
What’s New in Cryptococcus gattii: From Bench to Bedside and Beyond
 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Latest on Taxonomy and Molecular Epidemiology of Cryptococcus

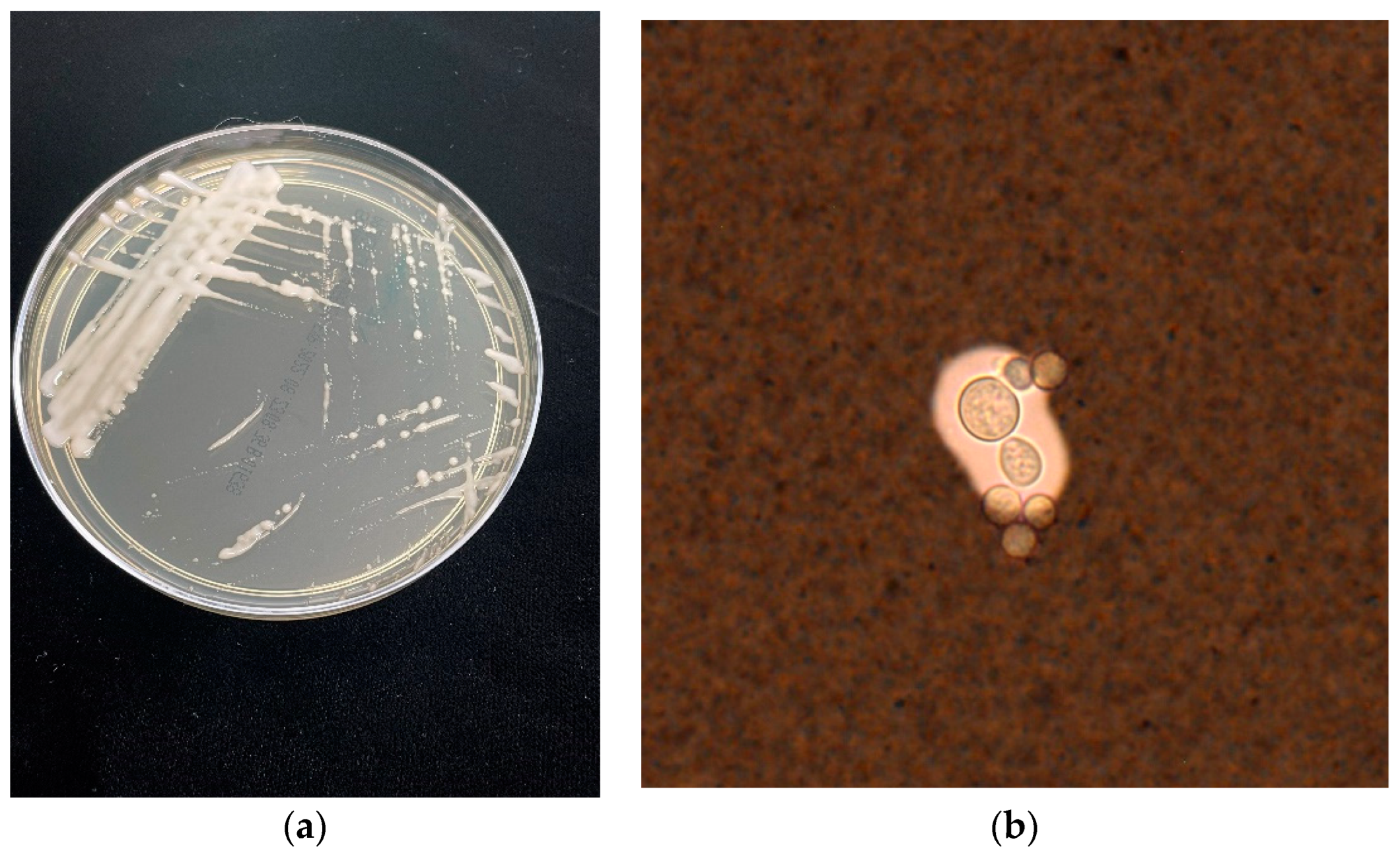{kind=link}
{kind=link}
| Species Complex | Proposed Species Name | Variety | Serotype | PCR Fingerprinting | Reference Strain—Accession ID of Reference Assembly |
|---|---|---|---|---|---|
| Cryptococcus neoformans species complex | C. neoformans | var. grubii | A | VNI | H99-ASM301198v1 |
| VNB | Bt88-BROAD_CneoA_Bt88_1 | ||||
| VNII | PMHc1023.ENR BROAD_CneoA_PMHc1023.Enr_1 | ||||
| C. deneoformans | var. neoformans | D | VNIV | JEC21-ASM9104v1 | |
| C. neoformans × C. deneoformans hybrid | AD hybrid | AD | VNIII | ||
| Cryptocccus gattii species complex | C. gattii | B/C | VGI | WM276-ASM18594v1 Ru294-Cryp_gatt_Ru294_V1 | |
| C. deuterogattii | B/C | VGII, VGIIa, b, c, d | R265-R265.1 | ||
| C. bacillisporus | B/C | VGIII | CA1280-Cryp_gatt_CA1280_V1 | ||
| B | CA1873-Cryp_gatt_CA1873_V1 | ||||
| C. tetragattii | B/C | VGIV | IND107-Cryp_gatt_IND107_V2 | ||
| B | MF34-Cryp_gatt_MF34 | ||||
| - | B/C | VGV | |||
| C. decagattii | B/C | VGVI |
3. Ecology, Epidemiology and Clinical Features
3.1. Ecology
3.2. Epidemiology
3.3. Clinical Features and Diagnosis
4. Advances in Understanding of Virulence and Pathogenesis
5. Novel Therapeutics for C. gattii
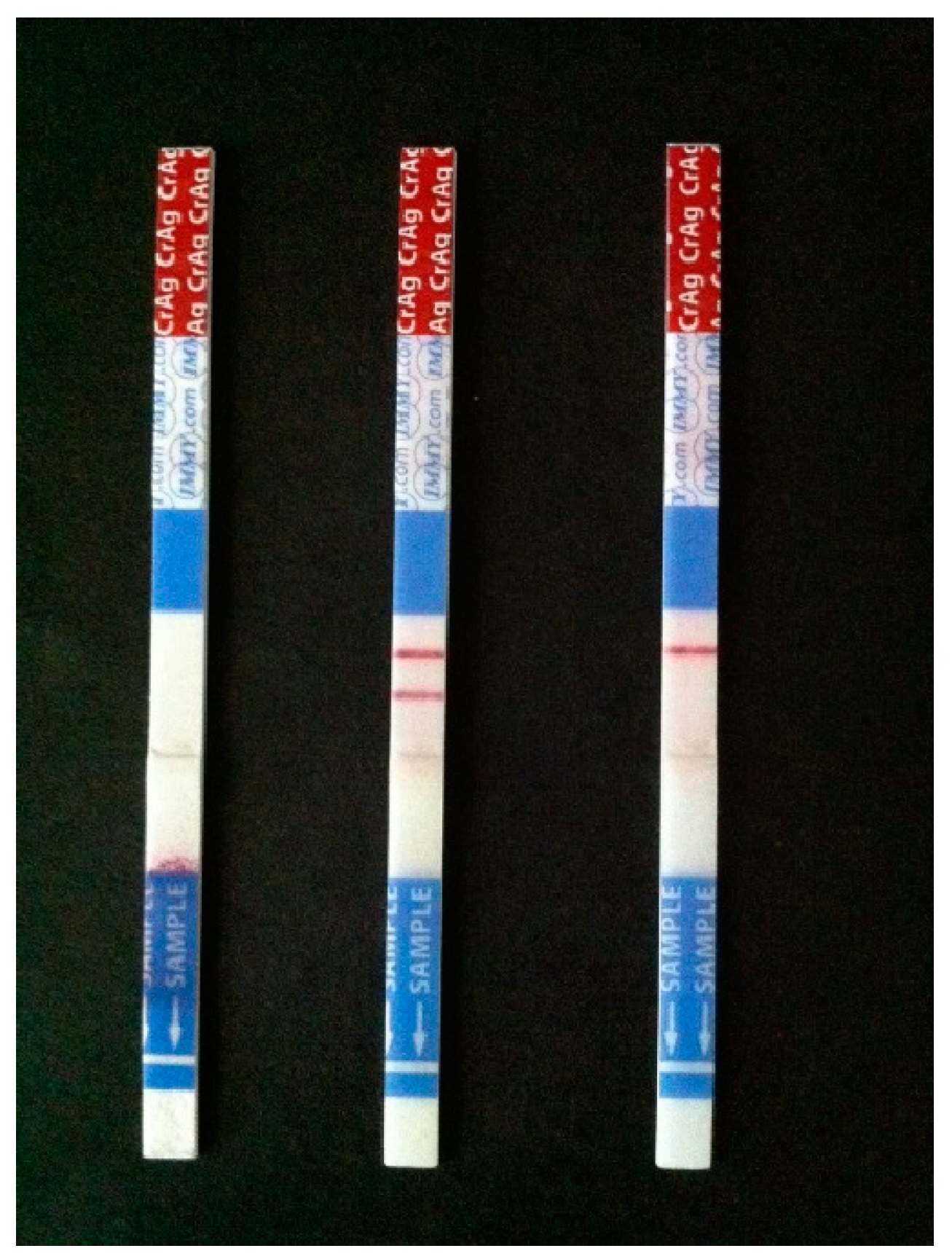
6. Updates in Diagnostics
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hurtado, J.C.; Castillo, P.; Fernandes, F.; Navarro, M.; Lovane, L.; Casas, I.; Quintó, L.; Marco, F.; Jordao, D.; Ismail, M.R.; et al. Mortality due to Cryptococcus neoformans and Cryptococcus gattii in low-income settings: An autopsy study. Sci. Rep. 2019, 9, 7493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.A.; Arthur, I.; Merritt, A.; Leung, M. Molecular types of Cryptococcus neoformans and Cryptococcus gattii in Western Australia and correlation with antifungal susceptibility. Med. Mycol. 2019, 57, 1004–1010. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, S.; Manjunath, N.; Bhat, M.; Hagen, F.; Bahubali, V.H.; Palaniappan, M.; Maji, S.; Chandrashekar, N. Clinical insights and epidemiology of central nervous system infection due to Cryptococcus neoformans/gattii species complexes: A prospective study from South India. Med. Mycol. 2020, 58, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.M.; Mba-Jonas, A.; Tourdjman, M.; Schimek, T.; DeBess, E.; Marsden-Haug, N.; Harris, J.R. Treatment and outcomes among patients with Cryptococcus gattii infections in the United States Pacific Northwest. PLoS ONE 2014, 9, e88875. [Google Scholar] [CrossRef]
- Chen, S.C.; Korman, T.M.; Slavin, M.A.; Marriott, D.; Byth, K.; Bak, N.; Currie, B.J.; Hajkowicz, K.; Heath, C.H.; Kidd, S.; et al. Antifungal therapy and management of complications of cryptococcosis due to Cryptococcus gattii. Clin. Infect. Dis. 2013, 57, 543–551. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.C.; Slavin, M.A.; Heath, C.H.; Playford, E.G.; Byth, K.; Marriott, D.; Kidd, S.E.; Bak, N.; Currie, B.; Hajkowicz, K.; et al. Clinical manifestations of Cryptococcus gattii infection: Determinants of neurological sequelae and death. Clin. Infect. Dis. 2012, 55, 789–798. [Google Scholar] [CrossRef] [Green Version]
- Phillips, P.; Galanis, E.; MacDougall, L.; Chong, M.Y.; Balshaw, R.; Cook, V.J.; Bowie, W.; Steiner, T.; Hoang, L.; Morshed, M.; et al. Longitudinal clinical findings and outcome among patients with Cryptococcus gattii infection in British Columbia. Clin. Infect. Dis. 2015, 60, 1368–1376. [Google Scholar] [CrossRef] [Green Version]
- WHO. Fungal Priority Pathogens List to Guide Research, Development and Public Health Action; World Health Organization: Geneva, Switzerland, 2022; pp. 1–48. [Google Scholar]
- Chen, S.C.; Meyer, W.; Sorrell, T.C. Cryptococcus gattii infections. Clin. Microbiol. Rev. 2014, 27, 980–1024. [Google Scholar] [CrossRef] [Green Version]
- Maziarz, E.K.; Perfect, J.R. Cryptococcosis. Infect. Dis. Clin. N. Am. 2016, 30, 179–206. [Google Scholar] [CrossRef] [Green Version]
- Firacative, C.; Roe, C.C.; Malik, R.; Ferreira-Paim, K.; Escandon, P.; Sykes, J.E.; Castanon-Olivares, L.R.; Contreras-Peres, C.; Samayoa, B.; Sorrell, T.C.; et al. MLST and Whole-Genome-Based Population Analysis of Cryptococcus gattii VGIII Links Clinical, Veterinary and Environmental Strains, and Reveals Divergent Serotype Specific Sub-populations and Distant Ancestors. PLoS Negl. Trop. Dis. 2016, 10, e0004861. [Google Scholar] [CrossRef]
- Kwon-Chung, K.J.; Bennett, J.E.; Wickes, B.L.; Meyer, W.; Cuomo, C.A.; Wollenburg, K.R.; Bicanic, T.A.; Castañeda, E.; Chang, Y.C.; Chen, J.; et al. The Case for Adopting the “Species Complex” Nomenclature for the Etiologic Agents of Cryptococcosis. mSphere 2017, 2, e00357-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firacative, C.; Trilles, L.; Meyer, W. Recent Advances in Cryptococcus and Cryptococcosis. Microorganisms 2021, 10, 13. [Google Scholar] [CrossRef] [PubMed]
- Ito-Kuwa, S.; Nakamura, K.; Aoki, S.; Vidotto, V. Serotype identification of Cryptococcus neoformans by multiplex PCR. Mycoses 2007, 50, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Rathore, S.S.; Sathiyamoorthy, J.; Lalitha, C.; Ramakrishnan, J. A holistic review on Cryptococcus neoformans. Microb. Pathog. 2022, 166, 105521. [Google Scholar] [CrossRef] [PubMed]
- Bive, B.Z.; Sacheli, R.; Situakibanza Nani-Tuma, H.; Kabututu Zakayi, P.; Ka, A.; Mbula Mambimbi, M.; Muendele, G.; Boreux, R.; Landu, N.; Nzanzu Mudogo, C.; et al. Clinical epidemiology and high genetic diversity amongst Cryptococcus spp. isolates infecting people living with HIV in Kinshasa, Democratic Republic of Congo. PLoS ONE 2022, 17, e0267842. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, M.M.; Lazera, M.S.; Barbosa, G.G.; Trilles, L.; Balassiano, B.R.; Macedo, R.C.; Bezerra, C.C.; Pérez, M.A.; Cardarelli, P.; Wanke, B. Serotyping of 467 Cryptococcus neoformans isolates from clinical and environmental sources in Brazil: Analysis of host and regional patterns. J. Clin. Microbiol. 2003, 41, 73–77. [Google Scholar] [CrossRef] [Green Version]
- Pfeiffer, T.J.; Ellis, D.H. Serotypes of Australian environmental and clinical isolates of Cryptococcus neoformans. J. Med. Vet. Mycol. 1993, 31, 401–404. [Google Scholar] [CrossRef]
- Desjardins, C.A.; Giamberardino, C.; Sykes, S.M.; Yu, C.H.; Tenor, J.L.; Chen, Y.; Yang, T.; Jones, A.M.; Sun, S.; Haverkamp, M.R.; et al. Population genomics and the evolution of virulence in the fungal pathogen Cryptococcus neoformans. Genome Res. 2017, 27, 1207–1219. [Google Scholar] [CrossRef] [Green Version]
- Hagen, F.; Khayhan, K.; Theelen, B.; Kolecka, A.; Polacheck, I.; Sionov, E.; Falk, R.; Parnmen, S.; Lumbsch, H.T.; Boekhout, T. Recognition of seven species in the Cryptococcus gattii/Cryptococcus neoformans species complex. Fungal Genet. Biol. 2015, 78, 16–48. [Google Scholar] [CrossRef] [Green Version]
- Farrer, R.A.; Chang, M.; Davis, M.J.; van Dorp, L.; Yang, D.H.; Shea, T.; Sewell, T.R.; Meyer, W.; Balloux, F.; Edwards, H.M.; et al. A New Lineage of Cryptococcus gattii (VGV) Discovered in the Central Zambezian Miombo Woodlands. mBio 2019, 10, e02306-19. [Google Scholar] [CrossRef]
- Meyer, W.; Aanensen, D.M.; Boekhout, T.; Cogliati, M.; Diaz, M.R.; Esposto, M.C.; Fisher, M.; Gilgado, F.; Hagen, F.; Kaocharoen, S.; et al. Consensus multi-locus sequence typing scheme for Cryptococcus neoformans and Cryptococcus gattii. Med. Mycol. 2009, 47, 561–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagen, F.; Lumbsch, H.T.; Arsic Arsenijevic, V.; Badali, H.; Bertout, S.; Billmyre, R.B.; Bragulat, M.R.; Cabañes, F.J.; Carbia, M.; Chakrabarti, A.; et al. Importance of Resolving Fungal Nomenclature: The Case of Multiple Pathogenic Species in the Cryptococcus Genus. mSphere 2017, 2, e00238-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McTaggart, L.R.; Lei, E.; Richardson, S.E.; Hoang, L.; Fothergill, A.; Zhang, S.X. Rapid identification of Cryptococcus neoformans and Cryptococcus gattii by matrix-assisted laser desorption ionization-time of flight mass spectrometry. J. Clin. Microbiol. 2011, 49, 3050–3053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, M.; Brown, Z.; Houldsworth, M.; Borman, A.M.; Johnson, E.M. Rapid identification of 6328 isolates of pathogenic yeasts using MALDI-ToF MS and a simplified, rapid extraction procedure that is compatible with the Bruker Biotyper platform and database. Med. Mycol. 2016, 54, 80–88. [Google Scholar] [CrossRef] [Green Version]
- Posteraro, B.; Vella, A.; Cogliati, M.; De Carolis, E.; Florio, A.R.; Posteraro, P.; Sanguinetti, M.; Tortorano, A.M. Matrix-assisted laser desorption ionization-time of flight mass spectrometry-based method for discrimination between molecular types of Cryptococcus neoformans and Cryptococcus gattii. J. Clin. Microbiol. 2012, 50, 2472–2476. [Google Scholar] [CrossRef] [Green Version]
- Siqueira, L.P.M.; Gimenes, V.M.F.; de Freitas, R.S.; Melhem, M.S.C.; Bonfietti, L.X.; da Silva, A.R., Jr.; Souza Santos, L.B.; Motta, A.L.; Rossi, F.; Benard, G.; et al. Evaluation of Vitek MS for Differentiation of Cryptococcus neoformans and Cryptococcus gattii Genotypes. J. Clin. Microbiol. 2019, 57, e01282-18. [Google Scholar] [CrossRef] [Green Version]
- Hong, N.; Chen, M.; Xu, J. Molecular Markers Reveal Epidemiological Patterns and Evolutionary Histories of the Human Pathogenic Cryptococcus. Front. Cell. Infect. Microbiol. 2021, 11, 683670. [Google Scholar] [CrossRef]
- Kan, A.; Schmertmann, L.J.; McArthur, C.; Mella, V.S.A.; Crowther, M.S.; Miranda, L.; Malik, R.; Meyer, W.; Krockenberger, M.B. A Possible Link between the Environment and Cryptococcus gattii Nasal Colonisation in Koalas (Phascolarctos cinereus) in the Liverpool Plains, New South Wales. Int. J. Environ. Res. Public Health 2022, 19, 4603. [Google Scholar] [CrossRef]
- Chen, Y.C.; Kuo, S.F.; Lin, S.Y.; Lin, Y.S.; Lee, C.H. Epidemiological and Clinical Characteristics, Antifungal Susceptibility, and MLST-Based Genetic Analysis of Cryptococcus Isolates in Southern Taiwan in 2013–2020. J. Fungi 2022, 8, 287. [Google Scholar] [CrossRef]
- Fan, X.; Xiao, M.; Chen, S.; Kong, F.; Dou, H.T.; Wang, H.; Xiao, Y.L.; Kang, M.; Sun, Z.Y.; Hu, Z.D.; et al. Predominance of Cryptococcus neoformans var. grubii multilocus sequence type 5 and emergence of isolates with non-wild-type minimum inhibitory concentrations to fluconazole: A multi-centre study in China. Clin. Microbiol. Infect. 2016, 22, 887.E1–887.E9. [Google Scholar] [CrossRef]
- Ferreira-Paim, K.; Andrade-Silva, L.; Fonseca, F.M.; Ferreira, T.B.; Mora, D.J.; Andrade-Silva, J.; Khan, A.; Dao, A.; Reis, E.C.; Almeida, M.T.; et al. MLST-Based Population Genetic Analysis in a Global Context Reveals Clonality amongst Cryptococcus neoformans var. grubii VNI Isolates from HIV Patients in Southeastern Brazil. PLoS Negl. Trop. Dis. 2017, 11, e0005223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loftus, B.J.; Fung, E.; Roncaglia, P.; Rowley, D.; Amedeo, P.; Bruno, D.; Vamathevan, J.; Miranda, M.; Anderson, I.J.; Fraser, J.A.; et al. The genome of the basidiomycetous yeast and human pathogen Cryptococcus neoformans. Science 2005, 307, 1321–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuomo, C.A.; Rhodes, J.; Desjardins, C.A. Advances in Cryptococcus genomics: Insights into the evolution of pathogenesis. Mem. Inst. Oswaldo Cruz. 2018, 113, e170473. [Google Scholar] [CrossRef] [Green Version]
- Gerstein, A.C.; Jackson, K.M.; McDonald, T.R.; Wang, Y.; Lueck, B.D.; Bohjanen, S.; Smith, K.D.; Akampurira, A.; Meya, D.B.; Xue, C.; et al. Identification of Pathogen Genomic Differences That Impact Human Immune Response and Disease during Cryptococcus neoformans Infection. mBio 2019, 10, e01440-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaragoza, O. Basic principles of the virulence of Cryptococcus. Virulence 2019, 10, 490–501. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, J.; Desjardins, C.A.; Sykes, S.M.; Beale, M.A.; Vanhove, M.; Sakthikumar, S.; Chen, Y.; Gujja, S.; Saif, S.; Chowdhary, A.; et al. Tracing Genetic Exchange and Biogeography of Cryptococcus neoformans var. grubii at the Global Population Level. Genetics 2017, 207, 327–346. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.M.; Tao, X.H.; Xu, D.F.; Yu, Y.; Teng, Y.; Xie, W.Q.; Fan, Y.B. HOG1 has an essential role in the stress response, virulence and pathogenicity of Cryptococcus gattii. Exp. Ther. Med. 2021, 21, 476. [Google Scholar] [CrossRef]
- Baddley, J.W.; Chen, S.C.; Huisingh, C.; Benedict, K.; DeBess, E.E.; Galanis, E.; Jackson, B.R.; MacDougall, L.; Marsden-Haug, N.; Oltean, H.; et al. MSG07: An International Cohort Study Comparing Epidemiology and Outcomes of Patients With Cryptococcus neoformans or Cryptococcus gattii Infections. Clin. Infect. Dis. 2021, 73, 1133–1141. [Google Scholar] [CrossRef]
- Cogliati, M. Global Molecular Epidemiology of Cryptococcus neoformans and Cryptococcus gattii: An Atlas of the Molecular Types. Scientifica 2013, 2013, 675213. [Google Scholar] [CrossRef] [Green Version]
- Hagen, F.; Colom, M.F.; Swinne, D.; Tintelnot, K.; Iatta, R.; Montagna, M.T.; Torres-Rodriguez, J.M.; Cogliati, M.; Velegraki, A.; Burggraaf, A.; et al. Autochthonous and dormant Cryptococcus gattii infections in Europe. Emerg. Infect. Dis. 2012, 18, 1618–1624. [Google Scholar] [CrossRef]
- Cogliati, M.; Puccianti, E.; Montagna, M.T.; De Donno, A.; Susever, S.; Ergin, C.; Velegraki, A.; Ellabib, M.S.; Nardoni, S.; Macci, C.; et al. Fundamental niche prediction of the pathogenic yeasts Cryptococcus neoformans and Cryptococcus gattii in Europe. Environ. Microbiol. 2017, 19, 4318–4325. [Google Scholar] [CrossRef] [PubMed]
- Cogliati, M. Global warming impact on the expansion of fundamental niche of Cryptococcus gattii VGI in Europe. Environ. Microbiol. Rep. 2021, 13, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Chase, B.M.; Scott, L.; Meadows, M.E.; Gil-Romera, G.; Boom, A.; Carr, A.S.; Reimer, P.J.; Truc, L.; Valsecchi, V.; Quick, L.J. Rock hyrax middens: A palaeoenvironmental archive for southern African drylands. Quat. Sci. Rev. 2012, 56, 107–125. [Google Scholar] [CrossRef]
- Chen, S.; Sorrell, T.; Nimmo, G.; Speed, B.; Currie, B.; Ellis, D.; Marriott, D.; Pfeiffer, T.; Parr, D.; Byth, K. Epidemiology and host- and variety-dependent characteristics of infection due to Cryptococcus neoformans in Australia and New Zealand. Australasian Cryptococcal Study Group. Clin. Infect. Dis. 2000, 31, 499–508. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.R.; Lockhart, S.R.; Debess, E.; Marsden-Haug, N.; Goldoft, M.; Wohrle, R.; Lee, S.; Smelser, C.; Park, B.; Chiller, T. Cryptococcus gattii in the United States: Clinical aspects of infection with an emerging pathogen. Clin. Infect. Dis. 2011, 53, 1188–1195. [Google Scholar] [CrossRef] [Green Version]
- Galanis, E.; Macdougall, L.; Kidd, S.; Morshed, M. Epidemiology of Cryptococcus gattii, British Columbia, Canada, 1999-2007. Emerg. Infect. Dis. 2010, 16, 251–257. [Google Scholar] [CrossRef]
- Saijo, T.; Chen, J.; Chen, S.C.; Rosen, L.B.; Yi, J.; Sorrell, T.C.; Bennett, J.E.; Holland, S.M.; Browne, S.K.; Kwon-Chung, K.J. Anti-granulocyte-macrophage colony-stimulating factor autoantibodies are a risk factor for central nervous system infection by Cryptococcus gattii in otherwise immunocompetent patients. mBio 2014, 5, e00912–e00914. [Google Scholar] [CrossRef] [Green Version]
- Crum-Cianflone, N.F.; Lam, P.V.; Ross-Walker, S.; Rosen, L.B.; Holland, S.M. Autoantibodies to Granulocyte-Macrophage Colony-Stimulating Factor Associated With Severe and Unusual Manifestations of Cryptococcus gattii Infections. Open Forum Infect. Dis. 2017, 4, ofx211. [Google Scholar] [CrossRef] [Green Version]
- Sim, B.Z.; Conway, L.; Smith, L.K.; Fairhead, L.; Der, Y.S.; Payne, L.; Binotto, E.; Smith, S.; Hanson, J. The aetiology and clinical characteristics of cryptococcal infections in Far North Queensland, tropical Australia. PLoS ONE 2022, 17, e0265739. [Google Scholar] [CrossRef]
- Piffer, A.C.; Santos, F.M.D.; Thomé, M.P.; Diehl, C.; Garcia, A.W.A.; Kinskovski, U.P.; Schneider, R.O.; Gerber, A.; Feltes, B.C.; Schrank, A.; et al. Transcriptomic analysis reveals that mTOR pathway can be modulated in macrophage cells by the presence of cryptococcal cells. Genet. Mol. Biol. 2021, 44, e20200390. [Google Scholar] [CrossRef]
- Holcomb, Z.E.; Steinbrink, J.M.; Zaas, A.K.; Betancourt, M.; Tenor, J.L.; Toffaletti, D.L.; Alspaugh, J.A.; Perfect, J.R.; McClain, M.T. Transcriptional Profiles Elucidate Differential Host Responses to Infection with Cryptococcus neoformans and Cryptococcus gattii. J. Fungi 2022, 8, 430. [Google Scholar] [CrossRef] [PubMed]
- Elsegeiny, W.; Marr, K.A.; Williamson, P.R. Immunology of Cryptococcal Infections: Developing a Rational Approach to Patient Therapy. Front. Immunol. 2018, 9, 651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva-Junior, E.B.; Firmino-Cruz, L.; Guimarães-de-Oliveira, J.C.; De-Medeiros, J.V.R.; de Oliveira Nascimento, D.; Freire-de-Lima, M.; de Brito-Gitirana, L.; Morrot, A.; Previato, J.O.; Mendonça-Previato, L.; et al. The role of Toll-like receptor 9 in a murine model of Cryptococcus gattii infection. Sci. Rep. 2021, 11, 1407. [Google Scholar] [CrossRef] [PubMed]
- Kuo, P.H.; Wu, U.I.; Pan, Y.H.; Wang, J.T.; Wang, Y.C.; Sun, H.Y.; Sheng, W.H.; Chen, Y.C.; Chang, S.C. Neutralizing Anti-Granulocyte-Macrophage Colony-Stimulating Factor Autoantibodies in Patients With Central Nervous System and Localized Cryptococcosis: Longitudinal Follow-up and Literature Review. Clin. Infect. Dis. 2022, 75, 278–287. [Google Scholar] [CrossRef]
- Huang, Y.; Zang, X.; Yang, C.; Deng, H.; Ma, X.; Xie, M.; Zhou, M.; Song, J.; Xue, X. Gene, virulence and related regulatory mechanisms in Cryptococcus gattii. Acta Biochim. Biophys. Sin. 2022, 54, 593–603. [Google Scholar] [CrossRef]
- Wright, L.C.; Payne, J.; Santangelo, R.T.; Simpanya, M.F.; Chen, S.C.; Widmer, F.; Sorrell, T.C. Cryptococcal phospholipases: A novel lysophospholipase discovered in the pathogenic fungus Cryptococcus gattii. Biochem. J. 2004, 384, 377–384. [Google Scholar] [CrossRef] [Green Version]
- Ueno, K.; Otani, Y.; Yanagihara, N.; Urai, M.; Nagamori, A.; Sato-Fukushima, M.; Shimizu, K.; Saito, N.; Miyazaki, Y. Cryptococcus gattii evades CD11b-mediated fungal recognition by coating itself with capsular polysaccharides. Eur. J. Immunol. 2021, 51, 2281–2295. [Google Scholar] [CrossRef]
- Urai, M.; Kaneko, Y.; Ueno, K.; Okubo, Y.; Aizawa, T.; Fukazawa, H.; Sugita, T.; Ohno, H.; Shibuya, K.; Kinjo, Y.; et al. Evasion of Innate Immune Responses by the Highly Virulent Cryptococcus gattii by Altering Capsule Glucuronoxylomannan Structure. Front. Cell. Infect. Microbiol. 2015, 5, 101. [Google Scholar] [CrossRef] [Green Version]
- Castelo-Branco, D.; da Rocha, M.G.; de Oliveira, J.S.; Araujo, G.D.S.; Martins, D.V.; Garcia, L.G.S.; Cordeiro, R.A.; Sidrim, J.J.C.; Pereira-Neto, W.A.; de Melo Guedes, G.M.; et al. The herbicide paraquat alters growth and melanin production on the Cryptococcus neoformans/Cryptococcus gattii species complex. Can. J. Microbiol. 2022, 68, 493–499. [Google Scholar] [CrossRef]
- de Sousa, H.R.; de Oliveira, G.P., Jr.; Frazão, S.O.; Gorgonha, K.C.M.; Rosa, C.P.; Garcez, E.M.; Lucas, J., Jr.; Correia, A.F.; de Freitas, W.F.; Borges, H.M.; et al. Faster Cryptococcus Melanization Increases Virulence in Experimental and Human Cryptococcosis. J. Fungi 2022, 8, 393. [Google Scholar] [CrossRef]
- Saidykhan, L.; Correia, J.; Romanyuk, A.; Peacock, A.F.A.; Desanti, G.E.; Taylor-Smith, L.; Makarova, M.; Ballou, E.R.; May, R.C. An in vitro method for inducing titan cells reveals novel features of yeast-to-titan switching in the human fungal pathogen Cryptococcus gattii. PLoS Pathog. 2022, 18, e1010321. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, M.H.; Machado, M.P.; Kumaresan, P.R.; da Silva, T.A. Titan Cells and Yeast Forms of Cryptococcus neoformans and Cryptococcus gattii Are Recognized by GXMR-CAR. Microorganisms 2021, 9, 1886. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.S.S.; Pinto, L.M.; de Medeiros, M.A.P.; Toffaletti, D.L.; Tenor, J.L.; Barros, T.F.; Neves, R.P.; Neto, R.G.L.; Milan, E.P.; Padovan, A.C.B.; et al. Comparison of Cryptococcus gattii/neoformans Species Complex to Related Genera (Papiliotrema and Naganishia) Reveal Variances in Virulence Associated Factors and Antifungal Susceptibility. Front. Cell. Infect. Microbiol. 2021, 11, 642658. [Google Scholar] [CrossRef] [PubMed]
- Freitas, G.J.C.; Santos, D.A. Cryptococcus gattii polysaccharide capsule: An insight on fungal-host interactions and vaccine studies. Eur. J. Immunol. 2021, 51, 2206–2209. [Google Scholar] [CrossRef]
- Zang, X.; Ke, W.; Wang, L.; Wu, H.; Huang, Y.; Deng, H.; Zhou, M.; Wu, N.; Xue, X.; Shen, D. Molecular epidemiology and microbiological characteristics of Cryptococcus gattii VGII isolates from China. PLoS Negl. Trop. Dis. 2022, 16, e0010078. [Google Scholar] [CrossRef] [PubMed]
- Arendrup, M.C.; Boekhout, T.; Akova, M.; Meis, J.F.; Cornely, O.A.; Lortholary, O. ESCMID and ECMM joint clinical guidelines for the diagnosis and management of rare invasive yeast infections. Clin. Microbiol. Infect. 2014, 20 (Suppl. 3), 76–98. [Google Scholar] [CrossRef] [Green Version]
- Baddley, J.W.; Forrest, G.N. Cryptococcosis in solid organ transplantation—Guidelines from the American Society of Transplantation Infectious Diseases Community of Practice. Clin. Transplant. 2019, 33, e13543. [Google Scholar] [CrossRef] [PubMed]
- Kung, H.C.; Huang, P.Y.; Chen, W.T.; Ko, B.S.; Chen, Y.C.; Chang, S.C.; Chuang, Y.C. 2016 guidelines for the use of antifungal agents in patients with invasive fungal diseases in Taiwan. J. Microbiol. Immunol. Infect. 2018, 51, 1–17. [Google Scholar] [CrossRef]
- Chang, C.C.; Blyth, C.C.; Chen, S.C.; Khanina, A.; Morrissey, C.O.; Roberts, J.A.; Thursky, K.A.; Worth, L.J.; Slavin, M.A. Introduction to the updated Australasian consensus guidelines for the management of invasive fungal disease and use of antifungal agents in the haematology/oncology setting, 2021. Intern. Med. J. 2021, 51 (Suppl. 7), 3–17. [Google Scholar] [CrossRef]
- Chen, S.C.; Sorrell, T.C.; Chang, C.C.; Paige, E.K.; Bryant, P.A.; Slavin, M.A. Consensus guidelines for the treatment of yeast infections in the haematology, oncology and intensive care setting, 2014. Intern. Med. J. 2014, 44, 1315–1332. [Google Scholar] [CrossRef]
- WHO. Guidelines for Diagnosing, Preventing and Managing Cryptococcal Disease among Adults, Adolescents and Children Living with HIV; World Health Organization: Geneva, Switzerland, 2022; pp. 1–64. [Google Scholar]
- Hoenigl, M.; Sprute, R.; Egger, M.; Arastehfar, A.; Cornely, O.A.; Krause, R.; Lass-Flörl, C.; Prattes, J.; Spec, A.; Thompson, G.R., 3rd; et al. The Antifungal Pipeline: Fosmanogepix, Ibrexafungerp, Olorofim, Opelconazole, and Rezafungin. Drugs 2021, 81, 1703–1729. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, M.; Guo, J.; Guo, W.; Zhong, N.; Shen, H.; Cai, J.; Zhu, Z.; Wu, W. In vitro activities of the tetrazole VT-1161 compared with itraconazole and fluconazole against Cryptococcus and non-albicans Candida species. Mycologia 2021, 113, 918–925. [Google Scholar] [CrossRef]
- Qian, A.; Zheng, Y.; Wang, R.; Wei, J.; Cui, Y.; Cao, X.; Yang, Y. Design, synthesis, and structure-activity relationship studies of novel tetrazole antifungal agents with potent activity, broad antifungal spectrum and high selectivity. Bioorg. Med. Chem. Lett. 2018, 28, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Mitsuyama, J.; Nomura, N.; Hashimoto, K.; Yamada, E.; Nishikawa, H.; Kaeriyama, M.; Kimura, A.; Todo, Y.; Narita, H. In vitro and in vivo antifungal activities of T-2307, a novel arylamidine. Antimicrob. Agents Chemother. 2008, 52, 1318–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiederhold, N.P.; Najvar, L.K.; Jaramillo, R.; Olivo, M.; Patterson, H.; Connell, A.; Fukuda, Y.; Mitsuyama, J.; Catano, G.; Patterson, T.F. The Novel Arylamidine T-2307 Demonstrates In Vitro and In Vivo Activity against Candida auris. Antimicrob. Agents Chemother. 2020, 64, e02198-19. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G.; Bustamante, B.; Ticona, E.; Hamill, R.J.; Johnson, P.C.; Reboli, A.; Aberg, J.; Hasbun, R.; Hsu, H.H. Recombinant interferon- gamma 1b as adjunctive therapy for AIDS-related acute cryptococcal meningitis. J. Infect. Dis. 2004, 189, 2185–2191. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, J.N.; Meintjes, G.; Rebe, K.; Williams, G.N.; Bicanic, T.; Williams, A.; Schutz, C.; Bekker, L.G.; Wood, R.; Harrison, T.S. Adjunctive interferon-gamma immunotherapy for the treatment of HIV-associated cryptococcal meningitis: A randomized controlled trial. AIDS 2012, 26, 1105–1113. [Google Scholar] [CrossRef] [Green Version]
- Thwaites, G.E.; Nguyen, D.B.; Nguyen, H.D.; Hoang, T.Q.; Do, T.T.; Nguyen, T.C.; Nguyen, Q.H.; Nguyen, T.T.; Nguyen, N.H.; Nguyen, T.N.; et al. Dexamethasone for the treatment of tuberculous meningitis in adolescents and adults. N. Engl. J. Med. 2004, 351, 1741–1751. [Google Scholar] [CrossRef] [Green Version]
- Beardsley, J.; Wolbers, M.; Kibengo, F.M.; Ggayi, A.B.; Kamali, A.; Cuc, N.T.; Binh, T.Q.; Chau, N.V.; Farrar, J.; Merson, L.; et al. Adjunctive Dexamethasone in HIV-Associated Cryptococcal Meningitis. N. Engl. J. Med. 2016, 374, 542–554. [Google Scholar] [CrossRef]
- Seaton, R.A.; Verma, N.; Naraqi, S.; Wembri, J.P.; Warrell, D.A. The effect of corticosteroids on visual loss in Cryptococcus neoformans var. gattii meningitis. Trans. R. Soc. Trop. Med. Hyg. 1997, 91, 50–52. [Google Scholar] [CrossRef]
- Lane, M.; McBride, J.; Archer, J. Steroid responsive late deterioration in Cryptococcus neoformans variety gattii meningitis. Neurology 2004, 63, 713–714. [Google Scholar] [CrossRef]
- Chang, C.C.; Hall, V.; Cooper, C.; Grigoriadis, G.; Beardsley, J.; Sorrell, T.C.; Heath, C.H. Consensus guidelines for the diagnosis and management of cryptococcosis and rare yeast infections in the haematology/oncology setting, 2021. Intern. Med. J. 2021, 51 (Suppl. 7), 118–142. [Google Scholar] [CrossRef] [PubMed]
- Rhein, J.; Huppler Hullsiek, K.; Tugume, L.; Nuwagira, E.; Mpoza, E.; Evans, E.E.; Kiggundu, R.; Pastick, K.A.; Ssebambulidde, K.; Akampurira, A.; et al. Adjunctive sertraline for HIV-associated cryptococcal meningitis: A randomised, placebo-controlled, double-blind phase 3 trial. Lancet Infect. Dis. 2019, 19, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Villanueva-Lozano, H.; Trevino-Rangel, R.J.; Gonzalez, G.M.; Hernandez-Rodriguez, P.A.; Camacho-Ortiz, A.; Castillo-Reyna, L.; Galindo-Alvarado, S.G.; Martinez-Resendez, M.F. Clinical evaluation of the antifungal effect of sertraline in the treatment of cryptococcal meningitis in HIV patients: A single Mexican center experience. Infection 2018, 46, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Boulware, D.R.; Nalintya, E.; Rajasingham, R.; Kirumira, P.; Naluyima, R.; Turya, F.; Namanda, S.; Rutakingirwa, M.K.; Skipper, C.P.; Nikweri, Y.; et al. Adjunctive sertraline for asymptomatic cryptococcal antigenemia: A randomized clinical trial. Med. Mycol. 2020, 58, 1037–1043. [Google Scholar] [CrossRef]
- Ngan, N.T.T.; Thanh Hoang Le, N.; Vi Vi, N.N.; Van, N.T.T.; Mai, N.T.H.; Van Anh, D.; Trieu, P.H.; Lan, N.P.H.; Phu, N.H.; Chau, N.V.V.; et al. An open label randomized controlled trial of tamoxifen combined with amphotericin B and fluconazole for cryptococcal meningitis. Elife 2021, 10, e68929. [Google Scholar] [CrossRef]
- Ueno, K.; Urai, M.; Takatsuka, S.; Abe, M.; Miyazaki, Y.; Kinjo, Y. Immunization with Antigen-Pulsed Dendritic Cells Against Highly Virulent Cryptococcus gattii Infection: Analysis of Cytokine-Producing T Cells. Methods Mol. Biol. 2017, 1625, 327–339. [Google Scholar] [CrossRef]
- Ueno, K.; Yanagihara, N.; Shimizu, K.; Miyazaki, Y. Vaccines and Protective Immune Memory against Cryptococcosis. Biol. Pharm. Bull. 2020, 43, 230–239. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, L.V.N.; Wang, R.; Specht, C.A.; Levitz, S.M. Vaccines for human fungal diseases: Close but still a long way to go. NPJ Vaccines 2021, 6, 33. [Google Scholar] [CrossRef]
- da Silva, T.A.; Hauser, P.J.; Bandey, I.; Laskowski, T.; Wang, Q.; Najjar, A.M.; Kumaresan, P.R. Glucuronoxylomannan in the Cryptococcus species capsule as a target for Chimeric Antigen Receptor T-cell therapy. Cytotherapy 2021, 23, 119–130. [Google Scholar] [CrossRef]
- Perfect, J.R.; Bicanic, T. Cryptococcosis diagnosis and treatment: What do we know now. Fungal Genet. Biol. 2015, 78, 49–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, S.; Gu, F.; Chen, M.; Wang, C.; Li, J.; Yang, J.; Wang, G.; Zhou, Z.; Yang, Y. A novel method for identifying and distinguishing Cryptococcus neoformans and Cryptococcus gattii by surface-enhanced Raman scattering using positively charged silver nanoparticles. Sci. Rep. 2020, 10, 12480. [Google Scholar] [CrossRef] [PubMed]
- Klein, K.R.; Hall, L.; Deml, S.M.; Rysavy, J.M.; Wohlfiel, S.L.; Wengenack, N.L. Identification of Cryptococcus gattii by use of L-canavanine glycine bromothymol blue medium and DNA sequencing. J. Clin. Microbiol. 2009, 47, 3669–3672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.C.; Ling, J.F.; Chang, F.C.; Wang, S.J.; Fuh, J.L.; Chen, S.S.; Teng, M.M.; Chang, C.Y. Radiological manifestations of cryptococcal infection in central nervous system. J. Chin. Med. Assoc. 2003, 66, 19–26. [Google Scholar]
- Duarte, S.B.L.; Oshima, M.M.; Mesquita, J.; do Nascimento, F.B.P.; de Azevedo, P.C.; Reis, F. Magnetic resonance imaging findings in central nervous system cryptococcosis: Comparison between immunocompetent and immunocompromised patients. Radiol. Bras. 2017, 50, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Song, K.D.; Lee, K.S.; Chung, M.P.; Kwon, O.J.; Kim, T.S.; Yi, C.A.; Chung, M.J. Pulmonary cryptococcosis: Imaging findings in 23 non-AIDS patients. Korean J. Radiol. 2010, 11, 407–416. [Google Scholar] [CrossRef] [Green Version]
- Tintelnot, K.; Hagen, F.; Han, C.O.; Seibold, M.; Rickerts, V.; Boekhout, T. Pitfalls in Serological Diagnosis of Cryptococcus gattii Infections. Med. Mycol. 2015, 53, 874–879. [Google Scholar] [CrossRef] [Green Version]
- Dhana, A. Diagnosis of Cryptococcosis and Prevention of Cryptococcal Meningitis Using a Novel Point-of-Care Lateral Flow Assay. Case Rep. Med. 2013, 2013, 640216. [Google Scholar] [CrossRef] [Green Version]
- Vidal, J.E.; Boulware, D.R. Lateral flow assay for cryptococcal antigen: An important advance to improve the continuum of hiv care and reduce cryptococcal meningitis-related mortality. Rev. Inst. Med. Trop. Sao Paulo 2015, 57 (Suppl. 19), 38–45. [Google Scholar] [CrossRef]
- Tay, E.; Chen, S.C.; Green, W.; Lopez, R.; Halliday, C.L. Development of a Real-Time PCR Assay to Identify and Distinguish between Cryptococcus neoformans and Cryptococcus gattii Species Complexes. J. Fungi 2022, 8, 462. [Google Scholar] [CrossRef]
- Ma, Q.; Yao, J.; Yuan, S.; Liu, H.; Wei, N.; Zhang, J.; Shan, W. Development of a lateral flow recombinase polymerase amplification assay for rapid and visual detection of Cryptococcus neoformans/C. gattii in cerebral spinal fluid. BMC Infect. Dis. 2019, 19, 108. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Y.; Wang, F.; Zhao, M.; Gao, X.; Chen, H.; Li, N.; Zhu, Q.; Liu, L.; Zhu, W.; et al. Development and Application of Rapid Clinical Visualization Molecular Diagnostic Technology for Cryptococcus neoformans/C. gattii Based on Recombinase Polymerase Amplification Combined With a Lateral Flow Strip. Front. Cell. Infect. Microbiol. 2021, 11, 803798. [Google Scholar] [CrossRef] [PubMed]
- Firacative, C.; Trilles, L.; Meyer, W. MALDI-TOF MS enables the rapid identification of the major molecular types within the Cryptococcus neoformans/C. gattii species complex. PLoS ONE 2012, 7, e37566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF mass spectrometry: An emerging technology for microbial identification and diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef] [PubMed]


| Pathogen | Hospitalization and Complications | Mortality by 12 Months | Antifungal Geometric Mean MIC |
|---|---|---|---|
| C. neoformans | Length of hospital stay: 2–210 days | PLWHIV: 20–61% | Fluconazole: 0.5–9.7 µg/mL |
| Renal impairment: 28% | Voriconazole: 0.021–0.5 µg/mL | ||
| Elevated ICP: 18% | Non-HIV: 8–50% | Posaconazole: 0.027–0.10 µg/mL | |
| Blindness: 12% | Amphotericin B: 0.098–0.69 µg/mL | ||
| C. gattii | 10–43% | Fluconazole: 1.46–8.6 µg/mL | |
| Length of ICU stay: 1–29 days. | Voriconazole: 0.02–0.10 µg/mL | ||
| Neurological sequelae: 17–27% at 12 months | Posaconazole: 0.04–0.36 µg/mL | ||
| IRIS: 44% | Amphotericin B: 0.2726–0.39 µg/mL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beardsley, J.; Dao, A.; Keighley, C.; Garnham, K.; Halliday, C.; Chen, S.C.-A.; Sorrell, T.C. What’s New in Cryptococcus gattii: From Bench to Bedside and Beyond. J. Fungi 2023, 9, 41. https://doi.org/10.3390/jof9010041
Beardsley J, Dao A, Keighley C, Garnham K, Halliday C, Chen SC-A, Sorrell TC. What’s New in Cryptococcus gattii: From Bench to Bedside and Beyond. Journal of Fungi. 2023; 9(1):41. https://doi.org/10.3390/jof9010041
Chicago/Turabian StyleBeardsley, Justin, Aiken Dao, Caitlin Keighley, Katherine Garnham, Catriona Halliday, Sharon C.-A. Chen, and Tania C. Sorrell. 2023. "What’s New in Cryptococcus gattii: From Bench to Bedside and Beyond" Journal of Fungi 9, no. 1: 41. https://doi.org/10.3390/jof9010041