
Specific Focus on Antifungal Peptides against Azole Resistant Aspergillus fumigatus: Current Status, Challenges, and Future Perspectives
 , and
, and
Abstract
:1. Introduction
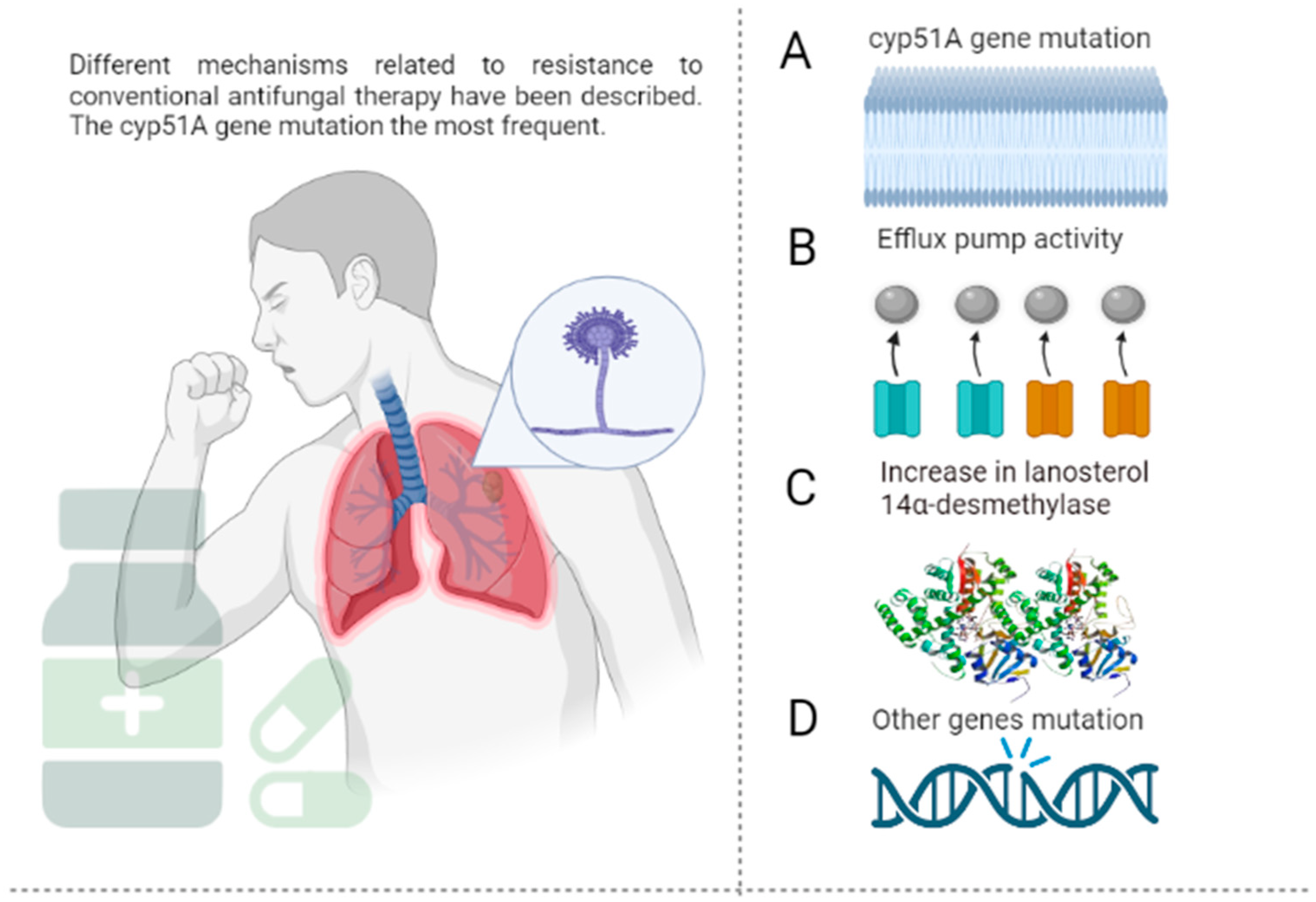
2. Epidemiology and Mechanism of Resistance of A. fumigatus to Antifungal Agents
3. Alternative Therapy
3.1. Antifungal Peptides

{kind=link}
{kind=link}
| Author/Year | Name | Source | Length | Net Charge | Hydrophobic Residues | Boman Index | MIC |
|---|---|---|---|---|---|---|---|
| Mignone et al., 2022 [65] | Sin a 1 | Seeds, white mustard, Brassica hirta | 145 | 10 | 32% | 2.09 | 63 µM |
| Seyedjavadi et al., 2019 [66] | M. chamomilla AMP 1 | Matricaria chamomilla L. | 23 | 3 | 47% | 0.68 | 6.66 µM |
| Khani et al., 2019 [67] | Skh-AMP1 | Leaves, Satureja khuzistanica | 25 | 5 | 28% | 3.19 | 20.7 µM |
| Xiaoxia et al., 2019 [68] | P. xylostella Moricin | Highly expressed in fat body and hemocyte, diamondback moth, Plutella xylostella | 42 | 9 | 35% | 1.67 | 8.9–23 µM |
| Park et al., 2016 [69] | Human alpha-synuclein | Brain, Homo sapiens | 140 | −9 | 35% | 1.3 | 0.8–3.2 µM |
| Bellmonte et al., 2012 [70] | Hb 98–114 | Tick midgut, Rhipicephalus (Boophilus) microplus | 17 | 3 | 52% | −0.62 | 6.3 µM |
| Rodríguez et al., 2010 [71] | PgAFP | Penicillium chrysogenum RP42C; also found in Penicillium chrysogenum Q176 | 58 | 4 | 27% | 2.63 | 0.12–1.0 µM |
| Gao et al., 2009 [72] | Meucin-18 | Mesobuthus eupeus | 18 | 2 | 55% | −0.66 | 1.9–8.3 µM |
| Simon et al., 2008 [73] | Human drosomycin-like defensin | Mainly expressed in skin (mRNA), Homo sapiens | 43 | 5 | 25% | 3.58 | 6.25 µM |
| Cabras et al., 2008 [74] | SP-B | Porcine salivary gland granules | 21 | 1 | 9% | 0.35 | 58.68 µM |
| Briolat et al., 2005 [75] | Catestatin | Skin, Homo sapiens | 21 | 4 | 33% | 1.98 | 80 µM |
| Briolat et al., 2005 [75] | Cateslytin | Chromaffin cells and in secretion medium, bovine | 15 | 5 | 33% | 4.3 | 10 µM |
| Landon et al., 2004 [76] | ARD1 | Archaeoprepona demophoon | 41 | 3 | 39% | 1.6 | ND |
| Kaiserer et al., 2003 [77] | Penicillium antifungal protein | Penicillium chrysogenum | 55 | 5 | 25% | 3.12 | ND |
| Lauth et al., 2002 [78] | wb-Moronecidin | Skin/gill, Morone saxatilis | 23 | 3 | 43% | 0.38 | 50–100 µM |
| Silva et al., 2000 [79] | Gomesin | Hemocytes, Acanthoscurria gomesiana | 18 | 6 | 33% | 4.39 | ND |
| Lugardon et al., 2000 [80] | Vasostatin-1 | Bovine chromaffin granules, Bos taurus | 76 | −1 | 35% | 2.03 | 1–10 µM |
| Gun et al., 1999 [81] | AnAFP | Aspergillus niger | 58 | 5 | 28% | 2.42 | 4–8 µM |
| Gallo et al., 1997 [82] | Mouse cathelin-related antimicrobial peptide | Adult testis, spleen, stomach, and intestine, mouse, Mus musculus | 34 | 6 | 29% | 1.74 | 100 µM |
| Lawyer et al., 1996 [83] | Tritrpticin | Synthetic fragment of porcine cathelicidin. | 13 | 4 | 53% | 2.9 | ND |
| Ehret et al., 1996 [84] | Androctonin | Androctonus australis | 25 | 8 | 28% | 3.9 | 25–50 µM |
| Mor et al., 1994 [85] | Dermaseptin-B2 | skin, giant leaf frog, Phyllomedusa bicolor, South America | 33 | 4 | 54% | 0.23 | 125 µg/mL |
| Mor et al., 1994 [85] | Dermaseptin-S2 | Sauvage’s leaf frog, Phyllomedusa sauvagii, South America | 34 | 3 | 52% | −0.14 | 20 µM |
| Mor et al., 1994 [86] | Dermaseptin-S3 | Sauvage’s leaf frog, Phyllomedusa sauvagii, South America | 30 | 6 | 53% | −0.25 | 10–20 µM |
| Mor et al., 1994 [86] | Dermaseptin-S4 | Sauvage’s leaf frog, Phyllomedusa sauvagii, South America | 28 | 4 | 71% | −0.91 | 20–30 µM |
| Mor et al., 1994 [87] | Skin peptide tyrosine-tyrosine | Skin, the South American arboreal frog Phyllomedusa bicolor | 36 | 1 | 22% | 2.69 | 100 µg/mL |
| Fehlbaum et al., 1994 [88] | Drosomycin | Fruitfly, Drosophila melanogaster | 44 | 1 | 34% | 2.56 | 6.25 µM |
| Bellamy et al., 1992 [89] | Lactoferricin B | Bos taurus | 25 | 8 | 48% | 2.75 | ND |
| Mor et al., 1991 [90] | Dermaseptin-S1 | Sauvage’s leaf frog, Phyllomedusa sauvagii, South America | 34 | 3 | 50% | 0.16 | 30 µM |
| Wnendt et al., 1990 [91] | Antifungal protein | Aspergillus giganteus | 51 | 9 | 31% | 2.1 | 1 µM |
| Miller et al., 1989 [92] | Secretory leukocyte protease inhibitor | Tears, saliva, airway, gastrointestines, genital tracts, Homo sapiens | 107 | 12 | 34% | 1.87 | ND |
3.1.1. Synthetic Antifungal Peptides
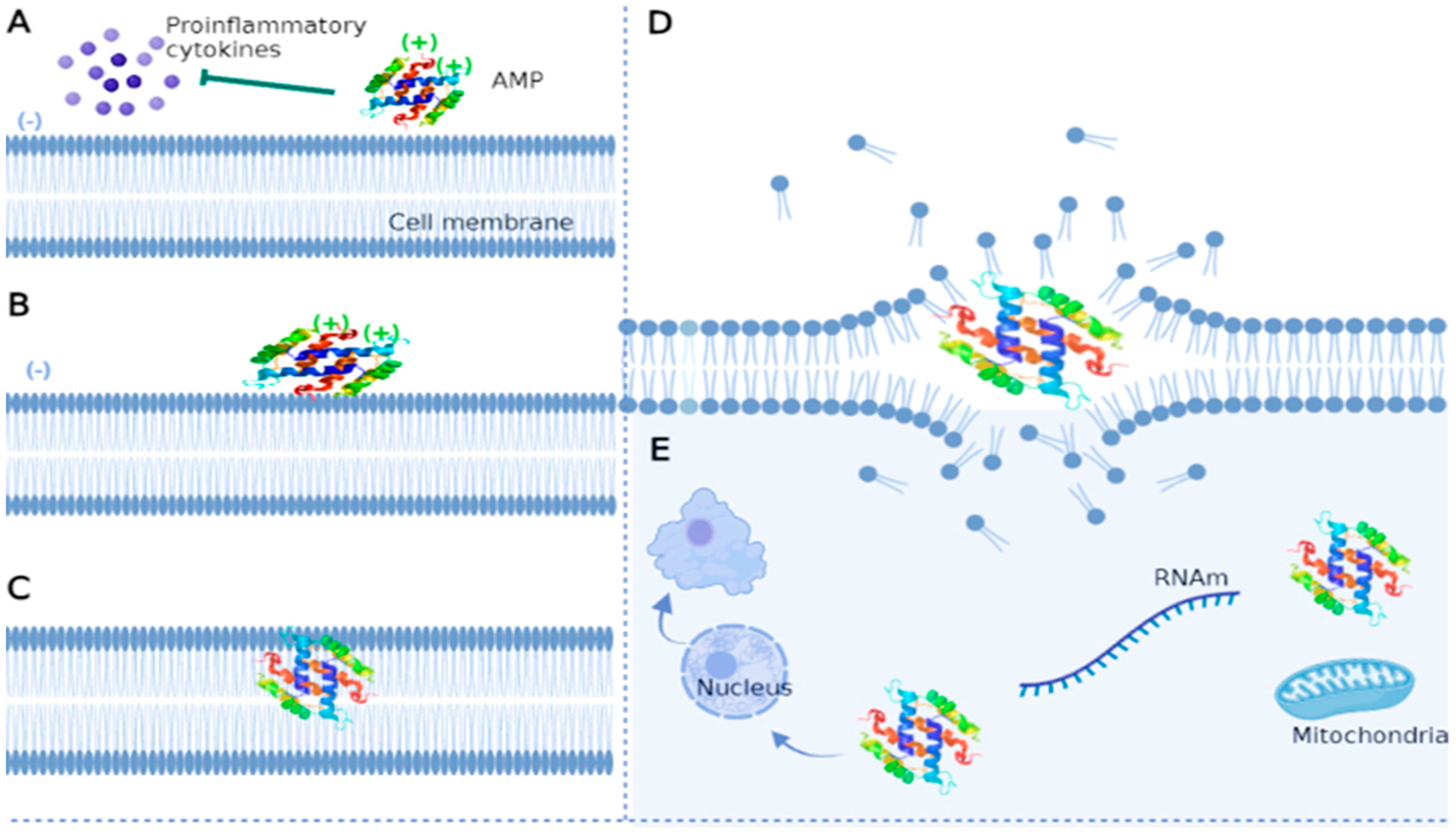
3.1.2. Mechanism of Action of Antifungal Peptides
Cell Membrane-Targeted Antifungal Peptides
Cell Wall-Targeted Antifungal Peptides
Antifungal Peptides Targeting Intracellular Molecules and Structures
3.1.3. Limitations in the Use of Peptides as Antifungals
Route of Administration
Selectivity and Toxicity
Peptide Stability
Pharmacological Resistance
4. Future Perspectives and Challenges Related to the Use of Antifungal Peptides
4.1. Other Potential Alternatives to Combat A. fumigatus
4.1.1. Vaccines for the Prevention of Aspergillosis
4.1.2. Nanotechnology to Combat Aspergillus fumigatus
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cadena, J.; Thompson, G.R.; Patterson, T.F. Aspergillosis. Infect. Dis. Clin. N. Am. 2021, 35, 415–434. [Google Scholar] [CrossRef] [PubMed]
- Marr, K.A.; Platt, A.; Tornheim, J.A.; Zhang, S.X.; Datta, K.; Cardozo, C.; Garcia-Vidal, C. Aspergillosis Complicating Severe Coronavirus Disease. Emerg. Infect. Dis. 2021, 27, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Spallone, A.; Schwartz, I.S. Emerging Fungal Infections. Infect. Dis. Clin. N. Am. 2021, 35, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Prattes, J.; Valentin, T.; Hoenigl, M.; Talakic, E.; Reisinger, A.C.; Eller, P. Invasive Pulmonary Aspergillosis Complicating COVID-19 in the ICU—A Case Report. Med. Mycol. Case Rep. 2021, 31, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Seagle, E.E.; Williams, S.L.; Chiller, T.M. Recent Trends in the Epidemiology of Fungal Infections. Infect. Dis. Clin. N. Am. 2021, 35, 237–260. [Google Scholar] [CrossRef] [PubMed]
- Girmenia, C.; Ferretti, A.; Barberi, W. Epidemiology and Risk Factors for Invasive Fungal Diseases in Hematopoietic Stem Cell Transplantation. Curr. Opin. Hematol. 2014, 21, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Everaerts, S.; Lagrou, K.; Dubbeldam, A.; Lorent, N.; Vermeersch, K.; Van Hoeyveld, E.; Bossuyt, X.; Dupont, L.; Vanaudenaerde, B.; Janssens, W. Sensitization to Aspergillus Fumigatus as a Risk Factor for Bronchiectasis in COPD. COPD 2017, 12, 2629–2638. [Google Scholar] [CrossRef] [Green Version]
- Segrelles-Calvo, G.; Araújo, G.R.S.; Llopis-Pastor, E.; Carrillo, J.; Hernández-Hernández, M.; Rey, L.; Rodríguez Melean, N.; Escribano, I.; Antón, E.; Zamarro, C.; et al. Prevalence of Opportunistic Invasive Aspergillosis in COVID-19 Patients with Severe Pneumonia. Mycoses 2021, 64, 144–151. [Google Scholar] [CrossRef]
- Namvar, S.; Labram, B.; Rowley, J.; Herrick, S. Aspergillus Fumigatus—Host Interactions Mediating Airway Wall Remodelling in Asthma. JoF 2022, 8, 159. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.; Tiseo, G.; Falcone, M.; Menichetti, F. Pulmonary Aspergillosis: An Evolving Challenge for Diagnosis and Treatment. Infect. Dis. Ther. 2020, 9, 511–524. [Google Scholar] [CrossRef]
- Patterson, T.F.; Thompson, G.R.; Denning, D.W.; Fishman, J.A.; Hadley, S.; Herbrecht, R.; Kontoyiannis, D.P.; Marr, K.A.; Morrison, V.A.; Nguyen, M.H.; et al. Practice Guidelines for the Diagnosis and Management of Aspergillosis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 63, e1–e60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulussen, C.; Hallsworth, J.E.; Álvarez-Pérez, S.; Nierman, W.C.; Hamill, P.G.; Blain, D.; Rediers, H.; Lievens, B. Ecology of Aspergillosis: Insights into the Pathogenic Potency of Aspergillus Fumigatus and Some Other Aspergillus Species. Microb. Biotechnol. 2017, 10, 296–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon-Chung, K.J.; Sugui, J.A. Aspergillus Fumigatus—What Makes the Species a Ubiquitous Human Fungal Pathogen? PLoS Pathog. 2013, 9, e1003743. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Elders, B.B.L.J.; Warris, A.; Caudri, D.; Ciet, P.; Tiddens, H.A.W.M. Aspergillus-Related Lung Disease in People with Cystic Fibrosis: Can Imaging Help Us to Diagnose Disease? Eur. Respir. Rev. 2021, 30, 210103. [Google Scholar] [CrossRef]
- Taccone, F.S.; Van den Abeele, A.-M.; Bulpa, P.; Misset, B.; Meersseman, W.; Cardoso, T.; Paiva, J.-A.; Blasco-Navalpotro, M.; De Laere, E.; Dimopoulos, G.; et al. Epidemiology of Invasive Aspergillosis in Critically Ill Patients: Clinical Presentation, Underlying Conditions, and Outcomes. Crit. Care 2015, 19, 7. [Google Scholar] [CrossRef] [Green Version]
- Moura, S.; Cerqueira, L.; Almeida, A. Invasive Pulmonary Aspergillosis: Current Diagnostic Methodologies and a New Molecular Approach. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1393–1403. [Google Scholar] [CrossRef]
- McCashney, A.; Robinson, P. Structural Lung Disease Following Allergic Bronchopulmonary Aspergillosis Complicating Pediatric Cystic Fibrosis. Pediatr. Pulmonol. 2021, 56, 3737–3744. [Google Scholar] [CrossRef]
- Denning, D.W.; Pleuvry, A.; Cole, D.C. Global Burden of Allergic Bronchopulmonary Aspergillosis with Asthma and Its Complication Chronic Pulmonary Aspergillosis in Adults. Med. Mycol. 2013, 51, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Zilberberg, M.D.; Nathanson, B.H.; Harrington, R.; Spalding, J.R.; Shorr, A.F. Epidemiology and Outcomes of Hospitalizations with Invasive Aspergillosis in the United States, 2009–2013. Clin. Infect. Dis. 2018, 67, 727–735. [Google Scholar] [CrossRef]
- Alvarez-Moreno, C.; Cortes, J.; Denning, D. Burden of Fungal Infections in Colombia. JoF 2018, 4, 41. [Google Scholar] [CrossRef]
- Alastruey-Izquierdo, A.; Cadranel, J.; Flick, H.; Godet, C.; Hennequin, C.; Hoenigl, M.; Kosmidis, C.; Lange, C.; Munteanu, O.; Page, I.; et al. Treatment of Chronic Pulmonary Aspergillosis: Current Standards and Future Perspectives. Respiration 2018, 96, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Jenks, J.; Hoenigl, M. Treatment of Aspergillosis. JoF 2018, 4, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbrecht, R.; Patterson, T.F.; Slavin, M.A.; Marchetti, O.; Maertens, J.; Johnson, E.M.; Schlamm, H.T.; Donnelly, J.P.; Pappas, P.G. Application of the 2008 Definitions for Invasive Fungal Diseases to the Trial Comparing Voriconazole Versus Amphotericin B for Therapy of Invasive Aspergillosis: A Collaborative Study of the Mycoses Study Group (MSG 05) and the European Organization for Research and Treatment of Cancer Infectious Diseases Group. Clin. Infect. Dis. 2015, 60, 713–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lestrade, P.P.; Bentvelsen, R.G.; Schauwvlieghe, A.F.A.D.; Schalekamp, S.; van der Velden, W.J.F.M.; Kuiper, E.J.; van Paassen, J.; van der Hoven, B.; van der Lee, H.A.; Melchers, W.J.G.; et al. Voriconazole Resistance and Mortality in Invasive Aspergillosis: A Multicenter Retrospective Cohort Study. Clin. Infect. Dis. 2019, 68, 1463–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebrehiwet, T.; Gebremichael, G. Development of Vaccination against Fungal Disease: A Review Article. Int. J. Trop. Dis. 2018, 1, 1–8. [Google Scholar] [CrossRef] [Green Version]
- El-Diasty, E.M.; Ahmed, M.A.; Okasha, N.; Mansour, S.F.; El-Dek, S.I.; El-Khalek, H.M.A.; Youssif, M.H. Antifungal activity of zinc oxide nanoparticles against dermatophytic lesions of cattle. Rom. J. Biophys. 2013, 23, 191–202. [Google Scholar]
- Lucca, A.J.D.; Walsh, T.J. Antifungal Peptides: Origin, Activity, and Therapeutic Potential. Rev. Iberoam. Micol. 2000, 17, 116–120. [Google Scholar]
- Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A.; Giulianotti, M.A.; Apidianakis, Y.; Bradfute, S.; Ferguson, A.L.; et al. The Value of Antimicrobial Peptides in the Age of Resistance. Lancet Infect. Dis. 2020, 20, e216–e230. [Google Scholar] [CrossRef]
- Ballard, E.; Yucel, R.; Melchers, W.J.G.; Brown, A.J.P.; Verweij, P.E.; Warris, A. Antifungal Activity of Antimicrobial Peptides and Proteins against Aspergillus Fumigatus. JoF 2020, 6, 65. [Google Scholar] [CrossRef]
- Lupetti, A.; van Dissel, J.T.; Brouwer, C.P.J.M.; Nibbering, P.H. Human Antimicrobial Peptides’ Antifungal Activity against Aspergillus Fumigatus. Eur. J. Clin. Microbiol. Infect. Dis. 2008, 27, 1125–1129. [Google Scholar] [CrossRef] [Green Version]
- Denning, D.W.; Venkateswarlu, K.; Oakley, K.L.; Anderson, M.J.; Manning, N.J.; Stevens, D.A.; Warnock, D.W.; Kelly, S.L. Itraconazole Resistance in Aspergillus Fumigatus. Antimicrob. Agents Chemother. 1997, 41, 1364–1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kidd, S.E.; Goeman, E.; Meis, J.F.; Slavin, M.A.; Verweij, P.E. Multi-Triazole-Resistant Aspergillus Fumigatus Infections in Australia. Mycoses 2015, 58, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Meis, J.F.; Chowdhary, A.; Rhodes, J.L.; Fisher, M.C.; Verweij, P.E. Clinical Implications of Globally Emerging Azole Resistance in Aspergillus Fumigatus. Phil. Trans. R. Soc. B 2016, 371, 20150460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verweij, P.E.; Chowdhary, A.; Melchers, W.J.G.; Meis, J.F. Azole Resistance in Aspergillus Fumigatus: Can We Retain the Clinical Use of Mold-Active Antifungal Azoles? Clin. Infect. Dis. 2016, 62, 362–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Linden, J.W.M.; Snelders, E.; Kampinga, G.A.; Rijnders, B.J.A.; Mattsson, E.; Debets-Ossenkopp, Y.J.; Kuijper, E.J.; Van Tiel, F.H.; Melchers, W.J.G.; Verweij, P.E. Clinical Implications of Azole Resistance in Aspergillus Fumigatus, the Netherlands, 2007–2009. Emerg. Infect. Dis. 2011, 17, 1846–1854. [Google Scholar] [CrossRef]
- van der Linden, J.W.M.; Arendrup, M.C.; Warris, A.; Lagrou, K.; Pelloux, H.; Hauser, P.M.; Chryssanthou, E.; Mellado, E.; Kidd, S.E.; Tortorano, A.M.; et al. Prospective Multicenter International Surveillance of Azole Resistance in Aspergillus Fumigatus. Emerg. Infect. Dis. 2015, 21, 1041–1044. [Google Scholar] [CrossRef]
- Perlin, D.S.; Rautemaa-Richardson, R.; Alastruey-Izquierdo, A. The Global Problem of Antifungal Resistance: Prevalence, Mechanisms, and Management. Lancet Infect. Dis. 2017, 17, e383–e392. [Google Scholar] [CrossRef]
- Resendiz Sharpe, A.; Lagrou, K.; Meis, J.F.; Chowdhary, A.; Lockhart, S.R.; Verweij, P.E.; on behalf of the ISHAM/ECMM. Aspergillus Resistance Surveillance working group Triazole Resistance Surveillance in Aspergillus Fumigatus. Med. Mycol. 2018, 56, S83–S92. [Google Scholar] [CrossRef] [Green Version]
- Hurst, S.F.; Berkow, E.L.; Stevenson, K.L.; Litvintseva, A.P.; Lockhart, S.R. Isolation of Azole-Resistant Aspergillus Fumigatus from the Environment in the South-Eastern USA. J. Antimicrob. Chemother. 2017, 72, 2443–2446. [Google Scholar] [CrossRef]
- Negri, C.E.; Gonçalves, S.S.; Sousa, A.C.P.; Bergamasco, M.D.; Martino, M.D.V.; Queiroz-Telles, F.; Aquino, V.R.; Castro, P.d.T.O.; Hagen, F.; Meis, J.F.; et al. Triazole Resistance Is Still Not Emerging in Aspergillus Fumigatus Isolates Causing Invasive Aspergillosis in Brazilian Patients. Antimicrob. Agents Chemother. 2017, 61, e00608-17. [Google Scholar] [CrossRef] [Green Version]
- Bustamante, B.; Illescas, L.R.; Posadas, A.; Campos, P.E. Azole Resistance among Clinical Isolates of Aspergillus Fumigatus in Lima-Peru. Med. Mycol. 2020, 58, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.; Messina, F.; Marin, E.; Arechavala, A.; Depardo, R.; Walker, L.; Negroni, R.; Santiso, G. Antifungal Resistance in Clinical Isolates of Aspergillus spp.: When Local Epidemiology Breaks the Norm. JoF 2019, 5, 41. [Google Scholar] [CrossRef] [PubMed]
- Le Pape, P.; Lavergne, R.; Morio, F.; Alvarez, C. Multiple Fungicide-Driven Alterations in Azole-Resistant Aspergillus Fumigatus, Colombia, 2015. Emerg. Infect. Dis. 2016, 22, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roundtree, M.T.; Juvvadi, P.R.; Shwab, E.K.; Cole, D.C.; Steinbach, W.J. Aspergillus Fumigatus Cyp51A and Cyp51B Proteins Are Compensatory in Function and Localize Differentially in Response to Antifungals and Cell Wall Inhibitors. Antimicrob. Agents Chemother. 2020, 64, e00735-20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, L.; Lv, Q.; Yan, L.; Wang, Y.; Jiang, Y. The Fungal CYP51s: Their Functions, Structures, Related Drug Resistance, and Inhibitors. Front. Microbiol. 2019, 10, 691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Liu, J.; Zeng, M.; Sang, H. Exploring the Molecular Mechanism of Azole Resistance in Aspergillus Fumigatus. J. Mycol. Médicale 2020, 30, 100915. [Google Scholar] [CrossRef]
- Fraczek, M.G.; Bromley, M.; Buied, A.; Moore, C.B.; Rajendran, R.; Rautemaa, R.; Ramage, G.; Denning, D.W.; Bowyer, P. The Cdr1B Efflux Transporter Is Associated with Non-Cyp51a-Mediated Itraconazole Resistance in Aspergillus Fumigatus. J. Antimicrob. Chemother. 2013, 68, 1486–1496. [Google Scholar] [CrossRef]
- Moazeni, M.; Aslani, N.; Nabili, M.; Badali, H. Overexpression of Efflux Pump Genes Is an Alternative Mechanism in Voriconazole Resistant Aspergillus Fumigatus Isolates Without Relative Mutations in CYP5A. IDDT 2021, 20, 860–866. [Google Scholar] [CrossRef]
- Quintana, S.C.; Sjostrom, P.D.; Baldeón, G.M.; Socarrás, D.A.; Paz, M.C.; Molina, A.H. Genome of Candida albicans and drug resistance. Sun 2017, 33, 438–450. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y.; Lu, L. Calcium Signaling Pathway Is Involved in Non-CYP51 Azole Resistance in Aspergillus Fumigatus. Med. Mycol. 2019, 57, S233–S238. [Google Scholar] [CrossRef]
- Liu, F.; Pu, L.; Zheng, Q.; Zhang, Y.; Gao, R.; Xu, X.; Zhang, S.; Lu, L. Calcium Signaling Mediates Antifungal Activity of Triazole Drugs in the Aspergilli. Fungal Genet. Biol. 2015, 81, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Chen, P.; Gao, R.; Li, Y.; Zhang, A.; Liu, F.; Lu, L. Screening and Characterization of a Non-Cyp51A Mutation in an Aspergillus Fumigatus Cox10 Strain Conferring Azole Resistance. Antimicrob. Agents Chemother. 2017, 61, e02101-16. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, D.; Arai, T.; Takahashi, H.; Kusuya, Y.; Watanabe, A.; Kamei, K. Non-Cyp51A Azole-Resistant Aspergillus Fumigatus Isolates with Mutation in HMG-CoA Reductase. Emerg. Infect. Dis. 2018, 24, 1889–1897. [Google Scholar] [CrossRef] [Green Version]
- Bahar, A.; Ren, D. Antimicrobial Peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Gallo, R.L. Antimicrobial Peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef]
- Lima, P.G.; Oliveira, J.T.A.; Amaral, J.L.; Freitas, C.D.T.; Souza, P.F.N. Synthetic Antimicrobial Peptides: Characteristics, Design, and Potential as Alternative Molecules to Overcome Microbial Resistance. Life Sci. 2021, 278, 119647. [Google Scholar] [CrossRef]
- Chen, Y.; Mant, C.T.; Hodges, R.S. Determination of Stereochemistry Stability Coefficients of Amino Acid Side-Chains in an Amphipathic Alpha-Helix. J. Pept. Res. 2002, 59, 18–33. [Google Scholar] [CrossRef]
- Kovacs, J.M.; Mant, C.T.; Hodges, R.S. Determination of Intrinsic Hydrophilicity/Hydrophobicity of Amino Acid Side Chains in Peptides in the Absence of Nearest-Neighbor or Conformational Effects. Biopolymers 2006, 84, 283–297. [Google Scholar] [CrossRef] [Green Version]
- Lyu, Y.; Yang, Y.; Lyu, X.; Dong, N.; Shan, A. Antimicrobial Activity, Improved Cell Selectivity and Mode of Action of Short PMAP-36-Derived Peptides against Bacteria and Candida. Sci. Rep. 2016, 6, 27258. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.; Park, S.-C.; Park, H.-K.; Shin, S.Y.; Kim, Y.; Hahm, K.-S. Structure-Activity Relationship of HP (2–20) Analog Peptide: Enhanced Antimicrobial Activity by N-Terminal Random Coil Region Deletion. Biopolymers 2007, 88, 199–207. [Google Scholar] [CrossRef]
- Benincasa, M.; Skerlavaj, B.; Gennaro, R.; Pellegrini, A.; Zanetti, M. In Vitro and in Vivo Antimicrobial Activity of Two α-Helical Cathelicidin Peptides and of Their Synthetic Analogs. Peptides 2003, 24, 1723–1731. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, R.I.; Ganz, T. Defensins of Vertebrate Animals. Curr. Opin. Immunol. 2002, 14, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.W.; Chapple, D.S. Peptide Antibiotics. Antimicrob. Agents Chemother. 1999, 43, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hancock, R.E. Cationic Antimicrobial Peptides: Towards Clinical Applications. Expert Opin. Investig. Drugs 2000, 9, 1723–1729. [Google Scholar] [CrossRef] [PubMed]
- Mignone, G.; Shwaiki, L.N.; Arendt, E.K.; Coffey, A. Isolation of the Mustard Napin Protein Allergen Sin a 1 and Characterisation of Its Antifungal Activity. Biochem. Biophys. Rep. 2022, 29, 101208. [Google Scholar] [CrossRef] [PubMed]
- Seyedjavadi, S.S.; Khani, S.; Zare-Zardini, H.; Halabian, R.; Goudarzi, M.; Khatami, S.; Imani Fooladi, A.A.; Amani, J.; Razzaghi-Abyaneh, M. Isolation, Functional Characterization, and Biological Properties of MCh-AMP1, a Novel Antifungal Peptide from Matricaria Chamomilla L. Chem. Biol. Drug Des. 2019, 93, 949–959. [Google Scholar] [CrossRef]
- Khani, S.; Seyedjavadi, S.S.; Zare-Zardini, H.; Hosseini, H.M.; Goudarzi, M.; Khatami, S.; Amani, J.; Fooladi, A.A.I.; Razzaghi-Abyaneh, M. Isolation and Functional Characterization of an Antifungal Hydrophilic Peptide, Skh-AMP1, Derived from Satureja Khuzistanica Leaves. Phytochemistry 2019, 164, 136–143. [Google Scholar] [CrossRef]
- Xu, X.; Zhong, A.; Wang, Y.; Lin, B.; Li, P.; Ju, W.; Zhu, X.; Yu, J.; De Mandal, S.; Jin, F. Molecular Identification of a Moricin Family Antimicrobial Peptide (Px-Mor) From Plutella Xylostella With Activities Against the Opportunistic Human Pathogen Aureobasidium Pullulans. Front. Microbiol. 2019, 10, 2211. [Google Scholar] [CrossRef]
- Park, S.-C.; Moon, J.C.; Shin, S.Y.; Son, H.; Jung, Y.J.; Kim, N.-H.; Kim, Y.-M.; Jang, M.-K.; Lee, J.R. Functional Characterization of Alpha-Synuclein Protein with Antimicrobial Activity. Biochem. Biophys. Res. Commun. 2016, 478, 924–928. [Google Scholar] [CrossRef]
- Belmonte, R.; Cruz, C.E.; Pires, J.R.; Daffre, S. Purification and Characterization of Hb 98–114: A Novel Hemoglobin-Derived Antimicrobial Peptide from the Midgut of Rhipicephalus (Boophilus) microplus. Peptides 2012, 37, 120–127. [Google Scholar] [CrossRef]
- Rodríguez-Martín, A.; Acosta, R.; Liddell, S.; Núñez, F.; Benito, M.J.; Asensio, M.A. Characterization of the Novel Antifungal Protein PgAFP and the Encoding Gene of Penicillium chrysogenum. Peptides 2010, 31, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Sherman, P.; Luo, L.; Bowie, J.; Zhu, S. Structural and Functional Characterization of Two Genetically Related Meucin Peptides Highlights Evolutionary Divergence and Convergence in Antimicrobial Peptides. FASEB J. 2009, 23, 1230–1245. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.; Kullberg, B.J.; Tripet, B.; Boerman, O.C.; Zeeuwen, P.; van der Ven-Jongekrijg, J.; Verweij, P.; Schalkwijk, J.; Hodges, R.; van der Meer, J.W.M.; et al. Drosomycin-Like Defensin, a Human Homologue of Drosophila Melanogaster Drosomycin with Antifungal Activity. Antimicrob. Agents Chemother. 2008, 52, 1407–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabras, T.; Longhi, R.; Secundo, F.; Nocca, G.; Conti, S.; Polonelli, L.; Fanali, C.; Inzitari, R.; Petruzzelli, R.; Messana, I.; et al. Structural and Functional Characterization of the Porcine Proline–Rich Antifungal Peptide SP-B Isolated from Salivary Gland Granules. J. Pept. Sci. 2008, 14, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Briolat, J.; Wu, S.D.; Mahata, S.K.; Gonthier, B.; Bagnard, D.; Chasserot-Golaz, S.; Helle, K.B.; Aunis, D.; Metz-Boutigue, M.H. New Antimicrobial Activity for the Catecholamine Release-Inhibitory Peptide from Chromogranin A. CMLS Cell. Mol. Life Sci. 2005, 62, 377–385. [Google Scholar] [CrossRef]
- Landon, C.; Barbaut, F.; Legrain, M.; Menin, L.; Guenneugues, M.; Schott, V.; Vovelle, F. Lead Optimization of Antifungal Peptides with 3D NMR Structures Analysis. Protein Sci. 2004, 13, 703–713. [Google Scholar] [CrossRef] [Green Version]
- Kaiserer, L.; Oberparleiter, C.; Weiler-Görz, R.; Burgstaller, W.; Leiter, E.; Marx, F. Characterization of the Penicillium chrysogenum Antifungal Protein PAF. Arch. Microbiol. 2003, 180, 204–210. [Google Scholar] [CrossRef]
- Lauth, X.; Shike, H.; Burns, J.C.; Westerman, M.E.; Ostland, V.E.; Carlberg, J.M.; Van Olst, J.C.; Nizet, V.; Taylor, S.W.; Shimizu, C.; et al. Discovery and Characterization of Two Isoforms of Moronecidin, a Novel Antimicrobial Peptide from Hybrid Striped Bass. J. Biol. Chem. 2002, 277, 5030–5039. [Google Scholar] [CrossRef] [Green Version]
- Silva, P.I.; Daffre, S.; Bulet, P. Isolation and Characterization of Gomesin, an 18-Residue Cysteine-Rich Defense Peptide from the Spider Acanthoscurria Gomesiana Hemocytes with Sequence Similarities to Horseshoe Crab Antimicrobial Peptides of the Tachyplesin Family. J. Biol. Chem. 2000, 275, 33464–33470. [Google Scholar] [CrossRef] [Green Version]
- Lugardon, K.; Raffner, R.; Goumon, Y.; Corti, A.; Delmas, A.; Bulet, P.; Aunis, D.; Metz-Boutigue, M.-H. Antibacterial and Antifungal Activities of Vasostatin-1, the N-Terminal Fragment of Chromogranin A. J. Biol. Chem. 2000, 275, 10745–10753. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.G.; Shin, S.Y.; Maeng, C.-Y.; Jin, Z.Z.; Kim, K.L.; Hahm, K.-S. Isolation and Characterization of a Novel Antifungal Peptide from Aspergillus Niger. Biochem. Biophys. Res. Commun. 1999, 263, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Gallo, R.L.; Kim, K.J.; Bernfield, M.; Kozak, C.A.; Zanetti, M.; Merluzzi, L.; Gennaro, R. Identification of CRAMP, a Cathelin-Related Antimicrobial Peptide Expressed in the Embryonic and Adult Mouse. J. Biol. Chem. 1997, 272, 13088–13093. [Google Scholar] [CrossRef] [PubMed]
- Lawyer, C.; Pai, S.; Watabe, M.; Borgia, P.; Mashimo, T.; Eagleton, L.; Watabe, K. Antimicrobial Activity of a 13 Amino Acid Tryptophan-Rich Peptide Derived from a Putative Porcine Precursor Protein of a Novel Family of Antibacterial Peptides. FEBS Lett. 1996, 390, 95–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehret-Sabatier, L.; Loew, D.; Goyffon, M.; Fehlbaum, P.; Hoffmann, J.A.; van Dorsselaer, A.; Bulet, P. Characterization of Novel Cysteine-Rich Antimicrobial Peptides from Scorpion Blood. J. Biol. Chem. 1996, 271, 29537–29544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mor, A.; Nicolas, P. Isolation and Structure of Novel Defensive Peptides from Frog Skin. Eur. J. Biochem. 1994, 219, 145–154. [Google Scholar] [CrossRef]
- Mor, A.; Hani, K.; Nicolas, P. The Vertebrate Peptide Antibiotics Dermaseptins Have Overlapping Structural Features but Target Specific Microorganisms. J. Biol. Chem. 1994, 269, 31635–31641. [Google Scholar] [CrossRef]
- Mor, A.; Chartrel, N.; Vaudry, H.; Nicolas, P. Skin Peptide Tyrosine-Tyrosine, a Member of the Pancreatic Polypeptide Family: Isolation, Structure, Synthesis, and Endocrine Activity. Proc. Natl. Acad. Sci. USA 1994, 91, 10295–10299. [Google Scholar] [CrossRef] [Green Version]
- Fehlbaum, P.; Bulet, P.; Michaut, L.; Lagueux, M.; Broekaert, W.F.; Hetru, C.; Hoffmann, J.A. Insect Immunity. Septic Injury of Drosophila Induces the Synthesis of a Potent Antifungal Peptide with Sequence Homology to Plant Antifungal Peptides. J. Biol. Chem. 1994, 269, 33159–33163. [Google Scholar] [CrossRef]
- Bellamy, W.; Takase, M.; Wakabayashi, H.; Kawase, K.; Tomita, M. Antibacterial Spectrum of Lactoferricin B, a Potent Bactericidal Peptide Derived from the N -Terminal Region of Bovine Lactoferrin. J. Appl. Bacteriol. 1992, 73, 472–479. [Google Scholar] [CrossRef]
- Mor, A.; Van Huong, N.; Delfour, A.; Migliore-Samour, D.; Nicolas, P. Isolation, Amino Acid Sequence and Synthesis of Dermaseptin, a Novel Antimicrobial Peptide of Amphibian Skin. Biochemistry 1991, 30, 8824–8830. [Google Scholar] [CrossRef] [PubMed]
- Wnendt, S.; Ulbrich’, N.; Stahl, U. Cloning and Nucleotide Sequence of a CDNA Encoding the Antifungal-Protein of Aspergillus Giganteus and Preliminary Characterization of the Native Gene. Nucleic Acids Res. 1990, 18, 3987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, K.W.; Evans, R.J.; Eisenberg, S.P.; Thompson, R.C. Secretory Leukocyte Protease Inhibitor Binding to MRNA and DNA as a Possible Cause of Toxicity to Escherichia coli. J. Bacteriol. 1989, 171, 2166–2172. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. New approaches to antimicrobial discovery. Biochem. Pharmacol. 2017, 134, 87–98. [Google Scholar] [CrossRef]
- Marr, A.; Gooderham, W.; Hancock, R. Antibacterial Peptides for Therapeutic Use: Obstacles and Realistic Outlook. Curr. Opin. Pharmacol. 2006, 6, 468–472. [Google Scholar] [CrossRef]
- Souza, P.F.N.; Marques, L.S.M.; Oliveira, J.T.A.; Lima, P.G.; Dias, L.P.; Neto, N.A.S.; Lopes, F.E.S.; Sousa, J.S.; Silva, A.F.B.; Caneiro, R.F.; et al. Synthetic Antimicrobial Peptides: From Choice of the Best Sequences to Action Mechanisms. Biochimie 2020, 175, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Dias, L.P.; Souza, P.F.N.; Oliveira, J.T.A.; Vasconcelos, I.M.; Araújo, N.M.S.; Tilburg, M.F.V.; Guedes, M.I.F.; Carneiro, R.F.; Lopes, J.L.S.; Sousa, D.O.B. RcAlb-PepII, a Synthetic Small Peptide Bioinspired in the 2S Albumin from the Seed Cake of Ricinus communis, Is a Potent Antimicrobial Agent against Klebsiella pneumoniae and Candida parapsilosis. Biochim. Biophys. Acta (BBA)-Biomembr. 2020, 1862, 183092. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, T.; Kelly, B.; Dobson, C.; d’Enfert, C. Endocytosis-Mediated Vacuolar Accumulation of the Human ApoE Apolipoprotein-Derived ApoEdpL-W Antimicrobial Peptide Contributes to Its Antifungal Activity in Candida albicans. Antimicrob. Agents Chemother. 2011, 55, 4670–4681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurya, I.K.; Thota, C.K.; Sharma, J.; Tupe, S.G.; Chaudhary, P.; Singh, M.K.; Thakur, I.S.; Deshpande, M.; Prasad, R.; Chauhan, V.S. Mechanism of action of novel synthetic dodecapeptides against Candida albicans. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2013, 1830, 5193–5203. [Google Scholar] [CrossRef]
- Fioriti, S.; Cirioni, O.; Simonetti, O.; Franca, L.; Candelaresi, B.; Pallotta, F.; Neubauer, D.; Kamysz, E.; Kamysz, W.; Canovari, B.; et al. In Vitro Activity of Novel Lipopeptides against Triazole-Resistant Aspergillus Fumigatus. JoF 2022, 8, 872. [Google Scholar] [CrossRef]
- De Lucca, A.J.; Walsh, T.J. Antifungal Peptides: Novel Therapeutic Compounds against Emerging Pathogens. Antimicrob. Agents Chemother. 1999, 43, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Swidergall, M.; Ernst, J.F. Interplay between Candida Albicans and the Antimicrobial Peptide Armory. Eukaryot. Cell 2014, 13, 950–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sierra, J.M.; Fusté, E.; Rabanal, F.; Vinuesa, T.; Viñas, M. An Overview of Antimicrobial Peptides and the Latest Advances in Their Development. Expert Opin. Biol. Ther. 2017, 17, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Brogden, K.A. Antimicrobial Peptides: Pore Formers or Metabolic Inhibitors in Bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, K. Membrane Permeabilization Mechanisms. In Antimicrobial Peptides; Matsuzaki, K., Ed.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2019; Volume 1117, pp. 9–16. ISBN 9789811335877. [Google Scholar]
- Christoffersen, H.F.; Hansen, S.K.; Vad, B.S.; Nielsen, E.H.; Nielsen, J.T.; Vosegaard, T.; Skrydstrup, T.; Otzen, D.E. The Natural, Peptaibolic Peptide SPF-5506-A 4 Adopts a β-Bend Spiral Structure, Shows Low Hemolytic Activity and Targets Membranes through Formation of Large Pores. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2015, 1854, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Shai, Y. Mechanism of the Binding, Insertion and Destabilization of Phospholipid Bilayer Membranes by K-Helical Antimicrobial and Cell Non-Selective Membrane-Lytic Peptides. Biochim. Biophys. Acta 1999, 1462, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Katragkou, A.; Williams, M.; Sternberg, S.; Pantazatos, D.; Roilides, E.; Walsh, T.J. Micafungin Alters the Amino Acid, Nucleic Acid and Central Carbon Metabolism of Candida Albicans at Subinhibitory Concentrations: Novel Insights into Mechanisms of Action. J. Antimicrob. Chemother. 2017, 72, 712–716. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-R.; Raman, N.; Ortiz-Bermúdez, P.; Lynn, D.M.; Palecek, S.P. 14-Helical β-Peptides Elicit Toxicity against C. Albicans by Forming Pores in the Cell Membrane and Subsequently Disrupting Intracellular Organelles. Cell Chem. Biol. 2019, 26, 289–299. [Google Scholar] [CrossRef]
- Sharma, S.; Sahoo, N.; Bhunia, A. Antimicrobial Peptides and Their Pore/Ion Channel Properties in Neutralization of Pathogenic Microbes. CTMC 2015, 16, 46–53. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Dou, X.; Song, J.; Lyu, Y.; Zhu, X.; Xu, L.; Li, W.; Shan, A. Antimicrobial Peptides: Promising Alternatives in the Post Feeding Antibiotic Era. Med. Res. Rev. 2019, 39, 831–859. [Google Scholar] [CrossRef]
- Haney, E.F.; Mansour, S.C.; Hancock, R.E.W. Antimicrobial Peptides: An Introduction. In Antimicrobial Peptides; Hansen, P.R., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2017; Volume 1548, pp. 3–22. ISBN 978-1-4939-6735-3. [Google Scholar]
- Buda De Cesare, G.; Cristy, S.A.; Garsin, D.A.; Lorenz, M.C. Antimicrobial Peptides: A New Frontier in Antifungal Therapy. mBio 2020, 11, e02123-20. [Google Scholar] [CrossRef] [PubMed]
- Debono, M.; Gordee, R.S. Antibiotics that inhibit fungal cell wall development. Annu. Rev. Microbiol. 1994, 48, 471–497. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, L.T.; Manavathu, E.K.; Cutright, J.L.; Alangaden, G.J.; Chandrasekar, P.H. In-Vitro Activity of Nikkomycin Z Alone and in Combination with Polyenes, Triazoles or Echinocandins against Aspergillus fumigatus. Clin. Microbiol. Infect. 2004, 10, 961–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, I.; Perkins, W.R.; Lupan, D.M.; Selsted, M.E.; Janoff, A.S. Liposomal Entrapment of the Neutrophil-Derived Peptide Indolicidin Endows It with in Vivo Antifungal Activity. Biochim. Biophys. Acta (BBA)-Biomembr. 1995, 1237, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.G.; Kim, H.K.; Kim, S.A.; Park, Y.; Park, S.-C.; Jang, S.-H.; Hahm, K.-S. Fungicidal Effect of Indolicidin and Its Interaction with Phospholipid Membranes. Biochem. Biophys. Res. Commun. 2003, 305, 305–310. [Google Scholar] [CrossRef]
- Peng, C.; Liu, Y.; Shui, L.; Zhao, Z.; Mao, X.; Liu, Z. Mechanisms of Action of the Antimicrobial Peptide Cecropin in the Killing of Candida Albicans. Life 2022, 12, 1581. [Google Scholar] [CrossRef]
- Kumar, P.; Kizhakkedathu, J.; Straus, S. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility In Vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Rautenbach, M.; Troskie, A.M.; Vosloo, J.A. Antifungal Peptides: To Be or Not to Be Membrane Active. Biochimie 2016, 130, 132–145. [Google Scholar] [CrossRef]
- Kyriakidis, I.; Tragiannidis, A.; Munchen, S.; Groll, A.H. Clinical Hepatotoxicity Associated with Antifungal Agents. Expert Opin. Drug Saf. 2017, 16, 149–165. [Google Scholar] [CrossRef]
- Fernández, D.U.M.; Arbulu, S.; Garcia-Gutierrez, E.; Cotter, P.D. Antifungal Peptides as Therapeutic Agents. Front. Cell. Infect. Microbiol. 2020, 10, 105. [Google Scholar] [CrossRef]
- Zhao, X.; Zhou, Z.; Han, Y.; Wang, Z.; Fan, J.; Xiao, H. Isolation and Identification of Antifungal Peptides from Bacillus BH072, a Novel Bacterium Isolated from Honey. Microbiol. Res. 2013, 168, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.N.; Yan, X.; Gao, X.N.; Niu, H.J.; Kang, Z.S.; Huang, L.L. Purification and Characterization of a Potential Antifungal Protein from Bacillus subtilis E1R-J against Valsa mali. World J. Microbiol. Biotechnol. 2016, 32, 63. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Wang, Q.; Wang, K.; Brian, K.; Liu, C.; Gu, Y. Study of the Antifungal Activity of Bacillus vallismortis ZZ185 in Vitro and Identification of Its Antifungal Components. Bioresour. Technol. 2010, 101, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.T.; Wencewicz, T.A. Prospects for New Antibiotics: A Molecule-Centered Perspective. J. Antibiot. 2014, 67, 7–22. [Google Scholar] [CrossRef]
- Yan, J.; Bhadra, P.; Li, A.; Sethiya, P.; Qin, L.; Tai, H.K.; Wong, K.H.; Siu, S.W.I. Deep-AmPEP30: Improve Short Antimicrobial Peptides Prediction with Deep Learning. Mol. Ther.-Nucleic Acids 2020, 20, 882–894. [Google Scholar] [CrossRef] [PubMed]
- Waghu, F.H.; Gopi, L.; Barai, R.S.; Ramteke, P.; Nizami, B.; Idicula-Thomas, S. CAMP: Collection of Sequences and Structures of Antimicrobial Peptides. Nucl. Acids Res. 2014, 42, D1154–D1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burdukiewicz, M.; Sidorczuk, K.; Rafacz, D.; Pietluch, F.; Chilimoniuk, J.; Rödiger, S.; Gagat, P. Proteomic Screening for Prediction and Design of Antimicrobial Peptides with AmpGram. Int. J. Mol. Sci. 2020, 21, 4310. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Wang, P.; Lin, W.-Z.; Jia, J.-H.; Chou, K.-C. IAMP-2L: A Two-Level Multi-Label Classifier for Identifying Antimicrobial Peptides and Their Functional Types. Anal. Biochem. 2013, 436, 168–177. [Google Scholar] [CrossRef]
- Lee, E.Y.; Lee, M.W.; Fulan, B.M.; Ferguson, A.L.; Wong, G.C.L. What Can Machine Learning Do for Antimicrobial Peptides, and What Can Antimicrobial Peptides Do for Machine Learning? Interface Focus. 2017, 7, 20160153. [Google Scholar] [CrossRef]
- Müller, A.T.; Hiss, J.A.; Schneider, G. Recurrent Neural Network Model for Constructive Peptide Design. J. Chem. Inf. Model. 2018, 58, 472–479. [Google Scholar] [CrossRef]
- Vélez, A.; Mera, C.; Orduz, S.; Branch, J.W. Synthetic antimicrobial peptides generation using recurrent neural networks. Dyna 2021, 10, 210–219. [Google Scholar]
- Capecchi, A.; Cai, X.; Personne, H.; Köhler, T.; van Delden, C.; Reymond, J.-L. Machine Learning Designs Non-Hemolytic Antimicrobial Peptides. Chem. Sci. 2021, 12, 9221–9232. [Google Scholar] [CrossRef] [PubMed]
- Otovic, E.; Njirjak, M.; Kalafatovic, D.; Mauša, G. Sequential Properties Representation Scheme for Recurrent Neural Network Based Prediction of Therapeutic Peptides Supplementary Information. J. Chem. Inf. Model. 2022, 62, 2961–2972. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, L.; Tian, Z.; Zhao, W.; Sun, C.; Zhu, L.; Huang, M.; Guo, G.; Liang, G. Large-Scale Screening of Antifungal Peptides Based on Quantitative Structure–Activity Relationship. ACS Med. Chem. Lett. 2022, 13, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Shrivastava, S.; Kumar Singh, S.; Kumar, A.; Saxena, S. Accelerating the Discovery of Antifungal Peptides Using Deep Temporal Convolutional Networks. Brief. Bioinform. 2022, 23, bbac008. [Google Scholar] [CrossRef]
- Kovalainen, M.; Mönkäre, J.; Riikonen, J.; Pesonen, U.; Vlasova, M.; Salonen, J.; Lehto, V.-P.; Järvinen, K.; Herzig, K.-H. Novel Delivery Systems for Improving the Clinical Use of Peptides. Pharm. Rev. 2015, 67, 541–561. [Google Scholar] [CrossRef] [Green Version]
- Jao, D.; Xue, Y.; Medina, J.; Hu, X. Protein-Based Drug-Delivery Materials. Materials 2017, 10, 517. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.Y.; Yuk, S.H. Polymeric Protein Delivery Systems. Prog. Polym. Sci. 2007, 32, 669–697. [Google Scholar] [CrossRef]
- Cassone, A. Fungal Vaccines: Real Progress from Real Challenges. Lancet Infect. Dis. 2008, 8, 114–124. [Google Scholar] [CrossRef]
- Filler, S.G.; Sheppard, D.C. Fungal Invasion of Normally Non-Phagocytic Host Cells. PLoS Pathog. 2006, 2, e129. [Google Scholar] [CrossRef]
- Latgé, J.-P.; Chamilos, G. Aspergillus fumigatus and Aspergillosis in 2019. Clin. Microbiol. Rev. 2019, 33, e00140-18. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; Pirofski, L. Polysaccharide-Containing Conjugate Vaccines for Fungal Diseases. Trends Mol. Med. 2006, 12, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Feldmesser, M. Prospects of Vaccines for Medically Important Fungi. Med. Mycol. 2005, 43, 571–587. [Google Scholar] [CrossRef] [PubMed]
- Torosantucci, A.; Bromuro, C.; Chiani, P.; De Bernardis, F.; Berti, F.; Galli, C.; Norelli, F.; Bellucci, C.; Polonelli, L.; Costantino, P.; et al. A Novel Glyco-Conjugate Vaccine against Fungal Pathogens. J. Exp. Med. 2005, 202, 597–606. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.; Hua, Y.-H.; Zhang, Y.-D.; Bao, D.; Lv, J.; Hu, H.-F. The Pathogenesis of Aspergillus fumigatus, Host Defense Mechanisms, and the Development of AFMP4 Antigen as a Vaccine. Pol. J. Microbiol. 2021, 70, 3–11. [Google Scholar] [CrossRef]
- Ito, J.I.; Lyons, J.M.; Hong, T.B.; Tamae, D.; Liu, Y.-K.; Wilczynski, S.P.; Kalkum, M. Vaccinations with Recombinant Variants of Aspergillus fumigatus Allergen Asp f 3 Protect Mice against Invasive Aspergillosis. Infect. Immun. 2006, 74, 5075–5084. [Google Scholar] [CrossRef] [Green Version]
- Bozza, S.; Perruccio, K.; Montagnoli, C.; Gaziano, R.; Bellocchio, S.; Burchielli, E.; Nkwanyuo, G.; Pitzurra, L.; Velardi, A.; Romani, L. A Dendritic Cell Vaccine against Invasive Aspergillosis in Allogeneic Hematopoietic Transplantation. Blood 2003, 102, 3807–3814. [Google Scholar] [CrossRef]
- Stevens, D.A. Vaccinate Against Aspergillosis! A Call to Arms of the Immune System. Clin. Infect. Dis. 2004, 38, 1131–1136. [Google Scholar] [CrossRef] [Green Version]
- Mestas, J.; Hughes, C.C.W. Of Mice and Not Men: Differences between Mouse and Human Immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef] [Green Version]
- Portuondo, D.L.F.; Ferreira, L.S.; Urbaczek, A.C.; Batista-Duharte, A.; Carlos, I.Z. Adjuvants and Delivery Systems for Antifungal Vaccines: Current State and Future Developments. Med. Mycol. 2015, 53, 69–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, L.V.N.; Wang, R.; Specht, C.A.; Levitz, S.M. Vaccines for Human Fungal Diseases: Close but Still a Long Way to Go. npj Vaccines 2021, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Szalewski, D.A.; Hinrichs, V.S.; Zinniel, D.K.; Barletta, R.G. The Pathogenicity of Aspergillus Fumigatus, Drug Resistance, and Nanoparticle Delivery. Can. J. Microbiol. 2018, 64, 439–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soliman, G.M. Nanoparticles as Safe and Effective Delivery Systems of Antifungal Agents: Achievements and Challenges. Int. J. Pharm. 2017, 523, 15–32. [Google Scholar] [CrossRef]
- Sahoo, S.K.; Parveen, S.; Panda, J.J. The Present and Future of Nanotechnology in Human Health Care. Nanomed. Nanotechnol. Biol. Med. 2007, 3, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Skanda, S.; Bharadwaj, P.S.J.; Datta Darshan, V.M.; Sivaramakrishnan, V.; Vijayakumar, B.S. Proficient Mycogenic Synthesis of Silver Nanoparticles by Soil Derived Fungus Aspergillus Melleus SSS-10 with Cytotoxic and Antibacterial Potency. J. Microbiol. Methods 2022, 199, 106517. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Jha, S.K.; Hasan, N.; Insaf, A.; Shrestha, J.; Shrestha, J.; Devkota, H.P.; Khan, S.; Panth, N.; Warkiani, M.E.; et al. Overcoming Multidrug Resistance of Antibiotics via Nanodelivery Systems. Pharmaceutics 2022, 14, 586. [Google Scholar] [CrossRef]
- Aguilar-Pérez, K.M.; Medina, D.I.; Parra-Saldívar, R.; Iqbal, H.M.N. Nano-Size Characterization and Antifungal Evaluation of Essential Oil Molecules-Loaded Nanoliposomes. Molecules 2022, 27, 5728. [Google Scholar] [CrossRef]
- Hassanpour, P.; Hamishehkar, H.; Bahari Baroughi, B.; Baradaran, B.; Sandoghchian Shotorbani, S.; Mohammadi, M.; Shomali, N.; Aghebati-Maleki, L.; Nami, S. Antifungal Effects of Voriconazole-Loaded Nano-Liposome on Fluconazole-Resistant Clinical Isolates of Candida Albicans, Biological Activity and ERG11, CDR1, and CDR2 Gene Expression. ASSAY Drug Dev. Technol. 2021, 19, 453–462. [Google Scholar] [CrossRef]
- Jafari, A.; Daneshamouz, S.; Ghasemiyeh, P.; Mohammadi-Samani, S. Ethosomes as Dermal/Transdermal Drug Delivery Systems: Applications, Preparation and Characterization. J. Liposome Res. 2022, 1–19. [Google Scholar] [CrossRef]
- Sguizzato, M.; Ferrara, F.; Hallan, S.S.; Baldisserotto, A.; Drechsler, M.; Malatesta, M.; Costanzo, M.; Cortesi, R.; Puglia, C.; Valacchi, G.; et al. Ethosomes and Transethosomes for Mangiferin Transdermal Delivery. Antioxidants 2021, 10, 768. [Google Scholar] [CrossRef]
- Aparajay, P.; Dev, A. Functionalized Niosomes as a Smart Delivery Device in Cancer and Fungal Infection. Eur. J. Pharm. Sci. 2022, 168, 106052. [Google Scholar] [CrossRef] [PubMed]
- Almansob, A.; Bahkali, A.H.; Ameen, F. Efficacy of Gold Nanoparticles against Drug-Resistant Nosocomial Fungal Pathogens and Their Extracellular Enzymes: Resistance Profiling towards Established Antifungal Agents. Nanomaterials 2022, 12, 814. [Google Scholar] [CrossRef] [PubMed]
- Majumder, N.; G Das, N.; Das, S.K. Polymeric Micelles for Anticancer Drug Delivery. Ther. Deliv. 2020, 11, 613–635. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Gulati, N.; Makhija, M.; Purohit, D.; Dureja, H. Nanoemulsion: A Novel Drug Delivery Approach for Enhancement of Bioavailability. Recent Pat. Nanotechnol. 2020, 14, 276–293. [Google Scholar] [CrossRef]
- Yu, K.-P.; Huang, Y.-T.; Yang, S.-C. The Antifungal Efficacy of Nano-Metals Supported TiO2 and Ozone on the Resistant Aspergillus Niger Spore. J. Hazard. Mater. 2013, 261, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-F.; Liu, Z.-G.; Shen, W.; Gurunathan, S. Silver Nanoparticles: Synthesis, Characterization, Properties, Applications, and Therapeutic Approaches. IJMS 2016, 17, 1534. [Google Scholar] [CrossRef]
- Palacios-Hernandez, T.; Diaz-Diestra, D.M.; Nguyen, A.K.; Skoog, S.A.; Chikkaveeraiah, B.V.; Tang, X.; Wu, Y.; Petrochenko, P.E.; Sussman, E.M.; Goering, P.L. Cytotoxicity, Cellular Uptake and Apoptotic Responses in Human Coronary Artery Endothelial Cells Exposed to Ultrasmall Superparamagnetic Iron Oxide Nanoparticles. J. Appl. Toxicol. 2020, 40, 918–930. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, Z.W.; Allaker, R.P.; Reip, P.; Oxford, J.; Ahmad, Z.; Ren, G. A Review of Nanoparticle Functionality and Toxicity on the Central Nervous System. J. R. Soc. Interface 2010, 7, S411–S422. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pimienta, D.A.; Cruz Mosquera, F.E.; Palacios Velasco, I.; Giraldo Rodas, M.; Oñate-Garzón, J.; Liscano, Y. Specific Focus on Antifungal Peptides against Azole Resistant Aspergillus fumigatus: Current Status, Challenges, and Future Perspectives. J. Fungi 2023, 9, 42. https://doi.org/10.3390/jof9010042
Pimienta DA, Cruz Mosquera FE, Palacios Velasco I, Giraldo Rodas M, Oñate-Garzón J, Liscano Y. Specific Focus on Antifungal Peptides against Azole Resistant Aspergillus fumigatus: Current Status, Challenges, and Future Perspectives. Journal of Fungi. 2023; 9(1):42. https://doi.org/10.3390/jof9010042
Chicago/Turabian StylePimienta, Dilan Andrés, Freiser Eceomo Cruz Mosquera, Isabella Palacios Velasco, María Giraldo Rodas, Jose Oñate-Garzón, and Yamil Liscano. 2023. "Specific Focus on Antifungal Peptides against Azole Resistant Aspergillus fumigatus: Current Status, Challenges, and Future Perspectives" Journal of Fungi 9, no. 1: 42. https://doi.org/10.3390/jof9010042