
Human Airway Epithelium Responses to Invasive Fungal Infections: A Critical Partner in Innate Immunity

 , ,
, ,
Abstract
:1. Should We Care about Pulmonary Lung Infections?
2. Viewing Human Airway Epithelium as a Monomorphic Barrier Is Outdated
2.1. Mouse Models and Cell Lines Do Not Phenocopy Human Respiratory Epithelial Biology

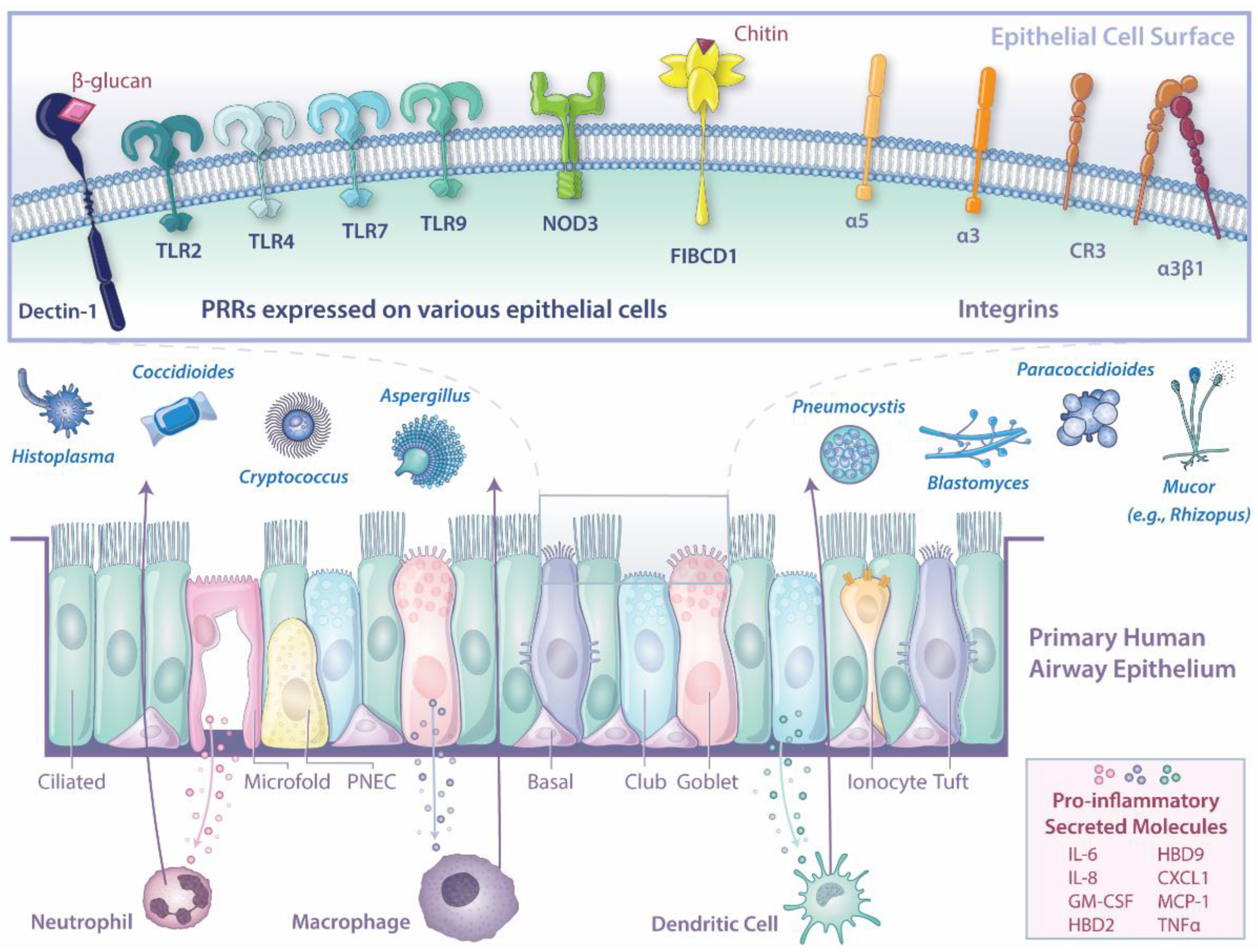{kind=link}
| Cell Line | Source | Immortalization Method | Sex & Ethnicity of Source | Ref |
|---|---|---|---|---|
| A549 | Explant culture of lung carcinomatous tissue; alveolar | N/A | Male; Caucasian | [58] |
| BEAS-2B | Normal human bronchial epithelium (non-cancerous individuals) | Adenovirus 12-SV40 virus | Male; No ethnicity noted | [59,60] |
| Calu-3 | Pleural effusion from an individual with lung adenocarcinoma; bronchial | N/A | Male; Caucasian | [61] |
| HBEC3-KT | Primary human bronchial epithelial cell | hTERT expression & mouse CDK4-expressing retrovirus | Female; No ethnicity noted | [62] |
| H292 | Lymph node metastasis of a pulmonary mucoepidermoid carcinoma | N/A | Female; Black | [63] |
| 16HBE | Human bronchial epithelial cell from a heart-lung patient | Origin-of-replication defective SV40 plasmid | Male; No ethnicity noted | [64] |
2.2. Towards a Better Model of Human Airway Epithelium
3. The Human Airway Epithelium Is a Key Player in the Host-Fungal Pathogen Battle
3.1. First Contact with Fungal Pathogens
3.1.1. Communication Is Key to a Strong and Coordinated Immune Response
3.1.2. DIY Local Immunity—A Novel Paradigm for Lung Immune Responses
3.2. Insights from Non-Fungal Pulmonary Infection Models
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benedict, K.; Jackson, B.R.; Chiller, T.; Beer, K.D. Estimation of direct healthcare costs of fungal diseases in the United States. Clin. Infect. Dis. 2018, 68, 1791–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Lu, G.; Meng, G. Pathogenic fungal infection in the lung. Front. Immunol. 2019, 10, 1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denning, D.W.; Pleuvry, A.; Cole, D.C. Global burden of allergic bronchopulmonary aspergillosis with asthma and its complication chronic pulmonary aspergillosis in adults. Med. Mycol. 2013, 51, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, R.; Muthu, V.; Sehgal, I.S.; Dhooria, S.; Prasad, K.T.; Aggarwal, A.N. Allergic bronchopulmonary aspergillosis. Clin. Chest Med. 2022, 43, 99–125. [Google Scholar] [CrossRef]
- Denning, D.W.; Pleuvry, A.; Cole, D.C. Global burden of chronic pulmonary aspergillosis as a sequel to pulmonary tuberculosis. Bull. World Health Organ. 2011, 89, 864–872. [Google Scholar] [CrossRef]
- Barac, A.; Kosmidis, C.; Alastruey-Izquierdo, A.; Salzer, H.J.F. Chronic pulmonary aspergillosis update: A year in review. Med. Mycol. 2019, 57, S104–S109. [Google Scholar] [CrossRef] [PubMed]
- El-Baba, F.; Gao, Y.; Soubani, A.O. Pulmonary aspergillosis: What the generalist needs to know. Am. J. Med. 2020, 133, 668–674. [Google Scholar] [CrossRef]
- Cadena, J.; Thompson, G.R., 3rd; Patterson, T.F. Aspergillosis: Epidemiology, diagnosis, and treatment. Infect. Dis. Clin. N. Am. 2021, 35, 415–434. [Google Scholar] [CrossRef]
- Dewi, I.M.; Janssen, N.A.; Rosati, D.; Bruno, M.; Netea, M.G.; Brüggemann, R.J.; Verweij, P.E.; van de Veerdonk, F.L. Invasive pulmonary aspergillosis associated with viral pneumonitis. Curr. Opin. Microbiol. 2021, 62, 21–27. [Google Scholar] [CrossRef]
- Schauwvlieghe, A.; Rijnders, B.J.A.; Philips, N.; Verwijs, R.; Vanderbeke, L.; Van Tienen, C.; Lagrou, K.; Verweij, P.E.; Van de Veerdonk, F.L.; Gommers, D.; et al. Invasive aspergillosis in patients admitted to the intensive care unit with severe influenza: A retrospective cohort study. Lancet. Respir. Med. 2018, 6, 782–792. [Google Scholar] [CrossRef]
- Setianingrum, F.; Rautemaa-Richardson, R.; Denning, D.W. Pulmonary cryptococcosis: A review of pathobiology and clinical aspects. Med. Mycol. 2019, 57, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Roux, A.; Canet, E.; Valade, S.; Gangneux-Robert, F.; Hamane, S.; Lafabrie, A.; Maubon, D.; Debourgogne, A.; Le Gal, S.; Dalle, F.; et al. Pneumocystis jirovecii pneumonia in patients with or without AIDS, France. Emerg. Infect. Dis. 2014, 20, 1490–1497. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Su, L.; Jiang, S.J.; Qu, H. Risk factors for mortality from Pneumocystis carinii pneumonia (PCP) in non-HIV patients: A meta-analysis. Oncotarget 2017, 8, 59729–59739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, H.; Chakrabarti, A. Global epidemiology of mucormycosis. J. Fungi 2019, 5, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazi, P.B.; Rauseo, A.M.; Spec, A. Blastomycosis. Infect. Dis. Clin. N. Am. 2021, 35, 515–530. [Google Scholar] [CrossRef] [PubMed]
- McCotter, O.Z.; Benedict, K.; Engelthaler, D.M.; Komatsu, K.; Lucas, K.D.; Mohle-Boetani, J.C.; Oltean, H.; Vugia, D.; Chiller, T.M.; Sondermeyer Cooksey, G.L.; et al. Update on the epidemiology of coccidioidomycosis in the United States. Med. Mycol. 2019, 57, S30–S40. [Google Scholar] [CrossRef]
- Azar, M.M.; Loyd, J.L.; Relich, R.F.; Wheat, L.J.; Hage, C.A. Current concepts in the epidemiology, diagnosis, and management of histoplasmosis syndromes. Semin. Respir. Crit. Care Med. 2020, 41, 13–30. [Google Scholar] [CrossRef]
- Alvarado, P.; Teixeira, M.M.; Cavallera, E.; Paes, H.C.; Guerra, G.; Santander, G.; Merino-Alado, R. Epidemiology of paracoccidioidomycosis in Venezuela: A retrospective study from 1954 to 2019. Mem. Inst. Oswaldo Cruz. 2021, 116, e210203. [Google Scholar] [CrossRef]
- Martinez, R. Epidemiology of paracoccidioidomycosis. Rev. Inst. Med. Trop Sao Paulo 2015, 57 (Suppl. S19), 11–20. [Google Scholar] [CrossRef] [Green Version]
- Zavala, S.; Baddley, J.W. Cryptococcosis. Semin. Respir. Crit. Care Med. 2020, 41, 69–79. [Google Scholar] [CrossRef]
- Schwarz, C.; Brandt, C.; Whitaker, P.; Sutharsan, S.; Skopnik, H.; Gartner, S.; Smazny, C.; Röhmel, J.F. Invasive pulmonary fungal infections in cystic fibrosis. Mycopathologia 2018, 183, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G.; Alexander, B.D.; Andes, D.R.; Hadley, S.; Kauffman, C.A.; Freifeld, A.; Anaissie, E.J.; Brumble, L.M.; Herwaldt, L.; Ito, J.; et al. Invasive fungal infections among organ transplant recipients: Results of the Transplant-Associated Infection Surveillance Network (TRANSNET). Clin. Infect. Dis. 2010, 50, 1101–1111. [Google Scholar] [CrossRef] [PubMed]
- Sipsas, N.V.; Kontoyiannis, D.P. Invasive fungal infections in patients with cancer in the Intensive Care Unit. Int. J. Antimicrob. Agents 2012, 39, 464–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Zhang, N.; Huang, X.; Xiong, S.; Feng, Y.; Zhang, Y.; Li, M.; Zhan, Q. Invasive pulmonary aspergillosis in patients with influenza infection: A retrospective study and review of the literature. Clin. Respir. J. 2019, 13, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Rawson, T.M.; Moore, L.S.P.; Zhu, N.; Ranganathan, N.; Skolimowska, K.; Gilchrist, M.; Satta, G.; Cooke, G.; Holmes, A. Bacterial and fungal coinfection in individuals with coronavirus: A rapid review to support COVID-19 antimicrobial prescribing. Clin. Infect. Dis. 2020, 71, 2459–2468. [Google Scholar] [CrossRef]
- Rayens, E.; Norris, K.A.; Cordero, J.F. Mortality trends in risk conditions and invasive mycotic disease in the United States, 1999–2018. Clin. Infect. Dis. 2021, 74, 309–318. [Google Scholar] [CrossRef]
- Baddley, J.W.; Cantini, F.; Goletti, D.; Gómez-Reino, J.J.; Mylonakis, E.; San-Juan, R.; Fernández-Ruiz, M.; Torre-Cisneros, J. ESCMID Study Group for Infections in Compromised Hosts (ESGICH) Consensus Document on the safety of targeted and biological therapies: An infectious diseases perspective (Soluble immune effector molecules [I]: Anti-tumor necrosis factor-α agents). Clin. Microbiol. Infect. 2018, 24, S10–S20. [Google Scholar] [CrossRef] [Green Version]
- Moldoveanu, B.; Gearhart, A.M.; Jalil, B.A.; Saad, M.; Guardiola, J.J. Pulmonary aspergillosis: Spectrum of disease. Am. J. Med. Sci. 2021, 361, 411–419. [Google Scholar] [CrossRef]
- Thompson, G.R.; Pasqualotto, A.C. Endemic mycoses: Expansion of traditional geographic ranges and pitfalls in management. Mycoses 2021, 64, 989–992. [Google Scholar] [CrossRef]
- Amburgy, J.W.; Miller, J.H.; Ditty, B.J.; Vande Lune, P.; Muhammad, S.; Fisher, W.S., 3rd. Cryptococcus gattii in an immunocompetent patient in the southeastern United States. Case Rep. Infect. Dis. 2016, 2016, 8280915. [Google Scholar] [CrossRef]
- Andreou, M.; Cogliati, M.; Kolonitsiou, F.; Stroumpos, C.; Stamouli, V.; Ravazoula, P.; Siagris, D.; Papadogeorgaki, H.; Christofidou, M.; Lekkou, A. Cryptococcus gattii infection in an immunocompetent host in Greece. Med. Mycol. Case Rep. 2020, 27, 1–3. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, C.A.; Kronstad, J.W.; Taylor, G.; Warren, R.; Yuen, M.; Hu, G.; Jung, W.H.; Sham, A.; Kidd, S.E.; Tangen, K.; et al. Genome variation in Cryptococcus gattii, an emerging pathogen of immunocompetent hosts. mBio 2011, 2, e00342-10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiller, H.B.; Montoro, D.T.; Simon, L.M.; Rawlins, E.L.; Meyer, K.B.; Strunz, M.; Vieira Braga, F.A.; Timens, W.; Koppelman, G.H.; Budinger, G.R.S.; et al. The Human Lung Cell Atlas: A high-resolution reference map of the human lung in health and disease. Am. J. Respir. Cell Mol. Biol. 2019, 61, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, R.J.; Lloyd, C.M. Regulation of immune responses by the airway epithelial cell landscape. Nat. Rev. Immunol. 2021, 21, 347–362. [Google Scholar] [CrossRef]
- Montoro, D.T.; Haber, A.L.; Biton, M.; Vinarsky, V.; Lin, B.; Birket, S.E.; Yuan, F.; Chen, S.; Leung, H.M.; Villoria, J.; et al. A revised airway epithelial hierarchy includes CFTR-expressing ionocytes. Nature 2018, 560, 319–324. [Google Scholar] [CrossRef]
- Zuo, W.-l.; Rostami, M.R.; Shenoy, S.A.; LeBlanc, M.G.; Salit, J.; Strulovici-Barel, Y.; O’Beirne, S.L.; Kaner, R.J.; Leopold, P.L.; Mezey, J.G.; et al. Cell-specific expression of lung disease risk-related genes in the human small airway epithelium. Respir. Res. 2020, 21, 200. [Google Scholar] [CrossRef]
- Rock, J.R.; Onaitis, M.W.; Rawlins, E.L.; Lu, Y.; Clark, C.P.; Xue, Y.; Randell, S.H.; Hogan, B.L. Basal cells as stem cells of the mouse trachea and human airway epithelium. Proc. Natl. Acad. Sci. USA 2009, 106, 12771–12775. [Google Scholar] [CrossRef] [Green Version]
- Plasschaert, L.W.; Žilionis, R.; Choo-Wing, R.; Savova, V.; Knehr, J.; Roma, G.; Klein, A.M.; Jaffe, A.B. A single-cell atlas of the airway epithelium reveals the CFTR-rich pulmonary ionocyte. Nature 2018, 560, 377–381. [Google Scholar] [CrossRef]
- Shah, V.S.; Chivukula, R.R.; Lin, B.; Waghray, A.; Rajagopal, J. Cystic fibrosis and the cells of the airway epithelium: What are ionocytes and what do they do? Annu. Rev. Pathol. Mech. Dis. 2022, 17, 23–46. [Google Scholar] [CrossRef]
- Branchfield, K.; Nantie, L.; Verheyden, J.M.; Sui, P.; Wienhold, M.D.; Sun, X. Pulmonary neuroendocrine cells function as airway sensors to control lung immune response. Science 2016, 351, 707–710. [Google Scholar] [CrossRef]
- Ualiyeva, S.; Hallen, N.; Kanaoka, Y.; Ledderose, C.; Matsumoto, I.; Junger, W.G.; Barrett, N.A.; Bankova, L.G. Airway brush cells generate cysteinyl leukotrienes through the ATP sensor P2Y2. Sci. Immunol. 2020, 5, eaax7224. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Mutoh, M.; Hisamoto, M.; Saito, H.; Takahashi, S.; Asakura, T.; Ishii, M.; Nakamura, Y.; Iida, J.; Hase, K.; et al. Airway M cells arise in the lower airway due to RANKL signaling and reside in the bronchiolar epithelium associated with iBALT in murine models of respiratory disease. Front. Immunol. 2019, 10, 1323. [Google Scholar] [CrossRef] [PubMed]
- Mutoh, M.; Kimura, S.; Takahashi-Iwanaga, H.; Hisamoto, M.; Iwanaga, T.; Iida, J. RANKL regulates differentiation of microfold cells in mouse nasopharynx-associated lymphoid tissue (NALT). Cell Tissue Res. 2016, 364, 175–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, V.R.; Franco, L.H.; Zacharia, V.M.; Khan, H.S.; Stamm, C.E.; You, W.; Marciano, D.K.; Yagita, H.; Levine, B.; Shiloh, M.U. Microfold cells actively translocate Mycobacterium tuberculosis to initiate infection. Cell Rep. 2016, 16, 1253–1258. [Google Scholar] [CrossRef] [Green Version]
- Obar, J.J.; Hohl, T.M.; Cramer, R.A. New advances in invasive aspergillosis immunobiology leading the way towards personalized therapeutic approaches. Cytokine 2016, 84, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Margalit, A.; Kavanagh, K. The innate immune response to Aspergillus fumigatus at the alveolar surface. FEMS Microbiol. Rev. 2015, 39, 670–687. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, R.; Muller, M.M.; Klassert, T.E.; Driesch, D.; Stock, M.; Heinrich, A.; Conrad, T.; Moore, C.; Schier, U.K.; Guthke, R.; et al. Differential regulation of the transcriptomic and secretomic landscape of sensor and effector functions of human airway epithelial cells. Mucosal. Immunol. 2018, 11, 627–642. [Google Scholar] [CrossRef] [Green Version]
- Desoubeaux, G.; Cray, C. Rodent models of invasive aspergillosis due to Aspergillus fumigatus: Still a long path toward standardization. Front. Microbiol. 2017, 8, 841. [Google Scholar] [CrossRef]
- Pappagianis, D. Evaluation of the protective efficacy of the killed Coccidioides immitis spherule vaccine in humans. The Valley Fever Vaccine Study Group. Am. Rev. Respir. Dis. 1993, 148, 656–660. [Google Scholar] [CrossRef]
- Kawamoto, Y.; Morinaga, Y.; Kimura, Y.; Kaku, N.; Kosai, K.; Uno, N.; Hasegawa, H.; Yanagihara, K. TNF-alpha inhibits the growth of Legionella pneumophila in airway epithelial cells by inducing apoptosis. J. Infect. Chemother. 2017, 23, 51–55. [Google Scholar] [CrossRef]
- Chen, F.; Zhang, C.; Jia, X.; Wang, S.; Wang, J.; Chen, Y.; Zhao, J.; Tian, S.; Han, X.; Han, L. Transcriptome profiles of human lung epithelial cells A549 interacting with Aspergillus fumigatus by RNA-seq. PLoS ONE 2015, 10, e0135720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oguma, T.; Asano, K.; Tomomatsu, K.; Kodama, M.; Fukunaga, K.; Shiomi, T.; Ohmori, N.; Ueda, S.; Takihara, T.; Shiraishi, Y.; et al. Induction of mucin and MUC5AC expression by the protease activity of Aspergillus fumigatus in airway epithelial cells. J. Immunol. 2011, 187, 999–1005.e616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiesner, D.L.; Merkhofer, R.M.; Ober, C.; Kujoth, G.C.; Niu, M.; Keller, N.P.; Gern, J.E.; Brockman-Schneider, R.A.; Evans, M.D.; Jackson, D.J.; et al. Club cell TRPV4 serves as a damage sensor driving lung allergic inflammation. Cell Host. Microbe 2020, 27, 614–628. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.Q.; Finkbeiner, W.E.; Wine, J.J.; Mrsny, R.J.; Widdicombe, J.H. Calu-3: A human airway epithelial cell line that shows cAMP-dependent Cl- secretion. Am. J. Physiol. 1994, 266, L493–L501. [Google Scholar] [CrossRef]
- Grainger, C.I.; Greenwell, L.L.; Lockley, D.J.; Martin, G.P.; Forbes, B. Culture of Calu-3 cells at the air interface provides a representative model of the airway epithelial barrier. Pharm. Res. 2006, 23, 1482–1490. [Google Scholar] [CrossRef]
- Ordonez, S.R.; van Eijk, M.; Escobar Salazar, N.; de Cock, H.; Veldhuizen, E.J.A.; Haagsman, H.P. Antifungal activities of surfactant protein D in an environment closely mimicking the lung lining. Mol. Immunol. 2019, 105, 260–269. [Google Scholar] [CrossRef]
- Kaur, R.; Dennison, S.R.; Burrow, A.J.; Rudramurthy, S.M.; Swami, R.; Gorki, V.; Katare, O.P.; Kaushik, A.; Singh, B.; Singh, K.K. Nebulised surface-active hybrid nanoparticles of voriconazole for pulmonary aspergillosis demonstrate clathrin-mediated cellular uptake, improved antifungal efficacy and lung retention. J. Nanobiotechnology 2021, 19, 19. [Google Scholar] [CrossRef]
- Giard, D.J.; Aaronson, S.A.; Todaro, G.J.; Arnstein, P.; Kersey, J.H.; Dosik, H.; Parks, W.P. In vitro cultivation of human tumors: Establishment of cell lines derived from a series of solid tumors. J Natl. Cancer Inst. 1973, 51, 1417–1423. [Google Scholar] [CrossRef]
- Reddel, R.R.; Ke, Y.; Gerwin, B.I.; McMenamin, M.G.; Lechner, J.F.; Su, R.T.; Brash, D.E.; Park, J.B.; Rhim, J.S.; Harris, C.C. Transformation of human bronchial epithelial cells by infection with SV40 or adenovirus-12 SV40 hybrid virus, or transfection via strontium phosphate coprecipitation with a plasmid containing SV40 early region genes. Cancer Res. 1988, 48, 1904–1909. [Google Scholar]
- Lechner, J.F.; LaVeck, M.A. A serum-free method for culturing normal human bronchial epithelial cells at clonal density. J. Tissue Cult. Methods 1985, 9, 43–48. [Google Scholar] [CrossRef]
- Fogh, J.; Fogh, J.M.; Orfeo, T. One hundred and twenty-seven cultured human tumor cell lines producing tumors in nude mice. J. Natl. Cancer Inst. 1977, 59, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, R.D.; Sheridan, S.; Girard, L.; Sato, M.; Kim, Y.; Pollack, J.; Peyton, M.; Zou, Y.; Kurie, J.M.; Dimaio, J.M.; et al. Immortalization of human bronchial epithelial cells in the absence of viral oncoproteins. Cancer Res. 2004, 64, 9027–9034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carney, D.N.; Gazdar, A.F.; Bepler, G.; Guccion, J.G.; Marangos, P.J.; Moody, T.W.; Zweig, M.H.; Minna, J.D. Establishment and identification of small cell lung cancer cell lines having classic and variant features. Cancer Res. 1985, 45, 2913–2923. [Google Scholar] [PubMed]
- Cozens, A.L.; Yezzi, M.J.; Kunzelmann, K.; Ohrui, T.; Chin, L.; Eng, K.; Finkbeiner, W.E.; Widdicombe, J.H.; Gruenert, D.C. CFTR expression and chloride secretion in polarized immortal human bronchial epithelial cells. Am. J. Respir. Cell Mol. Biol. 1994, 10, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Klimecki, W.T. Culture conditions profoundly impact phenotype in BEAS-2B, a human pulmonary epithelial model. J. Appl. Toxicol. 2015, 35, 945–951. [Google Scholar] [CrossRef] [Green Version]
- Stewart, C.E.; Torr, E.E.; Mohd Jamili, N.H.; Bosquillon, C.; Sayers, I. Evaluation of differentiated human bronchial epithelial cell culture systems for asthma research. J. Allergy 2012, 2012, 943982. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Na, T.; Wu, T.; Yuan, B.Z. Human lung epithelial BEAS-2B cells exhibit characteristics of mesenchymal stem cells. PLoS ONE 2020, 15, e0227174. [Google Scholar] [CrossRef]
- Callaghan, P.J.; Ferrick, B.; Rybakovsky, E.; Thomas, S.; Mullin, J.M. Epithelial barrier function properties of the 16HBE14o- human bronchial epithelial cell culture model. Biosci. Rep. 2020, 40, BSR20201532. [Google Scholar] [CrossRef]
- Ferreira Lopes, S.; Vacher, G.; Ciarlo, E.; Savova-Bianchi, D.; Roger, T.; Niculita-Hirzel, H. Primary and immortalized human respiratory cells display different patterns of cytotoxicity and cytokine release upon exposure to deoxynivalenol, nivalenol and fusarenon-X. Toxins 2017, 9, 337. [Google Scholar] [CrossRef] [Green Version]
- Nakauchi, M.; Nagata, N.; Takayama, I.; Saito, S.; Kubo, H.; Kaida, A.; Oba, K.; Odagiri, T.; Kageyama, T. Propagation of rhinovirus C in differentiated immortalized human airway HBEC3-KT epithelial cells. Viruses 2019, 11, 216. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, M.; Sivarajan, R.; Walles, T.; Hackenberg, S.; Steinke, M. Susceptibility of primary human airway epithelial cells to Bordetella pertussis adenylate cyclase toxin in two- and three-dimensional culture conditions. Innate Immun. 2021, 27, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.C.; Wang, S.C.; Chen, Y.T.; Tseng, H.H.; Liu, P.L.; Lin, T.C.; Wu, H.E.; Chen, Y.R.; Tseng, Y.H.; Hsu, J.H.; et al. Corylin ameliorates LPS-induced acute lung injury via suppressing the MAPKs and IL-6/STAT3 signaling pathways. Pharmaceuticals 2021, 14, 1046. [Google Scholar] [CrossRef]
- Liu, X.; Ory, V.; Chapman, S.; Yuan, H.; Albanese, C.; Kallakury, B.; Timofeeva, O.A.; Nealon, C.; Dakic, A.; Simic, V.; et al. ROCK Inhibitor and Feeder Cells Induce the Conditional Reprogramming of Epithelial Cells. Am. J. Pathol. 2012, 180, 599–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rheinwald, J.; Green, H. Serial cultivation of strains of human epidermal keratinocytes: The formation of keratinizing colonies from single cells. Cell 1975, 6, 331–343. [Google Scholar] [CrossRef]
- Mou, H.; Vinarsky, V.; Tata, P.R.; Brazauskas, K.; Choi, S.H.; Crooke, A.K.; Zhang, B.; Solomon, G.M.; Turner, B.; Bihler, H.; et al. Dual SMAD signaling inhibition enables long-term expansion of diverse epithelial basal cells. Cell Stem. Cell 2016, 19, 217–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldman, M.B.; Wood, M.; Lapey, A.; Mou, H. SMAD signaling restricts mucous cell differentiation in human airway epithelium. Am. J. Respir. Cell Mol. Biol. 2019, 61, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Feldman, M.B.; Dutko, R.A.; Wood, M.A.; Ward, R.A.; Leung, H.M.; Snow, R.F.; De La Flor, D.J.; Yonker, L.M.; Reedy, J.L.; Tearney, G.J.; et al. Aspergillus fumigatus cell wall promotes apical airway epithelial recruitment of human neutrophils. Infect. Immun. 2020, 88, e00813–e00819. [Google Scholar] [CrossRef]
- Yonker, L.M.; Marand, A.; Muldur, S.; Hopke, A.; Leung, H.M.; De La Flor, D.; Park, G.; Pinsky, H.; Guthrie, L.B.; Tearney, G.J.; et al. Neutrophil dysfunction in cystic fibrosis. J. Cyst. Fibros 2021, 20, 1062–1071. [Google Scholar] [CrossRef]
- Choi, W.; Yang, A.X.; Sieve, A.; Kuo, S.H.; Mudalagiriyappa, S.; Vieson, M.; Maddox, C.W.; Nanjappa, S.G.; Lau, G.W. Pulmonary mycosis drives forkhead box protein A2 degradation and mucus hypersecretion through activation of the spleen tyrosine kinase-epidermal growth factor receptor-AKT/extracellular signal-regulated kinase 1/2 signaling. Am. J. Pathol. 2021, 191, 108–130. [Google Scholar] [CrossRef]
- Hernández-Santos, N.; Wiesner, D.L.; Fites, J.S.; McDermott, A.J.; Warner, T.; Wüthrich, M.; Klein, B.S. Lung epithelial cells coordinate innate lymphocytes and immunity against pulmonary fungal infection. Cell Host. Microbe 2018, 23, 511–522.e515. [Google Scholar] [CrossRef] [Green Version]
- Kradin, R.L.; Mark, E.J. The pathology of pulmonary disorders due to Aspergillus spp. Arch. Pathol. Lab. Med. 2008, 132, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Amitani, R.; Taylor, G.; Elezis, E.N.; Llewellyn-Jones, C.; Mitchell, J.; Kuze, F.; Cole, P.J.; Wilson, R. Purification and characterization of factors produced by Aspergillus fumigatus which affect human ciliated respiratory epithelium. Infect. Immun. 1995, 63, 3266–3271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karnak, D.; Avery, R.K.; Gildea, T.R.; Sahoo, D.; Mehta, A.C. Endobronchial fungal disease: An under-recognized entity. Respiration 2007, 74, 88–104. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Lillehoj, E.P.; Kai, H.; Kim, K.C. MUC1 expression by human airway epithelial cells mediates Pseudomonas aeruginosa adhesion. Front. Biosci. (Elite Ed.) 2010, 2, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Kato, K.; Lillehoj, E.P.; Lu, W.; Kim, K.C. MUC1: The first respiratory mucin with an anti-inflammatory function. J. Clin. Med. 2017, 6, 110. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Davis, A.S.; Ma, L.; Bishop, L.; Cissé, O.H.; Kutty, G.; Kovacs, J.A. MUC1 mediates Pneumocystis murina binding to airway epithelial cells. Cell Microbiol. 2020, 22, e13182. [Google Scholar] [CrossRef]
- Basu, T.; Seyedmousavi, S.; Sugui, J.A.; Balenga, N.; Zhao, M.; Kwon Chung, K.J.; Biardel, S.; Laviolette, M.; Druey, K.M. Aspergillus fumigatus alkaline protease 1 (Alp1/Asp f13) in the airways correlates with asthma severity. J. Allergy Clin. Immunol. 2018, 141, 423–425.e427. [Google Scholar] [CrossRef] [Green Version]
- Namvar, S.; Warn, P.; Farnell, E.; Bromley, M.; Fraczek, M.; Bowyer, P.; Herrick, S. Aspergillus fumigatus proteases, Asp f 5 and Asp f 13, are essential for airway inflammation and remodelling in a murine inhalation model. Clin. Exp. Allergy 2015, 45, 982–993. [Google Scholar] [CrossRef]
- Homma, T.; Kato, A.; Bhushan, B.; Norton, J.E.; Suh, L.A.; Carter, R.G.; Gupta, D.S.; Schleimer, R.P. Role of Aspergillus fumigatus in triggering protease-activated receptor-2 in airway epithelial cells and skewing the cells toward a T-helper 2 bias. Am. J. Respir. Cell Mol. Biol 2016, 54, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Liu, X.; Chen, F.; Hu, Y.; Li, Z.; Liu, Y.; Han, X.; Sun, Y.; Han, L. Gliotoxin destructs the pulmonary epithelium barrier function by reducing cofilin oligomer formation to promote the dissolution of actin stress fibers. Microb. Pathog. 2018, 123, 169–176. [Google Scholar] [CrossRef]
- Ballard, E.; Yucel, R.; Melchers, W.J.G.; Brown, A.J.P.; Verweij, P.E.; Warris, A. Antifungal activity of antimicrobial peptides and proteins against Aspergillus fumigatus. J. Fungi 2020, 6, 65. [Google Scholar] [CrossRef] [PubMed]
- Alekseeva, L.; Huet, D.; Féménia, F.; Mouyna, I.; Abdelouahab, M.; Cagna, A.; Guerrier, D.; Tichanné-Seltzer, V.; Baeza-Squiban, A.; Chermette, R.; et al. Inducible expression of beta defensins by human respiratory epithelial cells exposed to Aspergillus fumigatus organisms. BMC Microbiol. 2009, 9, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Ahmad, N.; Varadarajan, A.; Vikram, N.; Singh, T.P.; Sharma, S.; Sharma, P. Lactoferrin, a potential iron-chelator as an adjunct treatment for mucormycosis—A comprehensive review. Int. J. Biol. Macromol. 2021, 187, 988–998. [Google Scholar] [CrossRef] [PubMed]
- Smole, U.; Kratzer, B.; Pickl, W.F. Soluble pattern recognition molecules: Guardians and regulators of homeostasis at airway mucosal surfaces. Eur. J. Immunol. 2020, 50, 624–642. [Google Scholar] [CrossRef] [Green Version]
- Koh, S.H.; Shin, S.G.; Andrade, M.J.; Go, R.H.; Park, S.; Woo, C.H.; Lim, J.H. Long pentraxin PTX3 mediates acute inflammatory responses against pneumococcal infection. Biochem. Biophys. Res. Commun. 2017, 493, 671–676. [Google Scholar] [CrossRef]
- D’Angelo, C.; De Luca, A.; Zelante, T.; Bonifazi, P.; Moretti, S.; Giovannini, G.; Iannitti, R.G.; Zagarella, S.; Bozza, S.; Campo, S.; et al. Exogenous pentraxin 3 restores antifungal resistance and restrains inflammation in murine chronic granulomatous disease. J. Immunol. 2009, 183, 4609–4618. [Google Scholar] [CrossRef] [Green Version]
- Cunha, C.; Aversa, F.; Lacerda, J.F.; Busca, A.; Kurzai, O.; Grube, M.; Löffler, J.; Maertens, J.A.; Bell, A.S.; Inforzato, A.; et al. Genetic PTX3 deficiency and aspergillosis in stem-cell transplantation. N. Engl. J. Med. 2014, 370, 421–432. [Google Scholar] [CrossRef]
- Kabbani, D.; Bhaskaran, A.; Singer, L.G.; Bhimji, A.; Rotstein, C.; Keshavjee, S.; Liles, W.C.; Husain, S. Pentraxin 3 levels in bronchoalveolar lavage fluid of lung transplant recipients with invasive aspergillosis. J. Heart Lung. Transplant. 2017, 36, 973–979. [Google Scholar] [CrossRef]
- Lambourne, J.; Agranoff, D.; Herbrecht, R.; Troke, P.F.; Buchbinder, A.; Willis, F.; Letscher-Bru, V.; Agrawal, S.; Doffman, S.; Johnson, E.; et al. Association of mannose-binding lectin deficiency with acute invasive aspergillosis in immunocompromised patients. Clin. Infect. Dis. 2009, 49, 1486–1491. [Google Scholar] [CrossRef]
- Ampel, N.M.; Dionne, S.O.; Giblin, A.; Podany, A.B.; Galgiani, J. Mannose-binding lectin serum levels are low in persons with clinically active coccidioidomycosis. Mycopathologia 2009, 167, 173–180. [Google Scholar] [CrossRef]
- Yanagisawa, K.; Ogawa, Y.; Uchiumi, H.; Gohda, F.; Mawatari, M.; Ishizaki, T.; Mitsui, T.; Yokohama, A.; Handa, H.; Tsukamoto, N.; et al. Gene polymorphisms of mannose-binding lectin confer susceptibility to Pneumocystis pneumonia in HIV-infected patients. J. Infect. Chemother. 2015, 21, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Pottratz, S.T.; Weir, A.L.; Wisniowski, P.E. Pneumocystis carinii attachment increases expression of fibronectin-binding integrins on cultured lung cells. Infect. Immun. 1994, 62, 5464–5469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, B.R.; Barros, B.; Araújo, A.C.L.; Alcantara, C.; Suzuki, E. Paracoccidioides species present distinct fungal adherence to epithelial lung cells and promote different IL-8 secretion levels. Med. Microbiol. Immunol. 2020, 209, 59–67. [Google Scholar] [CrossRef]
- Liu, H.; Lee, M.J.; Solis, N.V.; Phan, Q.T.; Swidergall, M.; Ralph, B.; Ibrahim, A.S.; Sheppard, D.C.; Filler, S.G. Aspergillus fumigatus CalA binds to integrin α(5)β(1) and mediates host cell invasion. Nat. Microbiol. 2016, 2, 16211. [Google Scholar] [CrossRef] [Green Version]
- Alqarihi, A.; Gebremariam, T.; Gu, Y.; Swidergall, M.; Alkhazraji, S.; Soliman, S.S.M.; Bruno, V.M.; Edwards, J.E., Jr.; Filler, S.G.; Uppuluri, P.; et al. GRP78 and integrins play different roles in host cell invasion during mucormycosis. mBio 2020, 11, e01087-20. [Google Scholar] [CrossRef]
- Watkins, T.N.; Gebremariam, T.; Swidergall, M.; Shetty, A.C.; Graf, K.T.; Alqarihi, A.; Alkhazraji, S.; Alsaadi, A.I.; Edwards, V.L.; Filler, S.G.; et al. Inhibition of EGFR signaling protects from mucormycosis. mBio 2018, 9, e01384-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Su, X.; Li, Z.; Liu, Y.; Wang, S.; Zhu, M.; Zhang, C.; Yang, F.; Zhao, J.; Li, X.; et al. Complement receptor 3 mediates Aspergillus fumigatus internalization into alveolar epithelial cells with the increase of intracellular phosphatidic acid by activating FAK. Virulence 2021, 12, 1980–1996. [Google Scholar] [CrossRef]
- McClure, R.; Massari, P. TLR-dependent human mucosal epithelial cell responses to microbial pathogens. Front. Immunol. 2014, 5, 386. [Google Scholar] [CrossRef] [Green Version]
- Jepsen, C.S.; Dubey, L.K.; Colmorten, K.B.; Moeller, J.B.; Hammond, M.A.; Nielsen, O.; Schlosser, A.; Templeton, S.P.; Sorensen, G.L.; Holmskov, U. FIBCD1 binds Aspergillus fumigatus and regulates lung epithelial response to cell wall components. Front. Immunol. 2018, 9, 1967. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.K.; Lu, X.; Li, X.; Sun, Q.Y.; Su, X.; Song, Y.; Sun, H.M.; Shi, Y. Dectin-1 is inducible and plays a crucial role in Aspergillus-induced innate immune responses in human bronchial epithelial cells. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 2755–2764. [Google Scholar] [CrossRef]
- Heyl, K.A.; Klassert, T.E.; Heinrich, A.; Müller, M.M.; Klaile, E.; Dienemann, H.; Grünewald, C.; Bals, R.; Singer, B.B.; Slevogt, H. Dectin-1 is expressed in human lung and mediates the proinflammatory immune response to nontypeable Haemophilus influenzae. mBio 2014, 5, e01492-14. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ghazzi, N.; Moreno-Velásquez, S.; Seidel, C.; Thomson, D.; Denning, D.W.; Read, N.D.; Bowyer, P.; Gago, S. Characterisation of Aspergillus fumigatus endocytic trafficking within airway epithelial cells using high-resolution automated quantitative confocal microscopy. J. Fungi 2021, 7, 454. [Google Scholar] [CrossRef]
- Culibrk, L.; Croft, C.A.; Toor, A.; Yang, S.J.; Singhera, G.K.; Dorscheid, D.R.; Moore, M.M.; Tebbutt, S.J. Phagocytosis of Aspergillus fumigatus by human bronchial epithelial cells is mediated by the Arp2/3 complex and WIPF2. Front. Cell Infect. Microbiol. 2019, 9, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rammaert, B.; Jouvion, G.; de Chaumont, F.; Garcia-Hermoso, D.; Szczepaniak, C.; Renaudat, C.; Olivo-Marin, J.C.; Chrétien, F.; Dromer, F.; Bretagne, S. Absence of fungal spore internalization by bronchial epithelium in mouse models evidenced by a new bioimaging approach and transmission electronic microscopy. Am. J. Pathol. 2015, 185, 2421–2430. [Google Scholar] [CrossRef] [PubMed]
- Merkel, G.J.; Scofield, B.A. The in vitro interaction of Cryptococcus neoformans with human lung epithelial cells. FEMS Immunol. Med. Microbiol. 1997, 19, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Guillot, L.; Carroll, S.F.; Badawy, M.; Qureshi, S.T. Cryptococcus neoformans induces IL-8 secretion and CXCL1 expression by human bronchial epithelial cells. Respir. Res. 2008, 9, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, A.B.M.; Oliveira, J.R.; Souza, A.L.J.; Andrade-Silva, L.; Silva, M.V.; Silva, P.R.; Silva-Vergara, M.L.; Rogerio, A.P. Effects of cigarette smoke extract on bronchial epithelial cells stimulated with Cryptococcus neoformans. Med. Microbiol. Immunol. 2021, 210, 221–233. [Google Scholar] [CrossRef]
- Seidel, C.; Moreno-Velásquez, S.D.; Ben-Ghazzi, N.; Gago, S.; Read, N.D.; Bowyer, P. Phagolysosomal survival enables non-lytic hyphal escape and ramification through lung epithelium during Aspergillus fumigatus infection. Front. Microbiol. 2020, 11, 1955. [Google Scholar] [CrossRef]
- Fernandes, J.; Hamidi, F.; Leborgne, R.; Beau, R.; Castier, Y.; Mordant, P.; Boukkerou, A.; Latgé, J.P.; Pretolani, M. Penetration of the human pulmonary epithelium by Aspergillus fumigatus hyphae. J. Infect. Dis. 2018, 218, 1306–1313. [Google Scholar] [CrossRef]
- Toor, A.; Culibrk, L.; Singhera, G.K.; Moon, K.M.; Prudova, A.; Foster, L.J.; Moore, M.M.; Dorscheid, D.R.; Tebbutt, S.J. Transcriptomic and proteomic host response to Aspergillus fumigatus conidia in an air-liquid interface model of human bronchial epithelium. PLoS ONE 2018, 13, e0209652. [Google Scholar] [CrossRef] [Green Version]
- Fekkar, A.; Balloy, V.; Pionneau, C.; Marinach-Patrice, C.; Chignard, M.; Mazier, D. Secretome of human bronchial epithelial cells in response to the fungal pathogen Aspergillus fumigatus analyzed by differential in-gel electrophoresis. J. Infect. Dis. 2012, 205, 1163–1172. [Google Scholar] [CrossRef] [PubMed]
- Benfield, T.L.; Lundgren, B.; Shelhamer, J.H.; Lundgren, J.D. Pneumocystis carinii major surface glycoprotein induces interleukin-8 and monocyte chemoattractant protein-1 release from a human alveolar epithelial cell line. Eur. J. Clin. Investig. 1999, 29, 717–722. [Google Scholar] [CrossRef]
- Barbosa, F.M.; Fonseca, F.L.; Holandino, C.; Alviano, C.S.; Nimrichter, L.; Rodrigues, M.L. Glucuronoxylomannan-mediated interaction of Cryptococcus neoformans with human alveolar cells results in fungal internalization and host cell damage. Microbes Infect. 2006, 8, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, F.M.; Fonseca, F.L.; Figueiredo, R.T.; Bozza, M.T.; Casadevall, A.; Nimrichter, L.; Rodrigues, M.L. Binding of glucuronoxylomannan to the CD14 receptor in human A549 alveolar cells induces interleukin-8 production. Clin. Vaccine Immunol. 2007, 14, 94–98. [Google Scholar] [CrossRef] [Green Version]
- de Barros, B.; Almeida, B.R.; Suzuki, E. Paracoccidioides brasiliensis downmodulates α3 integrin levels in human lung epithelial cells in a TLR2-dependent manner. Sci. Rep. 2020, 10, 19483. [Google Scholar] [CrossRef]
- de Oliveira, P.; Juliano, M.A.; Tanaka, A.S.; Carmona, A.K.; Dos Santos, S.M.; de Barros, B.C.; Maza, P.K.; Puccia, R.; Suzuki, E. Paracoccidioides brasiliensis induces cytokine secretion in epithelial cells in a protease-activated receptor-dependent (PAR) manner. Med. Microbiol. Immunol. 2017, 206, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Z.; Wang, S.; Zhang, C.; Han, L.; Sun, Q.; Han, X. Aspergillus fumigatus induces the release of IL-8 and MCP-1 by activating nuclear transcription through dectin-1 and CR3 receptors in alveolar epithelial cells. Curr. Microbiol. 2021, 78, 3474–3482. [Google Scholar] [CrossRef]
- Souza, H.I.; Pereira, A.B.M.; Oliveira, J.R.; Silva, P.R.; Teixeira, D.N.S.; Silva-Vergara, M.L.; Rogério, A.P. Cryptococcus neoformans in association with Dermatophagoides pteronyssinus has pro-(IL-6/STAT3 overproduction) and anti-inflammatory (CCL2/ERK1/2 downregulation) effects on human bronchial epithelial cells. Inflammation 2022, 45, 1269–1280. [Google Scholar] [CrossRef]
- Bonnett, C.R.; Cornish, E.J.; Harmsen, A.G.; Burritt, J.B. Early neutrophil recruitment and aggregation in the murine lung inhibit germination of Aspergillus fumigatus conidia. Infect. Immun. 2006, 74, 6528–6539. [Google Scholar] [CrossRef] [Green Version]
- Rowley, J.; Namvar, S.; Gago, S.; Labram, B.; Bowyer, P.; Richardson, M.D.; Herrick, S.E. Differential proinflammatory responses to Aspergillus fumigatus by airway epithelial cells in vitro are protease dependent. J. Fungi 2021, 7, 468. [Google Scholar] [CrossRef]
- Kottom, T.J.; Carmona, E.M.; Limper, A.H. Gene expression in lung epithelial cells following interaction with Pneumocystis carinii and its specific life forms yields insights into host gene responses to infection. Microbiol. Immunol. 2022, 66, 238–251. [Google Scholar] [CrossRef]
- Morton, C.O.; Fliesser, M.; Dittrich, M.; Mueller, T.; Bauer, R.; Kneitz, S.; Hope, W.; Rogers, T.R.; Einsele, H.; Loeffler, J. Gene expression profiles of human dendritic cells interacting with Aspergillus fumigatus in a bilayer model of the alveolar epithelium/endothelium interface. PLoS ONE 2014, 9, e98279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belic, S.; Page, L.; Lazariotou, M.; Waaga-Gasser, A.M.; Dragan, M.; Springer, J.; Loeffler, J.; Morton, C.O.; Einsele, H.; Ullmann, A.J.; et al. Comparative analysis of inflammatory cytokine release and alveolar epithelial barrier invasion in a Transwell® bilayer model of mucormycosis. Front. Microbiol. 2019, 9, 3204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelante, T.; Wong, A.Y.; Ping, T.J.; Chen, J.; Sumatoh, H.R.; Viganò, E.; Hong Bing, Y.; Lee, B.; Zolezzi, F.; Fric, J.; et al. CD103(+) dendritic cells control Th17 cell function in the lung. Cell Rep. 2015, 12, 1789–1801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panda, S.K.; Colonna, M. Innate lymphoid cells in mucosal immunity. Front. Immunol. 2019, 10, 861. [Google Scholar] [CrossRef] [Green Version]
- Kindermann, M.; Knipfer, L.; Obermeyer, S.; Müller, U.; Alber, G.; Bogdan, C.; Schleicher, U.; Neurath, M.F.; Wirtz, S. Group 2 innate lymphoid cells (ILC2) suppress beneficial type 1 immune responses during pulmonary cryptococcosis. Front. Immunol. 2020, 11, 209. [Google Scholar] [CrossRef] [Green Version]
- Gladiator, A.; Wangler, N.; Trautwein-Weidner, K.; LeibundGut-Landmann, S. Cutting edge: IL-17-secreting innate lymphoid cells are essential for host defense against fungal infection. J. Immunol. 2013, 190, 521–525. [Google Scholar] [CrossRef] [Green Version]
- Flaczyk, A.; Duerr, C.U.; Shourian, M.; Lafferty, E.I.; Fritz, J.H.; Qureshi, S.T. IL-33 signaling regulates innate and adaptive immunity to Cryptococcus neoformans. J. Immunol. 2013, 191, 2503–2513. [Google Scholar] [CrossRef] [Green Version]
- Heyen, L.; Müller, U.; Siegemund, S.; Schulze, B.; Protschka, M.; Alber, G.; Piehler, D. Lung epithelium is the major source of IL-33 and is regulated by IL-33-dependent and IL-33-independent mechanisms in pulmonary cryptococcosis. Pathog. Dis. 2016, 74, ftw086. [Google Scholar] [CrossRef] [Green Version]
- Carreto-Binaghi, L.E.; Tenorio, E.P.; Morales-Villarreal, F.R.; Aliouat, E.M.; Zenteno, E.; Martínez-Orozco, J.A.; Taylor, M.L. Detection of cytokines and collectins in bronchoalveolar fluid samples of patients infected with Histoplasma capsulatum and Pneumocystis jirovecii. J. Fungi 2021, 7, 938. [Google Scholar] [CrossRef]
- Sánchez-Maldonado, J.M.; Moñiz-Díez, A.; Ter Horst, R.; Campa, D.; Cabrera-Serrano, A.J.; Martínez-Bueno, M.; Garrido-Collado, M.D.P.; Hernández-Mohedo, F.; Fernández-Puerta, L.; López-Nevot, M.; et al. Polymorphisms within the TNFSF4 and MAPKAPK2 loci influence the risk of developing invasive aspergillosis: A two-stage case control study in the context of the aspBIOmics consortium. J. Fungi 2020, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Rivera, A.; Ro, G.; Van Epps, H.L.; Simpson, T.; Leiner, I.; Sant’Angelo, D.B.; Pamer, E.G. Innate immune activation and CD4+ T cell priming during respiratory fungal infection. Immunity 2006, 25, 665–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhushan, B.; Homma, T.; Norton, J.E.; Sha, Q.; Siebert, J.; Gupta, D.S.; Schroeder, J.W., Jr.; Schleimer, R.P. Suppression of epithelial signal transducer and activator of transcription 1 activation by extracts of Aspergillus fumigatus. Am. J. Respir. Cell Mol. Biol 2015, 53, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Nicolas de Lamballerie, C.; Pizzorno, A.; Fouret, J.; Szpiro, L.; Padey, B.; Dubois, J.; Julien, T.; Traversier, A.; Dulière, V.; Brun, P.; et al. Transcriptional profiling of immune and inflammatory responses in the context of SARS-CoV-2 fungal superinfection in a human airway epithelial model. Microorganisms 2020, 8, 1974. [Google Scholar] [CrossRef]
- Kobayashi, N.; Takahashi, D.; Takano, S.; Kimura, S.; Hase, K. The roles of Peyer’s patches and microfold cells in the gut immune system: Relevance to autoimmune diseases. Front. Immunol. 2019, 10, 2345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mabbott, N.A.; Donaldson, D.S.; Ohno, H.; Williams, I.R.; Mahajan, A. Microfold (M) cells: Important immunosurveillance posts in the intestinal epithelium. Mucosal. Immunol. 2013, 6, 666–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, H.S.; Nair, V.R.; Ruhl, C.R.; Alvarez-Arguedas, S.; Galvan Rendiz, J.L.; Franco, L.H.; Huang, L.; Shaul, P.W.; Kim, J.; Xie, Y.; et al. Identification of scavenger receptor B1 as the airway microfold cell receptor for Mycobacterium tuberculosis. Elife 2020, 9, e52551. [Google Scholar] [CrossRef] [PubMed]
- Albac, S.; Schmitz, A.; Lopez-Alayon, C.; d’Enfert, C.; Sautour, M.; Ducreux, A.; Labruère-Chazal, C.; Laue, M.; Holland, G.; Bonnin, A.; et al. Candida albicans is able to use M cells as a portal of entry across the intestinal barrier in vitro. Cell Microbiol. 2016, 18, 195–210. [Google Scholar] [CrossRef] [Green Version]
- Baughman, R.P.; Kim, C.K.; Vinegar, A.; Hendricks, D.E.; Schmidt, D.J.; Bullock, W.E. The pathogenesis of experimental pulmonary histoplasmosis. Correlative studies of histopathology, bronchoalveolar lavage, and respiratory function. Am. Rev. Respir. Dis. 1986, 134, 771–776. [Google Scholar] [CrossRef]
- Eddens, T.; Elsegeiny, W.; Garcia-Hernadez, M.L.; Castillo, P.; Trevejo-Nunez, G.; Serody, K.; Campfield, B.T.; Khader, S.A.; Chen, K.; Rangel-Moreno, J.; et al. Pneumocystis-driven inducible bronchus-associated lymphoid tissue formation requires Th2 and Th17 immunity. Cell Rep. 2017, 18, 3078–3090. [Google Scholar] [CrossRef]
- Li, L.X.; Hole, C.R.; Rangel-Moreno, J.; Khader, S.A.; Doering, T.L. Cryptococcus neoformans evades pulmonary immunity by modulating xylose precursor transport. Infect. Immun. 2020, 88, e00288-20. [Google Scholar] [CrossRef] [PubMed]
- Moyron-Quiroz, J.E.; Rangel-Moreno, J.; Kusser, K.; Hartson, L.; Sprague, F.; Goodrich, S.; Woodland, D.L.; Lund, F.E.; Randall, T.D. Role of inducible bronchus associated lymphoid tissue (iBALT) in respiratory immunity. Nat. Med. 2004, 10, 927–934. [Google Scholar] [CrossRef]
- Tan, H.X.; Esterbauer, R.; Vanderven, H.A.; Juno, J.A.; Kent, S.J.; Wheatley, A.K. Inducible bronchus-associated lymphoid tissues (iBALT) serve as sites of B cell selection and maturation following influenza infection in mice. Front. Immunol. 2019, 10, 611. [Google Scholar] [CrossRef] [PubMed]
- Sell, S.; Guest, I.; McKinstry, K.K.; Strutt, T.M.; Kohlmeier, J.E.; Brincks, E.; Tighe, M.; Blackman, M.A.; Woodland, D.L.; Dutton, R.W.; et al. Intraepithelial T-cell cytotoxicity, induced bronchus-associated lymphoid tissue, and proliferation of pneumocytes in experimental mouse models of influenza. Viral Immunol. 2014, 27, 484–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusek, M.E.; Pazos, M.A.; Pirzai, W.; Hurley, B.P. In vitro coculture assay to assess pathogen induced neutrophil trans-epithelial migration. J. Vis. Exp. 2014, 83, e50823. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Herbert, J.A.; Robinson, E.; Ren, L.; Smyth, R.L.; Smith, C.M. Neutrophil-airway epithelial interactions result in increased epithelial damage and viral clearance during respiratory syncytial virus infection. J. Virol. 2020, 94, e02161-19. [Google Scholar] [CrossRef] [Green Version]
- Kroes, M.M.; Miranda-Bedate, A.; Jacobi, R.H.J.; van Woudenbergh, E.; den Hartog, G.; van Putten, J.P.M.; de Wit, J.; Pinelli, E. Bordetella pertussis-infected innate immune cells drive the anti-pertussis response of human airway epithelium. Sci. Rep. 2022, 12, 3622. [Google Scholar] [CrossRef]
- Haghi, M.; Hittinger, M.; Zeng, Q.; Oliver, B.; Traini, D.; Young, P.M.; Huwer, H.; Schneider-Daum, N.; Lehr, C.M. Mono- and cocultures of bronchial and alveolar epithelial cells respond differently to proinflammatory stimuli and their modulation by salbutamol and budesonide. Mol. Pharm. 2015, 12, 2625–2632. [Google Scholar] [CrossRef]
- Ugonna, K.; Bingle, C.D.; Plant, K.; Wilson, K.; Everard, M.L. Macrophages are required for dendritic cell uptake of respiratory syncytial virus from an infected epithelium. PLoS ONE 2014, 9, e91855. [Google Scholar] [CrossRef]

| Disease | Fungal Species (**Most Common) | High-Risk Populations for Severe Disease | Global Prevalence | Mortality Rates | Ref |
|---|---|---|---|---|---|
| ABPA * | Aspergillus fumigatus | Asthma | 4.8 million | 10–30% | [3,4] |
| CF | |||||
| CPA * | A. fumigatus ** A. niger, A. flavus, A. nidulans, A. terreus | Chronic lung disease (COPD, CF, asthma), prior mycobacterial infection, ABPA | 3 million | 7–80% | [5,6] |
| IPA * | A. fumigatus ** A. flavus, A. niger, A. terreus | Neutropenia, hematopoietic stem-cell or organ transplant, critically ill, chronic corticosteroid treatment, AIDS | 250,000 | 40–90% | [7,8] |
| Acute Community- Acquired Pneumonia * | A. fumigatus | Prior viral infection (influenza, SARS-CoV-2), COPD | 1–34% of severe viral infections | ~50% | [9,10] |
| Pulmonary Cryptococcosis * | Cryptococcus neoformans, C. gattii | HIV, organ transplantation, malignancies, immuno-suppressive therapy, diabetes | 223,000 | 20–55% | [11] |
| PJP (previously PCP) * | Pneumocystis jirovecii | HIV/AIDS, immune- suppressive medications (e.g., corticosteroids) | 500,000 | 10–50% | [12,13] |
| Pulmonary Mucormycosis * | Rhizopus spp., Mucor spp., Lichtheimia spp. | Cancer, diabetes mellitus, hematopoietic stem cell or organ transplant | >10,000 | 38–82% | [14] |
| Blastomycosis # | Blastomyces dermatitidis; B. gilchristii | HIV, organ transplantation, immunomodulatory therapy | 1–40 cases per 100,000 (in endemic areas) | 50% | [15] |
| CM/Valley Fever # | Coccidioides immitis C. posadasii | HIV, organ transplantation, immunosuppressive therapy, diabetes, pregnancy, older age | 20,000 | 1–88% | [16] |
| Histoplasmosis # | Histoplasma capsulatum | HIV, organ transplantation, immunosuppressive therapy, infancy, older age | 100,000 | 5–8% | [17] |
| PCM # | Paracoccidioides brasiliensis; P. lutzii | Men; individuals in South America | 15,000 | 6–10% | [18,19] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crossen, A.J.; Ward, R.A.; Reedy, J.L.; Surve, M.V.; Klein, B.S.; Rajagopal, J.; Vyas, J.M. Human Airway Epithelium Responses to Invasive Fungal Infections: A Critical Partner in Innate Immunity. J. Fungi 2023, 9, 40. https://doi.org/10.3390/jof9010040
Crossen AJ, Ward RA, Reedy JL, Surve MV, Klein BS, Rajagopal J, Vyas JM. Human Airway Epithelium Responses to Invasive Fungal Infections: A Critical Partner in Innate Immunity. Journal of Fungi. 2023; 9(1):40. https://doi.org/10.3390/jof9010040
Chicago/Turabian StyleCrossen, Arianne J., Rebecca A. Ward, Jennifer L. Reedy, Manalee V. Surve, Bruce S. Klein, Jayaraj Rajagopal, and Jatin M. Vyas. 2023. "Human Airway Epithelium Responses to Invasive Fungal Infections: A Critical Partner in Innate Immunity" Journal of Fungi 9, no. 1: 40. https://doi.org/10.3390/jof9010040