Cladosporium resinae (NRL-6437) under different growth media and chemical inducers and their pharmaceutical activity. Pak. J. Pharm Sci. 2017, 30, 1617–1624. [Google Scholar] [PubMed]
- Ul-Hassan, S.R.; Strobel, G.A.; Booth, E.; Knighton, B.; Floerchinger, C.; Sears, J. Modulation of volatile organic compound formation in the mycodiesel-producing endophyte Hypoxylon sp. CI-4. Microbiology 2012, 158, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Kritsky, M.S.; Flippovich, S.Y.; Afanasieva, T.P.; Bachurina, G.P.; Russo, V.E.A. Effect of inhibitors of enzymatic DNA methylation on the formation of reproductive structures and carotenoid production in Neurospora crassa. Appl. Biochem. Microbiol. 2001, 37, 243–247. [Google Scholar] [CrossRef]
- Guo, D.-L.; Qiu, L.; Feng, D.; He, X.; Li, X.-H.; Cao, Z.X.; Gu, Y.-C.; Mei, L.; Deng, F.; Deng, Y. Three new α-pyrone derivatives induced by chemical epigenetic manipulation of Penicillium herquei, an endophytic fungus isolated from Cordyceps sinensis. Nat. Prod. Res. 2020, 34, 958–964. [Google Scholar] [CrossRef]
- Lei, H.; Zhang, D.; Ding, N.; Chen, S.; Song, C.; Luo, Y.; Fu, X.; Bi, X.; Niu, H. New cytotoxic natural products from the marine sponge-derived fungus Pestalotiopsis sp. by epigenetic modification. RSC Adv. 2020, 10, 37982–37988. [Google Scholar] [CrossRef]
- De Amorim, M.R.; Wijeratne, E.M.K.; Zhou, S.; Arnold, A.E.; Batista, A.N.L.; Batista, J.M.; Dos Santos, L.C. An epigenetic modifier induces production of 3-(4-oxopyrano)-chromen-2-ones in Aspergillus sp. AST0006, an endophytic fungus of Astragalus lentiginosus. Tetrahedron 2020, 76, 131525. [Google Scholar] [CrossRef] [PubMed]
- Aldholmi, M.; Wilkinson, B.; Ganesan, A. Epigenetic modulation of secondary metabolite profiles in Aspergillus calidoustus and Aspergillus westerdijkiae through histone deacetylase (HDAC) inhibition by vorinostat. J. Antibiot. 2020, 73, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Albright, J.C.; Henke, M.T.; Soukup, A.A.; McClure, R.A.; Thomson, R.J.; Keller, N.P.; Kelleher, N.L. Large-scale metabolomics reveals a complex response of Aspergillus nidulans to epigenetic perturbation. ACS Chem. Biol. 2015, 10, 1535–1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henke, M.; Soukup, A.A.; Goering, A.W.; McClure, R.A.; Thomason, R.J.; Keller, N.P.; Kelleher, N.L. New aspercryptins, lipopeptide natural products, revealed by HDAC inhibition in Aspergillus nidulans. ACS Chem. Biol. 2016, 11, 2117–2123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Yin, Y.; Sun, W.; Zhang, F.; Li, Z. Enhanced production of (+)-terrein by Aspergillus terreus strain PF26 with epigenetic modifier suberoylanilide hydroxamic acid. Process Biochem. 2013, 48, 1635–1639. [Google Scholar] [CrossRef]
- Wu, J.-S.; Shi, X.-H.; Zhang, Y.-H.; Shao, C.-L.; Fu, X.-M.; Li, X.; Yao, G.-S.; Wang, C.-Y. Benzyl furanones and pyrones from the marine-derived fungus Aspergillus terreus induced by chemical epigenetic modification. Molecules 2020, 25, 3927. [Google Scholar] [CrossRef]
- Zhu, J.X.; Ding, L.; He, S. Discovery of a new biphenyl derivative by epigenetic manipulation of marine-derived fungus Aspergillus versicolor. Nat. Prod. Res. 2019, 33, 1191–1195. [Google Scholar] [CrossRef]
- Liu, W.; Wang, L.; Wang, B.; Xu, Y.; Zhu, G.; Lan, M.; Zhu, W.; Sun, K. Diketopiperazine and diphenylether derivatives from marine algae-derived Aspergillus versicolor OUCMDZ-2738 by epigenetic activation. Mar. Drugs 2019, 17, 6. [Google Scholar] [CrossRef] [Green Version]
- Miao, F.-P.; Liang, X.R.; Liu, X.-H.; Ji, N.-Y. Aspewentins A-C, norditerpenes from a cryptic pathway in an algicolous strain of Aspergillus wentii. J. Nat. Prod. 2014, 77, 429–432. [Google Scholar] [CrossRef]
- Frisvad, J.C. A critical review of producers of small lactone mycotoxins: Patulin, penicillic acid and moniliformin. World Mycotoxin J. 2018, 11, 73–100. [Google Scholar] [CrossRef]
- Igboeli, H.; Marchbank, D.H.; Correa, H.; Overy, D.; Kerr, R.G. Discovery of primarolides A and B from marine fungus Asteromyces cruciatus using osmotic stress and treatment with suberoylanilide hydroxamic acid. Mar. Drugs 2019, 17, 435. [Google Scholar] [CrossRef] [Green Version]
- Chung, Y.-M.; El-Shazly, M.; Chuang, D.W.; Hwang, T.L.; Asai, T.; Oshima, Y.; Ashour, M.L.; Wu, Y.C.; Chang, F.-R. Suberoylanilide hydroxamic acid, a histone deacetylase inhibitor, induces the production of anti-inflammatory cyclodepsipeptides from Beauveria felina. J. Nat. Prod. 2013, 76, 1260–1266. [Google Scholar] [CrossRef]
- Huang, L.; Wang, S.-L.; Xu, Y.-L.; Yu, H.-F.; Zhan, Z.-J.; Shan, W.-G.; Wang, J.-W.; Ying, Y.-M. Induced production of tremulane sesquiterpenoids in Bjerkandera adusta by chemical epigenetic modification. Chem. Nat. Compd. 2020, 56, 754–756. [Google Scholar] [CrossRef]
- Adpressa, D.; Stalheim, K.J.; Proteau, P.J.; Loesgen, S. Unexpected biotransformation of the HDAC inhibitor vorinostat yields aniline-containing fungal metabolites. ACS Chem. Biol. 2017, 12, 1842–1847. [Google Scholar] [CrossRef]
- Zhang, Z.; He, X.; Wu, G.; Liu, C.; Lu, C.; Gu, Q.; Che, Q.; Zhu, T.; Zhang, G.; Li, D. Aniline-tetramic acids from the deep-sea-derived fungus Cladosporium sphaerospermum L3P3 cultured with the HDAC inhibitor SAHA. J. Nat. Prod. 2018, 81, 1651–1657. [Google Scholar] [CrossRef]
- Chen, J.-J.; Han, M.-Y.; Gong, T.; Qiao, Y.-M.; Yang, J.-L.; Zhu, P. Epigenetic modification enhances ergot alkaloid production of Claviceps purpurea. Biotechnol. Lett. 2019, 41, 1439–1449. [Google Scholar] [CrossRef]
- Liu, S.-L.; Zhou, L.; Chen, H.-P.; Liu, J.-K. Sesquiterpenes with diverse skeletons from histone deacetylase inhibitor modified cultures of the basidiomycete Cyathus stercoreus (Schwein.) De Toni HFG134. Phytochemistry 2022, 195, 113048. [Google Scholar] [CrossRef]
- Du, L.; King, J.B.; Cichewicz, R.H. Chlorinated polyketide obtained from a Daldinia sp. treated with the epigenetic modifier suberoylanilide hydroxamic acid. J. Nat. Prod. 2014, 77, 2454–2458. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Yuan, L.-Y.; Gu, D.-L.; Ye, Y.; Da-Wa, Z.M.; Wang, X.-L.; Ma, F.-W.; Chen, L.; Gu, Y.-C.; Ding, L.-S.; et al. Bioactive halogenated dihydroisocoumarins produced by the endophytic fungus Lachnum palmae isolated from Przewalskia tangutica. Phytochemistry 2018, 148, 97–103. [Google Scholar] [CrossRef]
- Ying, Y.-M.; Li, L.; Yu, H.-F.; Xu, Y.-L.; Huang, L.; Mao, W.; Tong, C.-P.; Zhang, Z.-D.; Zhan, Z.-J.; Zhang, Y. Induced production of a new polyketide in Penicillium sp. HS-11 by chemical epigenetic manipulation. Nat. Prod. Res. 2021, 35, 3446–3451. [Google Scholar] [CrossRef]
- Li, C.; Shao, Y.; Li, W.; Yin, T.; Li, H.; Yan, H.; Guo, X.; Liu, B.; He, B. Hybrid diterpenic meroterpenoids from an endophytic Penicillium sp. induced by chemical epigenetic manipulation. J. Nat. Prod. 2022, 85, 1486–1494. [Google Scholar] [CrossRef]
- Zhang, S.; Fang, H.; Yin, C.; Wei, C.; Hu, J.; Zhang, Y. Antimicrobial metabolites produced by Penicillium mallochii CCH01 isolated from the gut of Ectropis oblique, cultivated in the presence of a histone deacetylase inhibitor. Front. Microbiol. 2019, 10, 2186. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Zhang, Z.; Che, Q.; Zhu, T.; Gu, Q.; Li, D. Varilactones and wortmannilactones produced by Penicillium variabile cultured with histone deacetylase inhibitor. Arch. Pharm. Res. 2018, 41, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Gubiani, J.R.; Wijeratne, E.M.K.; Shi, T.; Araujo, A.R.; Arnold, A.E.; Chapman, E.; Gunatilaka, A.A.L. An epigenetic modifier induces production of (10′S)-verruculide B, an inhibitor of protein tyrosine phosphatases by Phoma sp. nov. LG0217, a fungal endophyte of Parkinsonia microphylla. Bioorg. Med. Chem. 2017, 25, 1860–1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Meng, Q.; Niu, S.; Liu, J.; Guo, X.; Sun, Z.; Liu, D.; Gu, Y.; Huang, J.; Fan, A.; et al. Epigenetic manipulation to trigger production of guaiane-type sesquiterpenes from a marine-derived Spiromastix sp. fungus with antineuroinflammatory effects. J. Nat. Prod. 2021, 84, 1993–2003. [Google Scholar] [CrossRef]
- Wang, Z.; He, X.; Niu, C.; Zhou, J.; Zhang, Z. Induction of funitatin A, a new polyketide from the Yellow River wetland-derived fungus Talaromyces funiculosus. Phytochem. Lett. 2022, 47, 42–45. [Google Scholar] [CrossRef]
- Liu, W.-C.; Wang, Y.-Y.; Liu, J.-H.; Ke, A.-B.; Zheng, Z.-H.; Lu, X.-H.; Luan, Y.-S.; Xiu, Z.-L.; Dong, Y.-S. Wortmannilactones I–L, new NADH-fumarate reductase inhibitors, induced by adding suberoylanilide hydroxamic acid to the culture medium of Talaromyces wortmannii. Bioorg. Med. Chem. Lett. 2016, 26, 5328–5333. [Google Scholar] [CrossRef]
- Fisch, K.M.; Gillaspy, A.F.; Gipson, M.; Henrikson, J.C.; Hoover, A.R.; Jackson, L.; Najar, F.Z.; Wagele, H.; Cicherwicz, R.H. Chemical induction of silent biosynthetic pathway transcription in Aspergillus niger. J. Ind. Microbiol. Biotechnol. 2009, 36, 1199–1213. [Google Scholar] [CrossRef]
- Li, C.-Y.; Lo, I.-W.; Hsueh, Y.P.; Chung, Y.M.; Wang, S.W.; Korinek, M.; Tsai, Y.H.; Cheng, Y.B.; Hwang, T.L.; Wang, C.C.C.; et al. Epigenetic manipulation induces the production of coumarin-type secondary metabolite from Arthrobotrys foliicola. Isr. J. Chem. 2019, 59, 432–438. [Google Scholar] [CrossRef]
- Asai, T.; Yamamoto, T.; Shirata, N.; Taniguchi, T.; Monde, K.; Fuji, I.; Gomi, K.; Oshima, Y. Structurally diverse chaetophenol productions induced by chemically mediated epigenetic manipulation of fungal gene expression. Org. Lett. 2013, 15, 3346–3349. [Google Scholar] [CrossRef] [PubMed]
- Asai, T.; Taniguchi, T.; Yamamoto, T.; Monde, K.; Oshima, Y. Structures of spiroindicumides A and B, unprecedented carbon skeletal spirolactones, and determination of the absolute configuration by vibrational circular dichroism exciton approach. Org. Lett. 2013, 15, 4320–4323. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.-Z.; Liang, B.-W.; Li, X.-F.; Yu, Z.-Y. Epigenetic modifier-induced biosynthesis of novel acetylenic sterols from Cladosporium colocasiae. Steroids 2014, 87, 35–38. [Google Scholar] [CrossRef] [PubMed]
- Asai, T.; Luo, D.; Obara, Y.; Taniguchi, T.; Monde, K.; Yamashita, K.; Osima, Y. Dihydrobenzofurans as cannabinoid receptor ligands from Cordyceps annullata, an entomopathogenic fungus cultivated in the presence of an HDAC inhibitor. Tetrahedron Lett. 2012, 53, 2239–2243. [Google Scholar] [CrossRef]
- Asai, T.; Yamamoto, T.; Oshima, Y. Aromatic polyketide production in Cordyceps indigotica, an entomopathogenic fungus, induced by exposure to a histone deacetylase inhibitor. Org. Lett. 2012, 14, 2006–2009. [Google Scholar] [CrossRef] [PubMed]
- Niu, S.; Liu, D.; Shao, Z.; Proksch, P.; Lin, W. Eremophilane-type sesquiterpenoids in a deep-sea fungus Eutypella sp. activated by chemical epigenetic manipulation. Tetrahedron 2018, 74, 7310–7325. [Google Scholar] [CrossRef]
- Chen, H.J.; Awakawa, T.; Sun, J.Y.; Wakimoto, T.; Abe, I. Epigenetic modifier-induced biosynthesis of novel fusaric acid derivatives in endophytic fungi from Datura stramonium L. Nat. Prod. Bioprospect. 2013, 3, 20–23. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wei, T.; Luo, L.; Zhang, Q.; Yu, X.; Guo, Z.; He, H.; Liu, C.; Zou, K. Chemical epigenetic modification of symbiotic fungus Paraconiothyrium brasiliense. Biot. Resour. 2017, 39, 113–117. [Google Scholar]
- Sheng, S.-L.; Li, Y.-P.; Xiang, H.-Y.; Liu, Y.; Wang, Y.-D.; Kong, L.-P.; Du, G.; Hu, Q.-F.; Chen, Y.-J.; Wang, W.-G. Histone deacetylase inhibitor induced lipase inhibitors from endophytic Phomopsis sp. 0391. Rec. Nat. Prod. 2020, 14, 42–47. [Google Scholar] [CrossRef]
- Asai, T.; Yamamoto, T.; Oshima, Y. Histone deacetylase inhibitor induced the production of three novel prenylated tryptophan analogs in the entomopathogenic fungus, Torrubiella luteorostrata. Tetrahedron Lett. 2011, 52, 7042–7045. [Google Scholar] [CrossRef]
- Magotra, A.; Kumar, M.; Kushwaha, M.; Awasthi, P.; Raina, C.; Gupta, A.P.; Shah, B.A.; Gandhi, S.G.; Chaubey, A. Epigenetic modifier induced enhancement of fumiquinazoline C production in Aspergillus fumigatus (GA-L7): An endophytic fungus from Grewia asiatica L. AMB Expr. 2017, 7, 43. [Google Scholar] [CrossRef] [Green Version]
- Kunhorm, P.; Chueaphromsri, P.; Chicharoenaudomrung, N.; Noisa, P. Enhancement of cordycepin production from Cordyceps militaris culture by epigenetic modification. Biotechnol. Lett. 2022, 44, 581–593. [Google Scholar] [CrossRef]
- Sharma, V.; Singamaneni, V.; Sharma, N.; Kumar, A.; Arora, D.; Kushwaha, M.; Bhushan, S.; Jaglan, S.; Gupta, P. Valproic acid induces three novel cytotoxic secondary metabolites in Diaporthe sp., an endophytic fungus from Datura inoxia Mill. Bioorg. Med. Chem. Lett. 2018, 28, 2217–2221. [Google Scholar] [CrossRef] [PubMed]
- Jasim, B.; Sahadevan, N.; Chithra, S.; Mathew, J.; Radhakrishnan, E.K. Epigenetic modifier based enhancement of piperine production in endophytic Diaporthe sp. PF20. Proc. Natl. Acad. Sci. India Sec. B Biol. Sci. 2019, 89, 671–677. [Google Scholar] [CrossRef]
- Singh, G.; Kumar, A.; Verma, M.K.; Gupta, P.; Katoch, M. Secondary metabolites produced by Macrophomina phaseolina, a fungal root endophyte of Brugmansia aurea, using classical and epigenetic manipulation approach. Folia Microbiol. 2022, 67, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Zhen, X.; Gong, T.; Wen, Y.-H.; Yan, D.-J.; Chen, J.-J.; Zhu, P. Chrysoxanthones A–C, three new xanthone–chromanone heterdimers from sponge-associated Penicillium chrysogenum HLS111 treated with histone deacetylase inhibitor. Mar. Drugs 2018, 16, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ameen, F.; Almansob, A.; Tami, M.A.; Al-Enazi, N.; Al-Sabri, A.; Orfali, R. Epigenetic modifiers affect the bioactive compounds secreted by an endophyte of the tropical plant Piper longum. Molecules 2021, 26, 29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Shao, C.-L.; Chen, M.; Liu, Q.-A.; Wang, C.-Y. Brominated resorcylic acid lactones from the marine-derived fungus Cochliobolus lunatus induced by histone deacetylase inhibitors. Tetrahedron Lett. 2014, 55, 4888–4891. [Google Scholar] [CrossRef]
- Beau, J.; Mahid, N.; Burda, W.N.; Harrington, L.; Shaw, L.N.; Mutka, T.; Kyle, D.E.; Barisic, B.; Van Olphen, A.; Baker, B.J. Epigenetic tailoring for the production of anti-infective cytosporones from the marine fungus Leucostoma persoonii. Mar. Drugs 2012, 10, 762–774. [Google Scholar] [CrossRef]
- El-Hawary, S.; Sayed, A.M.; Mohammed, R.; Hassan, H.M.; Zaki, M.A.; Rateb, M.E.; Mohammed, T.A.; Amin, E.; Abdelmohsen, U.R. Epigenetic modifiers induce bioactive phenolic metabolites in the marine-derived fungus Penicillium brevicompactum. Mar. Drugs 2018, 16, 253. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Yan, L.; Xu, X.; Zhang, Y.; Shi, J.; Jiang, C.; Shao, D. Strategies to enhance the production of pinoresinol and its glucosides by endophytic fungus (Phomopsis sp. XP-8) isolated from Tu-chung bark. AMB Expr. 2018, 8, 55. [Google Scholar] [CrossRef]
- Feng, T.; Wei, C.; Deng, X.; Chen, D.; Wen, Z.; Xu, J. Epigenetic manipulation induced production of immunosuppressive chromones and cytochalasins from the mangrove endophytic fungus Phomopsis asparagi DHS-48. Mar. Drugs 2022, 20, 616. [Google Scholar] [CrossRef]
- Shi, T.; Shao, C.-Y.; Liu, Y.; Zhao, D.-L.; Cao, F.; Fu, X.-M.; Yu, J.-Y.; Wu, J.-S.; Zhang, Z.-K.; Wang, C.-Y. Terpenoids from the coral-derived fungus Trichoderma harzianum (XS-20090075) induced by chemical epigenetic manipulation. Front. Microbiol. 2020, 11, 572. [Google Scholar] [CrossRef] [PubMed]
- Lotfy, M.M.; Sayed, A.M.; AboulMagd, A.M.; Hassan, H.M.; El Amir, D.; Abouzid, S.F.; El-Gendy, A.O.; Rateb, M.E.; Abdelmohsen, U.R.; Alhadrami, H.; et al. Metabolomic profiling, biological evaluation of Aspergillus awamori, the river Nile-derived fungus using epigenetic and OSMAC approaches. RSC Adv. 2021, 11, 6709–6719. [Google Scholar] [CrossRef] [PubMed]
- Asai, T.; Morita, S.; Taniguchi, T.; Monde, K.; Oshima, Y. Epigenetic stimulation of polyketide production in Chaetomium cancroideum by an NAD+-dependent HDAC inhibitor. Org. Biomol. Chem. 2016, 14, 646–651. [Google Scholar] [CrossRef]
- Asai, T.; Morita, S.; Shirata, N.; Taniguchi, T.; Monde, K.; Sakurai, H.; Ozeki, T.; Oshima, Y. Structural diversity of new C13 -polyketides produced by Chaetomium mollipilium cultivated in the presence of a NAD+-dependent histone deacetylase inhibitor. Org. Lett. 2012, 14, 5456–5459. [Google Scholar] [CrossRef]
- Li, G.; Kusari, S.; Golz, C.; Laatsch, H.; Strohmann, C.; Spiteller, M. Epigenetic modulation of endophytic Eupenicillium sp. LG41 by a histone deacetylase inhibitor for production of decalin-containing compounds. J. Nat. Prod. 2017, 80, 983–988. [Google Scholar] [CrossRef]
- Sidiq, Y.; Nakano, M.; Mori, Y.; Yaeno, T.; Kimura, M.; Nishiuchi, T. Nicotinamide effectively suppresses Fusarium head blight in wheat plants. Int. J. Mol. Sci. 2021, 22, 2968. [Google Scholar] [CrossRef] [PubMed]
- Asai, T.; Otsuki, S.; Sakurai, H.; Yamashita, K.; Ozeki, T.; Oshima, Y. Benzophenones from an endophytic fungus, Graphiopsis chlorocephala, from Paeonia lactiflora cultivated in the presence of an NAD+-dependent HDAC inhibitor. Org. Lett. 2013, 15, 2058–2061. [Google Scholar] [CrossRef] [PubMed]
- Graessle, S.; Dangl, M.; Haas, H.; Mair, K.; Trojer, P.; Brandtner, E.-M.; Walton, J.D.; Loidl, P.; Brosch, G. Characterization of two putative histone deacetylase genes from Aspergillus nidulans. Biochim. Biophys. Acta 2000, 1492, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Zhu, G.; Hao, J.; Wang, Y.; Zhu, W. Chemical-epigenetic method to enhance the chemodiversity of the marine algicolous fungus, Aspergillus terreus OUCMDZ-2739. Tetrahedron 2018, 74, 83–87. [Google Scholar] [CrossRef]
- Lou, S.; Zhu, Y.; Ma, X.; Li, W.; Du, G.; Yang, H.; Wang, W. Epigenetic-induced secondary metabolites from the endophytic fungi Hypomyces sp. CLG4 isolated from Paris polyphylla var. yunnanensis. J. Yunnan Minzu Univ. (Nat. Sci. Ed.) 2021, 30, 17–21. [Google Scholar]
- Gulyamova, T.G.; Abdulmyanova, L.I.; Ruzieva, D.M.; Rasulova, G.A.; Yusupov, U.K.; Sattarova, R.S. Effect of epigenetic modifiers on fermentation parameters of endophytic fungi from plants growing in Uzbekistan. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 851–860. [Google Scholar] [CrossRef]
- Yang, X.L.; Awakawa, T.; Wakimoto, T.; Abe, I. Induced biosyntheses of a novel butyrophenone and two aromatic polyketides in the plant pathogen Stagonospora nodorum. Nat. Prod. Bioprospect. 2013, 3, 141–144. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, F.; Banakar, S.; Li, Z. Bortezomib-induced new bergamotene derivatives xylariterpenoids H–K from sponge-derived fungus Pestalotiopsis maculans 16F-12. RSC Adv. 2019, 9, 599–608. [Google Scholar] [CrossRef]
- Hemshekhar, M.; Santhosh, M.S.; Kemparaju, K.; Girish, K.S. Emerging roles of anacardic acid and its derivatives: A pharmacological overview. Basic Clin. Pharmacol. Toxicol. 2012, 110, 122–132. [Google Scholar] [CrossRef]
- Wang, L.; Li, M.; Tang, J.; Li, X. Eremophilane sesquiterpenes from a deep marine-derived fungus, Aspergillus sp. SCSIOW2, cultivated in the presence of epigenetic modifying agents. Molecules 2016, 21, 473. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xia, Z.; Tang, J.; Wu, J.; Tong, J.; Li, M.; Ju, J.; Chen, H.; Wang, L. Identification and biological evaluation of secondary metabolites from marine derived fungi-Aspergillus sp. SCSIOW3, cultivated in the presence of epigenetic modifying agents. Molecules 2017, 22, 1302. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Li, J.; Wei, C.; Deng, X.; Xu, J. Chemical epigenetic modifiers enhance the production of immunosuppressants from the endophytic fungus Aspergillus fumigatus isolated from Cynodon dactylon. Nat. Prod. Res. 2022, 36, 4481–4485. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.-S.; Yao, G.-S.; Shi, X.-H.; Ur Rehman, S.; Xu, Y.; Fu, X.-M.; Zhang, X.-L.; Liu, Y.; Wang, C.-Y. Epigenetic agents trigger the production of bioactive nucleoside derivatives and bisabolane sesquiterpenes from the marine-derived fungus Aspergillus versicolor. Front. Microbiol. 2020, 11, 85. [Google Scholar] [CrossRef] [Green Version]
- Niu, S.; Liu, D.; Shao, Z.; Liu, J.; Fan, A.; Lin, W. Chemical epigenetic manipulation triggers the production of sesquiterpenes from the deep-sea derived Eutypella fungus. Phytochemistry 2021, 192, 112978. [Google Scholar] [CrossRef]
- Asai, T.; Chung, Y.-M.; Sakurai, H.; Ozeki, T.; Chang, F.-R.; Yamashita, K.; Oshima, Y. Tenuipyrone, a novel skeletal polyketide from the entomopathogenic fungus, Isaria tenuipes, cultivated in the presence of epigenetic modifiers. Org. Lett. 2012, 14, 513–515. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.L.; Awakawa, T.; Wakimoto, T.; Abe, I. Induced production of novel prenyldepside and coumarins in endophytic fungi Pestalotiopsis acaciae. Tetrahedron Lett. 2013, 54, 5814–5817. [Google Scholar] [CrossRef]
- Yang, X.L.; Awakawa, T.; Wakimoto, T.; Abe, I. Induced production of the novel glycolipid ustilagic acid C in the plant pathogen Ustilago maydis. Tetrahedron Lett. 2013, 54, 3655–3657. [Google Scholar] [CrossRef]
- Pfannenstiel, B.T.; Keller, N.P. On top of biosynthetic gene clusters: How epigenetic machinery influences secondary metabolism in fungi. Bioetechnol. Adv. 2019, 37, 107345. [Google Scholar] [CrossRef] [PubMed]
- Bind, S.; Bind, S.; Sharma, A.K.; Chaturvedi, P. Epigenetic modification: A key tool for secondary metabolite production in microorganisms. Front. Microbiol. 2022, 13, 784109. [Google Scholar] [CrossRef]
- Pinedo-Rivilla, C.; Aleu, J.; Duran-Patron, R. Cryptic metabolites from marine-derived microorganisms using OSMAC and epigenetic approaches. Mar. Drugs 2022, 20, 84. [Google Scholar] [CrossRef]
- Yang, K.; Tian, J.; Keller, N.P. Post-translational modifications drive secondary metabolite biosynthesis in Aspergillus: A review. Environ. Microbiol. 2022, 24, 2857–2881. [Google Scholar] [CrossRef]
- Ansari, S.; Kumar, V.; Bhatt, D.N.; Irfan, M.; Datta, A. N-Acetylglucosamine sensing and metabolic engineering for attenuating human and plant pathogens. Bioengineering 2022, 9, 64. [Google Scholar] [CrossRef]
- Yan, Y.; Liao, Z.; Shen, J.; Zhu, Z.; Cao, Y. Nicotinamide potentiates amphotericin B activity against Candida albicans. Virulence 2022, 13, 1533–1542. [Google Scholar] [CrossRef]
- Mou, Y.; Luo, H.; Mao, Z.; Shan, T.; Sun, W.; Zhou, K.; Zhou, L. Enhancement of palmarumycins C12 and C13 production in liquid culture of endophytic fungus Berkleasmium sp. Dzf12 after treatments with metal ions. Int. J. Mol. Sci. 2013, 14, 979–998. [Google Scholar] [CrossRef]
- Luo, H.; Xu, D.; Xie, R.; Zhang, X.; Wang, J.; Dong, X.; Lai, D.; Zhou, L.; Liu, Y. Enhancement of botrallin and TMC-264 production in liquid culture of endophytic fungus Hyalodendriella sp. Ponipodef12 after treatments with metal ions. Electron. J. Biotechnol. 2016, 24, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Mou, Y.; Meng, J.; Fu, X.; Wang, X.; Tian, J.; Wang, M.; Peng, Y.; Zhou, L. Antimicrobial and antioxidant activities and effect of 1-hexadecene addition on palmarumycin C2 and C3 yields in liquid culture of endophytic fungus Berkleasmium sp. Dzf12. Moleulces 2013, 18, 15587–15599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mou, Y.; Xu, D.; Mao, Z.; Dong, X.; Lin, F.; Wang, A.; Lai, D.; Zhou, L.; Xie, B. Enhancement of palmarumycins C12 and C13 production by endophytic fungus Berkleasmium sp. Dzf12 in an aqueous-organic solvent system. Molecules 2015, 20, 20320–20333. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, M.; Hou, X.; Fu, J.; Zhang, J.; Wang, J.; Zhao, Z.; Xu, D.; Lai, D.; Zhou, L. Recent Advances in Search of Bioactive Secondary Metabolites from Fungi Triggered by Chemical Epigenetic Modifiers. J. Fungi 2023, 9, 172. https://doi.org/10.3390/jof9020172
Xue M, Hou X, Fu J, Zhang J, Wang J, Zhao Z, Xu D, Lai D, Zhou L. Recent Advances in Search of Bioactive Secondary Metabolites from Fungi Triggered by Chemical Epigenetic Modifiers. Journal of Fungi. 2023; 9(2):172. https://doi.org/10.3390/jof9020172
Chicago/Turabian StyleXue, Mengyao, Xuwen Hou, Jiajin Fu, Jiayin Zhang, Jiacheng Wang, Zhitong Zhao, Dan Xu, Daowan Lai, and Ligang Zhou. 2023. "Recent Advances in Search of Bioactive Secondary Metabolites from Fungi Triggered by Chemical Epigenetic Modifiers" Journal of Fungi 9, no. 2: 172. https://doi.org/10.3390/jof9020172