
Inhibition of Aflatoxin Production by Citrinin and Non-Enzymatic Formation of a Novel Citrinin-Kojic Acid Adduct
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Media
2.3. Detection of Microorganisms That are Inhibitors of Aflatoxin Production
2.3.1. Visual Agar Plate Assay
2.3.2. Microtiter Agar Plate Assay
2.4. Identification of the Isolated Fungi
2.5. Tip Culture Method
2.6. Feeding Experiment
2.7. RT-PCR
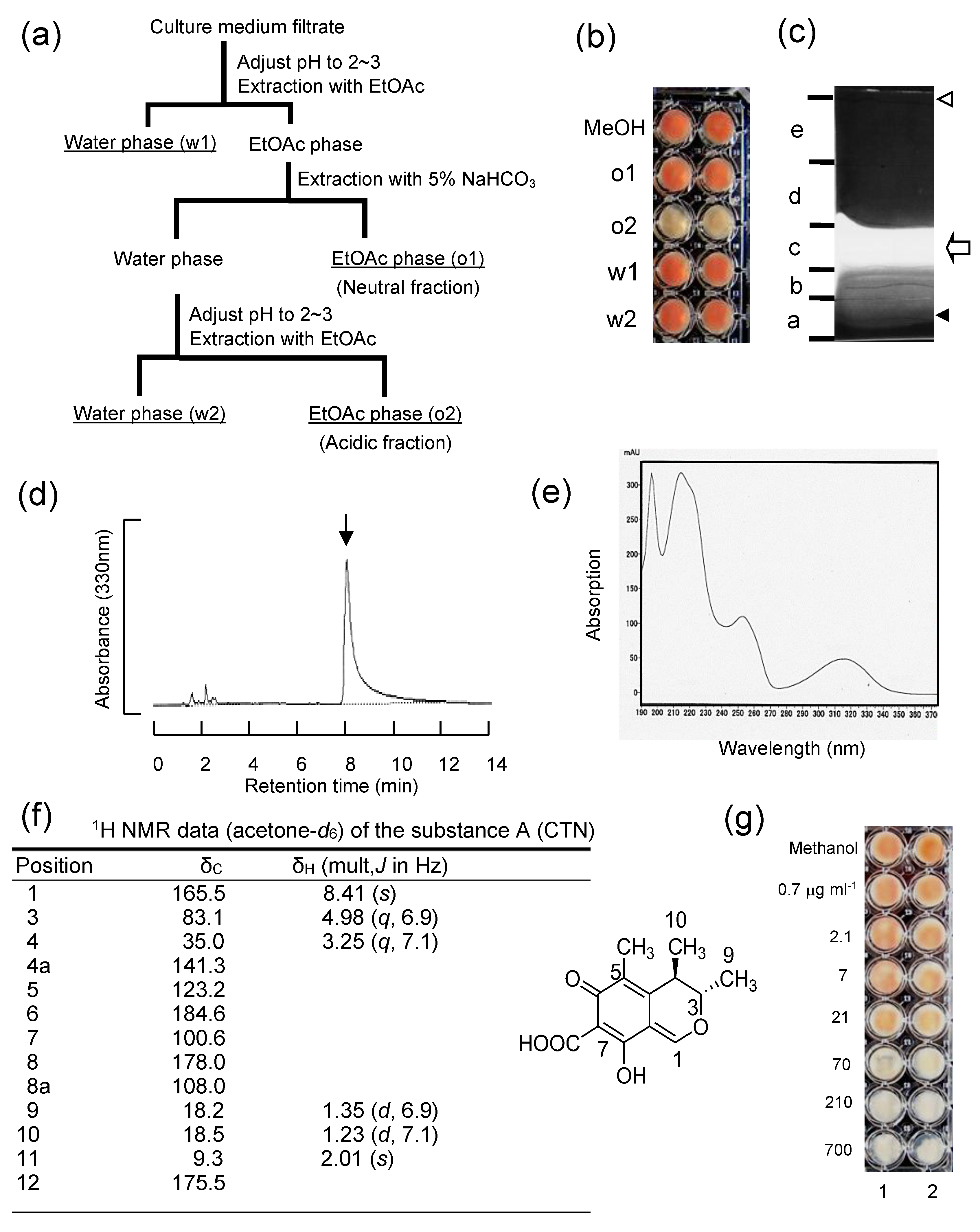
2.8. Purification and Characterization
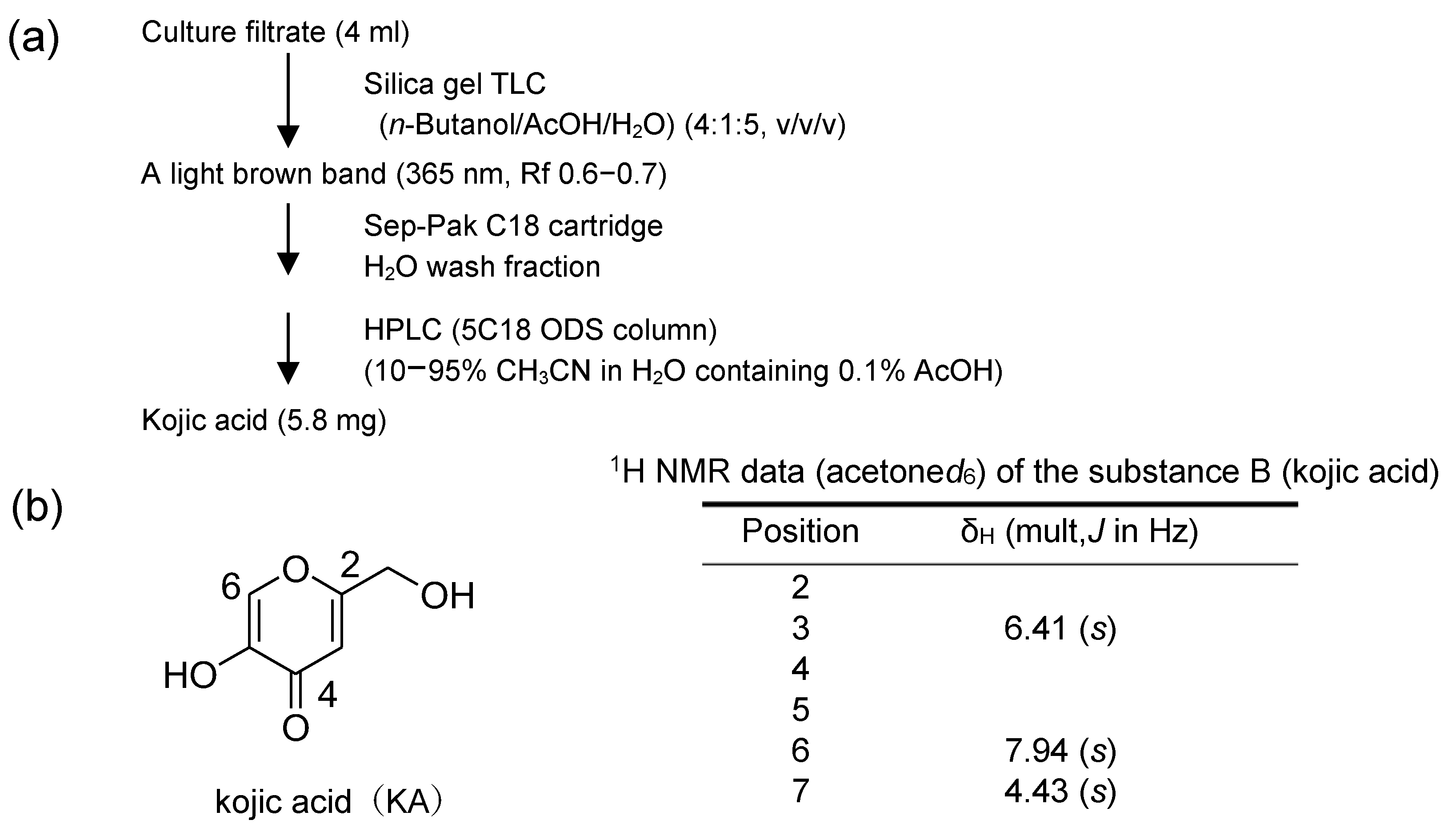
2.8.1. Inhibitory Substance to Aflatoxin Biosynthesis (Substance A: CTN)
2.8.2. The Substance Causing a Decrease in the Yellow Fluorescence of CTN (Substance B: KA)
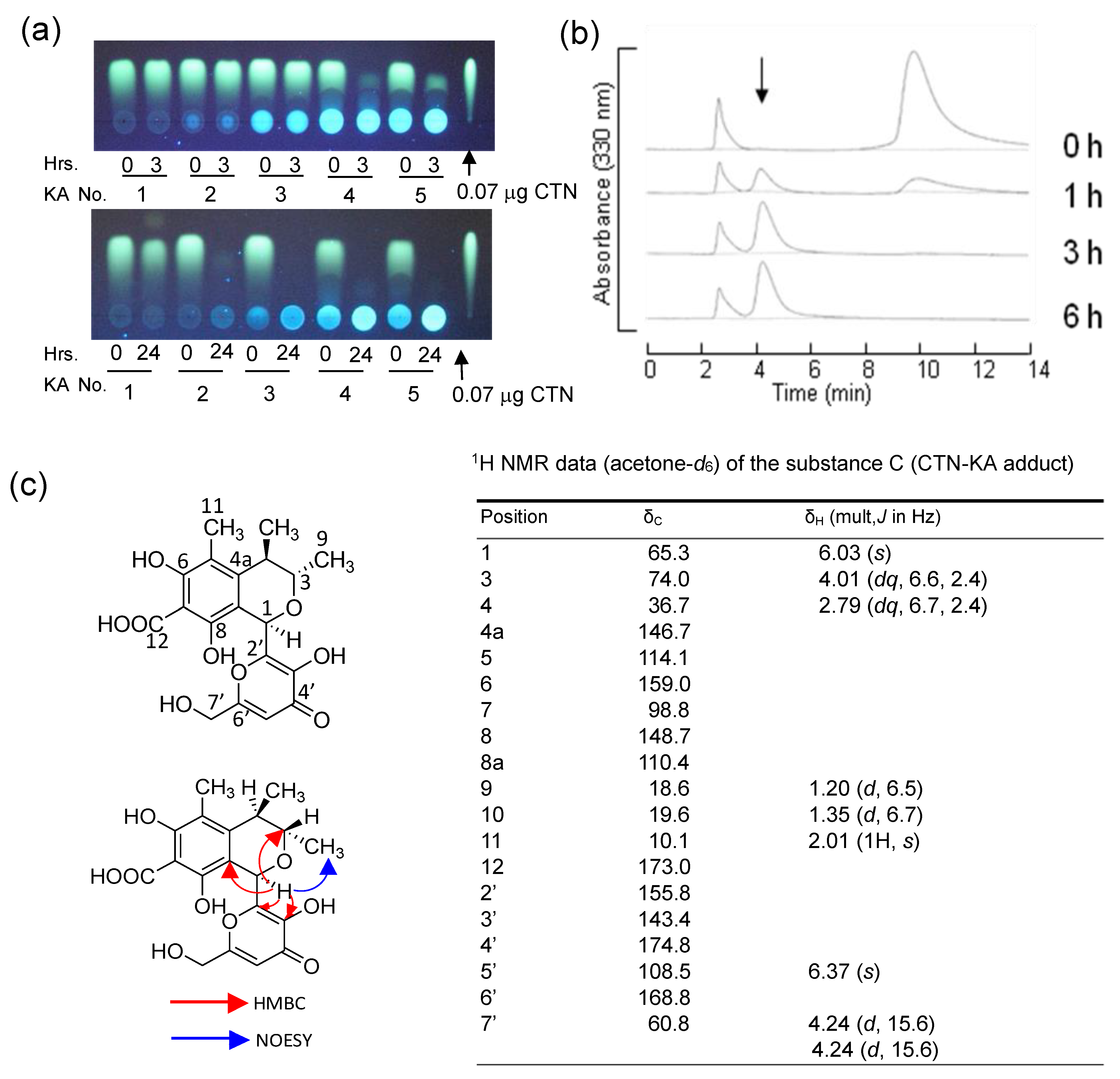
2.8.3. The Reaction Product Caused by the Incubation of CTN and Substance B (Substance C: CTN-KA Adduct)
2.9. Physico-Chemical Analyses
3. Results
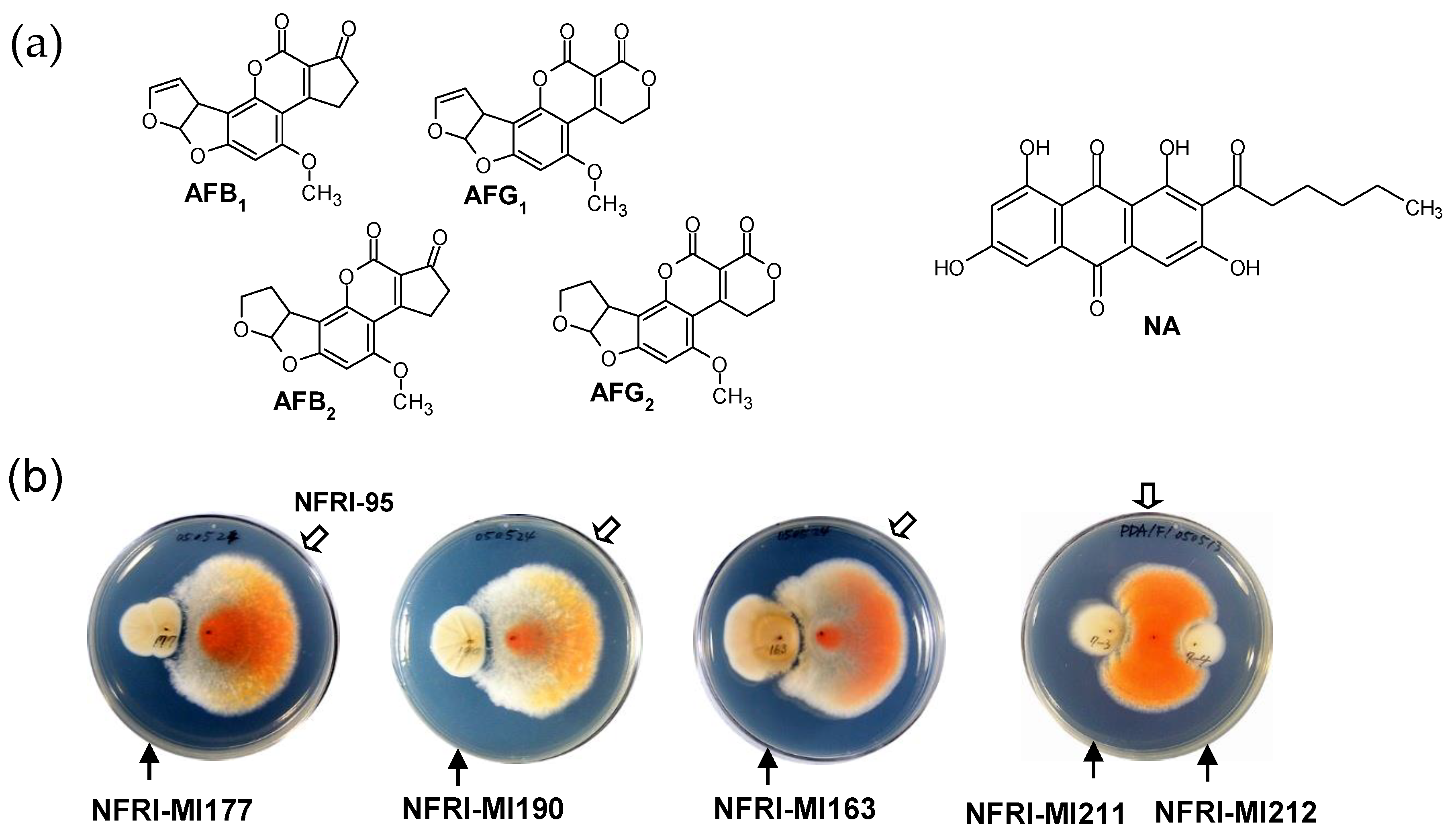
3.1. Isolation of Microorganisms That Inhibit the Production of AFs
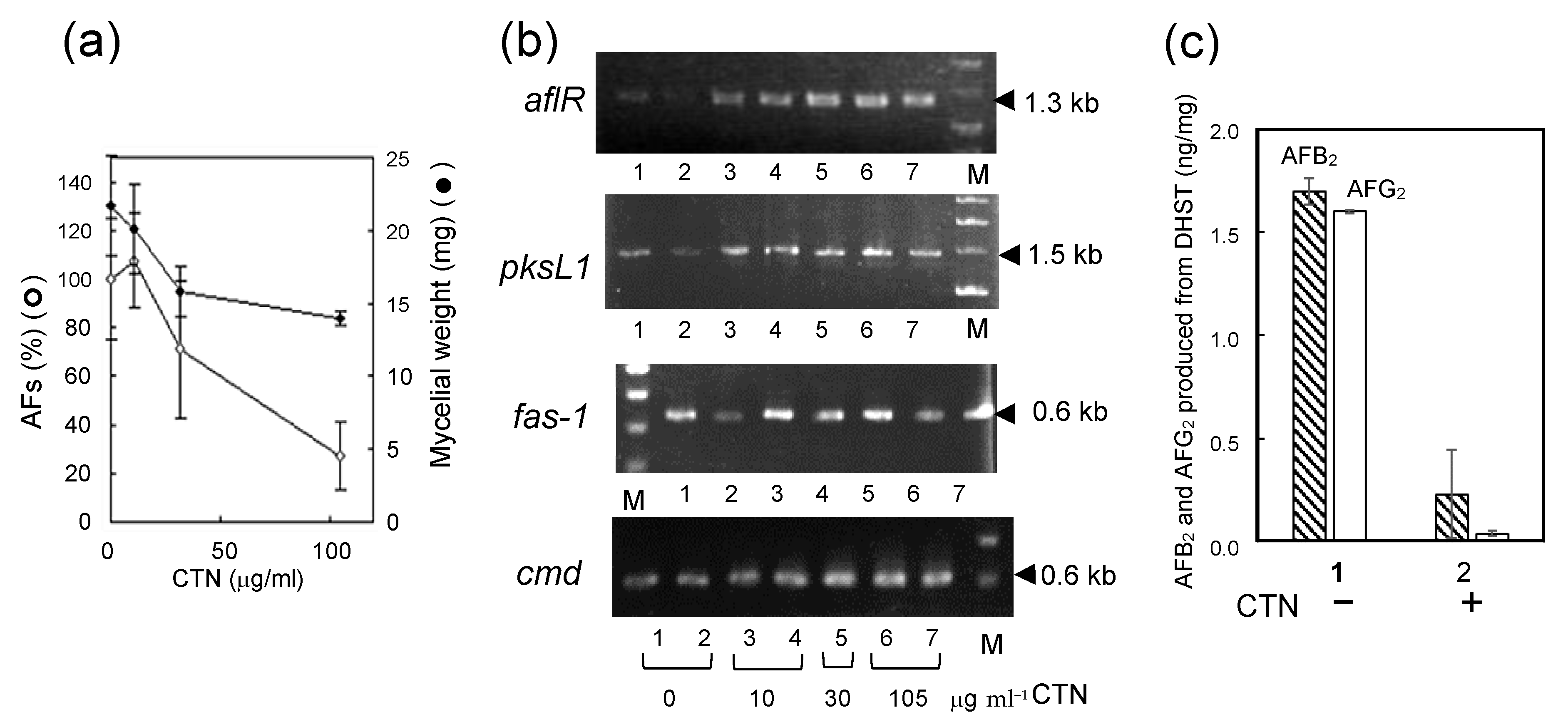
3.2. The Substance Inhibiting the Biosynthesis of AFs
3.3. The Mechanism Underlying the Inhibitory Effect of CTN on the Biosynthesis of AFs
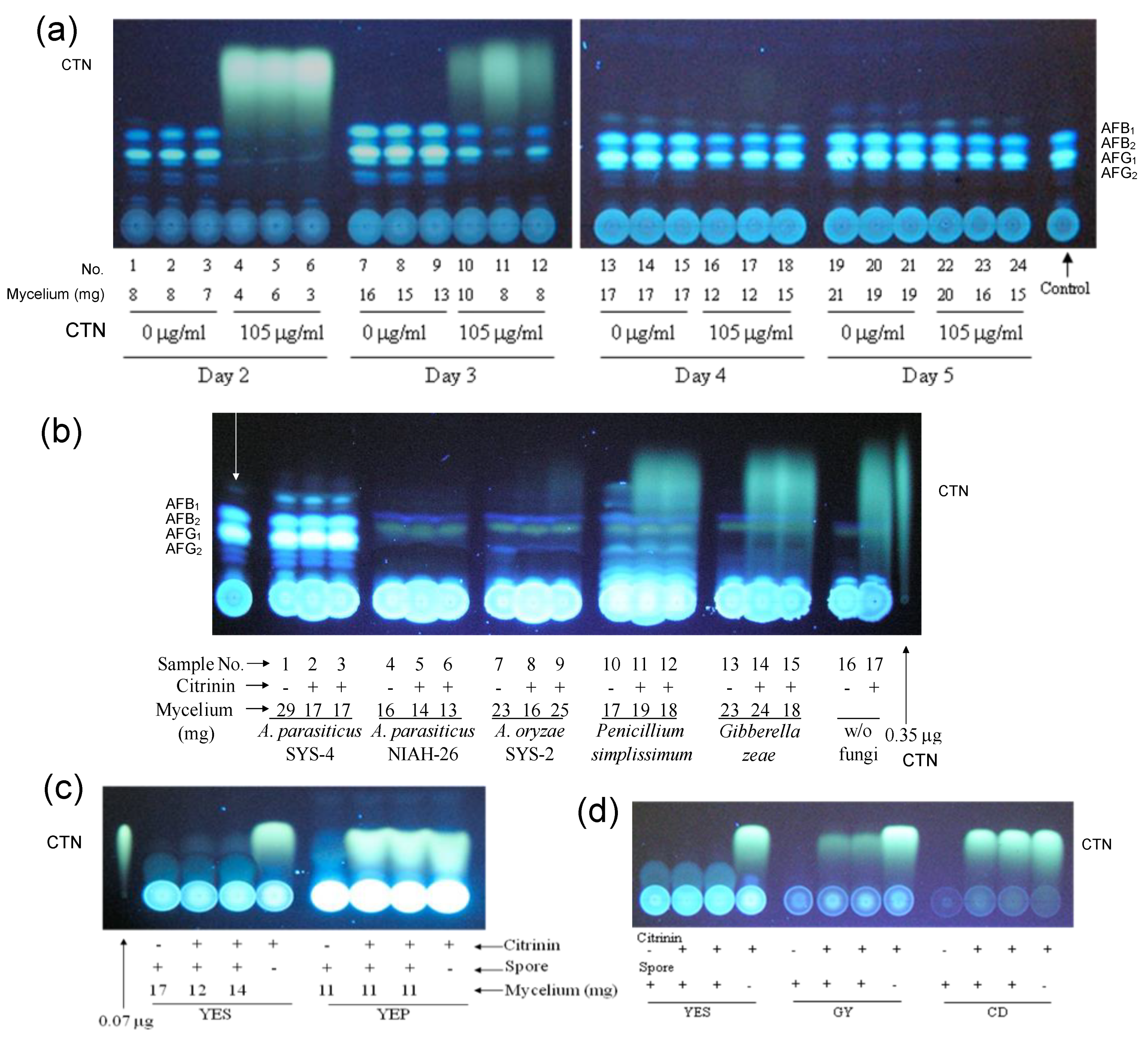
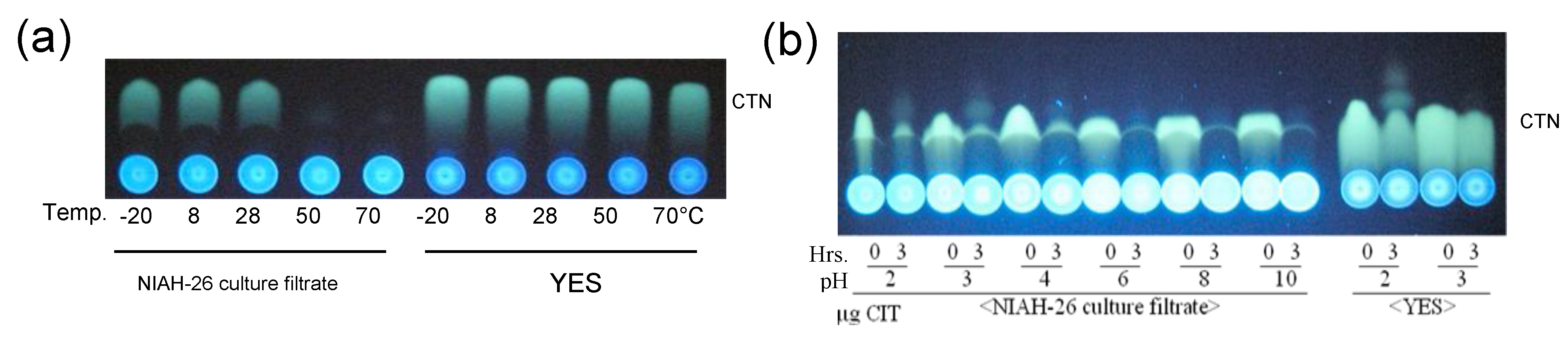
3.4. Decrease in the Fluorescence of CTN by Aspergillus Species and the Inhibition of Fungal Growth by CTN
3.5. Identification of the Substance Causing a Decrease in the Yellow Fluorescence of CTN
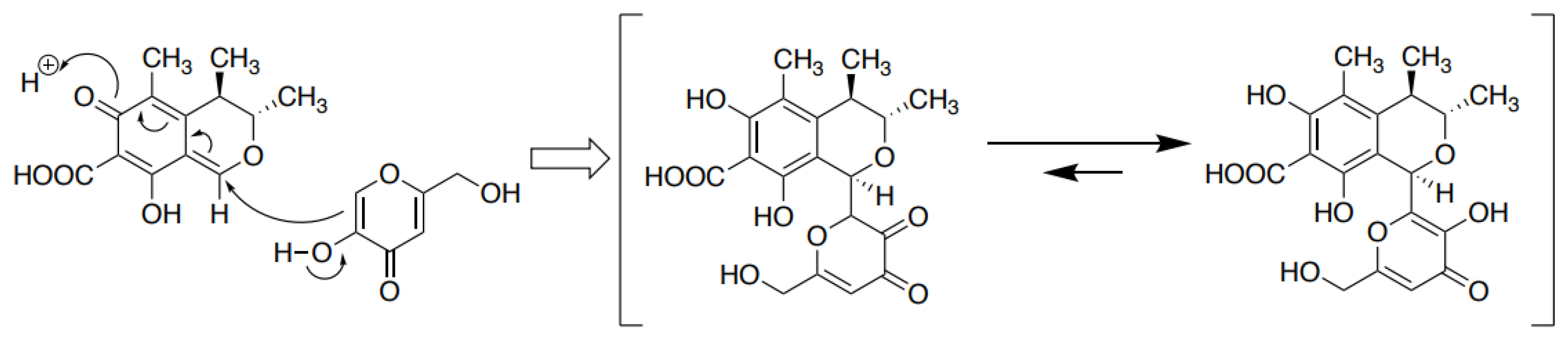
3.6. The Formation of the CTN-KA Adduct
3.7. Stability of the CTN-KA Adduct
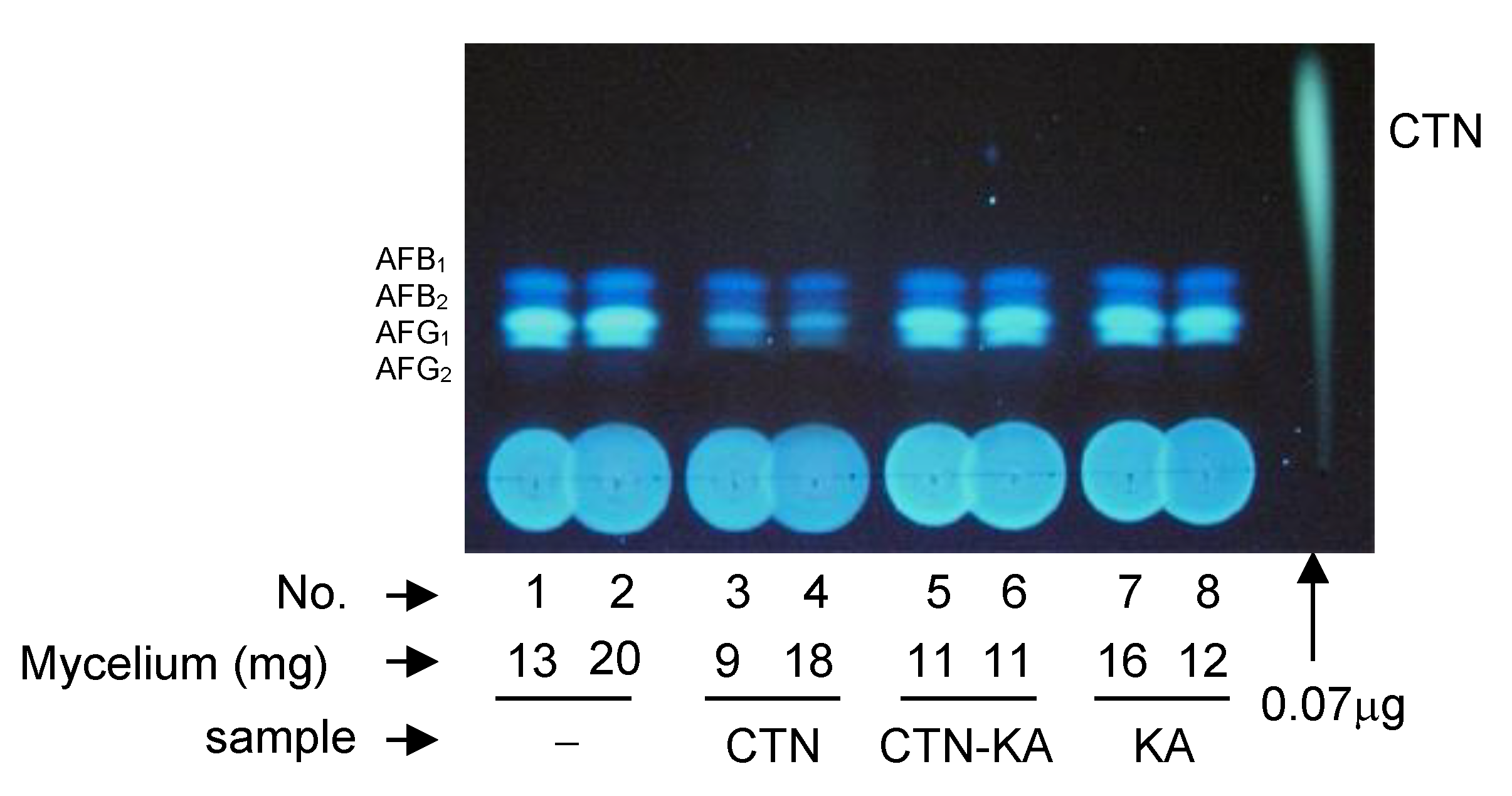
3.8. Effect of the CTN-KA Adduct on the Production of AFs
4. Discussion
4.1. CTN’s Inhibition of Aflatoxin Production and Fungal Growth
4.2. Formation of CTN-KA Adduct
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tola, M.; Kebede, B. Occurrence, Importance and Control of Mycotoxins: A Review. Cogent Food Agric. 2016, 2, 1191103. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Mycotoxin Regulations in 2003 and Current Developments. In Worldwide Regulations for Mycotoxins in Food and Feed; Food and Agriculture Organization of the United Nations: Rome, Italy, 2004; pp. 9–28. [Google Scholar]
- Higginson, J.; DeVita, V.T. IARC Monographs on the Evaluation of Carcinogenic Risk of Chemicals to Humans. Am. Ind. Hyg. Assoc. J. 1980, 41, A26–A28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, J.H.; Phillips, T.D.; Jolly, P.E.; Stiles, J.K.; Jolly, C.M.; Aggarwal, D. Human Aflatoxin in Developing Countries: A Review of Toxicology, Exposure, Potential Health Consequences, and Interventions. Am. Soc. Clin. Nutr. 2004, 80, 1106–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.; Liu, Y.; Bhatnagar, D. Cost-Effectiveness of Aflatoxin Control Methods: Economic Incentives. Toxin Rev. 2008, 27, 203–225. [Google Scholar] [CrossRef]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [Green Version]
- Yu, J. Current Understanding on Aflatoxin Biosynthesis and Future Perspective in Reducing Aflatoxin Contamination. Toxins 2012, 4, 1024–1057. [Google Scholar] [CrossRef] [Green Version]
- Yabe, K.; Chihaya, N.; Hatabayashi, H.; Kito, M.; Hoshino, S.; Zeng, H.; Cai, J.; Nakajima, H. Production of M-/GM-Group Aflatoxins Catalyzed by the OrdA Enzyme in Aflatoxin Biosynthesis. Fungal Genet. Biol. 2012, 49, 744–754. [Google Scholar] [CrossRef]
- Ehrlich, K.C. Non-Aflatoxigenic Aspergillus flavus to Prevent Aflatoxin Contamination in Crops: Advantages and Limitations. Front. Microbiol. 2014, 5, 50. [Google Scholar] [CrossRef]
- Yoshinari, T.; Akiyama, T.; Nakamura, K.; Kondo, T.; Takahashi, Y.; Muraoka, Y.; Nonomura, Y.; Nagasawa, H.; Sakuda, S. Dioctatin A Is a Strong Inhibitor of Aflatoxin Production by Aspergillus parasiticus. Microbiology 2007, 153, 2774–2780. [Google Scholar] [CrossRef] [Green Version]
- Rajani, P.; Sridevi, V.; Lakshmi, M.V.V.C. A Review on Biological Control of Aflatoxin Crop Contamination. Int. J. Chem. Environ. Pharm. Res. 2012, 3, 83–86. [Google Scholar] [CrossRef]
- Abbas, H.K.; Zablotowicz, R.M.; Bruns, H.A.; Abel, C.A. Biocontrol of Aflatoxin in Corn by Inoculation with Non-Aflatoxigenic Aspergillus flavus Isolates. Biocontrol Sci. Technol. 2006, 16, 437–449. [Google Scholar] [CrossRef]
- Yabe, K.; Yan, P.S.; Song, Y.; Ichinomiya, M.; Nakagawa, H.; Shima, Y.; Ando, Y.; Sakuno, E.; Nakajima, H. Isolation of Microorganisms and Substances Inhibitory to Aflatoxin Production. Food Addit. Contam. 2008, 25, 1111–1117. [Google Scholar] [CrossRef]
- Yan, P.; Song, Y.; Sakuno, E.; Nakajima, H. Cyclo (l-Leucyl-l-Prolyl) Produced by Achromobacter xylosoxidans Inhibits Aflatoxin Production by Aspergillus parasiticus. Appl. Environ. Microbiol. 2004, 70, 7466–7473. [Google Scholar] [CrossRef] [Green Version]
- Iimura, K.; Furukawa, T.; Yamamoto, T.; Negishi, L.; Suzuki, M.; Sakuda, S. The Mode of Action of Cyclo(L-Ala-L-Pro) in Inhibiting Aflatoxin Production of Aspergillus flavus. Toxins 2017, 9, 219. [Google Scholar] [CrossRef] [Green Version]
- Kamle, M.; Mahato, D.K.; Gupta, A.; Pandhi, S.; Sharma, B.; Sharma, N.; Sharma, B.; Mishra, S.; Arora, S.; Selvakumar, R.; et al. Citrinin Mycotoxin Contamination in Food and Feed: Impact on Agriculture, Human Health, and Detection and Management Strategies. Toxins 2022, 14, 85. [Google Scholar] [CrossRef]
- Silva, L.J.G.; Pereira, A.M.P.T.; Pena, A.; Lino, C.M. Citrinin in Foods and Supplements: A Review of Occurrence and Analytical Methodologies. Foods 2021, 10, 14. [Google Scholar] [CrossRef]
- Doughari, J.H. The Occurrence, Properties and Significance of Citrinin Mycotoxin. J. Plant Pathol. Microbiol. 2015, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Yabe, K.; Nakamura, H.; Ando, Y.; Terakado, N.; Nakajima, H.; Hamasaki, T. Isolation and Characterization of Aspergillus parasiticus Mutants with Impaired Aflatoxin Production by a Novel Tip Culture Method. Appl. Environ. Microbiol. 1988, 54, 2096–2100. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Zeng, H.; Shima, Y.; Hatabayashi, H.; Nakagawa, H.; Ito, Y.; Adachi, Y.; Nakajima, H.; Yabe, K. Involvement of the NadA Gene in Formation of G-Group Aflatoxins in Aspergillus parasiticus. Fungal Genet. Biol. 2008, 45, 1081–1093. [Google Scholar] [CrossRef]
- Bentley, R. Preparation and Analysis of Kojic Acid. Methods Enzymol. 1957, 3, 238–241. [Google Scholar] [CrossRef]
- Davies, J.E.; Kirkaldy, D.; Roberts, J.C. 437. Studies in Mycological Chemistry. Part VII. Sterigmatocystin, a Metabolite of Aspergillus versicolor(Vuillemin) Tiraboschi. J. Chem. Soc. 1960, 1960, 2169–2178. [Google Scholar] [CrossRef]
- Yabe, K.; Ando, Y.; Hamasaki, T. Biosynthetic Relationship among Aflatoxins B1, B2, G1, and G2. Appl. Environ. Microbiol. 1988, 54, 2101–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sankawa, U.; Ebizuka, Y.; Noguchi, H.; Isikawa, Y.; Kitaghawa, S.; Yamamoto, Y.; Kobayashi, T.; Iitak, Y.; Seto, H. Biosynthesis of Citrinin in Aspergillus Terreus. Tetrahedron 1983, 39, 3583–3591. [Google Scholar] [CrossRef]
- Poupko, R.; Luz, Z.; Destro, R. Carbon-13 NMR of Citrinin in the Solid State and in Solutions. J. Phys. Chem. A 1997, 101, 5097–5102. [Google Scholar] [CrossRef]
- Kushiro, M. Historical Review of Researches on Yellow Rice and Mycotoxigenic Fungi Adherent to Rice in Japan. Mycotoxins 2015, 65, 19–23. [Google Scholar] [CrossRef] [Green Version]
- Flajs, D.; Peraica, M. Toxicological Properties of Citrinin. Arh. Hig. Rada Toksikol. 2009, 60, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Iwahashi, H.; Kitagawa, E.; Suzuki, Y.; Ueda, Y.; Ishizawa, Y.; Nobumasa, H.; Kuboki, Y.; Hosoda, H.; Iwahashi, Y. Evaluation of Toxicity of the Mycotoxin Citrinin Using Yeast ORF DNA Microarray and Oligo DNA Microarray. BMC Genom. 2007, 8, 95. [Google Scholar] [CrossRef] [Green Version]
- Leite, M.C.A.; De Brito Bezerra, A.P.; De Sousa, J.P.; Guerra, F.Q.S.; De Oliveira Lima, E. Evaluation of Antifungal Activity and Mechanism of Action of Citral against Candida albicans. Evid.-Based Complement. Altern. Med. 2014, 2014, 378280. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Dwivedi, P.D.; Dhawan, A.; Das, M.; Ansari, K.M. Citrinin-Generated Reactive Oxygen Species Cause Cell Cycle Arrest Leading to Apoptosis via the Intrinsic Mitochondrial Pathway in Mouse Skin. Toxicol. Sci. 2011, 122, 557–566. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.C.; Chan, W.H. Inhibition of Citrinin-Induced Apoptotic Biochemical Signaling in Human Hepatoma G2 Cells by Resveratrol. Int. J. Mol. Sci. 2009, 10, 3338–3357. [Google Scholar] [CrossRef]
- Da Lozzo, E.J.; Oliveira, M.B.M.; Carnieri, E.G.S. Citrinin-Induced Mitochondrial Permeability Transition. J. Biochem. Mol. Toxicol. 1998, 12, 291–297. [Google Scholar] [CrossRef]
- Haraguchi, H.; Tanaka, T.; Taniguchi, M.; Oi, S.; Hashimoto, K. Inhibitory Effects of Citrinin on Mitochondrial Function. Agric. Biol. Chem. 1986, 51, 1453–1454. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, A.; Doi, E.; Kitabatake, N. Toxic Compounds Formed on Prolonged Heating of Citrinin under Watery Conditions. J. Food Sci. 1993, 58, 229–232. [Google Scholar] [CrossRef]
- Hirota, M.; Menta, A.B.; Yoneyama, K.; Kitabatake, N. A Major Decomposition Product, Citrinin H2, from Citrinin on Heating with Moisture. Biosci. Biotechnol. Biochem. 2002, 66, 206–210. [Google Scholar] [CrossRef]
- Commission Regulation (EU) 2019/1901 of 7 November 2019 Amending Regulation (EC) No 1881/2006 as Regards Maximum Levels of Citrinin in Food Supplements Based on Rice Fermented with Red Yeast Monascus purpureus. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32019R1901&from=EN (accessed on 1 June 2021).
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin Whitening Agents: Medicinal Chemistry Perspective of Tyrosinase Inhibitors. J. Enzyme Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef] [Green Version]
- Ashooriha, M.; Khoshneviszadeh, M.; Khoshneviszadeh, M.; Rafiei, A.; Kardan, M.; Yazdian-Robati, R.; Emami, S. Kojic Acid–Natural Product Conjugates as Mushroom Tyrosinase Inhibitors. Eur. J. Med. Chem. 2020, 201, 112480. [Google Scholar] [CrossRef]
- Poór, M.; Lemli, B.; Bálint, M.; Hetényi, C.; Sali, N.; Kőszegi, T.; Kunsági-Máté, S. Interaction of Citrinin with Human Serum Albumin. Toxins 2015, 7, 5155–5166. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ichinomiya, M.; Fukushima-Sakuno, E.; Kawamoto, A.; Nakagawa, H.; Hatabayashi, H.; Nakajima, H.; Yabe, K. Inhibition of Aflatoxin Production by Citrinin and Non-Enzymatic Formation of a Novel Citrinin-Kojic Acid Adduct. J. Fungi 2023, 9, 29. https://doi.org/10.3390/jof9010029
Ichinomiya M, Fukushima-Sakuno E, Kawamoto A, Nakagawa H, Hatabayashi H, Nakajima H, Yabe K. Inhibition of Aflatoxin Production by Citrinin and Non-Enzymatic Formation of a Novel Citrinin-Kojic Acid Adduct. Journal of Fungi. 2023; 9(1):29. https://doi.org/10.3390/jof9010029
Chicago/Turabian StyleIchinomiya, Masayuki, Emi Fukushima-Sakuno, Ayaka Kawamoto, Hiroyuki Nakagawa, Hidemi Hatabayashi, Hiromitsu Nakajima, and Kimiko Yabe. 2023. "Inhibition of Aflatoxin Production by Citrinin and Non-Enzymatic Formation of a Novel Citrinin-Kojic Acid Adduct" Journal of Fungi 9, no. 1: 29. https://doi.org/10.3390/jof9010029