
Antifungal Activity and Biocontrol Potential of Simplicillium lamellicola JC-1 against Multiple Fungal Pathogens of Oilseed Rape

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Cultural Media
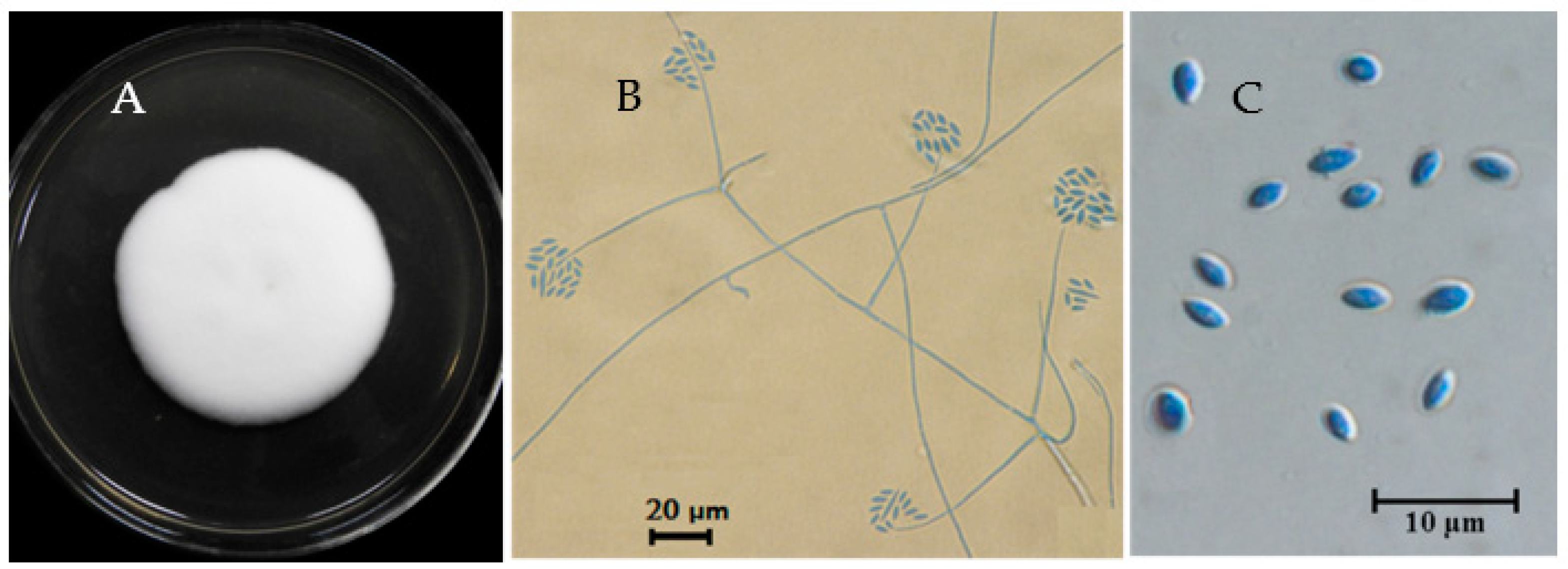
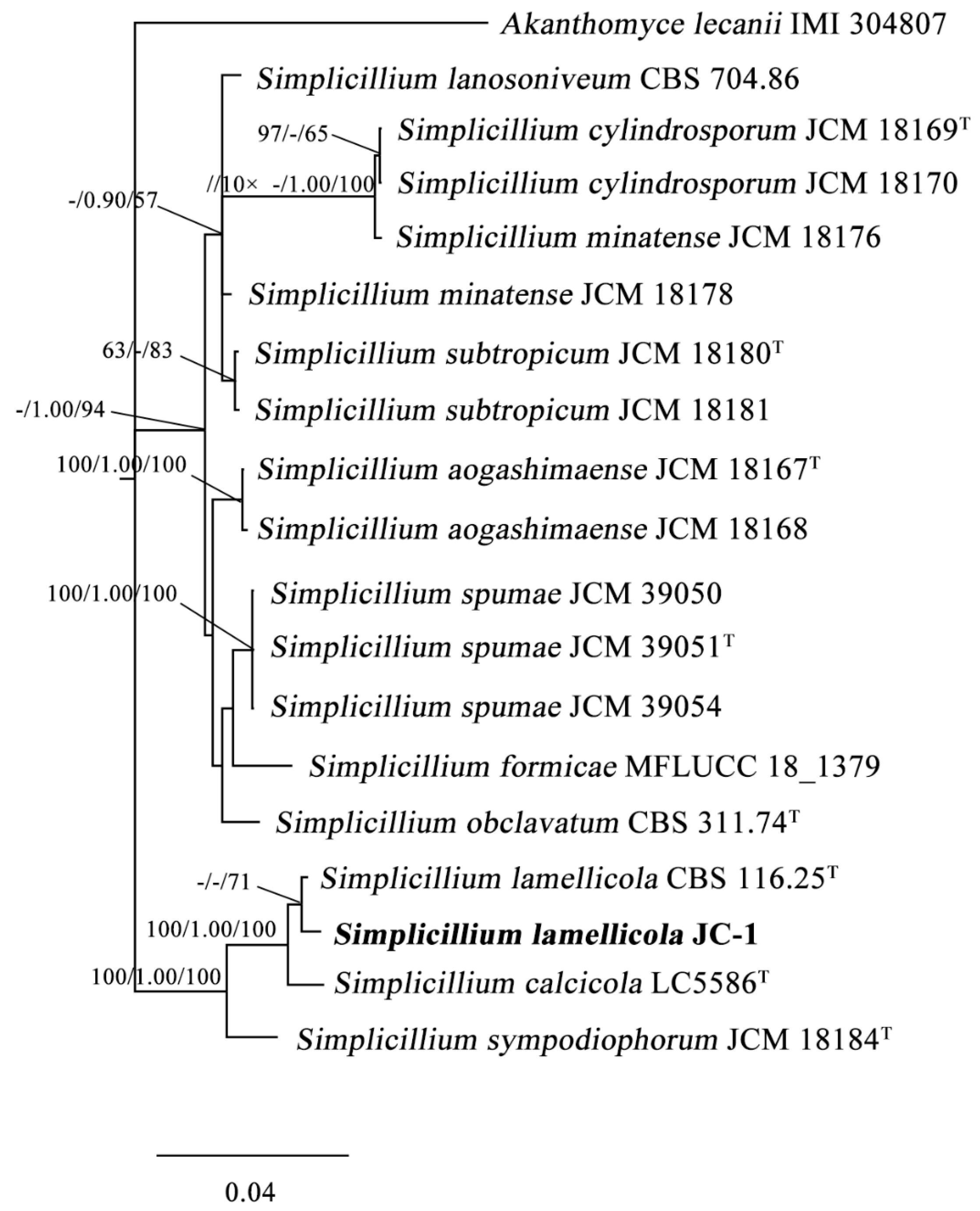
2.2. Fungal Identification
2.3. Genome Sequencing and Identification of Biosynthetic Gene Clusters for the Secondary Metabolites
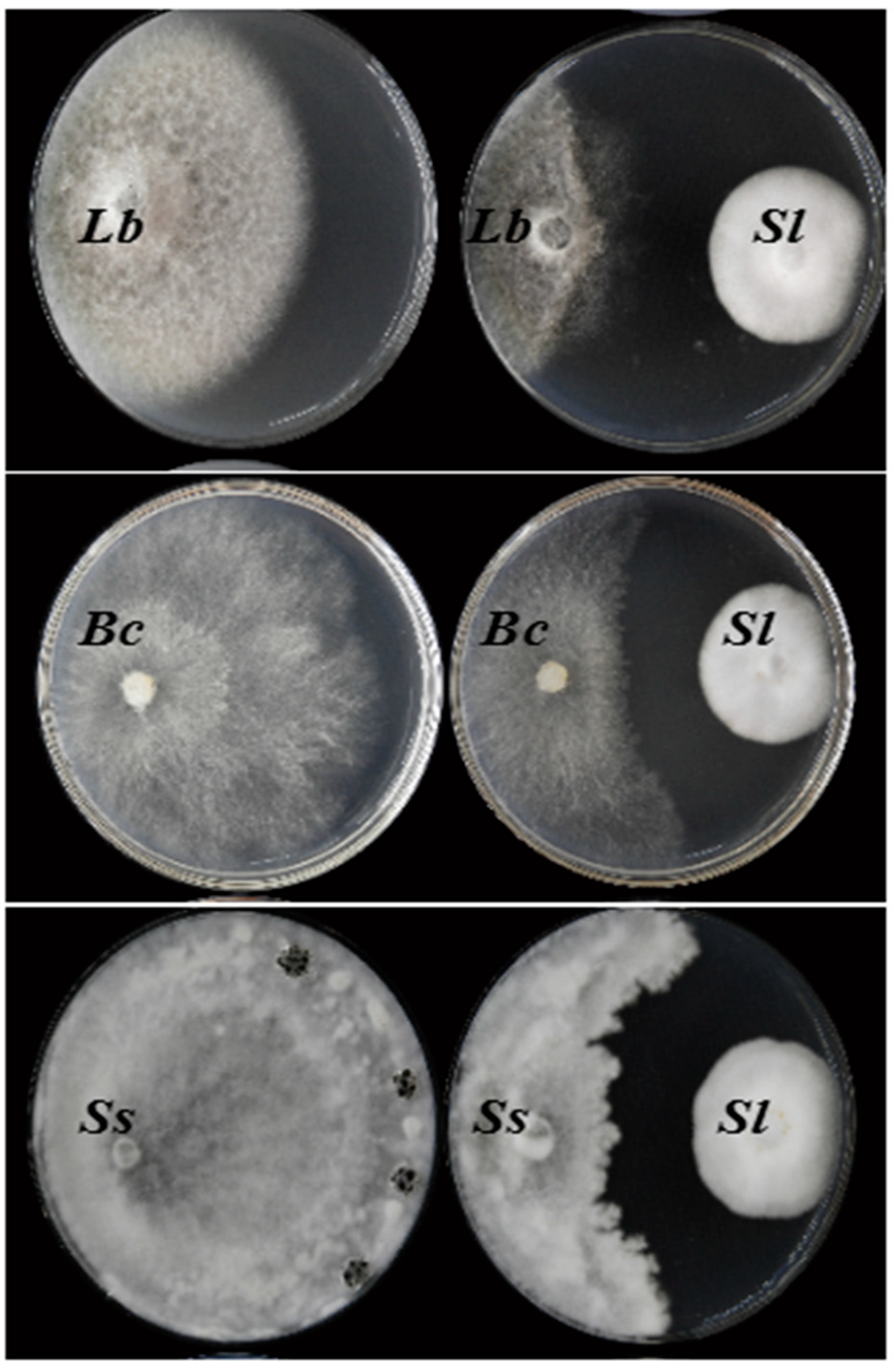
2.4. Dual Culturing
2.5. Determination of the Antifungal Activity of the JC-1 Cultures
2.6. Extraction of the Antifungal Metabolites
2.7. Determination of Control Efficacy of the Antifungal Metabolites
2.8. Detection of Endophytic Growth
2.9. Induction of Systemic Resistance
2.10. Quantification of Anthocyanins
2.11. Quantitative Detection of Expression of the Defense-Related Genes
2.12. Data Analysis
3. Results
3.1. Taxonomic Identity
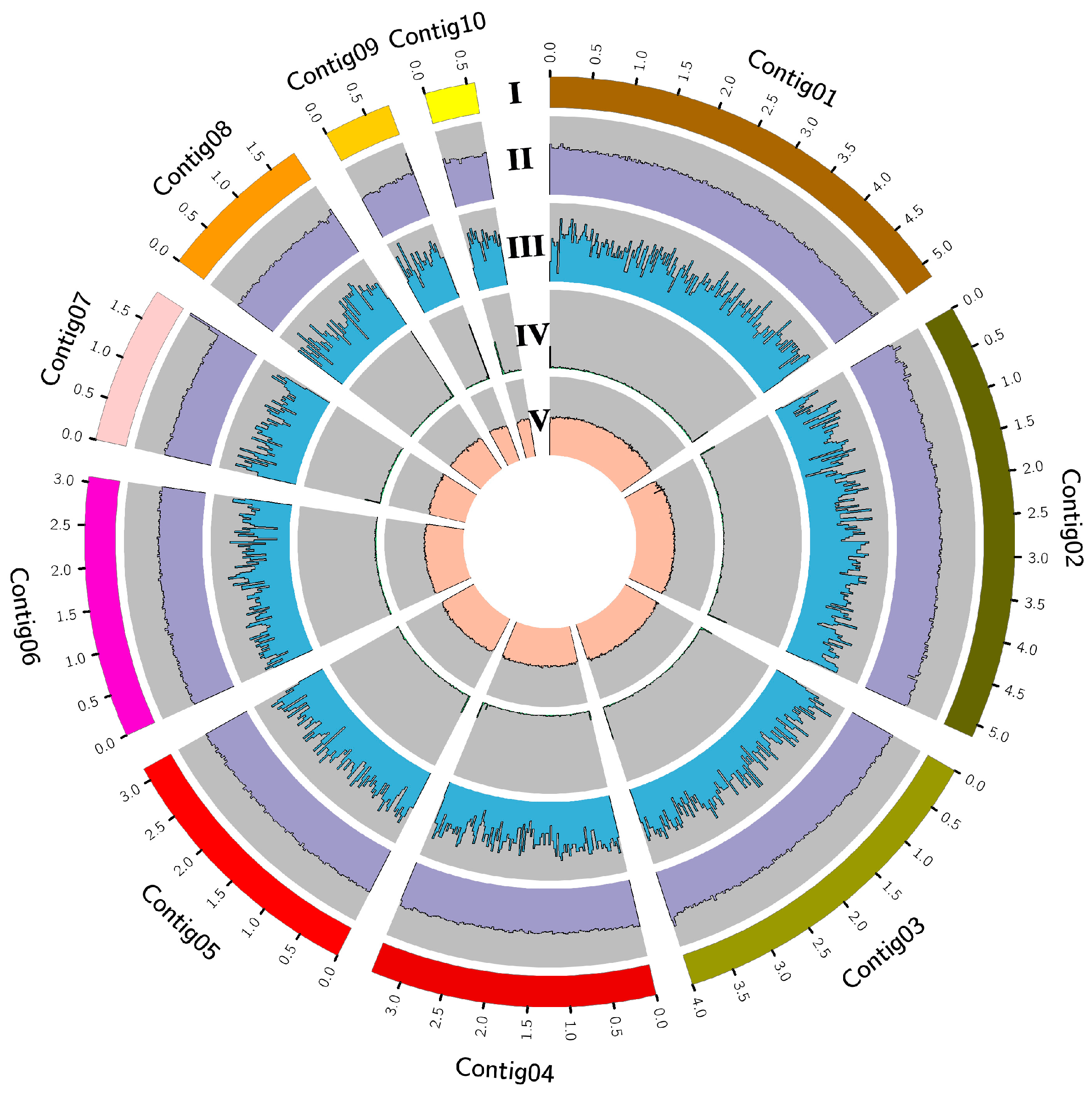
3.2. The Genome of JC-1 and the Gene Clusters for the Secondary Metabolites
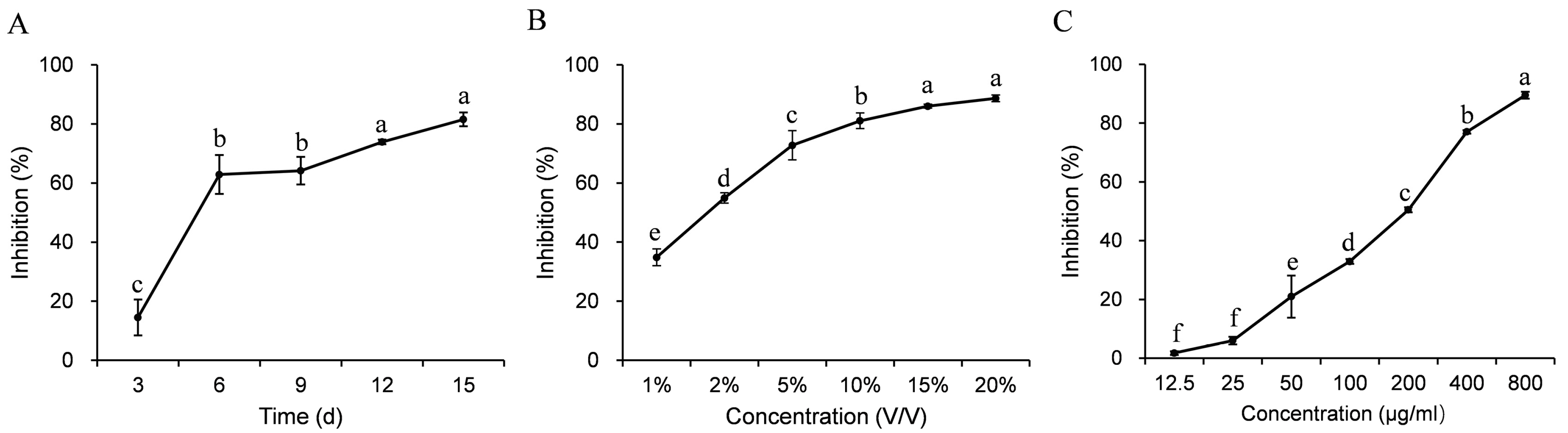
3.3. Antifungal Activity
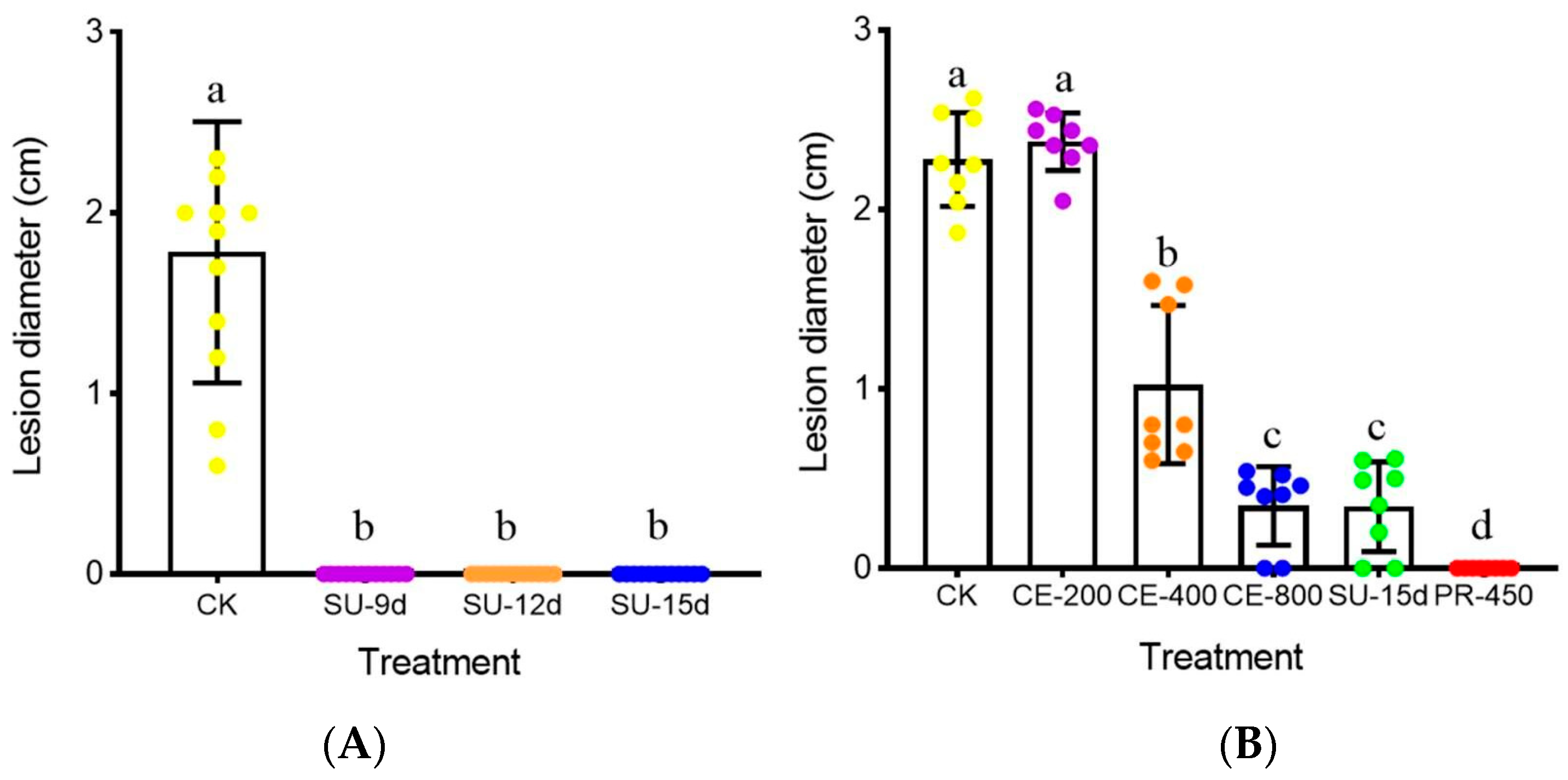
3.4. Control Efficacy of the Cultures and the Crude Extract
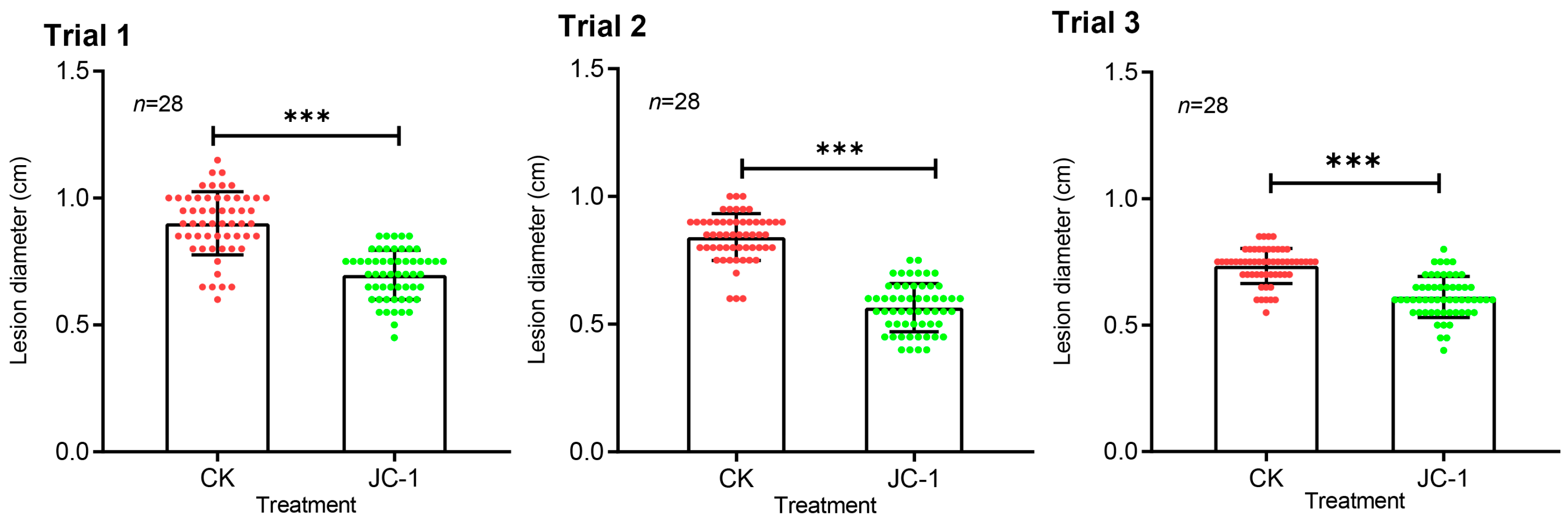
3.5. Endophytic Growth and Induced Systemic Resistance
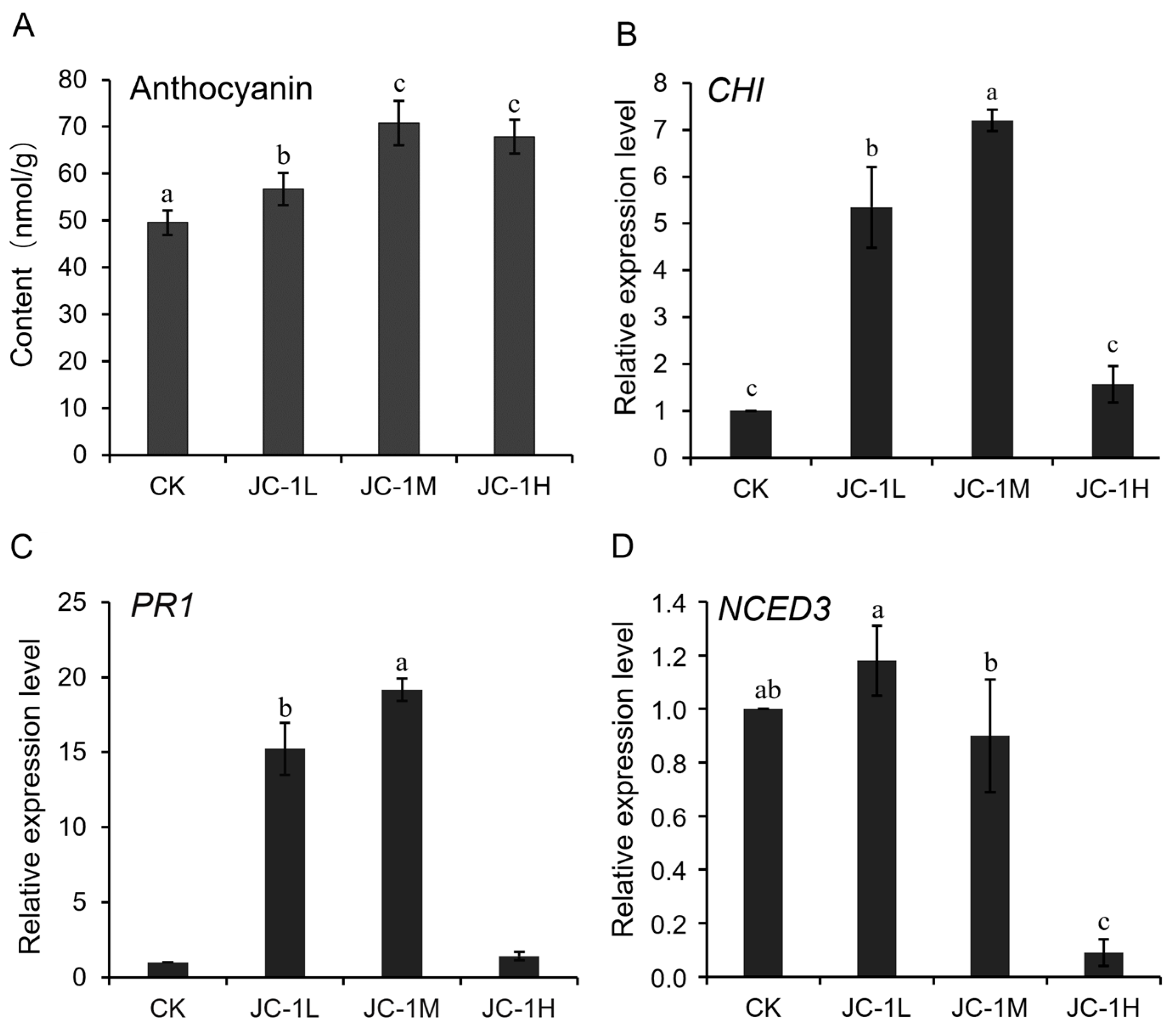
3.6. Mechanisms for the Enhanced Systemic Resistance
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zare, R.; Gams, W. A revision of Verticillium section Prostrata. IV. The genera Lecanicillium and Simplicillium gen. nov. Nova Hedwigia 2001, 73, 1–50. [Google Scholar] [CrossRef]
- Chen, W.H.; Liu, C.; Han, Y.F.; Liang, J.D.; Tian, W.Y.; Liang, Z.Q. Three novel insect-associated species of Simplicillium (Cordycipitaceae, Hypocreales) from Southwest China. MycoKeys 2019, 58, 83–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.H.; Han, Y.F.; Liang, J.D.; Liang, Z.Q. Taxonomic and phylogenetic characterizations reveal four new species of Simplicillium (Cordycipitaceae, Hypocreales, Ascomycota) from Guizhou, China. Sci. Rep. 2021, 11, 15300. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Luangsa-ard, J.J.; Wingfield, M.J.; Carnegie, A.J.; Hernández-Restrepo, M.; Lombard, L.; Roux, J.; Barreto, R.W.; Baseia, I.G.; Cano-Lira, J.F.; et al. Fungal Planet description sheets: 785–867. Persoonia 2018, 41, 238–417. [Google Scholar] [CrossRef]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Thangavel, R.; Wingfield, M.J.; Noordeloos, M.E.; Dima, B.; Brandrud, T.E.; Jansen, G.M.; et al. Fungal Planet description sheets: 1182–1283. Persoonia 2021, 46, 313–528. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.L.; Dong, R.Z.; Xing, X.Y.; Li, Y.K. A new antibiotic produced by the cyanobacterium-symbiotic fungus Simplicillium lanosoniveum. Nat. Prod. Res. 2018, 32, 1348–1352. [Google Scholar] [CrossRef]
- Liu, F.; Cai, L. Morphological and molecular characterization of a novel species of Simplicillium from China. Cryptogamie Mycologie 2012, 33, 137–144. [Google Scholar] [CrossRef]
- Nonaka, K.; Kaifuchi, S.; Omura, S.; Masuma, R. Five new Simplicillium species (Cordycipitaceae) from soils in Tokyo, Japan. Mycoscience 2013, 54, 42–53. [Google Scholar] [CrossRef]
- Wei, D.P.; Wanasinghe, D.N.; Hyde, K.D.; Mortimer, P.E.; Xu, J.C.; Xiao, Y.P.; Bhunjun, C.S.; To-anun, C. The genus Simplicillium. MycoKeys 2019, 60, 69–92. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.Z.; Liu, S.J.; Cai, L. Culturable mycobiota from Karst caves in China, with descriptions of 20 new species. Persoonia 2017, 39, 1–31. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Zhou, S.Y.; Eurwilaichitr, L.; Ingsriswang, S.; Raza, M.; Chen, Q.; Zhao, P.; Liu, F.; Cai, L. Culturable mycobiota from Karst caves in China II, with descriptions of 33 new species. Fung. Divers. 2021, 106, 29–136. [Google Scholar] [CrossRef]
- Baiswar, P.; Ngachan, S.V.; Rymbai, H.; Chandra, S. Simplicillium lanosoniveum, a hyperparasite on Aecidium elaeagni-latifoliae in India. Austral. Plant Dis. Notes 2014, 9, 144. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, N.W.; Maruthachalam, K.; Subbarao, K.V.; Brown, M.; Xiao, Y.; Robertson, C.L.; Schneider, R.W. Mycoparasitism of Phakopsora pachyrhizi, the soybean rust pathogen, by Simplicillium lanosoniveum. Biol. Control 2014, 76, 87–94. [Google Scholar] [CrossRef]
- Sun, J.Z.; Liu, X.Z.; McKenzie, E.H.C.; Jeewon, R.; Liu, J.K.; Zhang, X.L.; Zhao, Q.; Hyde, K.D. Fungicolous fungi: Terminology, diversity, distribution, evolution, and species checklist. Fung. Divers. 2019, 95, 337–430. [Google Scholar] [CrossRef]
- Wang, N.; Fan, X.; Zhang, S.; Liu, B.; He, M.; Chen, X.; Tang, C.; Kang, Z.; Wang, X. Identification of a hyperparasitic Simplicillium obclavatum strain affecting the infection dynamics of Puccinia striiformis f. sp. tritici on wheat. Front. Microbiol. 2020, 11, 1277. [Google Scholar] [CrossRef]
- Ward, N.A.; Schneider, R.W.; Aime, M.C. Colonization of soybean rust sori by Simplicillium lanosoniveum. Fung. Ecol. 2011, 4, 303–308. [Google Scholar] [CrossRef]
- Zhu, M.; Duan, X.; Cai, P.K.; Li, Y.F.; Qiu, Z.B. Deciphering the genome of Simplicillium aogashimaense to understand its mechanisms against the wheat powdery mildew fungus Blumeria graminis f. sp. tritici. Phytopathol. Res. 2022, 4, 16. [Google Scholar] [CrossRef]
- Lim, S.Y.; Lee, S.; Kong, H.G.; Lee, J.K. Entomopathogenicity of Simplicillium lanosoniveum isolated in Korea. Mycobiology 2014, 42, 317–321. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Liu, B.; Li, L.Y.; Zhu, X.F.; Wang, Y.Y.; Wang, Q.J.; Duan, Y.X.; Chen, L.J. Simplicillium chinense: A biological control agent against plant parasitic nematodes. Biocontrol Sci. Technol. 2013, 23, 980–986. [Google Scholar] [CrossRef]
- Chen, R.S.; Huang, C.C.; Li, J.C.; Tsay, J.G. First report of Simplicillium lanosoniveum causing brown spot on Salvinia auriculata and S. molesta in Taiwan. Plant Dis. 2008, 92, 1589. [Google Scholar] [CrossRef]
- Shen, X.Y.; Zheng, D.Q.; Gao, J.; Hou, C.L. Isolation and evaluation of endophytic fungi with antimicrobial ability from Phyllostachys edulis. Bangladesh J. Pharmacol. 2012, 7, 249–257. [Google Scholar] [CrossRef]
- Teasdale, S.E.; Caradus, J.R.; Johnson, L.J. Fungal endophyte diversity from tropical forage grass Brachiaria. Plant Ecol. Divers. 2018, 11, 611–624. [Google Scholar] [CrossRef]
- Liang, X.; Nong, X.H.; Huang, Z.H.; Qi, S.H. Antifungal and antiviral cyclic peptides from the deep-sea-derived fungus Simplicillium obclavatum EIODSF 020. J. Agric. Food Chem. 2017, 65, 5114–5121. [Google Scholar] [CrossRef]
- Ward, N.A.; Robertson, C.L.; Chanda, A.K.; Schneider, R.W. Effects of Simplicillium lanosoniveum on Phakopsora pachyrhizi, the soybean rust pathogen, and its use as a biological control agent. Phytopathology 2012, 102, 749–760. [Google Scholar] [CrossRef] [Green Version]
- Shin, T.S.; Yu, N.H.; Lee, J.; Choi, G.J.; Kim, J.C.; Shin, C.S. Development of a biofungicide using a mycoparasitic fungus Simplicillium lamellicola BCP and its control efficacy against gray mold diseases of tomato and ginseng. Plant Pathol. J. 2017, 33, 337–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, G.J.; Kim, J.C.; Jang, K.S.; Cho, K.Y.; Kim, H.T. Mycoparasitism of Acremonium strictum BCP on Botrytis cinerea, the gray mold pathogen. J. Microbiol. Biotechnol. 2008, 18, 167–170. [Google Scholar] [PubMed]
- Kim, J.C.; Choi, G.J.; Kim, H.J.; Kim, H.T.; Ahn, J.W.; Cho, K.Y. Verlamelin, an antifungal compound produced by a mycoparasite, Acremonium strictum. Plant Pathol. J. 2002, 18, 102–105. [Google Scholar] [CrossRef]
- Le Dang, Q.; Shin, T.S.; Park, M.S.; Choi, Y.H.; Choi, G.J.; Jang, K.S.; Kim, I.S.; Kim, J.C. Antimicrobial activities of novel mannosyl lipids isolated from the biocontrol fungus Simplicillium lamellicola BCP against phytopathogenic bacteria. J. Agric. Food Chem. 2014, 62, 3363–3370. [Google Scholar] [CrossRef]
- Ishidoh, K.; Kinoshita, H.; Nihira, T. Identification of a gene cluster responsible for the biosynthesis of cyclic lipopeptide verlamelin. Appl. Microbiol. Biotechnol. 2014, 98, 7501–7510. [Google Scholar] [CrossRef]
- Zhang, Q.H.; Zhang, J.; Yang, L.; Zhang, L.; Jiang, D.H.; Chen, W.D.; Li, G.Q. Diversity and biocontrol potential of endophytic fungi in Brassica napus. Biol. Control 2014, 72, 98–108. [Google Scholar] [CrossRef]
- Li, J.C. Fixed-Site Monitoring of Occurrence of Blackleg of Oilseed Rape and Characterization of Infection by Conidia of the Pathogen. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2019; pp. IV + 61. [Google Scholar]
- Büttner, P.; Koch, F.; Voigt, K.; Quidde, T.; Risch, S.; Blaich, R.; Bruckner, B.; Tudzynski, P. Variations in ploidy among isolates of Botrytis cinerea: Implications for genetic and molecular analyses. Curr. Genet. 1994, 25, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Huang, Y.J.; Jiang, D.H.; Fitt, B.D.L.; Li, G.Q.; Yang, L. Evaluation of oilseed rape seed yield losses caused by Leptosphaeria biglobosa in central China. Eur. J. Plant Pathol. 2018, 150, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Du, R.; Huang, Y.J.; Zhang, J.; Yang, L.; Wu, M.D.; Li, G.Q. LAMP detection and identification of the blackleg pathogen Leptosphaeria biglobosa ‘brassicae’. Plant Dis. 2021, 105, 3192–3200. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Evol. Genet. Anal. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.S.; Crous, P.W.; Bai, Q.; Fu, M.; Yang, M.M.; Wang, X.H.; Du, Y.M.; Hong, N.; Xu, W.X.; Wang, G.P. High diversity of Diaporthe species associated with pear shoot canker in China. Persoonia 2020, 45, 132–162. [Google Scholar] [CrossRef]
- Xiao, C.L.; Chen, Y.; Xie, S.Q.; Chen, K.N.; Wang, Y.; Han, Y.; Luo, F.; Xie, Z. MECAT: Fast mapping, error correction, and de novo assembly for single-molecule sequencing reads. Nat. Methods 2017, 14, 1072–1074. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Hoff, K.J.; Lomsadze, A.; Borodovsky, M.; Stanke, M. Whole-genome annotation with BRAKER. Methods Mol. Biol. 2019, 1962, 65–95. [Google Scholar]
- Borodovsky, M.; Lomsadze, A. Eukaryotic gene prediction using GeneMark.hmm-E and GeneMark-ES. Curr. Protoc. Bioinf. 2011, 35, 4.6.1–4.6.10. [Google Scholar] [CrossRef] [Green Version]
- Stanke, M.; Diekhans, M.; Baertsch, R.; Haussler, D. Using native and syntenically mapped cDNA alignments to improve de novo gene finding. Bioinformatics 2008, 24, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, R.M.; Seppey, M.; Simao, F.A.; Manni, M.; Ioannidis, P.; Klioutchnikov, G.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO applications from quality assessments to gene prediction and phylogenomics. Mol. Biol. Evol. 2018, 35, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Han, Y.C.; Li, G.Q.; Jiang, D.H.; Huang, H.C. Suppression of Sclerotinia sclerotiorum by antifungal substances produced by the mycoparasite Coniothyrium minitans. Eur. J. Plant Pathol. 2007, 119, 411–420. [Google Scholar] [CrossRef]
- Yang, R.; Han, Y.C.; Yang, L.; Zhang, J.; Jiang, D.H.; Li, G.Q. Characterization of the antifungal substances produced by Coniothyrium minitans strain Chy-1. Chin. J. Biol. Control 2014, 30, 520–527. [Google Scholar]
- Methrtens, F.; Kranz, H.; Bednarek, P.; Weisshaar, B. The Arabidopsis transcription factor MYB12 is a flavonol-specific regulator of phenylpropanoid biosynthesis. Plant Physiol. 2005, 138, 1083–1096. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.J.; Yang, L.; Wu, M.D.; Chen, W.D.; Li, G.Q.; Zhang, J. A single-nucleotide deletion in the transcription factor gene bcsmr1 causes sclerotial-melanogenesis defificiency in Botrytis cinerea. Front. Microbiol. 2017, 8, 2492. [Google Scholar] [CrossRef]
- Dawson, M.J.; Farthing, J.E.; Marshall, P.S.; Middleton, R.F.; O’Neill, M.J.; Shuttleworth, A.; Stylli, C.; Tait, R.M.; Taylorm, P.M.; Wildman, H.G.; et al. Squalestatins, novel inhibitors of squalene synthase produced by a species of Phoma I. Taxonomy, fermantation isolation physico-chemical properties and biological activity. J. Antibiot. 1992, 45, 614–622. [Google Scholar] [CrossRef] [Green Version]
- Takesako, K.; Ikai, K.; Haruna, F.; Endo, M.; Shimanaka, K.; Sonon, E.; Nakamura, T.; Kato, I. Aureobasidins, new antifungal antibiotics. taxonomy, fermentation, isolation and properties. J. Antibiot. 1991, 44, 919–924. [Google Scholar] [CrossRef]
- Bonsch, B.; Belt, V.; Bartel, C.; Duensing, N.; Koziol, M.; Lazarus, C.M.; Bailey, A.M.; Simpson, T.J.; Cox, R.J. Identification of genes encoding squalestatin S1 biosynthesis and in vitro production of new squalestatin analogues. Chem. Comm. 2016, 52, 6777–6780. [Google Scholar] [CrossRef] [Green Version]
- Slightom, J.L.; Metzger, B.P.; Luu, H.T.; Elhammer, A.P. Cloning and molecular characterization of the gene encoding the aureobasidin A biosynthesis complex in Aureobasidium pullulans BP-1938. Gene 2009, 431, 67–79. [Google Scholar] [CrossRef]
- Li, W.T. Characterization of the Taxonomic Identity, Biocontrol Potential and Mechanisms of Simplicillium sp. JC-1. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2020; pp. IV + 103. [Google Scholar]
- Peng, G.; Liu, X.J.; McLaren, D.L.; McGregor, L.; Yu, F.Q. Seed treatment with the fungicide fluopyram limits cotyledon infection by Leptosphaeria maculans and reduces blackleg of canola. Can. J. Plant Pathol. 2020, 42, 480–492. [Google Scholar] [CrossRef]
- Lowe, R.G.T.; Cassin, A.; Grandaubert, J.; Clark, B.L.; Van de Wouw, A.P.; Rouxel, T.; Howlett, B.J. Genomes and transcriptomes of partners in plant-fungal-interactions between canola (Brassica napus) and two Leptosphaeria species. PLoS ONE 2014, 9, e103098. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-alpha sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [PubMed]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- White, T.J. PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Ed.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Collection No. | Host or Niche | Country or Region | GenBank Acc. No. | |||
|---|---|---|---|---|---|---|---|
| ITS | SSU | LSU | TEF1a | ||||
| Akanthomyces lecanii | CBS 101,247 = IMI 304807 | Coccus viridis | West Indies | AJ292382 | AF339604 | AF339555 | DQ522359 |
| S. aogashimaense | JCM 18167 | Soil | Japan | AB604002 | LC496889 | LC496874 | LC496904 |
| S. aogashimaense | JCM 18168 | Soil | Japan | AB604004 | LC496890 | LC496875 | LC496905 |
| S. calcicola | LC5586 = CGMCC3.17943 | Calcaire | China | KU746706 | KY883301 | KU746752 | KX855252 |
| S. chinense | LC 1342 | Freshwater | China | JQ410323 | - | JQ410321 | - |
| S. chinense | LC 1345 | Freshwater | China | JQ410324 | - | JQ410322 | - |
| S. coffeanum | CDA 734 | Coffea arabica | Brazil | MF066034 | - | MF066032 | - |
| S. cylindrosporum | JCM 18169 | Soil | Japan | AB603989 | LC496891 | LC496876 | LC496906 |
| S. cylindrosporum | JCM 18170 | Soil | Japan | AB603994 | LC496892 | LC496877 | LC496907 |
| S. filiforme | URM 7918 | Citrullus lanatus | Brazil | MH979338 | - | MH979399 | - |
| S. formicae | MFLUCC 18-1379 | Formicidae | Thailand | NR_168789 | NG_070121 | NG_068624 | MK926451 |
| S. lamellicola | JC-1 | Brassica napus | China | MT807906 | MT807908 | MT807907 | MT826785 |
| S. lamellicola | CBS 116.25 T | Agaricus bisporus | UK | MH854806 | AF339601 | AF339552 | DQ522356 |
| S. lamellicola | CBS 454.70 | - | USA | MH859793 | - | MH871559 | - |
| S. lanosoniveum | CBS 704.86 | Hemileia vastatrix | Venezuela | AJ292396 | AF339602 | AF339553 | DQ522358 |
| S. lanosoniveum var. tianjinienss | CGMCC 4460 | Blue-green alga | China | HM989951 | - | - | - |
| S. minatense | JCM 18176 | Soil | Japan | AB603992 | LC496893 | LC496878 | LC496908 |
| S. minatense | JCM 18178 | Soil | Japan | AB603993 | LC496894 | LC496879 | LC496909 |
| S. obclavatum | CBS 311.74 | Air | India | AJ292394 | AF339567 | AF339517 | EF468798 |
| S. obclavatum | JCM 18179 | Soil | Japan | AB604000 | - | - | - |
| S. spumae | JCM 39050 | Foam | Japan | LC496869 | LC496898 | LC496883 | LC496913 |
| S. spumae | JCM 39051 | Foam | Japan | LC496870 | LC496899 | LC496884 | LC496914 |
| S. spumae | JCM 39054 | Foam | Japan | LC496871 | LC496902 | LC496887 | LC496917 |
| S. subtropicum | JCM 18180 | Soil | Japan | AB603990 | LC496895 | LC496880 | LC496910 |
| S. subtropicum | JCM 18181 | Soil | Japan | AB603995 | LC496896 | LC496881 | LC496911 |
| S. sympodiophorum | JCM 18184 | Soil | Japan | AB604003 | LC496897 | LC496882 | LC496912 |
| Variable | Statistics |
|---|---|
| Genome size | 29.14 Mb |
| Genome coverage | 427.93× |
| Number of scaffolds | 22 |
| Average contig length | 1.32 Mb |
| Length of the largest contig | 5.180 Mb |
| N50 | 3.35 Mb |
| GC content | 48.37% |
| Protein coding genes | 10,488 |
| Gene clusters for secondary metabolites (SM) | 38 |
| Known SM gene clusters | 8 |
| Treatment 1 | No. Tissues Colonized by S. lamellicola/Total Tissues (Percentage) | ||
|---|---|---|---|
| Root Segments | Stem Segments | Leaf Pieces | |
| Water alone | 0/21 (0) | 0/21 (0) | 0/21 (0) |
| Conidial suspension of JC-1 | 0/21 (0) | 5/21 (23.8%) | 3/21 (14.3%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Luo, T.; Li, J.; Zhang, J.; Wu, M.; Yang, L.; Li, G. Antifungal Activity and Biocontrol Potential of Simplicillium lamellicola JC-1 against Multiple Fungal Pathogens of Oilseed Rape. J. Fungi 2023, 9, 57. https://doi.org/10.3390/jof9010057
Li W, Luo T, Li J, Zhang J, Wu M, Yang L, Li G. Antifungal Activity and Biocontrol Potential of Simplicillium lamellicola JC-1 against Multiple Fungal Pathogens of Oilseed Rape. Journal of Fungi. 2023; 9(1):57. https://doi.org/10.3390/jof9010057
Chicago/Turabian StyleLi, Wenting, Tao Luo, Juncheng Li, Jing Zhang, Mingde Wu, Long Yang, and Guoqing Li. 2023. "Antifungal Activity and Biocontrol Potential of Simplicillium lamellicola JC-1 against Multiple Fungal Pathogens of Oilseed Rape" Journal of Fungi 9, no. 1: 57. https://doi.org/10.3390/jof9010057