
More than Just Protein Degradation: The Regulatory Roles and Moonlighting Functions of Extracellular Proteases Produced by Fungi Pathogenic for Humans
 , , and
, , and
Abstract
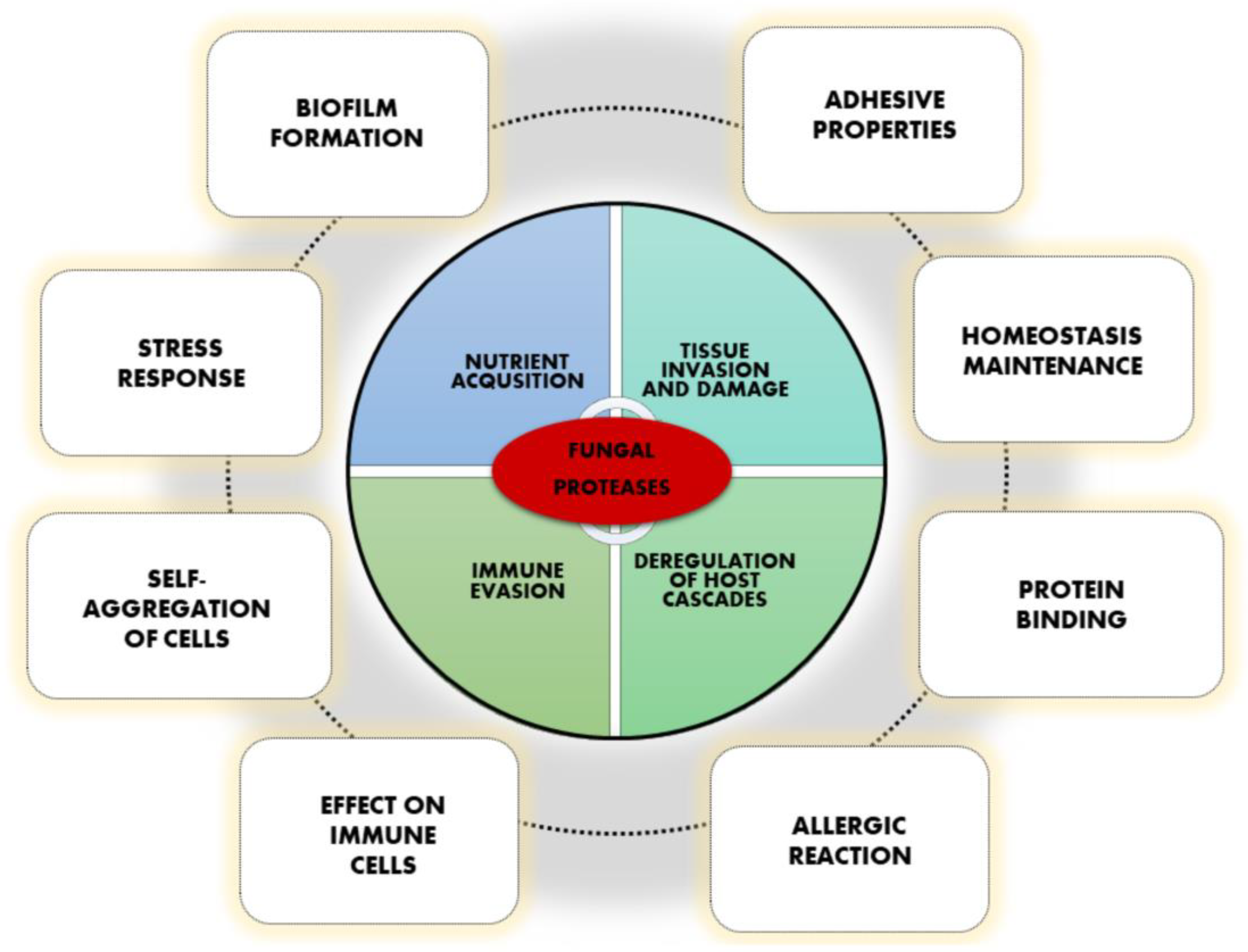
:1. Fungal Proteases Are Important Virulence Factors
2. Involvement in Binding, Adhesion, Self-Aggregation, and Biofilm Formation
3. Effect on Cells of the Host Immune System
4. Fungal Proteases as Allergens
5. Multiple Roles in Cellular Homeostasis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hogan, L.H.; Klein, B.S.; Levitz, S.M. Virulence factors of medically important fungi. Clin. Microbiol. Rev. 1996, 9, 469–488. [Google Scholar] [CrossRef] [PubMed]
- Naglik, J.R.; Challacombe, S.J.; Hube, B. Candida albicans secreted aspartyl proteinases in virulence and pathogenesis. Microbiol. Mol. Biol. Rev. 2003, 67, 400–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutherford, J.C. The emerging role of urease as a general microbial virulence factor. PLoS Pathog. 2014, 10, e1004062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapala-Kozik, M.; Bochenska, O.; Zajac, D.; Karkowska-Kuleta, J.; Gogol, M.; Zawrotniak, M.; Kozik, A. Extracellular proteinases of Candida species pathogenic yeasts. Mol. Oral Microbiol. 2018, 33, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Aoki, W.; Kitahara, N.; Miura, N.; Morisaka, H.; Yamamoto, Y.; Kuroda, K.; Ueda, M. Comprehensive characterization of secreted aspartic proteases encoded by a virulence gene family in Candida albicans. J. Biochem. 2011, 150, 431–438. [Google Scholar] [CrossRef]
- Jeffery, C.J. Moonlighting proteins. Trends Biochem. Sci. 1999, 24, 8–11. [Google Scholar] [CrossRef]
- Satala, D.; Karkowska-Kuleta, J.; Zelazna, A.; Rapala-Kozik, M.; Kozik, A. Moonlighting Proteins at the Candidal Cell Surface. Microorganisms 2020, 8, 1046. [Google Scholar] [CrossRef]
- Mani, M.; Chen, C.; Amblee, V.; Liu, H.; Mathur, T.; Zwicke, G.; Zabad, S.; Patel, B.; Thakkar, J.; Jeffery, C.J. MoonProt: A database for proteins that are known to moonlight. Nucleic Acids Res. 2015, 43, D277–D282. [Google Scholar] [CrossRef] [Green Version]
- Jarocki, V.M.; Tacchi, J.L.; Djordjevic, S.P. Non-proteolytic functions of microbial proteases increase pathological complexity. Proteomics 2015, 15, 1075–1088. [Google Scholar] [CrossRef]
- López-Otín, C.; Bond, J.S. Proteases: Multifunctional enzymes in life and disease. J. Biol. Chem. 2008, 283, 30433–30437. [Google Scholar] [CrossRef]
- Copley, S.D. Moonlighting is mainstream: Paradigm adjustment required. Bioessays 2012, 34, 578–588. [Google Scholar] [CrossRef]
- Rayens, E.; Norris, K.A.; Cordero, J.C.D.S.F. Mortality Trends in Risk Conditions and Invasive Mycotic Disease in the United States, 1999-2018. Clin. Infect. Dis. 2022, 74, 309–318. [Google Scholar] [CrossRef]
- Köhler, J.R.; Casadevall, A.; Perfect, J. The spectrum of fungi that infects humans. Cold Spring Harb. Perspect. Med. 2014, 5, a019273. [Google Scholar] [CrossRef] [Green Version]
- Lockhart, S.R.; Toda, M.; Benedict, K.; Caceres, D.H.; Litvintseva, A.P. Endemic and Other Dimorphic Mycoses in The Americas. J. Fungi 2021, 7, 151. [Google Scholar] [CrossRef]
- Modrzewska, B.; Kurnatowski, P. Adherence of Candida sp. to host tissues and cells as one of its pathogenicity featurese. Ann. Parasitol. 2015, 61, 3–9. [Google Scholar]
- Marcos, C.M.; de Oliveira, H.C.; da Silva, J.d.F.; Assato, P.A.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. The multifaceted roles of metabolic enzymes in the Paracoccidioides species complex. Front. Microbiol. 2014, 5, 719. [Google Scholar] [CrossRef]
- Kumar, R.; Saraswat, D.; Tati, S.; Edgerton, M. Novel Aggregation Properties of Candida albicans Secreted Aspartyl Proteinase Sap6 Mediate Virulence in Oral Candidiasis. Infect. Immun. 2015, 83, 2614–2626. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, R.; Robertson, D.P.; Hodge, P.J.; Lappin, D.F.; Ramage, G. Hydrolytic enzyme production is associated with Candida albicans biofilm formation from patients with type 1 diabetes. Mycopathologia 2010, 170, 229–235. [Google Scholar] [CrossRef]
- Schaller, M.; Schackert, C.; Korting, H.C.; Januschke, E.; Hube, B. Invasion of Candida albicans correlates with expression of secreted aspartic proteinases during experimental infection of human epidermis. J. Investig. Dermatol. 2000, 114, 712–717. [Google Scholar] [CrossRef] [Green Version]
- Korting, H.C.; Hube, B.; Oberbauer, S.; Januschke, E.; Hamm, G.; Albrecht, A.; Borelli, C.; Schaller, M. Reduced expression of the hyphal-independent Candida albicans proteinase genes SAP1 and SAP3 in the efg1 mutant is associated with attenuated virulence during infection of oral epithelium. J. Med. Microbiol. 2003, 52, 623–632. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, A.; Felk, A.; Pichova, I.; Naglik, J.R.; Schaller, M.; De Groot, P.; MacCallum, D.; Odds, F.C.; Schäfer, W.; Klis, F.; et al. Glycosylphosphatidylinositol-anchored proteases of Candida albicans target proteins necessary for both cellular processes and host-pathogen interactions. J. Biol. Chem. 2006, 281, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Schild, L.; Heyken, A.; de Groot, P.W.J.; Hiller, E.; Mock, M.; de Koster, C.; Horn, U.; Rupp, S.; Hube, B. Proteolytic cleavage of covalently linked cell wall proteins by Candida albicans Sap9 and Sap10. Eukaryot. Cell 2011, 10, 98–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoldt, V.R.; Sonneborn, A.; Leuker, C.E.; Ernst, J.F. Efg1p, an essential regulator of morphogenesis of the human pathogen Candida albicans, is a member of a conserved class of bHLH proteins regulating morphogenetic processes in fungi. EMBO J. 1997, 16, 1982–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Tsang, P.C.S.; Pow, E.H.N.; Lam, O.L.T.; Tsang, P.W.K. Potential role of Candida albicans secreted aspartic protease 9 in serum induced-hyphal formation and interaction with oral epithelial cells. Microb. Pathog. 2020, 139, 103896. [Google Scholar] [CrossRef] [PubMed]
- Dutton, L.C.; Jenkinson, H.F.; Lamont, R.J.; Nobbs, A.H. Role of Candida albicans secreted aspartyl protease Sap9 in interkingdom biofilm formation. Pathog. Dis. 2016, 74, ftw005. [Google Scholar] [CrossRef] [Green Version]
- Staniszewska, M.; Bondaryk, M.; Zukowski, K.; Chudy, M. Role of SAP7-10 and Morphological Regulators (EFG1, CPH1) in Candida albicans’ Hypha Formation and Adhesion to Colorectal Carcinoma Caco-2. Polish J. Microbiol. 2015, 64, 203–210. [Google Scholar] [CrossRef]
- Ray, T.L.; Payne, C.D. Scanning electron microscopy of epidermal adherence and cavitation in murine candidiasis: A role for Candida acid proteinase. Infect. Immun. 1988, 56, 1942–1949. [Google Scholar] [CrossRef] [Green Version]
- Kvaal, C.; Lachke, S.A.; Srikantha, T.; Daniels, K.; Mccoy, J.; Soll, D.R. Misexpression of the opaque-phase-specific gene PEP1 (SAP1) in the white phase of Candida albicans confers increased virulence in a mouse model of cutaneous infection. Infect. Immun. 1999, 67, 6652–6662. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Downs, D.; Ghosh, K.; Ghosh, A.K.; Staib, P.; Monod, M.; Tang, J. Candida albicans secreted aspartic proteases 4-6 induce apoptosis of epithelial cells by a novel Trojan horse mechanism. FASEB J. 2013, 27, 2132–2144. [Google Scholar] [CrossRef]
- Kumar, R.; Breindel, C.; Saraswat, D.; Cullen, P.J.; Edgerton, M. Candida albicans Sap6 amyloid regions function in cellular aggregation and zinc binding, and contribute to zinc acquisition. Sci. Rep. 2017, 7, 2908. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.K.; Németh, T.; Papp, A.; Tóth, R.; Lukácsi, S.; Heidingsfeld, O.; Dostal, J.; Vágvölgyi, C.; Bajtay, Z.; Józsi, M.; et al. Functional Characterization of Secreted Aspartyl Proteases in Candida parapsilosis. mSphere 2019, 4, e00484-19. [Google Scholar] [CrossRef]
- Vinterová, Z.; Šanda, M.; Dostál, J.; Hrušková-Heidingsfeldová, O.; Pichová, I. Evidence for the presence of proteolytically active secreted aspartic proteinase 1 of Candida parapsilosis in the cell wall. Protein Sci. 2011, 20, 2004–2012. [Google Scholar] [CrossRef] [Green Version]
- Kaur, R.; Ma, B.; Cormack, B.P. A family of glycosylphosphatidylinositol-linked aspartyl proteases is required for virulence of Candida glabrata. Proc. Natl. Acad. Sci. USA 2007, 104, 7628–7633. [Google Scholar] [CrossRef] [Green Version]
- Monod, M.; Hube, B.; Hess, D.; Sanglard, D. Differential regulation of SAP8 and SAP9, which encode two new members of the secreted aspartic proteinase family in Candida albicans. Microbiology 1998, 144, 2731–2737. [Google Scholar] [CrossRef] [Green Version]
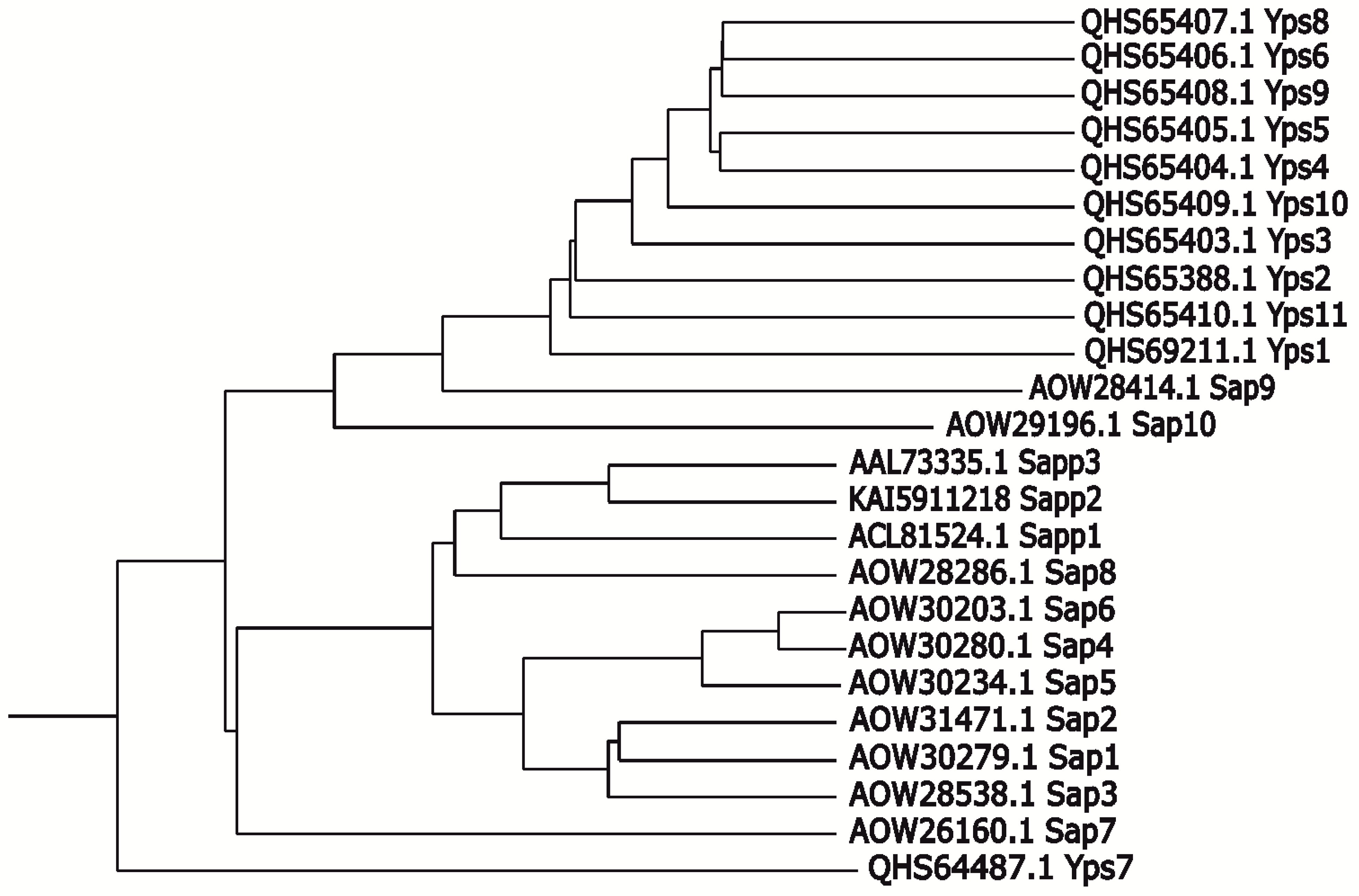
- Gagnon-Arsenault, I.; Tremblay, J.; Bourbonnais, Y. Fungal yapsins and cell wall: A unique family of aspartic peptidases for a distinctive cellular function. FEMS Yeast Res. 2006, 6, 966–978. [Google Scholar] [CrossRef]
- Rasheed, M.; Battu, A.; Kaur, R. Aspartyl proteases in Candida glabrata are required for suppression of the host innate immune response. J. Biol. Chem. 2018, 293, 6410–6433. [Google Scholar] [CrossRef] [Green Version]
- Boc, A.; Diallo, A.B.; Makarenkov, V. T-REX: A web server for inferring, validating and visualizing phylogenetic trees and networks. Nucleic Acids Res. 2012, 40, W573–W579. [Google Scholar] [CrossRef] [Green Version]
- Baldo, A.; Tabart, J.; Vermout, S.; Mathy, A.; Collard, A.; Losson, B.; Mignon, B. Secreted subtilisins of Microsporum canis are involved in adherence of arthroconidia to feline corneocytes. J. Med. Microbiol. 2008, 57, 1152–1156. [Google Scholar] [CrossRef] [Green Version]
- Baldo, A.; Mathy, A.; Tabart, J.; Camponova, P.; Vermout, S.; Massart, L.; Maréchal, F.; Galleni, M.; Mignon, B. Secreted subtilisin Sub3 from Microsporum canis is required for adherence to but not for invasion of the epidermis. Br. J. Dermatol. 2010, 162, 990–997. [Google Scholar] [CrossRef]
- Baldo, A.; Monod, M.; Mathy, A.; Cambier, L.; Bagut, E.T.; Defaweux, V.; Symoens, F.; Antoine, N.; Mignon, B. Mechanisms of skin adherence and invasion by dermatophytes. Mycoses 2012, 55, 218–223. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Gautam, P.; Pandit, H.; Singh, Y.; Basir, S.F.; Madan, T. Identification of fibrinogen-binding proteins of Aspergillus fumigatus using proteomic approach. Mycopathologia 2012, 173, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.Y.; Zhu, H.M.; Wu, J.H.; Wen, H.; Liu, C.J. Increased permeability of blood-brain barrier is mediated by serine protease during Cryptococcus meningitis. J. Int. Med. Res. 2014, 42, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Na Pombejra, S.; Jamklang, M.; Uhrig, J.P.; Vu, K.; Gelli, A. The structure-function analysis of the Mpr1 metalloprotease determinants of activity during migration of fungal cells across the blood-brain barrier. PLoS ONE 2018, 13, e0203020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mjokane, N.; Folorunso, O.S.; Ogundeji, A.O.; Sebolai, O.M. The Possible Role of Microbial Proteases in Facilitating SARS-CoV-2 Brain Invasion. Biology 2021, 10, 966. [Google Scholar] [CrossRef] [PubMed]
- Mjokane, N.; Maliehe, M.; Folorunso, O.S.; Ogundeji, A.O.; Gcilitshana, O.M.N.; Albertyn, J.; Pohl, C.H.; Sebolai, O.M. Cryptococcal Protease(s) and the Activation of SARS-CoV-2 Spike (S) Protein. Cells 2022, 11, 437. [Google Scholar] [CrossRef]
- Romani, L. Immunity to fungal infections. Nat. Rev. Immunol. 2004, 4, 11–23. [Google Scholar] [CrossRef]
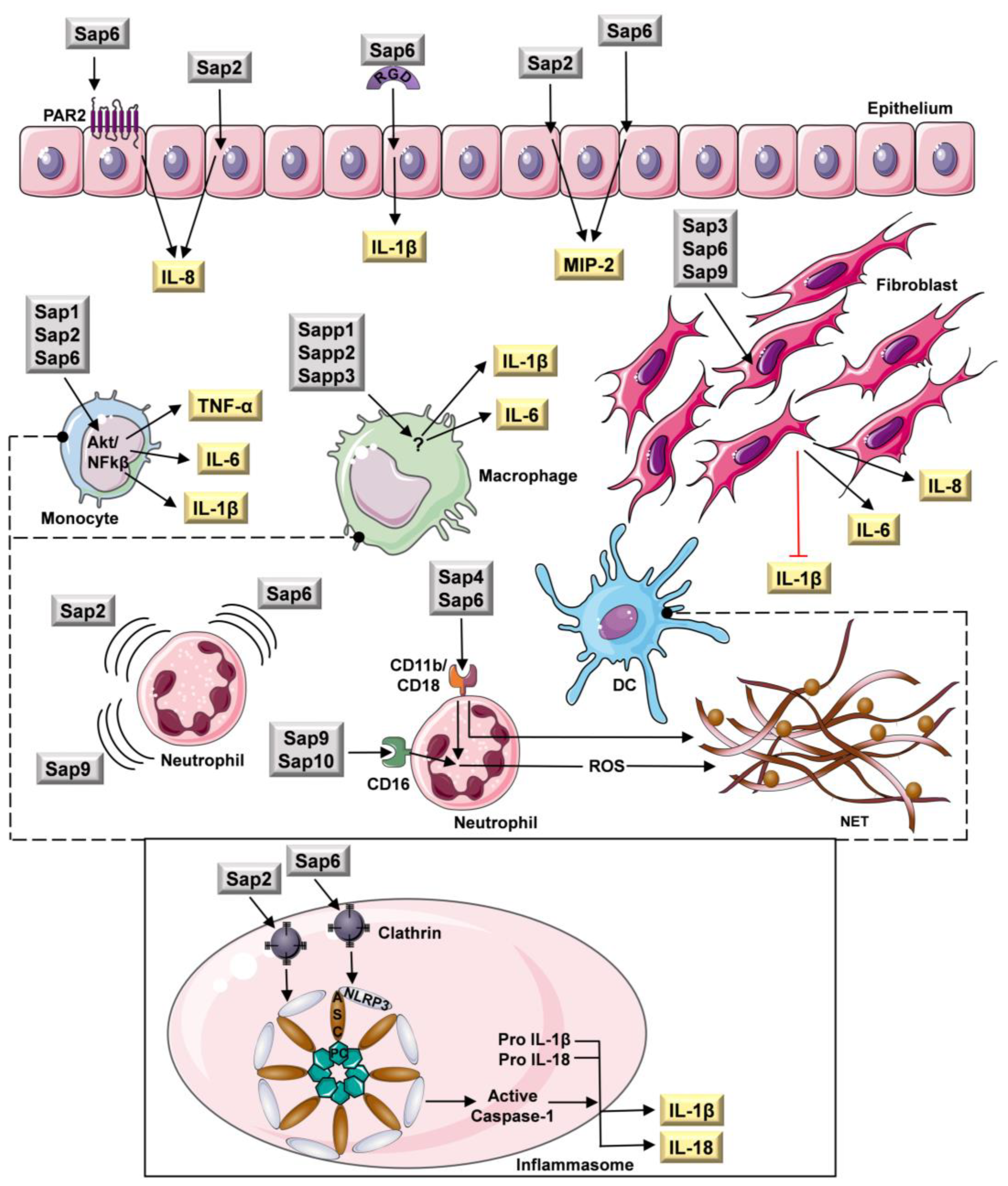
- Beauséjour, A.; Grenier, D.; Goulet, J.P.; Deslauriers, N. Proteolytic Activation of the Interleukin-1β Precursor by Candida albicans. Infect. Immun. 1998, 66, 676. [Google Scholar] [CrossRef] [Green Version]
- Schaller, M.; Borelli, C.; Korting, H.C.; Hube, B. Hydrolytic enzymes as virulence factors of Candida albicans. Mycoses 2005, 48, 365–377. [Google Scholar] [CrossRef]
- Jeong, S.K.; Kim, H.J.; Youm, J.K.; Ahn, S.K.; Choi, E.H.; Sohn, M.H.; Kim, K.E.; Hong, J.H.; Shin, D.M.; Lee, S.H. Mite and cockroach allergens activate protease-activated receptor 2 and delay epidermal permeability barrier recovery. J. Investig. Dermatol. 2008, 128, 1930–1939. [Google Scholar] [CrossRef] [Green Version]
- Lambrecht, B.N.; Hammad, H. Biology of lung dendritic cells at the origin of asthma. Immunity 2009, 31, 412–424. [Google Scholar] [CrossRef]
- Carey, R.M.; Freund, J.R.; Hariri, B.M.; Adappa, N.D.; Palmer, J.N.; Lee, R.J. Polarization of protease-activated receptor 2 (PAR-2) signaling is altered during airway epithelial remodeling and deciliation. J. Biol. Chem. 2020, 295, 6721–6740. [Google Scholar] [CrossRef]
- Reed, C.E.; Kita, H. The role of protease activation of inflammation in allergic respiratory diseases. J. Allergy Clin. Immunol. 2004, 114, 997–1008. [Google Scholar] [CrossRef]
- Pietrella, D.; Rachini, A.; Pandey, N.; Schild, L.; Netea, M.; Bistoni, F.; Hube, B.; Vecchiarelli, A. The Inflammatory response induced by aspartic proteases of Candida albicans is independent of proteolytic activity. Infect. Immun. 2010, 78, 4754–4762. [Google Scholar] [CrossRef] [Green Version]
- Ruoslahti, E. RGD and other recognition sequences for integrins. Annu. Rev. Cell Dev. Biol. 1996, 12, 697–715. [Google Scholar] [CrossRef]
- Kumar, R.; Rojas, I.G.; Edgerton, M. Candida albicans Sap6 Initiates Oral Mucosal Inflammation via the Protease Activated Receptor PAR2. Front. Immunol. 2022, 13, 912748. [Google Scholar] [CrossRef]
- Bartnicka, D.; Gonzalez-Gonzalez, M.; Sykut, J.; Koziel, J.; Ciaston, I.; Adamowicz, K.; Bras, G.; Zawrotniak, M.; Karkowska-Kuleta, J.; Satala, D.; et al. Candida albicans Shields the Periodontal Killer Porphyromonas gingivalis from Recognition by the Host Immune System and Supports the Bacterial Infection of Gingival Tissue. Int. J. Mol. Sci. 2020, 21, 1984. [Google Scholar] [CrossRef] [Green Version]
- Gabrielli, E.; Sabbatini, S.; Roselletti, E.; Kasper, L.; Perito, S.; Hube, B.; Cassone, A.; Vecchiarelli, A.; Pericolini, E. In vivo induction of neutrophil chemotaxis by secretory aspartyl proteinases of Candida albicans. Virulence 2016, 7, 819–825. [Google Scholar] [CrossRef] [Green Version]
- Ran, Y.; Iwabuchi, K.; Yamazaki, M.; Tsuboi, R.; Ogawa, H. Secreted aspartic proteinase from Candida albicans acts as a chemoattractant for peripheral neutrophils. J. Dermatol. Sci. 2013, 72, 191–193. [Google Scholar] [CrossRef]
- Hornbach, A.; Heyken, A.; Schild, L.; Hube, B.; Löffler, J.; Kurzai, O. The glycosylphosphatidylinositol-anchored protease Sap9 modulates the interaction of Candida albicans with human neutrophils. Infect. Immun. 2009, 77, 5216–5224. [Google Scholar] [CrossRef] [Green Version]
- Pericolini, E.; Gabrielli, E.; Amacker, M.; Kasper, L.; Roselletti, E.; Luciano, E.; Sabbatini, S.; Kaeser, M.; Moser, C.; Hube, B.; et al. Secretory Aspartyl Proteinases Cause Vaginitis and Can Mediate Vaginitis Caused by Candida albicans in Mice. MBio 2015, 6, e00724. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wang, H.; Kouadir, M.; Song, H.; Shi, F. Recent advances in the mechanisms of NLRP3 inflammasome activation and its inhibitors. Cell Death Dis. 2019, 10, 128. [Google Scholar] [CrossRef] [PubMed]
- Pietrella, D.; Pandey, N.; Gabrielli, E.; Pericolini, E.; Perito, S.; Kasper, L.; Bistoni, F.; Cassone, A.; Hube, B.; Vecchiarelli, A. Secreted aspartic proteases of Candida albicans activate the NLRP3 inflammasome. Eur. J. Immunol. 2013, 43, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Gabrielli, E.; Pericolini, E.; Luciano, E.; Sabbatini, S.; Roselletti, E.; Perito, S.; Kasper, L.; Hube, B.; Vecchiarelli, A. Induction of caspase-11 by aspartyl proteinases of Candida albicans and implication in promoting inflammatory response. Infect. Immun. 2015, 83, 1940–1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zawrotniak, M.; Bochenska, O.; Karkowska-Kuleta, J.; Seweryn-Ozog, K.; Aoki, W.; Ueda, M.; Kozik, A.; Rapala-Kozik, M. Aspartic Proteases and Major Cell Wall Components in Candida albicans Trigger the Release of Neutrophil Extracellular Traps. Front. Cell. Infect. Microbiol. 2017, 7, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krysan, D.J.; Ting, E.L.; Abeijon, C.; Kroos, L.; Fuller, R.S. Yapsins are a family of aspartyl proteases required for cell wall integrity in Saccharomyces cerevisiae. Eukaryot. Cell 2005, 4, 1364–1374. [Google Scholar] [CrossRef] [Green Version]
- Bairwa, G.; Kaur, R. A novel role for a glycosylphosphatidylinositol-anchored aspartyl protease, CgYps1, in the regulation of pH homeostasis in Candida glabrata. Mol. Microbiol. 2011, 79, 900–913. [Google Scholar] [CrossRef]
- Askari, F.; Rasheed, M.; Kaur, R. The yapsin family of aspartyl proteases regulate glucose homeostasis in Candida glabrata. J. Biol. Chem. 2022, 298, 101593. [Google Scholar] [CrossRef]
- Kasper, L.; Seider, K.; Hube, B. Intracellular survival of Candida glabrata in macrophages: Immune evasion and persistence. FEMS Yeast Res. 2015, 15, fov042. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.D.; Taylor, P.R.; Reid, D.M.; Willment, J.A.; Williams, D.L.; Martinez-Pomares, L.; Wong, S.Y.C.; Gordon, S. Dectin-1 is a major beta-glucan receptor on macrophages. J. Exp. Med. 2002, 196, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.M.; Shen, H.; Zhang, T.; Huang, X.; Liu, X.Q.; Guo, S.Y.; Zhao, J.J.; Wang, C.F.; Yan, L.; Xu, G.T.; et al. Dectin-1 plays an important role in host defense against systemic Candida glabrata infection. Virulence 2017, 8, 1643–1656. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, Y.; Tsai, H.F.; Myers, T.G.; Bennett, J.E. Transcriptional profiling of Candida glabrata during phagocytosis by neutrophils and in the infected mouse spleen. Infect. Immun. 2013, 81, 1325–1333. [Google Scholar] [CrossRef]
- Gross, O.; Poeck, H.; Bscheider, M.; Dostert, C.; Hannesschläger, N.; Endres, S.; Hartmann, G.; Tardivel, A.; Schweighoffer, E.; Tybulewicz, V.; et al. Syk kinase signalling couples to the Nlrp3 inflammasome for anti-fungal host defence. Nature 2009, 459, 433–436. [Google Scholar] [CrossRef]
- Tucey, T.M.; Verma, J.; Harrison, P.F.; Snelgrove, S.L.; Lo, T.L.; Scherer, A.K.; Barugahare, A.A.; Powell, D.R.; Wheeler, R.T.; Hickey, M.J.; et al. Glucose Homeostasis Is Important for Immune Cell Viability during Candida Challenge and Host Survival of Systemic Fungal Infection. Cell Metab. 2018, 27, 988–1006.e7. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.; Askari, F.; Sahu, M.S.; Kaur, R. Candida glabrata: A Lot More Than Meets the Eye. Microorganisms 2019, 7, 39. [Google Scholar] [CrossRef] [Green Version]
- Buu, L.M.; Chen, Y.C. Sap6, a secreted aspartyl proteinase, participates in maintenance the cell surface integrity of Candida albicans. J. Biomed. Sci. 2013, 20, 101. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Tao, A. Antigenicity, Immunogenicity, Allergenicity. Allergy Bioinforma. 2015, 8, 175. [Google Scholar] [CrossRef]
- Yike, I. Fungal proteases and their pathophysiological effects. Mycopathologia 2011, 171, 299–323. [Google Scholar] [CrossRef]
- Rick, E.M.; Woolnough, K.; Pashley, C.H.; Wardlaw, A.J. Allergic Fungal Airway Disease. J. Investig. Allergol. Clin. Immunol. 2016, 26, 344–354. [Google Scholar] [CrossRef] [Green Version]
- Furlong-Silva, J.; Cook, P.C. Fungal-mediated lung allergic airway disease: The critical role of macrophages and dendritic cells. PLoS Pathog. 2022, 18, e1010608. [Google Scholar] [CrossRef]
- Kay, A.B. Allergy and allergic diseases. First of two parts. N. Engl. J. Med. 2001, 344, 30–37. [Google Scholar] [CrossRef]
- Galli, S.J.; Tsai, M.; Piliponsky, A.M. The development of allergic inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.; Lambrecht, B.N. Dendritic cells and airway epithelial cells at the interface between innate and adaptive immune responses. Allergy 2011, 66, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Busse, W.W.; Lemanske, R.F. Asthma. N. Engl. J. Med. 2001, 344, 350–362. [Google Scholar] [CrossRef] [PubMed]
- Bush, R.K. Indoor allergens, environmental avoidance, and allergic respiratory disease. Allergy Asthma Proc. 2008, 29, 575–579. [Google Scholar] [CrossRef] [PubMed]
- GBD 2019 Diseases and Injuries Collaborators. Global burden of 369 diseases and injuries in 204 countries and territories, 1990-2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222, Erratum: Department of Error (The Lancet (2020) 396(10258) (1204–1222), (S0140673620309259), (10.1016/S0140-6736(20)30925-9)). Lancet 2020, 396, 1562. https://doi.org/10.1016/S0140-6736(20)32226-1. [Google Scholar] [CrossRef]
- Robinson, B.W.S.; Venaille, T.J.; Mendis, A.H.W.; McAleer, R. Allergens as proteases: An Aspergillus fumigatus proteinase directly induces human epithelial cell detachment. J. Allergy Clin. Immunol. 1990, 86, 726–731. [Google Scholar] [CrossRef]
- Chen, J.C.; Chuang, J.G.; Su, Y.Y.; Chiang, B.L.; Lin, Y.S.; Chow, L.P. The protease allergen Pen c 13 induces allergic airway inflammation and changes in epithelial barrier integrity and function in a murine model. J. Biol. Chem. 2011, 286, 26667–26679. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, Y. Role of Allergen Source-Derived Proteases in Sensitization via Airway Epithelial Cells. J. Allergy 2012, 2012, 903659. [Google Scholar] [CrossRef] [Green Version]
- Kauffman, H.F.; Christomee, J.F.; Van De Riet, M.A.; Timmerman, A.J.B.; Borger, P. Protease-dependent activation of epithelial cells by fungal allergens leads to morphologic changes and cytokine production. J. Allergy Clin. Immunol. 2000, 105, 1185–1193. [Google Scholar] [CrossRef]
- Shen, H.D.; Tam, M.F.; Tang, R.B.; Chou, H. Aspergillus and Penicillium allergens: Focus on proteases. Curr. Allergy Asthma Rep. 2007, 7, 351–356. [Google Scholar] [CrossRef]
- Boitano, S.; Flynn, A.N.; Sherwood, C.L.; Schulz, S.M.; Hoffman, J.; Gruzinova, I.; Daines, M.O. Alternaria alternata serine proteases induce lung inflammation and airway epithelial cell activation via PAR2. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 300, L605–L614. [Google Scholar] [CrossRef]
- Tiwary, M.; Samarasinghe, A.E. Initiation and Pathogenesis of Severe Asthma with Fungal Sensitization. Cells 2021, 10, 913. [Google Scholar] [CrossRef]
- Pomés, A.; Davies, J.M.; Gadermaier, G.; Hilger, C.; Holzhauser, T.; Lidholm, J.; Lopata, A.L.; Mueller, G.A.; Nandy, A.; Radauer, C.; et al. WHO/IUIS Allergen Nomenclature: Providing a common language. Mol. Immunol. 2018, 100, 3–13. [Google Scholar] [CrossRef]
- WHO/IUIS Allergen Nomenclature Home Page. Available online: http://www.allergen.org/index.php (accessed on 27 December 2022).
- Redes, J.L.; Basu, S.; Ram-Mohan, S.; Ghosh, C.C.; Chan, E.C.; Sek, A.C.; Zhao, M.; Krishnan, R.; Rosenberg, H.F.; Druey, K.M. Aspergillus fumigatus–Secreted Alkaline Protease 1 Mediates Airways Hyperresponsiveness in Severe Asthma. ImmunoHorizons 2019, 3, 368–377. [Google Scholar] [CrossRef]
- Blango, M.G.; Pschibul, A.; Rivieccio, F.; Krüger, T.; Rafiq, M.; Jia, L.J.; Zheng, T.; Goldmann, M.; Voltersen, V.; Li, J.; et al. Dynamic Surface Proteomes of Allergenic Fungal Conidia. J. Proteome Res. 2020, 19, 2092–2104. [Google Scholar] [CrossRef]
- Rowley, J.; Namvar, S.; Gago, S.; Labram, B.; Bowyer, P.; Richardson, M.D.; Herrick, S.E. Differential Proinflammatory Responses to Aspergillus fumigatus by Airway Epithelial Cells In Vitro Are Protease Dependent. J. Fungi 2021, 7, 468. [Google Scholar] [CrossRef]
- Behnsen, J.; Lessing, F.; Schindler, S.; Wartenberg, D.; Jacobsen, I.D.; Thoen, M.; Zipfel, P.F.; Brakhage, A.A. Secreted Aspergillus fumigatus protease Alp1 degrades human complement proteins C3, C4, and C5. Infect. Immun. 2010, 78, 3585–3594. [Google Scholar] [CrossRef] [Green Version]
- Balenga, N.A.; Klichinsky, M.; Xie, Z.; Chan, E.C.; Zhao, M.; Jude, J.; Laviolette, M.; Panettieri, R.A.; Druey, K.M. A fungal protease allergen provokes airway hyper-responsiveness in asthma. Nat. Commun. 2015, 6, 6763. [Google Scholar] [CrossRef] [Green Version]
- Wiesner, D.L.; Merkhofer, R.M.; Ober, C.; Kujoth, G.C.; Niu, M.; Keller, N.P.; Gern, J.E.; Brockman-Schneider, R.A.; Evans, M.D.; Jackson, D.J.; et al. Club Cell TRPV4 Serves as a Damage Sensor Driving Lung Allergic Inflammation. Cell Host Microbe 2020, 27, 614–628.e6. [Google Scholar] [CrossRef]
- Druey, K.M.; McCullough, M.; Krishnan, R. Aspergillus fumigatus Protease Alkaline Protease 1 (Alp1): A New Therapeutic Target for Fungal Asthma. J. Fungi 2020, 6, 88. [Google Scholar] [CrossRef]
- Chiu, L.-L.; Perng, D.-W.; Yu, C.-H.; Su, S.-N.; Chow, L.-P. Mold allergen, pen C 13, induces IL-8 expression in human airway epithelial cells by activating protease-activated receptor 1 and 2. J. Immunol. 2007, 178, 5237–5244. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.; Lai, H.Y.; Tam, M.F.; Chou, M.Y.; Wang, S.R.; Han, S.H.; Shen, H. Der cDNA cloning, biological and immunological characterization of the alkaline serine protease major allergen from Penicillium chrysogenum. Int. Arch. Allergy Immunol. 2002, 127, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Tai, H.Y.; Tam, M.F.; Chou, H.; Peng, H.J.; Su, S.N.; Perng, D.W.; Shen, H.D. Pen ch 13 allergen induces secretion of mediators and degradation of occludin protein of human lung epithelial cells. Allergy 2006, 61, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Denis, O.; Vincent, M.; Havaux, X.; De Prins, S.; Treutens, G.; Huygen, K. Induction of the specific allergic immune response is independent of proteases from the fungus Alternaria alternata. Eur. J. Immunol. 2013, 43, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Daines, M.; Zhu, L.; Pereira, R.; Zhou, X.; Bondy, C.; Pryor, B.M.; Zhou, J.; Chen, Y. Alternaria induces airway epithelial cytokine expression independent of protease-activated receptor. Respirology 2020, 25, 502–510. [Google Scholar] [CrossRef]
- Rivas, C.M.; Schiff, H.V.; Moutal, A.; Khanna, R.; Kiela, P.R.; Dussor, G.; Price, T.J.; Vagner, J.; DeFea, K.A.; Boitano, S. Alternaria alternata-induced airway epithelial signaling and inflammatory responses via protease-activated receptor-2 expression. Biochem. Biophys. Res. Commun. 2022, 591, 13–19. [Google Scholar] [CrossRef]
- Sircar, G.; Saha, B.; Mandal, R.S.; Pandey, N.; Saha, S.; Bhattacharya, S.G. Purification, Cloning and Immuno-Biochemical Characterization of a Fungal Aspartic Protease Allergen Rhi o 1 from the Airborne Mold Rhizopus oryzae. PLoS ONE 2015, 10, e0144547. [Google Scholar] [CrossRef] [Green Version]
- Monod, M. Secreted proteases from dermatophytes. Mycopathologia 2008, 166, 285–294. [Google Scholar] [CrossRef] [Green Version]
- Woodfolk, J.A. Allergy and dermatophytes. Clin. Microbiol. Rev. 2005, 18, 30–43. [Google Scholar] [CrossRef] [Green Version]
- Descamps, F.F.; Brouta, F.; Vermout, S.M.; Willame, C.; Losson, B.J.; Mignon, B.R. A recombinant 31.5 kDa keratinase and a crude exo-antigen from Microsporum canis fail to protect against a homologous experimental infection in guinea pigs. Vet. Dermatol. 2003, 14, 305–312. [Google Scholar] [CrossRef]
- Descamps, F.; Brouta, F.; Vermout, S.; Monod, M.; Losson, B.; Mignon, B. Recombinant expression and antigenic properties of a 31.5-kDa keratinolytic subtilisin-like serine protease from Microsporum canis. FEMS Immunol. Med. Microbiol. 2003, 38, 29–34. [Google Scholar] [CrossRef]
- Brouta, F.; Descamps, F.; Vermout, S.; Monod, M.; Losson, B.; Mignon, B. Humoral and cellular immune response to a Microsporum canis recombinant keratinolytic metalloprotease (r-MEP3) in experimentally infected guinea pigs. Med. Mycol. 2003, 41, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Vermout, S.M.; Brouta, F.D.; Descamps, F.F.; Losson, B.J.; Mignon, B.R. Evaluation of immunogenicity and protective efficacy of a Microsporum canis metalloprotease subunit vaccine in guinea pigs. FEMS Immunol. Med. Microbiol. 2004, 40, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Goh, J.P.Z.; Ruchti, F.; Poh, S.E.; Koh, W.L.C.; Tan, K.Y.; Lim, Y.T.; Thng, S.T.G.; Sobota, R.M.; Hoon, S.S.; Liu, C.; et al. The human pathobiont Malassezia furfur secreted protease Mfsap1 regulates cell dispersal and exacerbates skin inflammation. Proc. Natl. Acad. Sci. USA 2022, 119, e2212533119. [Google Scholar] [CrossRef]
- de Oliveira, P.; Juliano, M.A.; Tanaka, A.S.; Carmona, A.K.; dos Santos, S.M.B.; de Barros, B.C.S.C.; Maza, P.K.; Puccia, R.; Suzuki, E. Paracoccidioides brasiliensis induces cytokine secretion in epithelial cells in a protease-activated receptor-dependent (PAR) manner. Med. Microbiol. Immunol. 2017, 206, 149–156. [Google Scholar] [CrossRef]
- Moreira, A.L.E.; Oliveira, M.A.P.; Silva, L.O.S.; Inácio, M.M.; Bailão, A.M.; Parente-Rocha, J.A.; Cruz-Leite, V.R.M.; Paccez, J.D.; de Almeida Soares, C.M.; Weber, S.S.; et al. Immunoproteomic Approach of Extracellular Antigens From Paracoccidioides Species Reveals Exclusive B-Cell Epitopes. Front. Microbiol. 2020, 10, 2968. [Google Scholar] [CrossRef]
- Toyotome, T.; Watanabe, A.; Ochiai, E.; Kamei, K. N-acetylated α-linked acidic dipeptidase is identified as an antigen of Histoplasma capsulatum. Biochem. Biophys. Res. Commun. 2015, 458, 483–487. [Google Scholar] [CrossRef]
- Rosa, D.D.A.; Gezuele, E.; Calegari, L.; Goñi, F. Excretion-secretion products and proteases from live Sporothrix schenckii yeast phase: Immunological detection and cleavage of human IgG. Rev. Inst. Med. Trop. Sao Paulo 2009, 51, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kling, H.M.; Norris, K.A. Vaccine-Induced Immunogenicity and Protection Against Pneumocystis Pneumonia in a Nonhuman Primate Model of HIV and Pneumocystis Coinfection. J. Infect. Dis. 2016, 213, 1586–1595. [Google Scholar] [CrossRef] [Green Version]
- Tomás, A.L.; Cardoso, F.; de Sousa, B.; Matos, O. Detection of anti-Pneumocystis jirovecii antibodies in human serum using a recombinant synthetic multi-epitope kexin-based antigen. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 2205–2209. [Google Scholar] [CrossRef]
- Rayens, E.; Noble, B.; Vicencio, A.; Goldman, D.L.; Bunyavanich, S.; Norris, K.A. Relationship of Pneumocystis antibody responses to paediatric asthma severity. BMJ open Respir. Res. 2021, 8, e000842. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zeng, Z.; Guo, Y.; Song, L.; Weatherhead, J.E.; Huang, X.; Zeng, Y.; Bimler, L.; Chang, C.Y.; Knight, J.M.; et al. Candida albicans elicits protective allergic responses via platelet mediated T helper 2 and T helper 17 cell polarization. Immunity 2021, 54, 2595–2610.e7. [Google Scholar] [CrossRef] [PubMed]
- Sandini, S.; La Valle, R.; Deaglio, S.; Malavasi, F.; Cassone, A.; De Bernardis, F. A highly immunogenic recombinant and truncated protein of the secreted aspartic proteases family (rSap2t) of Candida albicans as a mucosal anticandidal vaccine. FEMS Immunol. Med. Microbiol. 2011, 62, 215–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, M.; Chandley, P.; Rohatgi, S. The Role of B-Cells and Antibodies against Candida Vaccine Antigens in Invasive Candidiasis. Vaccines 2021, 9, 1159. [Google Scholar] [CrossRef] [PubMed]
- Shukla, M.; Rohatgi, S. Vaccination with Secreted Aspartyl Proteinase 2 Protein from Candida parapsilosis Can Enhance Survival of Mice during C. tropicalis-Mediated Systemic Candidiasis. Infect. Immun. 2020, 88, e00312-20. [Google Scholar] [CrossRef]
- Miyazaki, T.; Izumikawa, K.; Yamauchi, S.; Inamine, T.; Nagayoshi, Y.; Saijo, T.; Seki, M.; Kakeya, H.; Yamamoto, Y.; Yanagihara, K.; et al. The glycosylphosphatidylinositol-linked aspartyl protease Yps1 is transcriptionally regulated by the calcineurin-Crz1 and Slt2 MAPK pathways in Candida glabrata. FEMS Yeast Res. 2011, 11, 449–456.e7. [Google Scholar] [CrossRef] [Green Version]
- Bairwa, G.; Rasheed, M.; Taigwal, R.; Sahoo, R.; Kaur, R. GPI (glycosylphosphatidylinositol)-linked aspartyl proteases regulate vacuole homoeostasis in Candida glabrata. Biochem. J. 2014, 458, 323–334. [Google Scholar] [CrossRef]
- Clarke, S.C.; Dumesic, P.A.; Homer, C.M.; O’Donoghue, A.J.; La Greca, F.; Pallova, L.; Majer, P.; Madhani, H.D.; Craik, C.S. Integrated Activity and Genetic Profiling of Secreted Peptidases in Cryptococcus neoformans Reveals an Aspartyl Peptidase Required for Low pH Survival and Virulence. PLoS Pathog. 2016, 12, e1006051. [Google Scholar] [CrossRef] [Green Version]
- Levitz, S.M.; Nong, S.H.; Seetoo, K.F.; Harrison, T.S.; Speizer, R.A.; Simons, E.R. Cryptococcus neoformans resides in an acidic phagolysosome of human macrophages. Infect. Immun. 1999, 67, 885–890. [Google Scholar] [CrossRef] [Green Version]
- King, J.; Henriet, S.S.V.; Warris, A. Aspergillosis in Chronic Granulomatous Disease. J. Fungi 2016, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Parsania, C.; Tan, K.; Todd, R.B.; Wong, K.H. Co-option of an extracellular protease for transcriptional control of nutrient degradation in the fungus Aspergillus nidulans. Commun. Biol. 2021, 4, 1409. [Google Scholar] [CrossRef]
- Zhao, X.; Hume, S.L.; Johnson, C.; Thompson, P.; Huang, J.; Gray, J.; Lamb, H.K.; Hawkins, A.R. The transcription repressor NmrA is subject to proteolysis by three Aspergillus nidulans proteases. Protein Sci. 2010, 19, 1405–1419. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.C.; Padovan, A.C.B.; Pimenta, D.C.; Ferreira, R.C.; da Silva, C.V.; Briones, M.R.S. Extracellular enolase of Candida albicans is involved in colonization of mammalian intestinal epithelium. Front. Cell. Infect. Microbiol. 2014, 4, 66. [Google Scholar] [CrossRef] [Green Version]
- Feng, W.; Yang, J.; Pan, Y.; Xi, Z.; Qiao, Z.; Ma, Y. The correlation of virulence, pathogenicity, and itraconazole resistance with SAP activity in Candida albicans strains. Can. J. Microbiol. 2016, 62, 173–178. [Google Scholar] [CrossRef]
- Kadry, A.A.; El-Ganiny, A.M.; El-Baz, A.M. Relationship between Sap prevalence and biofilm formation among resistant clinical isolates of Candida albicans. Afr. Health Sci. 2018, 18, 1166–1174. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Fungal Species | Identified Proteases | Protease Classification |
|---|---|---|
| Candida spp. | ||
| C. albicans | Sap1-Sap10 | aspartic proteases |
| C. glabrata | yapsins (CgYps1-11) | aspartic proteases |
| C. parapsilosis | Sapp1, Sapp2 | aspartic proteases |
| Aspergillus spp. | ||
| secreted subtilisins (i.e., Alp1/Asp f 13) | serine proteases | |
| elastinolytic proteases Mep, fungalysins (i.e., Asp f 5) | metalloproteases | |
| secreted aspartyl endopeptidase (i.e., aspergillopepsin-1/ Asp f 10) | aspartic proteases | |
| protease PnmB | serine protease | |
| Penicillium spp. | ||
| secreted subtilisin-like proteases (i.e., Pen c 13, Pen ch 13) | serine proteases | |
| Cryptococcus neoformans | ||
| major aspartyl peptidase 1 May1 | aspartic protease | |
| endopeptidase Prb1 | serine protease | |
| secreted metalloprotease Mpr1 | metalloprotease | |
| Pneumocystis jiroveci | kexin (Kex1) | serine protease |
| Rhizopus oryzae | rhizopuspepsin, Rhi o 1 | aspartic protease |
| Malassezia furfur | secretory aspartyl protease Mfsap1 | aspartic protease |
| Dermatophytes | ||
| secreted subtilisins (Sub2–Sub7) | serine proteases | |
| fungalysins (Mep1, Mep3, Mep4) | metalloproteases | |
| dipeptidyl peptidases (DppIV, DppV) | serine exoproteases | |
| Endemic fungi | ||
| Paracoccidioides brasiliensis | extracellular subtilisin-like protease (PbSP) | serine protease |
| Histoplasma capsulatum | N-acetylated α-linked acidic dipeptidase (NAALADase) | metallocarboxypeptidase |
| Sporothrix schenckii | proteinase I | serine protease |
| proteinase II | cysteine protease |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Satala, D.; Bras, G.; Kozik, A.; Rapala-Kozik, M.; Karkowska-Kuleta, J. More than Just Protein Degradation: The Regulatory Roles and Moonlighting Functions of Extracellular Proteases Produced by Fungi Pathogenic for Humans. J. Fungi 2023, 9, 121. https://doi.org/10.3390/jof9010121
Satala D, Bras G, Kozik A, Rapala-Kozik M, Karkowska-Kuleta J. More than Just Protein Degradation: The Regulatory Roles and Moonlighting Functions of Extracellular Proteases Produced by Fungi Pathogenic for Humans. Journal of Fungi. 2023; 9(1):121. https://doi.org/10.3390/jof9010121
Chicago/Turabian StyleSatala, Dorota, Grazyna Bras, Andrzej Kozik, Maria Rapala-Kozik, and Justyna Karkowska-Kuleta. 2023. "More than Just Protein Degradation: The Regulatory Roles and Moonlighting Functions of Extracellular Proteases Produced by Fungi Pathogenic for Humans" Journal of Fungi 9, no. 1: 121. https://doi.org/10.3390/jof9010121