
Lignin and Its Pathway-Associated Phytoalexins Modulate Plant Defense against Fungi
 , ,
, ,  ,
,
Abstract
:1. Introduction
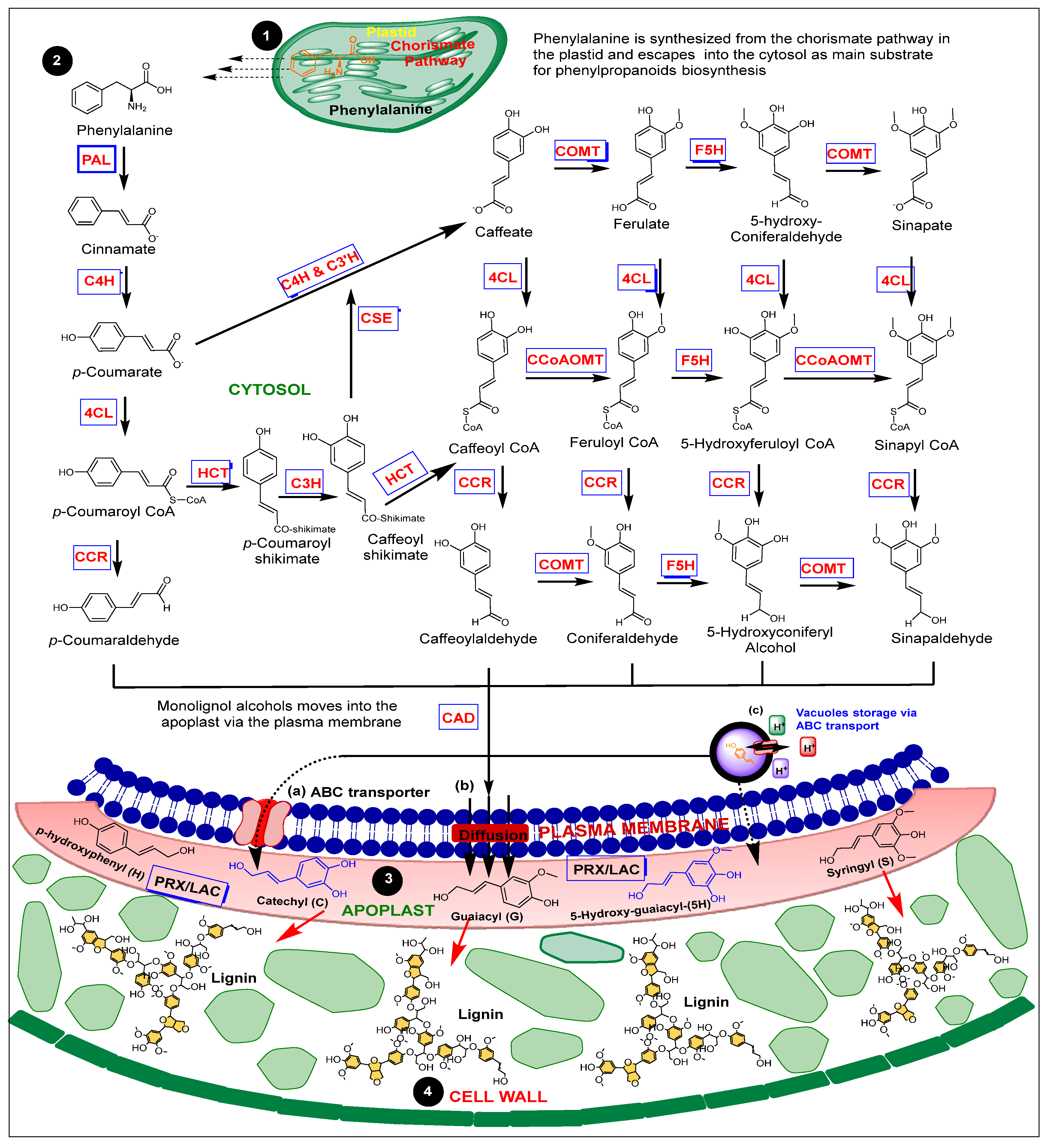
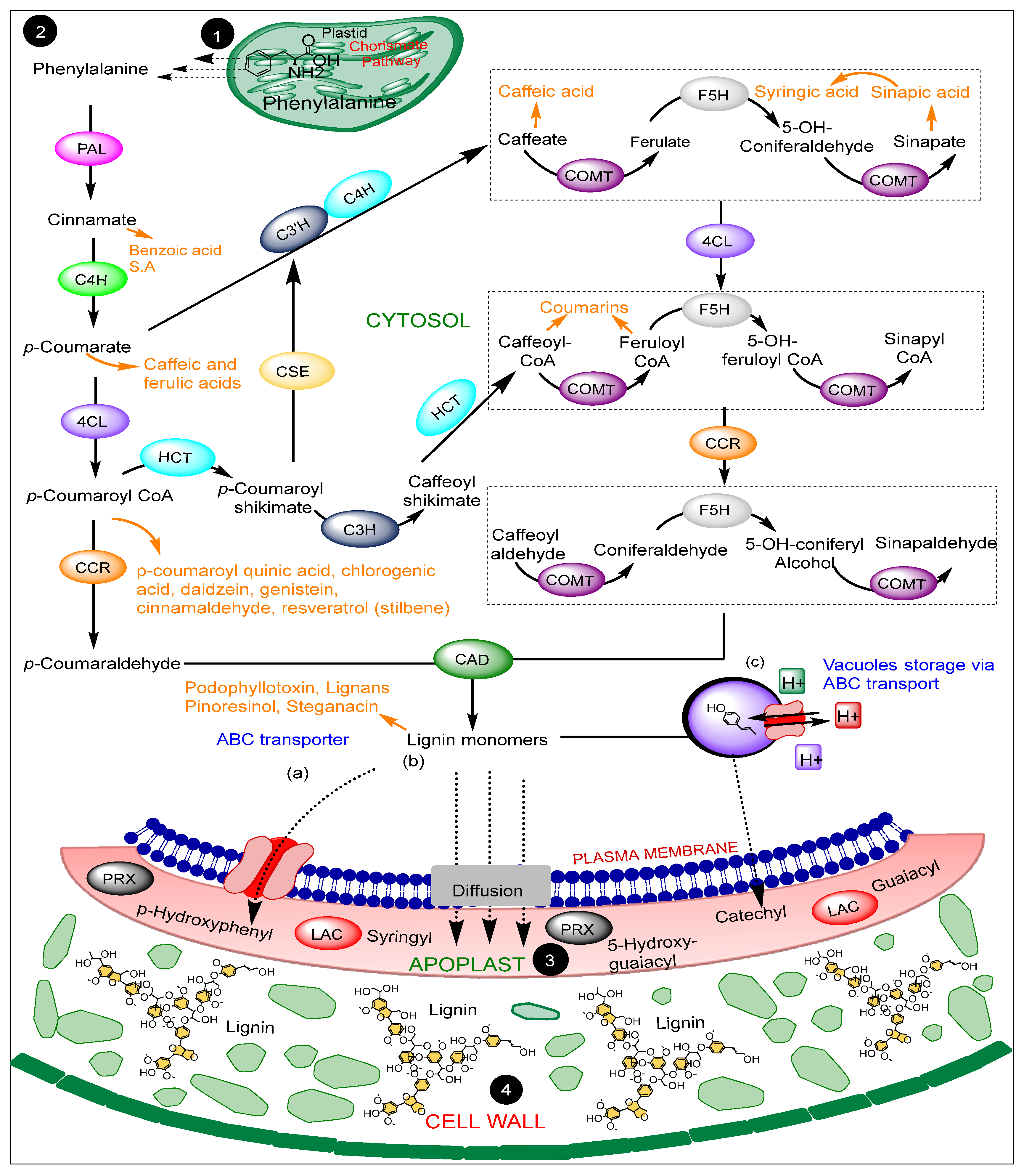
2. A Brief Overview of Monolignols Biosynthesis and Lignification
2.1. PHENYLALANINE AMMONIA-LYASE (PAL)
2.2. CINNAMATE 4-HYDROXYLASE (C4H)
2.3. 4-COUMARATE-COA –COENZYME A LIGASE (4CL)

{kind=link}
{kind=link}
{kind=link}
| No. | Gene/Protein | Plant | Research Strategy | Results Obtained | Metabolites | References |
|---|---|---|---|---|---|---|
| 1 | MdMRLK2 | Malus mellana | Overexpression MdMRLK2 cucurbits | Suppressed PAL, β-1,3- glucanase, chitinase | Inhibited polyphenol synthesis | [45] |
| 2 | AtERF114 | A. thaliana | RNAseq, overexpression, knockout | ERF114 activates PAL1 to mediate P. syringae pv tomato (Pst) defense | Lignin and SA | [46] |
| 3 | PAL1, 4CL5, MYB308 | Prunus persica | Overexpression MYB308 | PAL1 and 4CL5 enhanced expression-induced resistance against R. stolonifer | Chlorogenic, gallic acid, and rutin | [47] |
| 4 | POX, PAL | Zea mays | Inoculated maize genotypes (P1630H, AG3700, SCS156 Colorado and 30K75Y) with Bipolaris maydis | POX, PAL transcript abundance conferred resistance to B. maydis in AG3700 | phenolic and flavonoids | [48] |
| 5 | PAL, POD | Nicotiana tobaccum | Thiamine (vitamin B1, VB1) treatment | Increased PAL, POD, H2O2 accumulation, and catalase and peroxidase activities conferred resistance against Phytophthora nicotianae | - | [49] |
| 6 | WRKY1 | Ocimum sanctum, A. thaliana | Overexpression and VIGS OF WRKY1 | WRKY1 regulates PAL and C4H resistance to P. syringae pv. tomato Pst DC3000 | - | [50] |
| 7 | PAL | Phoenix dactylifera | Alginate extract from Bifurcaria bifurcata was tested agaisnt F. oxysporum | Alginate treatment triggered PAL expression against F. oxysporum f. sp. Albedinis | - | [51] |
| 8 | C4H , CAD, POD | Prunus persica | RNAseq, transient overexpression of PpMYB306 | P. guilliermondii inhibits PpMYB306 repressed lignin genes in peach after R. stolonifer infection. | Inhibited lignin content | [52] |
| 9 | C4H, COMT, BAK1, WRKY5 | Olea europaea | Analysis of defense mechanism of tolerant and susceptible olive cultivars to V. dahliae | V. dahlia-tolerant cultivar significantly accumulated root lignin after V. dahlia inoculation | Lignin | [53] |
| 10 | PALs, Cl4Cls, CYP73A, CCR ClHCTs | Citrullus lanatus | RNA-Seq of resistant ZXG1755 and susceptible ZXG1996 lines inoculated with powdery mildew during the early seedling stage | Hormonal, lignin and peroxidase transcripts were significantly expressed | Lignin and phytohormone biosynthesis | [54] |
| 11 | ScAPD1- like | Syntrichia caninervis | Overexpression of ScAPD1-like in Arabidopsis and S. caninervis | Defense against V. dahliae, decreased ROS synthesis, improved ROS scavenging activity, enhanced lignin (PAL, C4H) transcripts | High lignin accumulation | [55] |
| 12 | Hrip1 | Oryza sativa | RNAseq and metabolic analysis of Hrip1-treated rice leaves | Hrip1 mediates defense against rice blast fungi by activating PAL, C4H, 4CL, HCT, C3H, COMT, CAD, PRX, diterpene synthases (CPS2, -4, KSL4, 5, -6, -7, 10, cytochromes (CYP71Z, CYP7M, momilactone synthases), benzoxazinoids biosynthetic genes (BX1-BX7) | Lignin, diterpenoids | [56] |
| 13 | WRKY , PAL, CHI | Vigna angularies | Transcriptome and histological analysis of Vigna angularies against Uromyces vignae | PRRs recognize U. vignae invasion and activities PAL, WRKY, CHI defense | - | [57] |
| 14 | CAD35, CAD45, CAD43 | G. hirsutum | VIGS and overexpression of GhCAD35, GhCAD45, or GhCAD43 | VIGS of CAD genes inhibited S-lignin production, ultimately affecting the syringyl/guaiacyl (S/G) ratio, while OE-lines enhanced V. dahliae defense | Lignin, SA | [58] |
| 15 |
PAL
, 4CL,
COMT
, CAD POX | Panax notoginseng | Transcriptomic and proteomic technologies | Alternaria panax inoculation activated PAL, 4CL, COMT, CAD, POX expression | Lignin | [59] |
| 16 | PAL | Cajanus cajan | Metabolic analysis | Fusarium udum induced the expression of lignin-related transcripts and enzyme activities for lignin and phenolic acids accumulation | Phenolics, lignin | [60] |
| 17 | COMT, PRX, CAD, HCT | Malus domestica | Comparative RNA-seq analysis | Malus domestica inoculated with Fpmd MR5 induced the expression of several lignin genes, antimicrobial and antioxidants genes | - | [61] |
| 18 | COMT1 | Triticum aestivum | Transcription profiling of genes involved in Triticum aestivum- Puccinia striiformis interaction | COMT1 was highly expressed in response to Puccinia striiformis inoculation | - | [62] |
| 19 | GhODO1 Gh4CL1, GhCAD3 | G. hirsutum | GhODO1-GFP transient expression in onion, qPCR, lignin quantification | GhODO1 binds to Gh4CL1 and GhCAD3 promoters to activate lignin-enhanced resistance to V. dahliae | Lignin, JA | [63] |
| 20 | LCC24 , ROMT, LCC24, | Elaeis guineensis | Analysis of oil palm defense against Ganoderma boninense inoculation, qPCR, and metabolic analysis | oil palm cultivar, C08 exhibited high resistance by activating Ganoderma boninense | SA, lignin | [64] |
| 21 | Xylogen-like arabinogalactan protein1 and -2 | Capsicum annuum | Genome-wide studies, phylogenetics, and VIGS analysis | Enhanced expression of lignin genes and lignin accumulation in pepper stem. | Lignin | [65] |
| 22 | Ammonia-lysases (ALs) | B. distachylon | Proteomics, RNAi knockdown, metabolic analysis | Ammonia-lysases performed a central role in carbon allocation for lignin accumulation and shikimate ester does not contribute to lignin synthesis in B. distachylon | Lignin | [66] |
2.4. HYDROXYCINNAMOYL TRANSFERASE (HCT)
2.5. CAFFEOYL SHIKIMATE ESTERASE (CSE)
2.6. CAFFEOYL-COENZYME A 3-O-METHYLTRANSFERASE (CCoAOMTs) and CAFFEIC ACID 3-O-METHYLTRANSFERASE (COMTs)
2.7. FERULATE 5-HYDROXYLASE (F5H)
2.8. CINNAMOYL COA REDUCTASE (CCR)
2.9. CINNAMYL ALCOHOL DEHYDROGENASE (CAD)
2.10. PEROXIDASES and LACCASES (PRX and LACs)
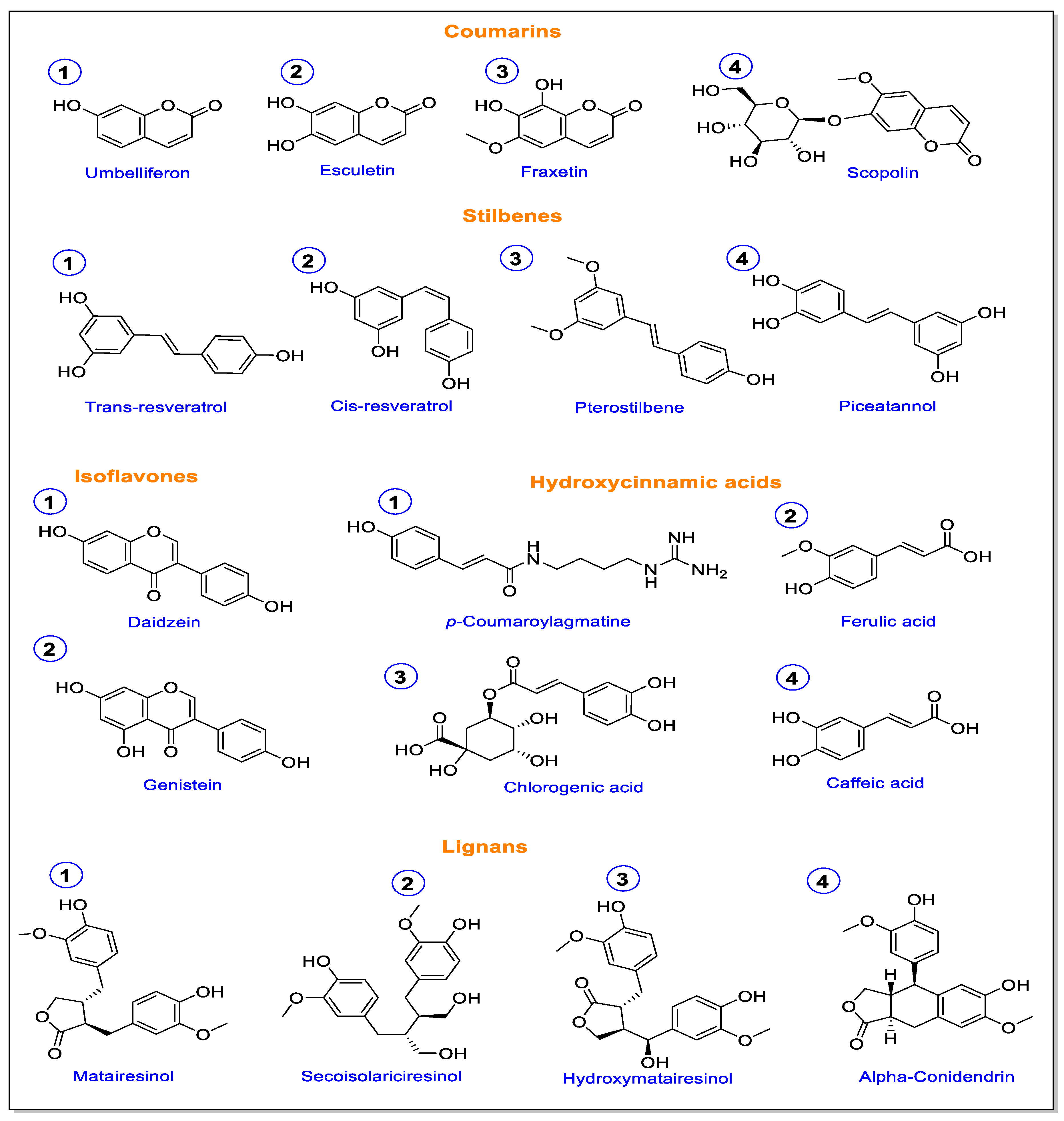
3. Phytoalexins Associated with the Lignin Pathway Enzymes
4. Missing Links in the Lignin Research, Prospects, and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Citores, L.; Iglesias, R.; Gay, C.; Ferreras, J.M. Antifungal activity of the ribosome-inactivating protein BE27 from sugar beet (Beta vulgaris L.) against the green mould Penicillium digitatum. Mol. Plant Pathol. 2016, 17, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Landi, N.; Ragucci, S.; Citores, L.; Clemente, A.; Hussain, H.Z.F.; Iglesias, R.; Ferreras, J.M.; Di Maro, A. Isolation, Characterization and Biological Action of Type-1 Ribosome-Inactivating Proteins from Tissues of Salsola soda L. Toxins 2022, 14, 566. [Google Scholar] [CrossRef] [PubMed]
- Molina, A.; Miedes, E.; Bacete, L.; Rodríguez, T.; Mélida, H.; Denancé, N.; Sánchez-Vallet, A.; Rivière, M.-P.; López, G.; Freydier, A.; et al. Arabidopsis cell wall composition determines disease resistance specificity and fitness. Proc. Natl. Acad. Sci. USA 2021, 118, e2010243118. [Google Scholar] [CrossRef] [PubMed]
- Silva-Sanzana, C.; Estevez, J.M.; Blanco-Herrera, F. Influence of cell wall polymers and their modifying enzymes during plant–aphid interactions. J. Exp. Bot. 2019, 71, 3854–3864. [Google Scholar] [CrossRef] [PubMed]
- Vaahtera, L.; Schulz, J.; Hamann, T. Cell wall integrity maintenance during plant development and interaction with the environment. Nat. Plants 2019, 5, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; He, M.; Hou, Q.; Zou, L.; Yang, Y.; Wei, Y.; Chen, X. Cell wall associated immunity in plants. Stress Biol. 2021, 1, 1–15. [Google Scholar] [CrossRef]
- Pruitt, R.N.; Gust, A.A.; Nürnberger, T. Plant immunity unified. Nat. Plants 2021, 7, 382–383. [Google Scholar] [CrossRef]
- Dong, N.-Q.L.; Lin, H.-X. Contribution of phenylpropanoid metabolism to plant development and plant-environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef]
- Lavhale, S.G.; Kalunke, R.M.; Giri, A.P. Structural, functional and evolutionary diversity of 4-coumarate-CoA ligase in plants. Planta 2018, 248, 1063–1078. [Google Scholar] [CrossRef]
- Dos Santos, A.C.; Ximenes, E.; Kim, Y.; Ladisch, M.R. Lignin-Enzyme Interactions in the Hydrolysis of Lignocellulosic Biomass. Trends Biotechnol. 2019, 37, 518–531. [Google Scholar] [CrossRef]
- Kiselev, K.V.; Dubrovina, A.S. Overexpression of stilbene synthase genes to modulate the properties of plants and plant cell cultures. Biotechnol. Appl. Biochem. 2021, 68, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.P.; Matthews, M.L.; Naik, P.P.; Williams, C.M.; Ducoste, J.J.; Sederoff, R.R.; Chiang, V.L. Flux modeling for monolignol biosynthesis. Curr. Opin. Biotechnol. 2019, 56, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Luan, Y.; Shi, W.; Zhang, X.; Meng, J.; Tao, J. A Paeonia ostii caffeoyl-CoA O-methyltransferase confers drought stress tolerance by promoting lignin synthesis and ROS scavenging. Plant Sci. 2021, 303, 110765. [Google Scholar] [CrossRef]
- Lin, S.J.; Yang, Y.Z.; Teng, R.M.; Liu, H.; Li, H.; Zhuang, J. Identification and expression analysis of caffeoyl-coenzyme A O-methyltransferase family genes related to lignin biosynthesis in tea plant (Camellia sinensis). Protoplasma 2021, 258, 115–127. [Google Scholar] [CrossRef]
- Barros, J.; Temple, S.; Dixon, R.A. Development and commercialization of reduced lignin alfalfa. Curr. Opin. Biotechnol. 2019, 56, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Barros, J.; Dixon, R.A. Plant phenylalanine/tyrosine ammonia-lyases. Trends Plant Sci. 2020, 25, 66–79. [Google Scholar] [CrossRef]
- Vanholme, R.; De Meester, B.; Ralph, J.; Boerjan, W. Lignin biosynthesis and its integration into metabolism. Curr. Opin. Biotechnol. 2019, 56, 230–239. [Google Scholar] [CrossRef]
- Quan, M.; Du, Q.; Xiao, L.; Lu, W.; Wang, L.; Xie, J.; Song, Y.; Xu, B.; Zhang, D. Genetic architecture underlying the lignin biosynthesis pathway involves noncoding RNAs and transcription factors for growth and wood properties in Populus. Plant Biotechnol. J. 2019, 17, 302–315. [Google Scholar] [CrossRef] [Green Version]
- Zhong, R.; Cui, D.; Ye, Z.-H. Secondary cell wall biosynthesis. New Phytol. 2019, 221, 1703–1723. [Google Scholar] [CrossRef] [Green Version]
- Chao, N.; Qi, Q.; Li, S.; Ruan, B.; Jiang, X.; Gai, Y. Characterization and functional analysis of the Hydroxycinnamoyl-CoA: Shikimate hydroxycinnamoyl transferase (HCT) gene family in poplar. PeerJ 2021, 9, e10741. [Google Scholar] [CrossRef] [PubMed]
- Barros, J.; Escamilla-Trevino, L.; Song, L.; Rao, X.; Serrani-Yarce, J.C.; Palacios, M.D.; Engle, N.; Choudhury, F.K.; Tschaplinski, T.J.; Venables, B.J.; et al. 4-Coumarate 3-hydroxylase in the lignin biosynthesis pathway is a cytosolic ascorbate peroxidase. Nat. Commun. 2019, 10, 1994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, E.M.; Stone, M.L.; Katahira, R.; Reed, M.; Muchero, W.; Ramirez, K.J.; Beckham, G.T.; Román-Leshkov, Y. Differences in S/G ratio in natural poplar variants do not predict catalytic depolymerization monomer yields. Nat. Commun. 2019, 10, 2033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Wu, F.; Zhang, J. NAC and MYB Families and Lignin Biosynthesis-Related Members Identification and Expression Analysis in Melilotus albus. Plants 2021, 10, 303. [Google Scholar] [CrossRef]
- Yadav, V.; Wang, Z.; Wei, C.; Amo, A.; Ahmed, B.; Yang, X.; Zhang, X. Phenylpropanoid Pathway Engineering: An Emerging Approach towards Plant Defense. Pathogens 2020, 9, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Lam, P.Y.; Lee, M.-H.; Jeon, H.S.; Tobimatsu, Y.; Park, O.K. The Arabidopsis R2R3 MYB Transcription Factor MYB15 Is a Key Regulator of Lignin Biosynthesis in Effector-Triggered Immunity. Front. Plant Sci. 2020, 11, 583153. [Google Scholar] [CrossRef]
- von Tiedemann, A.; Koopmann, B.; Hoech, K. Lignin composition and timing of cell wall lignification are involved in Brassica napus resistance to Sclerotinia sclerotiorum stem rot. Phytopathology® 2021, 111, 1438–1448. [Google Scholar] [CrossRef]
- Saur, I.M.L.; Hückelhoven, R. Recognition and defence of plant-infecting fungal pathogens. J. Plant Physiol. 2021, 256, 153324. [Google Scholar] [CrossRef]
- Heldt, H.-W.P.; Piechulla, B. Chapter 18—Phenylpropanoids Comprise a Multitude of Plant-Specialized Metabolites and Cell Wall Components. In Plant Biochemistry, 5th ed.; Heldt, H.-W., Piechulla, B., Eds.; Academic Press: Cambridge, MA, USA, 2021. [Google Scholar] [CrossRef]
- Trivedi, V.D.; Chappell, T.C.; Krishna, N.B.; Anuj, S.; Sigamani, G.G.; Mohan, K.; Ramesh, A.; Pravin, K.R.; Nair, N.U. In-depth sequence-function characterization reveals multiple paths to enhance phenylalanine ammonia-lyase (PAL) activity. ACS Catal. 2022, 12, 2381–2396. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, X.; Zhang, F.; Dong, L.; Wu, J.; Cheng, Q.; Qi, D.; Yan, X.; Jiang, L.; Fan, S.; et al. Phenylalanine ammonia-lyase2.1 contributes to the soybean response towards Phytophthora sojae infection. Sci. Rep. 2017, 7, 7242. [Google Scholar] [CrossRef]
- Wang, R.; Wang, G.-L.; Ning, Y. PALs: Emerging key players in broad-spectrum disease resistance. Trends Plant Sci. 2019, 24, 785–787. [Google Scholar] [CrossRef]
- Pant Shankar, R.; Irigoyen, S.; Liu, J.; Bedre, R.; Christensen Shawn, A.; Schmelz Eric, A.; Sedbrook John, C.; Scholthof Karen-Beth, G.; Mandadi Kranthi, K.; Ausubel Frederick, M. Brachypodium Phenylalanine Ammonia Lyase (PAL) Promotes Antiviral Defenses against Panicum mosaic virus and Its Satellites. mBio 2021, 12, e03518–e03520. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yu, T.; Wu, T.; Wang, R.; Wang, H.; Du, H.; Xu, X.; Xie, D.; Xu, X. The dynamic transcriptome of pepper (Capsicum annuum) whole roots reveals an important role for the phenylpropanoid biosynthesis pathway in root resistance to Phytophthora capsici. Gene 2020, 728, 144288. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Si, J.; Cui, X.; Peng, H.; Chen, X.; Xing, H.; Dou, D. The soybean cinnamate 4-hydroxylase gene GmC4H1 contributes positively to plant defense via increasing lignin content. Plant Growth Regul. 2019, 88, 139–149. [Google Scholar] [CrossRef]
- Li, G.; Liu, X.; Zhang, Y.; Muhammad, A.; Han, W.; Li, D.; Cheng, X.; Cai, Y. Cloning and functional characterization of two cinnamate 4-hydroxylase genes from Pyrus bretschneideri. Plant Physiol. Biochem. 2020, 156, 135–145. [Google Scholar] [CrossRef]
- Jannoey, P.; Channei, D.; Kotcharerk, J.; Pongprasert, W.; Nomura, M. Expression Analysis of Genes Related to Rice Resistance Against Brown Planthopper, Nilaparvata lugens. Rice Sci. 2017, 24, 163–172. [Google Scholar] [CrossRef]
- Wang, R.-S. Perturbation. In Encyclopedia of Systems Biology; Dubitzky, W., Wolkenhauer, O., Cho, K.-H., Yokota, H., Eds.; Springer: New York, NY, USA, 2013. [Google Scholar] [CrossRef]
- Desmedt, W.; Jonckheere, W.; Nguyen, V.H.; Ameye, M.; De Zutter, N.; De Kock, K.; Debode, J.; Van Leeuwen, T.; Audenaert, K.; Vanholme, B.; et al. The phenylpropanoid pathway inhibitor piperonylic acid induces broad-spectrum pest and disease resistance in plants. Plant Cell Environ. 2021, 44, 3122–3139. [Google Scholar] [CrossRef]
- Chen, X.; Su, W.; Zhang, H.; Zhan, Y.; Zeng, F. Fraxinus mandshurica 4-coumarate-CoA ligase 2 enhances drought and osmotic stress tolerance of tobacco by increasing coniferyl alcohol content. Plant Physiol. Biochem. 2020, 155, 697–708. [Google Scholar] [CrossRef]
- Li, S.-S.; Chang, Y.; Li, B.; Shao, S.-L. Functional analysis of 4-coumarate: CoA ligase from Dryopteris fragrans in transgenic tobacco enhances lignin and flavonoids. Genet. Mol. Biol. 2020, 43, e20180355. [Google Scholar] [CrossRef]
- Awasthi, P.; Mahajan, V.; Jamwal, V.L.; Chouhan, R.; Kapoor, N.; Bedi, Y.S.; Gandhi, S.G. Characterization of the gene encoding 4-coumarate:CoA ligase in Coleus forskohlii. J. Plant Biochem. Biotechnol. 2019, 28, 203–210. [Google Scholar] [CrossRef]
- Ji, N.; Wang, J.; Li, Y.; Li, M.; Jin, P.; Zheng, Y. Involvement of PpWRKY70 in the methyl jasmonate primed disease resistance against Rhizopus stolonifer of peaches via activating phenylpropanoid pathway. Postharvest Biol. Technol. 2021, 174, 111466. [Google Scholar] [CrossRef]
- Hwang, H.-S.; Han, J.Y.; Choi, Y.E. Enhanced accumulation of pinosylvin stilbenes and related gene expression in Pinus strobus after infection of pine wood nematode. Tree Physiol. 2021, 41, 1972–1987. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Kou, X.; Wu, C.; Fan, G.; Li, T. Methyl jasmonate induces the resistance of postharvest blueberry to gray mold caused by Botrytis cinerea. J. Sci. Food Agric. 2020, 100, 4272–4281. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Zhan, M.; Li, C.; Pei, T.; Wang, Q.; Li, P.; Ma, F.; Liu, C. The apple FERONIA receptor-like kinase MdMRLK2 negatively regulates Valsa canker resistance by suppressing defence responses and hypersensitive reaction. Mol. Plant Pathol. 2022, 23, 1170–1186. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, Y.; Ren, J.; Jia, F.; Zeng, H.; Li, G.; Yang, X. Ethylene-responsive factor ERF114 mediates fungal pathogen effector PevD1-induced disease resistance in Arabidopsis thaliana. Mol. Plant Pathol. 2022, 23, 819–831. [Google Scholar] [CrossRef]
- Li, Y.; Ji, N.; Zuo, X.; Hou, Y.; Zhang, J.; Zou, Y.; Jin, P.; Zheng, Y. PpMYB308 is involved in Pichia guilliermondii-induced disease resistance against Rhizopus rot by activating the phenylpropanoid pathway in peach fruit. Postharvest Biol. Technol. 2023, 195, 112115. [Google Scholar] [CrossRef]
- Schauffler, G.P.; dos Anjos Verzutti Fonseca, J.; Di Piero, R.M. Defense mechanisms involved in the resistance of maize cultivars to Bipolaris maydis. Eur. J. Plant Pathol. 2022, 163, 269–277. [Google Scholar] [CrossRef]
- Suohui, T.; Yanping, C.; Shuhui, Z.; Zhihua, L.; Honggang, J.; Jun, L.; Tao, L. Thiamine induces resistance in tobacco against black shank. Australas. Plant Pathol. 2022, 51, 231–243. [Google Scholar] [CrossRef]
- Joshi, A.; Jeena, G.S.; Shikha; Kumar, R.S.; Pandey, A.; Shukla, R.K. Ocimum sanctum, OscWRKY1, regulates phenylpropanoid pathway genes and promotes resistance to pathogen infection in Arabidopsis. Plant Mol. Biol. 2022, 110, 235–251. [Google Scholar] [CrossRef]
- Bouissil, S.; Guérin, C.; Roche, J.; Dubessay, P.; El Alaoui-Talibi, Z.; Pierre, G.; Michaud, P.; Mouzeyar, S.; Delattre, C.; El Modafar, C. Induction of Defense Gene Expression and the Resistance of Date Palm to Fusarium oxysporum f. sp. Albedinis in Response to Alginate Extracted from Bifurcaria bifurcata. Mar. Drugs 2022, 20, 88. [Google Scholar] [CrossRef]
- Li, Y.; Ji, N.; Zuo, X.; Zhang, J.; Zou, Y.; Ru, X.; Wang, K.; Jin, P.; Zheng, Y. Involvement of PpMYB306 in Pichia guilliermondii-induced peach fruit resistance against Rhizopus stolonifer. Biol. Control 2023, 177, 105130. [Google Scholar] [CrossRef]
- Cardoni, M.; Gómez-Lama Cabanás, C.; Valverde-Corredor, A.; Villar, R.; Mercado-Blanco, J. Unveiling Differences in Root Defense Mechanisms Between Tolerant and Susceptible Olive Cultivars to Verticillium dahliae. Front. Plant Sci. 2022, 13, 1052. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.; Wang, Z.; Guo, Y.; Zhang, X. Comparative transcriptome profiling reveals the role of phytohormones and phenylpropanoid pathway in early-stage resistance against powdery mildew in watermelon (Citrullus lanatus L.). Front. Plant Sci. 2022, 13, 1016822. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, R.; Liang, Y.; Gao, B.; Li, S.; Bai, W.; Oliver, M.J.; Zhang, D. The ScAPD1-like gene from the desert moss Syntrichia caninervis enhances resistance to Verticillium dahliae via phenylpropanoid gene regulation. Plant J. 2022, in press. [CrossRef]
- Ninkuu, V.; Yan, J.; Zhang, L.; Fu, Z.; Yang, T.; Li, S.; Li, B.; Duan, J.; Ren, J.; Li, G. Hrip1 mediates rice cell wall fortification and phytoalexins elicitation to confer immunity against Magnaporthe oryzae. Front. Plant Sci. 2022, 13, 980821. [Google Scholar] [CrossRef] [PubMed]
- Ke, X.; Wang, J.; Xu, X.; Guo, Y.; Zuo, Y.; Yin, L. Histological and molecular responses of Vigna angularis to Uromyces vignae infection. BMC Plant Biol. 2022, 22, 489. [Google Scholar] [CrossRef]
- Li, H.; Zhang, S.; Zhao, Y.; Zhao, X.; Xie, W.; Guo, Y.; Wang, Y.; Li, K.; Guo, J.; Zhu, Q.-H. Identification and Characterization of Cinnamyl Alcohol Dehydrogenase Encoding Genes Involved in Lignin Biosynthesis and Resistance to Verticillium dahliae in Upland Cotton (Gossypium hirsutum L.). Front. Plant Sci. 2022, 13, 840397. [Google Scholar] [CrossRef]
- Yang, Q.; Li, J.; Sun, J.; Cui, X. Comparative transcriptomic and proteomic analyses to determine the lignin synthesis pathway involved in the fungal stress response in Panax notoginseng. Physiol. Mol. Plant Pathol. 2022, 119, 101814. [Google Scholar] [CrossRef]
- Hussain, K.; Jaweed, T.H.; Kamble, A.C. Modulation of phenylpropanoid and lignin biosynthetic pathway is crucial for conferring resistance in pigeon pea against Fusarium wilt. Gene 2023, 851, 146994. [Google Scholar] [CrossRef]
- Duan, Y.; Ma, S.; Chen, X.; Shen, X.; Yin, C.; Mao, Z. Transcriptome changes associated with apple (Malus domestica) root defense response after Fusarium proliferatum f. sp. malus domestica infection. BMC Genom. 2022, 23, 484. [Google Scholar] [CrossRef]
- Lata, C.; Prasad, P.; Gangwar, O.P.; Adhikari, S.; Thakur, R.K.; Savadi, S.; Kumar, K.; Kumar, S.; Singh, G.P.; Bhardwaj, S.C. Temporal behavior of wheat—Puccinia striiformis interaction prompted defense-responsive genes. J. Plant Interact. 2022, 17, 674–684. [Google Scholar] [CrossRef]
- Zhu, Y.; Hu, X.; Wang, P.; Wang, H.; Ge, X.; Li, F.; Hou, Y. GhODO1, an R2R3-type MYB transcription factor, positively regulates cotton resistance to Verticillium dahliae via the lignin biosynthesis and jasmonic acid signaling pathway. Int. J. Biol. Macromol. 2022, 201, 580–591. [Google Scholar] [CrossRef] [PubMed]
- Faizah, R.; Putranto, R.A.; Raharti, V.R.; Supena, N.; Sukma, D.; Budiani, A.; Wening, S.; Sudarsono, S. Defense response changes in roots of oil palm (Elaeis guineensis Jacq.) seedlings after internal symptoms of Ganoderma boninense Pat. infection. BMC Plant Biol. 2022, 22, 139. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, Q.; Cheng, L.; Li, Q.; He, X.; Wang, K.; Liu, J.; Li, F.; Deng, Y. Pepper (Capsicum annuum) xylogen-like arabinogalactan protein (XYLP) 1 and XYLP2 promote synthesis of lignin during stem development to cope with stresses. Veg. Res. 2022, 2, 1–10. [Google Scholar] [CrossRef]
- Barros, J.; Shrestha, H.K.; Serrani-Yarce, J.C.; Engle, N.L.; Abraham, P.E.; Tschaplinski, T.J.; Hettich, R.L.; Dixon, R.A. Proteomic and metabolic disturbances in lignin-modified Brachypodium distachyon. Plant Cell 2022, 34, 3339–3363. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.-P.; Sun, S.-C.; Zhu, Q.-H.; Zhang, X.-Y.; Li, Y.-J.; Liu, F.; Xue, F.; Sun, J. The Cotton Lignin Biosynthetic Gene Gh4CL30 Regulates Lignification and Phenolic Content and Contributes to Verticillium Wilt Resistance. Mol. Plant-Microbe Interact. 2021, 34, 240–254. [Google Scholar] [CrossRef]
- Serrani-Yarce, J.C.; Escamilla-Trevino, L.; Barros, J.; Gallego-Giraldo, L.; Pu, Y.; Ragauskas, A.; Dixon, R.A. Targeting hydroxycinnamoyl CoA: Shikimate hydroxycinnamoyl transferase for lignin modification in Brachypodium distachyon. Biotechnol. Biofuels 2021, 14, 50. [Google Scholar] [CrossRef] [PubMed]
- Kriegshauser, L.; Knosp, S.; Grienenberger, E.; Tatsumi, K.; Gütle, D.D.; Sørensen, I.; Herrgott, L.; Zumsteg, J.; Rose, J.K.C.; Reski, R.; et al. Function of the Hydroxycinnamoyl-Coa:Shikimate Hydroxycinnamoyl Transferase is evolutionarily conserved in embryophytes. Plant Cell 2021, 33, 1472–1491. [Google Scholar] [CrossRef]
- Fang, H.; Zhang, F.; Zhang, C.; Wang, D.; Shen, S.; He, F.; Tao, H.; Wang, R.; Wang, M.; Wang, D.; et al. Function of hydroxycinnamoyl transferases for the biosynthesis of phenolamides in rice resistance to Magnaporthe oryzae. J. Genet. Genom. 2022, 49, 776–786. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, Y.; Zheng, K.; Xie, M.; Feng, K.; Jawdy, S.S.; Gunter, L.E.; Ranjan, P.; Singan, V.R.; Engle, N.; et al. Genome-wide association studies and expression-based quantitative trait loci analyses reveal roles of HCT2 in caffeoylquinic acid biosynthesis and its regulation by defense-responsive transcription factors in Populus. New Phytol. 2018, 220, 502–516. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chao, N.; Zhang, M.; Jiang, X.; Gai, Y. Functional Characteristics of Caffeoyl Shikimate Esterase in Larix Kaempferi and Monolignol Biosynthesis in Gymnosperms. Int. J. Mol. Sci. 2019, 20, 6071. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Huang, X.; Huang, H.; Huo, H.; Nguyen, C.D.; Pian, R.; Li, H.; Ouyang, K.; Chen, X. Cloning and characterization of the lignin biosynthesis genes NcCSE and NcHCT from Neolamarckia cadamba. AMB Express 2019, 9, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleme, M.d.L.S.; Cesarino, I.; Vargas, L.; Kim, H.; Vanholme, R.; Goeminne, G.; Van Acker, R.; Fonseca, F.C.d.A.; Pallidis, A.; Voorend, W.; et al. Silencing CAFFEOYL SHIKIMATE ESTERASE Affects Lignification and Improves Saccharification in Poplar. Plant Physiol. 2017, 175, 1040–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Tao, X.; Xie, Z.; Gong, X.; Qi, K.; Zhang, S.; Shiratake, K.; Tao, S. PbCSE1 promotes lignification during stone cell development in pear (Pyrus bretschneideri) fruit. Sci. Rep. 2021, 11, 9450. [Google Scholar] [CrossRef] [PubMed]
- de Vries, S.; Fürst-Jansen, J.M.R.; Irisarri, I.; Dhabalia Ashok, A.; Ischebeck, T.; Feussner, K.; Abreu, I.N.; Petersen, M.; Feussner, I.; de Vries, J. The evolution of the phenylpropanoid pathway entailed pronounced radiations and divergences of enzyme families. Plant J. 2021, 107, 975–1002. [Google Scholar] [CrossRef] [PubMed]
- Hafeez, A.; Gě, Q.; Zhāng, Q.; Lǐ, J.; Gōng, J.; Liú, R.; Shí, Y.; Shāng, H.; Liú, À.; Iqbal, M.S.; et al. Multi-responses of O-methyltransferase genes to salt stress and fiber development of Gossypium species. BMC Plant Biol. 2021, 21, 37. [Google Scholar] [CrossRef]
- Parizotto, A.V.; Ferro, A.P.; Marchiosi, R.; Finger-Teixeira, A.; Bevilaqua, J.M.; dos Santos, W.D.; Seixas, F.A.V.; Ferrarese-Filho, O. Inhibition of Maize Caffeate 3-O-Methyltransferase by Nitecapone as a Possible Approach to Reduce Lignocellulosic Biomass Recalcitrance. Plant Mol. Biol. Report. 2021, 39, 179–191. [Google Scholar] [CrossRef]
- Fu, Y.; Zhu, Y.; Yang, W.; Xu, W.; Li, Q.; Chen, M.; Yang, L. Isolation and functional identification of a Botrytis cinerea-responsive caffeoyl-CoA O-methyltransferase gene from Lilium regale wilson. Plant Physiol. Biochem. 2020, 157, 379–389. [Google Scholar] [CrossRef]
- Wang, M.; Zhu, X.; Wang, K.; Lu, C.; Luo, M.; Shan, T.; Zhang, Z. A wheat caffeic acid 3-O-methyltransferase TaCOMT-3D positively contributes to both resistance to sharp eyespot disease and stem mechanical strength. Sci. Rep. 2018, 8, 6543. [Google Scholar] [CrossRef]
- Narnoliya, L.K.; Sangwan, N.; Jadaun, J.S.; Bansal, S.; Sangwan, R.S. Defining the role of a caffeic acid 3-O-methyltransferase from Azadirachta indica fruits in the biosynthesis of ferulic acid through heterologous over-expression in Ocimum species and Withania somnifera. Planta 2021, 253, 20. [Google Scholar] [CrossRef]
- Figueiredo, R.; Portilla Llerena, J.P.; Kiyota, E.; Ferreira, S.S.; Cardeli, B.R.; de Souza, S.C.R.; Dos Santos Brito, M.; Sodek, L.; Cesarino, I.; Mazzafera, P. The sugarcane ShMYB78 transcription factor activates suberin biosynthesis in Nicotiana benthamiana. Plant Mol. Biol. 2020, 104, 411–427. [Google Scholar] [CrossRef]
- Hegde, N.; Joshi, S.; Soni, N.; Kushalappa, A.C. The caffeoyl-CoA O-methyltransferase gene SNP replacement in Russet Burbank potato variety enhances late blight resistance through cell wall reinforcement. Plant Cell Rep. 2021, 40, 237–254. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Pan, W.; Zhang, R.; Pan, Y.; Guo, Q.; Song, W.; Zheng, W.; Nie, X. Genome-wide identification and characterization of caffeoyl-coenzyme A O-methyltransferase genes related to the Fusarium head blight response in wheat. BMC Genom. 2021, 22, 504. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Zeng, Q.; Jiang, Y.; Gai, Y.; Jiang, X. Molecular and functional characterization of ferulate-5-hydroxylase in Populus tomentosa. J. Plant Biochem. Biotechnol. 2021, 30, 92–98. [Google Scholar] [CrossRef]
- Cao, Y.; Yan, X.; Ran, S.; Ralph, J.; Smith, R.; Chen, X.; Qu, C.; Li, J.; Liu, L. Knockout of the lignin pathway gene BnF5H decreases the S/G lignin composition ratio and improves S. sclerotiorum resistance in B. napus. Plant Cell Environ. 2022, 45, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Jeena, G.S.; Joshi, A.; Shukla, R.K. Bm-miR172c-5p regulates lignin biosynthesis and secondary xylem thickness by altering Ferulate 5 hydroxylase gene in Bacopa monnieri. Plant Cell Physiol. 2021, 62, 894–912. [Google Scholar] [CrossRef]
- Mutuku, J.M.; Cui, S.; Hori, C.; Takeda, Y.; Tobimatsu, Y.; Nakabayashi, R.; Mori, T.; Saito, K.; Demura, T.; Umezawa, T.; et al. The Structural Integrity of Lignin Is Crucial for Resistance against Striga hermonthica Parasitism in Rice. Plant Physiol. 2019, 179, 1796–1809. [Google Scholar] [CrossRef] [Green Version]
- Yin, N.; Li, B.; Liu, X.; Liang, Y.; Lian, J.; Xue, Y.; Qu, C.; Lu, K.; Wei, L.; Wang, R.; et al. Cinnamoyl-CoA Reductase 1 (CCR1) and CCR2 Function Divergently in Tissue Lignification, Flux Control and Cross-talk with Glucosinolate Pathway in Brassica napus. bioRxiv 2021. [Google Scholar] [CrossRef]
- Sugiyama, R.; Hirai, M.Y. Atypical Myrosinase as a Mediator of Glucosinolate Functions in Plants. Front. Plant Sci. 2019, 10, 1008. [Google Scholar] [CrossRef]
- Ting, H.-M.; Cheah, B.H.; Chen, Y.-C.; Yeh, P.-M.; Cheng, C.-P.; Yeo, F.K.S.; Vie, A.K.; Rohloff, J.; Winge, P.; Bones, A.M.; et al. The Role of a Glucosinolate-Derived Nitrile in Plant Immune Responses. Front. Plant Sci. 2020, 11, 257. [Google Scholar] [CrossRef]
- Madloo, P.; Lema, M.; Francisco, M.; Soengas, P. Role of Major Glucosinolates in the Defense of Kale Against Sclerotinia sclerotiorum and Xanthomonas campestris pv. campestris. Phytopathology® 2019, 109, 1246–1256. [Google Scholar] [CrossRef]
- Park, H.L.; Kim, T.L.; Bhoo, S.H.; Lee, T.H.; Lee, S.W.; Cho, M.H. Biochemical Characterization of the Rice Cinnamyl Alcohol Dehydrogenase Gene Family. Molecules 2018, 23, 2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, Y.; Kim, H.; Kang, M.; Lee, G.; Choung, S.; Kaur, H.; Oh, S.; Choi, J.W.; Ralph, J.; Baldwin, I.T.; et al. Pith-specific lignification in Nicotiana attenuata as a defense against a stem-boring herbivore. New Phytol. 2021, 232, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Mao, Q.; Wang, Y.; Zhao, J.; Fu, Y.; Yang, Z.; Peng, X.; Zhang, M.; Bai, B.; Liu, A.; et al. Trichoderma harzianum induces resistance to root-knot nematodes by increasing secondary metabolite synthesis and defense-related enzyme activity in Solanum lycopersicum L. Biol. Control 2021, 158, 104609. [Google Scholar] [CrossRef]
- Li, M.; Cheng, C.; Zhang, X.; Zhou, S.; Li, L.; Yang, S. Overexpression of Pear (Pyrus pyrifolia) CAD2 in Tomato Affects Lignin Content. Molecules 2019, 24, 2595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashyap, A.; Planas-Marquès, M.; Capellades, M.; Valls, M.; Coll, N.S. Blocking intruders: Inducible physico-chemical barriers against plant vascular wilt pathogens. J. Exp. Bot. 2020, 72, 184–198. [Google Scholar] [CrossRef]
- Xie, T.; Liu, Z.; Wang, G. Structural basis for monolignol oxidation by a maize laccase. Nat. Plants 2020, 6, 231–237. [Google Scholar] [CrossRef]
- Kidwai, M.; Ahmad, I.Z.; Chakrabarty, D. Class III peroxidase: An indispensable enzyme for biotic/abiotic stress tolerance and a potent candidate for crop improvement. Plant Cell Rep. 2020, 39, 1381–1393. [Google Scholar] [CrossRef]
- Li, Q.; Qin, X.; Qi, J.; Dou, W.; Dunand, C.; Chen, S.; He, Y. CsPrx25, a class III peroxidase in Citrus sinensis, confers resistance to citrus bacterial canker through the maintenance of ROS homeostasis and cell wall lignification. Hortic. Res. 2020, 7, 192. [Google Scholar] [CrossRef]
- Reitz, N.F.; Mitcham, E.J. Lignification of tomato (Solanum lycopersicum) pericarp tissue during blossom-end rot development. Sci. Hortic. 2021, 276, 109759. [Google Scholar] [CrossRef]
- Park, S.-C.; Pyun, J.W.; Jeong, Y.J.; Park, S.H.; Kim, S.; Kim, Y.-H.; Lee, J.R.; Kim, C.Y.; Jeong, J.C. Overexpression of VlPRX21 and VlPRX35 genes in Arabidopsis plants leads to bioconversion of trans-resveratrol to δ-viniferin. Plant Physiol. Biochem. 2021, 162, 556–563. [Google Scholar] [CrossRef]
- Meng, G.; Fan, W.; Rasmussen, S.K. Characterisation of the class III peroxidase gene family in carrot taproots and its role in anthocyanin and lignin accumulation. Plant Physiol. Biochem. 2021, 167, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Soni, N.; Hegde, N.; Dhariwal, A.; Kushalappa, A.C. Role of laccase gene in wheat NILs differing at QTL-Fhb1 for resistance against Fusarium head blight. Plant Sci. 2020, 298, 110574. [Google Scholar] [CrossRef] [PubMed]
- Bleau, J.R.; Spoel, S.H. Selective redox signaling shapes plant-pathogen interactions. Plant Physiol. 2021, 186, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Robe, K.; Izquierdo, E.; Vignols, F.; Rouached, H.; Dubos, C. The Coumarins: Secondary Metabolites Playing a Primary Role in Plant Nutrition and Health. Trends Plant Sci. 2021, 26, 248–259. [Google Scholar] [CrossRef]
- Stassen, M.J.J.; Hsu, S.-H.; Pieterse, C.M.J.; Stringlis, I.A. Coumarin Communication Along the Microbiome–Root–Shoot Axis. Trends Plant Sci. 2021, 26, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Perkowska, I.; Siwinska, J.; Olry, A.; Grosjean, J.; Hehn, A.; Bourgaud, F.; Lojkowska, E.; Ihnatowicz, A. Identification and Quantification of Coumarins by UHPLC-MS in Arabidopsis thaliana Natural Populations. Molecules 2021, 26, 1804. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, R.P.; Koprivova, A.; Kopriva, S. Pinpointing secondary metabolites that shape the composition and function of the plant microbiome. J. Exp. Bot. 2021, 72, 57–69. [Google Scholar] [CrossRef]
- Suksungworn, R.; Roytrakul, S.; Gomes, N.G.M.; Duangsrisai, S. A shotgun proteomic approach reveals protein expression in morphological changes and programmed cell death in Mimosa pigra seedlings after treatment with coumarins. South Afr. J. Bot. 2021, 142, 370–379. [Google Scholar] [CrossRef]
- Gabaston, J.; Valls Fonayet, J.; Franc, C.; Waffo-Teguo, P.; de Revel, G.; Hilbert, G.; Gomès, E.; Richard, T.; Mérillon, J.-M. Characterization of Stilbene Composition in Grape Berries from Wild Vitis Species in Year-To-Year Harvest. J. Agric. Food Chem. 2020, 68, 13408–13417. [Google Scholar] [CrossRef]
- Wang, D.; Jiang, C.; Liu, W.; Wang, Y. The WRKY53 transcription factor enhances stilbene synthesis and disease resistance by interacting with MYB14 and MYB15 in Chinese wild grape. J. Exp. Bot. 2020, 71, 3211–3226. [Google Scholar] [CrossRef]
- Ziegler, T. Identification and Characterization of Genes Involved in Stilbene Biosynthesis and Modification in Vitis Vinifera; 2021. Available online: https://archiv.ub.uni-heidelberg.de/volltextserver/30000/ (accessed on 16 December 2022).
- Valletta, A.; Iozia, L.M.; Leonelli, F. Impact of Environmental Factors on Stilbene Biosynthesis. Plants 2021, 10, 90. [Google Scholar] [CrossRef] [PubMed]
- El Khawand, T.; Gabaston, J.; Taillis, D.; Iglesias, M.-L.; Pedrot, É.; Pinto, A.P.; Fonayet, J.V.; Merillon, J.M.; Decendit, A.; Cluzet, S.A. dimeric stilbene extract produced by oxidative coupling of resveratrol active against Plasmopara viticola and Botrytis cinerea for vine treatments. OENO One 2020, 54, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Song, P.; Yu, X.; Yang, W.; Wang, Q. Natural phytoalexin stilbene compound resveratrol and its derivatives as anti-tobacco mosaic virus and anti-phytopathogenic fungus agents. Sci. Rep. 2021, 11, 16509. [Google Scholar] [CrossRef] [PubMed]
- Bezhuashvili, M.; Tskhvedadze, L.; Surguladze, M.; Shoshiashvili, G.; Elanidze, L.; Vashakidze, P. Change of phytoalexins-stilbenoids of vine leave Tsitska variety (Vitis vinifera L.) in condition Downy mildew. EurAsian J. BioSci. 2020, 14, 167–171. [Google Scholar]
- Marchiosi, R.; dos Santos, W.D.; Constantin, R.P.; de Lima, R.B.; Soares, A.R.; Finger-Teixeira, A.; Mota, T.R.; de Oliveira, D.M.; Foletto-Felipe, M.d.P.; Abrahao, J. Biosynthesis and metabolic actions of simple phenolic acids in plants. Phytochem. Rev. 2020, 19, 865–906. [Google Scholar] [CrossRef]
- Valanciene, E.; Jonuskiene, I.; Syrpas, M.; Augustiniene, E.; Matulis, P.; Simonavicius, A.; Malys, N. Advances and Prospects of Phenolic Acids Production, Biorefinery and Analysis. Biomolecules 2020, 10, 874. [Google Scholar] [CrossRef]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef] [Green Version]
- Anguraj Vadivel, A.K.; Renaud, J.; Kagale, S.; Dhaubhadel, S. GmMYB176 Regulates Multiple Steps in Isoflavonoid Biosynthesis in Soybean. Front. Plant Sci. 2019, 10, 562. [Google Scholar] [CrossRef] [Green Version]
- Hano, C.F.; Dinkova-Kostova, A.T.; Davin, L.B.; Cort, J.R.; Lewis, N.G. Editorial: Lignans: Insights Into Their Biosynthesis, Metabolic Engineering, Analytical Methods and Health Benefits. Front. Plant Sci. 2021, 11, 630327. [Google Scholar] [CrossRef]
- Markulin, L.; Corbin, C.; Renouard, S.; Drouet, S.; Gutierrez, L.; Mateljak, I.; Auguin, D.; Hano, C.; Fuss, E.; Lainé, E. Pinoresinol–lariciresinol reductases, key to the lignan synthesis in plants. Planta 2019, 249, 1695–1714. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ninkuu, V.; Yan, J.; Fu, Z.; Yang, T.; Ziemah, J.; Ullrich, M.S.; Kuhnert, N.; Zeng, H. Lignin and Its Pathway-Associated Phytoalexins Modulate Plant Defense against Fungi. J. Fungi 2023, 9, 52. https://doi.org/10.3390/jof9010052
Ninkuu V, Yan J, Fu Z, Yang T, Ziemah J, Ullrich MS, Kuhnert N, Zeng H. Lignin and Its Pathway-Associated Phytoalexins Modulate Plant Defense against Fungi. Journal of Fungi. 2023; 9(1):52. https://doi.org/10.3390/jof9010052
Chicago/Turabian StyleNinkuu, Vincent, Jianpei Yan, Zenchao Fu, Tengfeng Yang, James Ziemah, Matthias S. Ullrich, Nikolai Kuhnert, and Hongmei Zeng. 2023. "Lignin and Its Pathway-Associated Phytoalexins Modulate Plant Defense against Fungi" Journal of Fungi 9, no. 1: 52. https://doi.org/10.3390/jof9010052