

Biofilm Formation by Chromoblastomycosis Fungi Fonsecaea pedrosoi and Phialophora verrucosa: Involvement with Antifungal Resistance
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Fungi and Growth Conditions
2.2. Adhesion and Germination Capability of Conidial Cells to Polystyrene Surface
2.3. Biofilm Formation Detection on the Polystyrene Surface
2.4. Confocal Laser Scanning Microscopy (CLSM) Analysis
2.5. Scanning Electron Microscopy (SEM) Assay
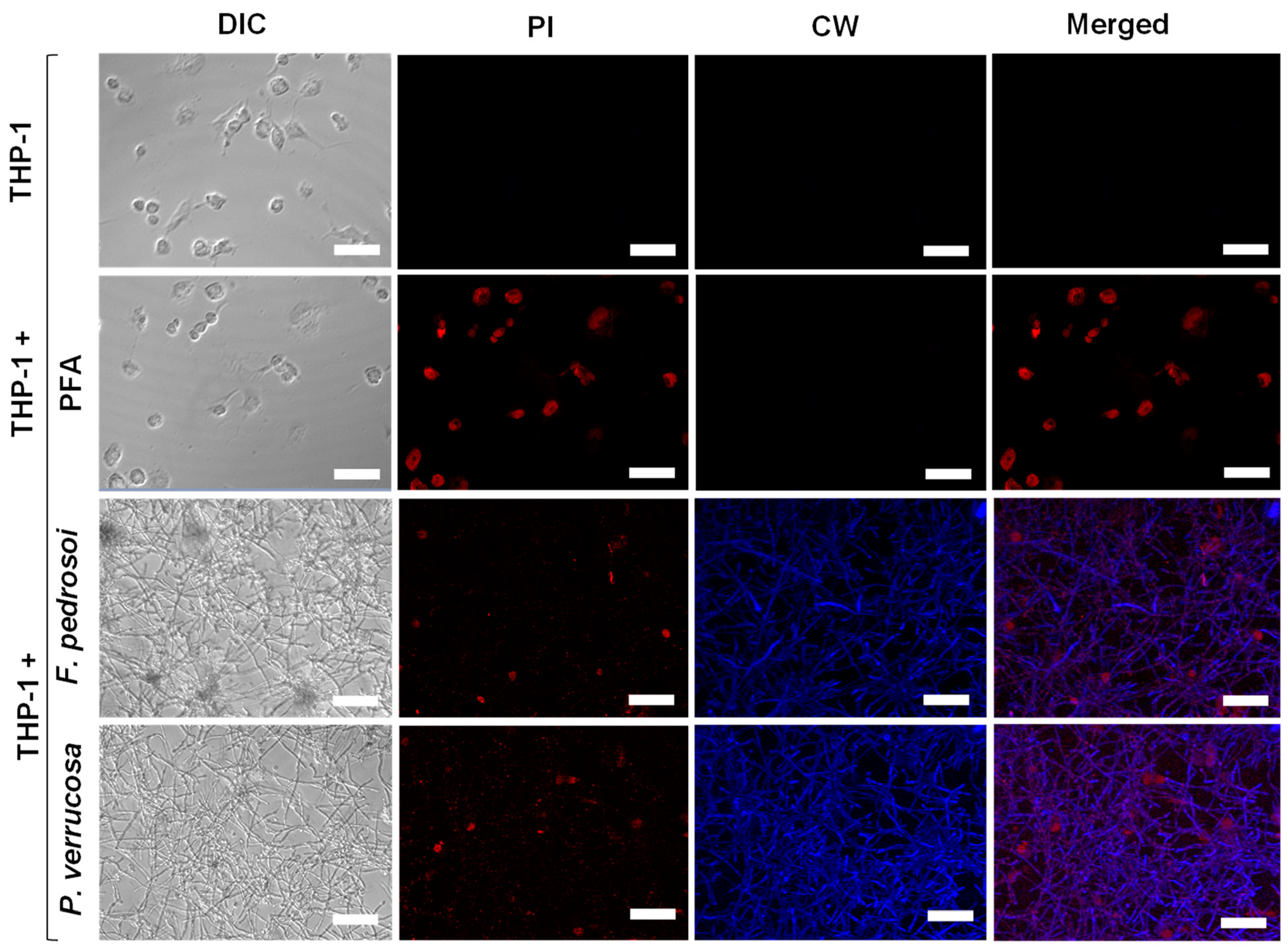
2.6. Biofilm Formation on THP-1 Cells
2.7. Antifungal Susceptibility Testing
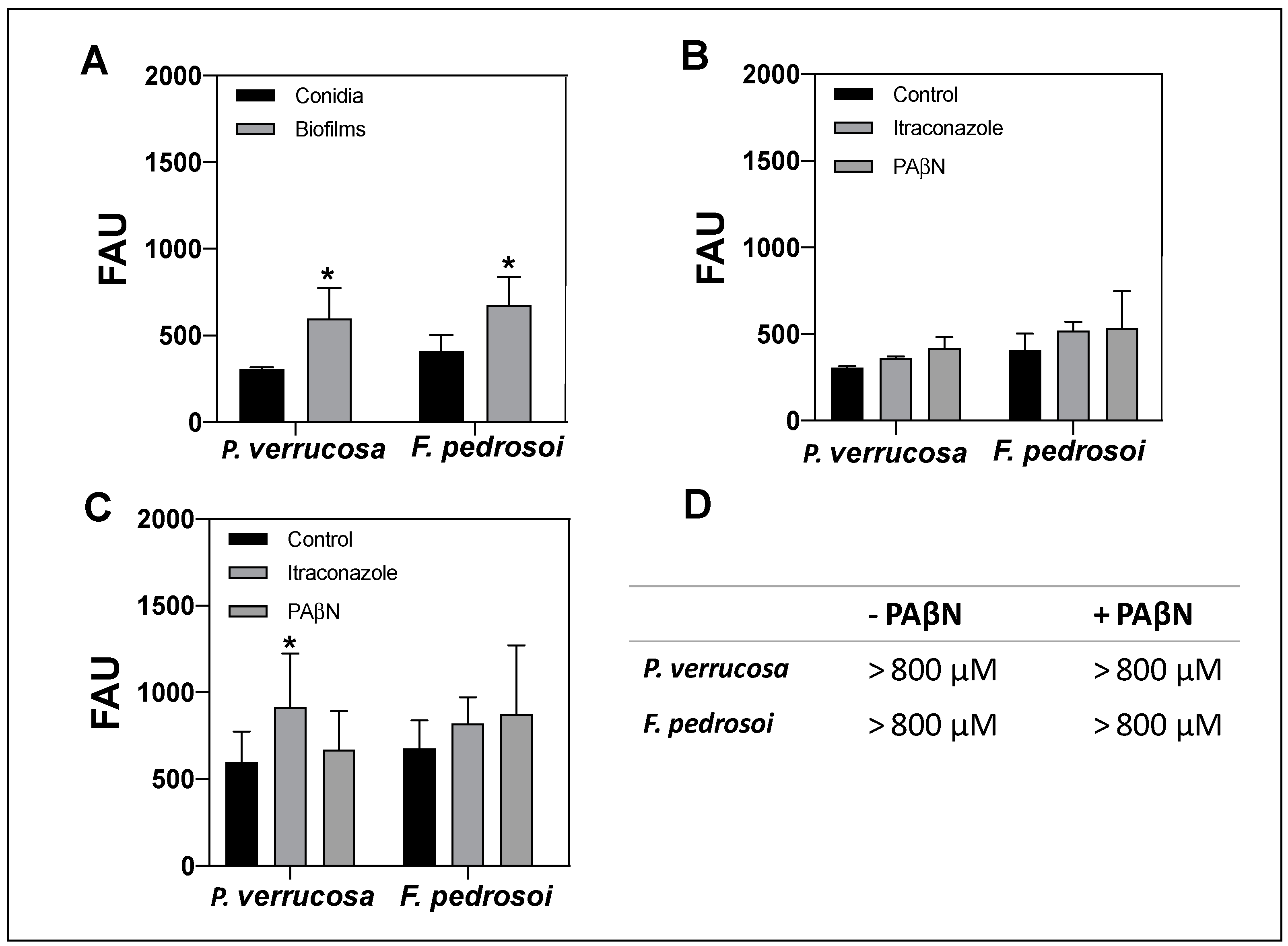
2.8. Activity of ABC Efflux Pumps on Conidial- and Biofilm-Forming Cells
2.9. Statistical Analysis
3. Results and Discussion
3.1. Adhesion and Germination of Fungi on Polystyrene Substrate
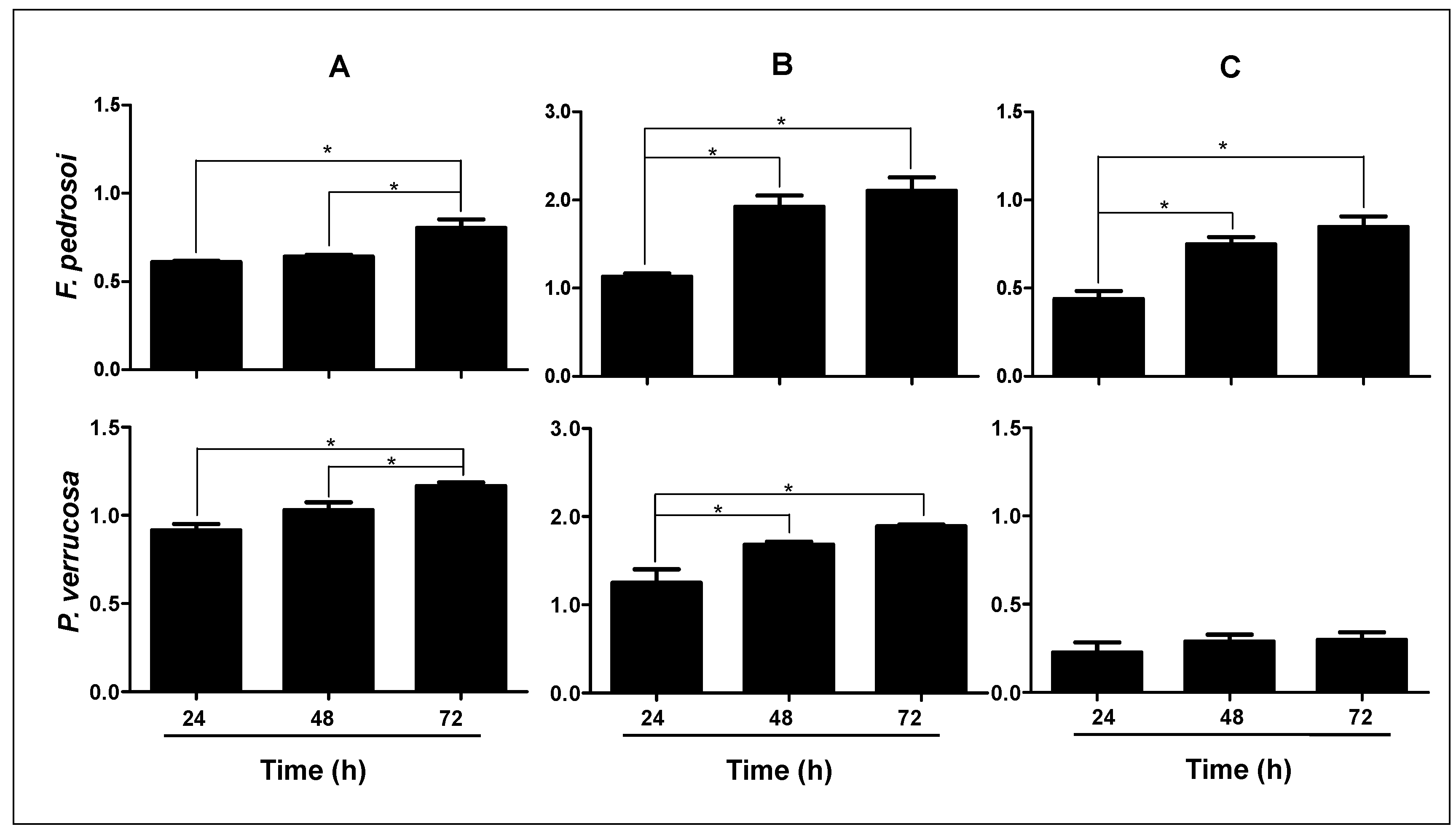
3.2. Measurement of Classical Biofilm Parameters
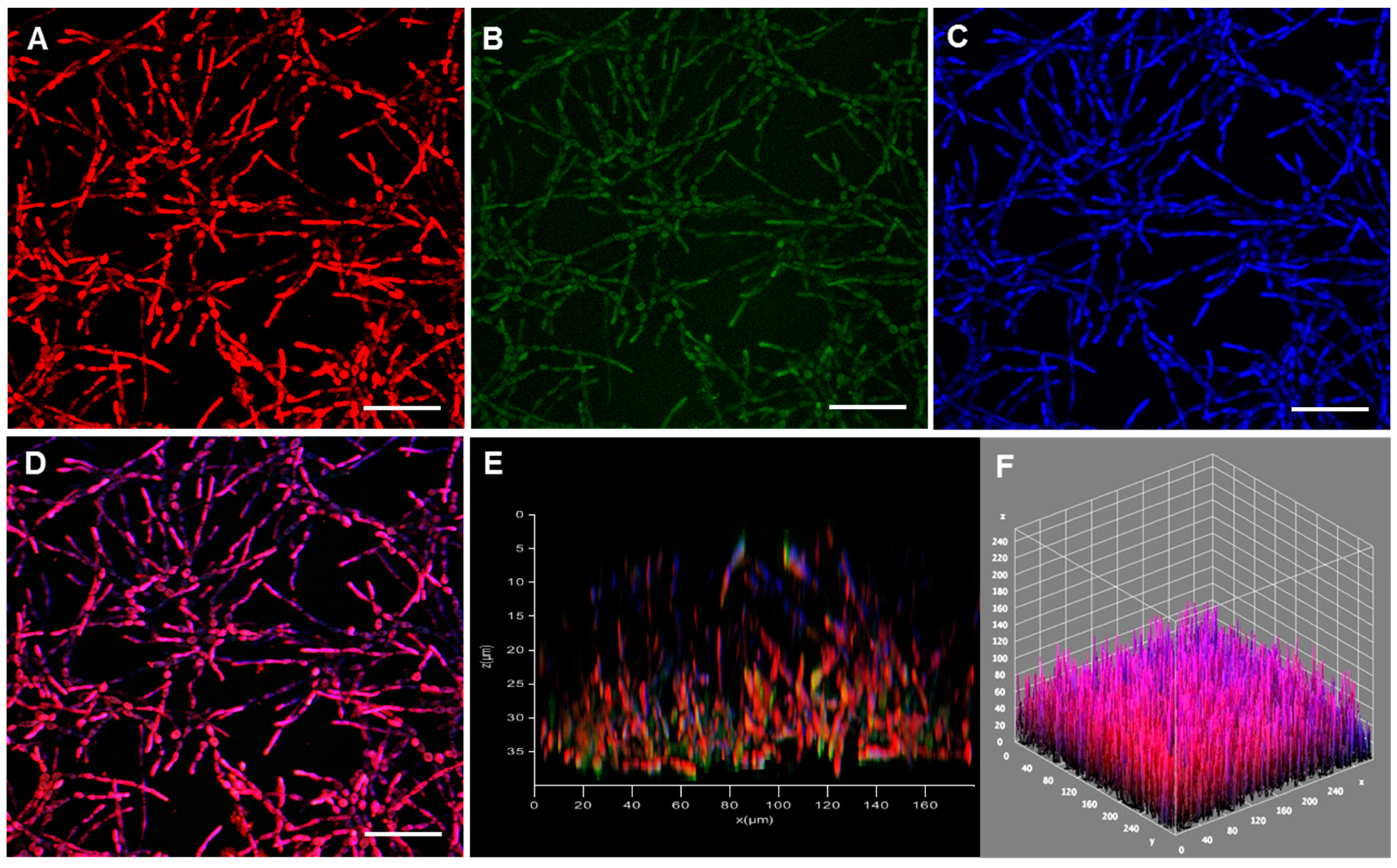
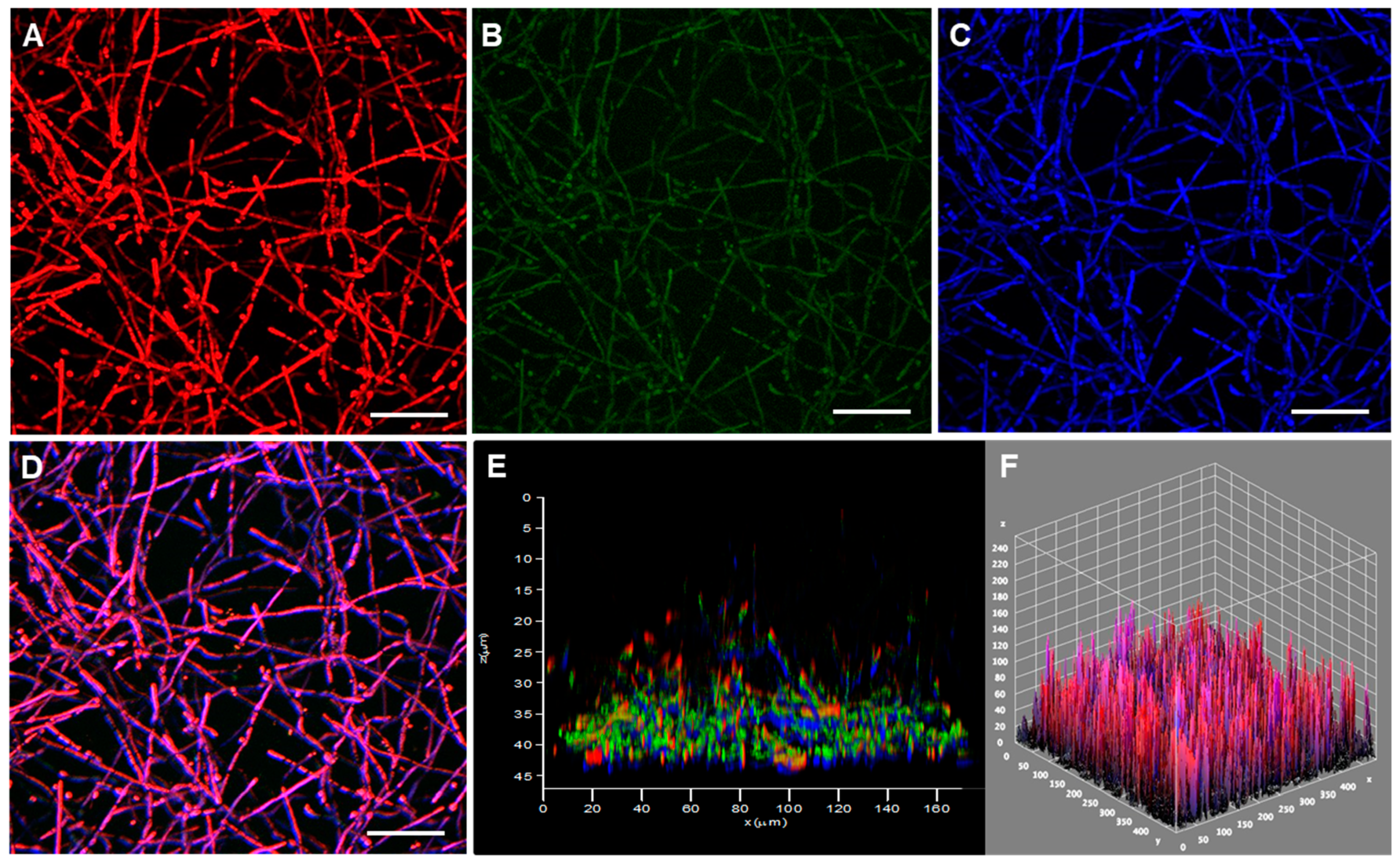
3.3. Fungal Biofilm Structural Distribution and 3-D Organization
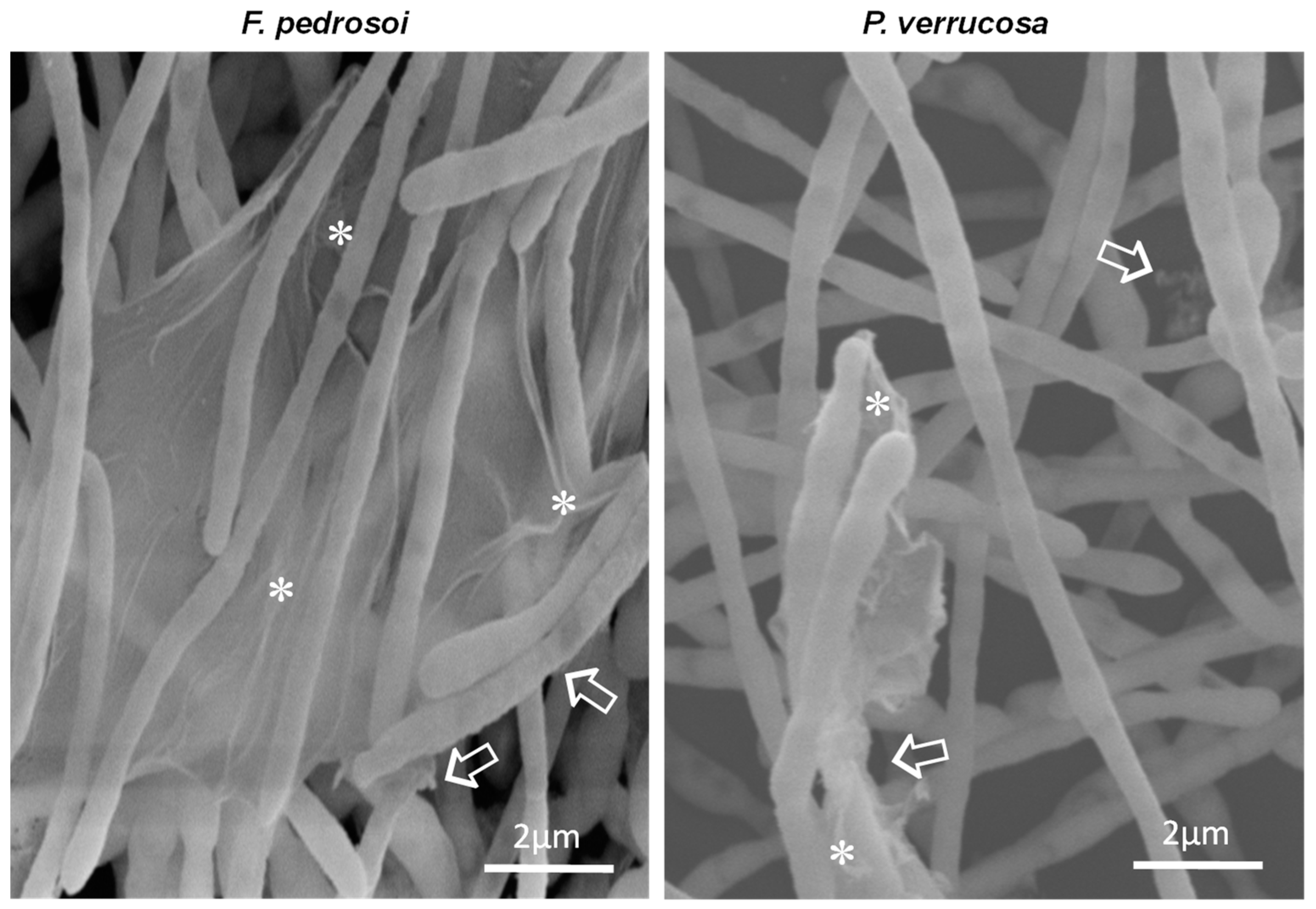
3.4. Ultrastructure of Biofilm-Growing Cells
3.5. Biofilm Formation on Animal Cells
3.6. Susceptibility of Planktonic and Biofilm Cells to Antifungal Agents
3.7. Activity of Efflux Pumps on Conidia and Biofilm-Forming Cells
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Queiroz-Telles, F.; de Hoog, S.; Santos, D.W.C.L.; Salgado, C.G.; Vicente, V.A.; Bonifaz, A.; Roilides, E.; Xi, L.; Azevedo, C.M.P.S.; da Silva, M.B.; et al. Chromoblastomycosis. Clin. Microbiol. Rev. 2017, 30, 233–276. [Google Scholar] [CrossRef] [PubMed]
- Torres-Guerrero, E.; Isa-Isa, R.; Isa, M.; Arenas, R. Chromoblastomycosis. Clin. Dermatol. 2012, 30, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Guevara, A.; Siqueira, N.P.; Nery, A.F.; Cavalcante, L.R.S.; Hagen, F.; Hahn, R.C. Chromoblastomycosis in Latin America and the Caribbean: Epidemiology over the past 50 years. Med. Mycol. 2021, 60, myab062. [Google Scholar] [CrossRef]
- Santos, D.W.C.L.; de Azevedo, C.M.P.S.; Vicente, V.A.; Queiroz-Telles, F.; Rodrigues, A.M.; de Hoog, G.S.; Denning, D.W.; Colombo, A.L. The global burden of chromoblastomycosis. PLoS Negl. Trop. Dis. 2021, 15, e0009611. [Google Scholar] [CrossRef]
- Krzyściak, P.M.; Pindycka-Piaszczyńska, M.; Piaszczyński, M. Chromoblastomycosis. Postepy Dermatol. Alergol. 2014, 31, 310–321. [Google Scholar] [CrossRef]
- Kneipp, L.F.; Rodrigues, M.L.; Holandino, C.; Esteves, F.F.; Souto-Padrón, T.; Alviano, C.S.; Travassos, L.R.; Meyer-Fernandes, J.R. Ectophosphatase activity in conidial forms of Fonsecaea pedrosoi is modulated by exogenous phosphate and influences fungal adhesion to mammalian cells. Microbiology 2004, 150, 3355–3362. [Google Scholar] [CrossRef]
- Kneipp, L.F.; Magalhães, A.S.; Abi-Chacra, E.A.; Souza, L.O.P.; Alviano, C.S.; Santos, A.L.S.; Meyer-Fernandes, J.R. Surface phosphatase in Rhinocladiella aquaspersa: Biochemical properties and its involvement with adhesion. Med. Mycol. 2012, 50, 570–578. [Google Scholar] [CrossRef]
- Santos, A.L.S.; Palmeira, V.F.; Rozental, S.; Kneipp, L.F.; Nimrichter, L.; Alviano, D.S.; Rodrigues, M.L.; Alviano, C.S. Biology and pathogenesis of Fonsecaea pedrosoi, the major etiologic agent of chromoblastomycosis. FEMS Microbiol. Rev. 2007, 31, 570–591. [Google Scholar] [CrossRef] [PubMed]
- Palmeira, V.F.; Kneipp, L.F.; Rozental, S.; Alviano, C.S.; Santos, A.L.S. Beneficial Effects of HIV peptidase inhibitors on Fonsecaea pedrosoi: Promising compounds to arrest key fungal biological processes and virulence. PLoS ONE 2008, 3, e3382. [Google Scholar] [CrossRef]
- Palmeira, V.F.; Alviano, D.S.; Braga-Silva, L.A.; Goulart, F.R.V.; Granato, M.Q.; Rozental, S.; Alviano, C.S.; Santos, A.L.S.; Kneipp, L.F. HIV aspartic peptidase inhibitors modulate surface molecules and enzyme activities involved with physiopathological events in Fonsecaea pedrosoi. Front. Microbiol. 2017, 8, 918. [Google Scholar] [CrossRef]
- Palmeira, V.F.; Goulart, F.R.V.; Granato, M.Q.; Alviano, D.S.; Alviano, C.S.; Kneipp, L.F.; Santos, A.L.S. Fonsecaea pedrosoi sclerotic cells: Secretion of aspartic-type peptidase and susceptibility to peptidase inhibitors. Front. Microbiol. 2018, 9, 1383. [Google Scholar] [CrossRef] [PubMed]
- Granato, M.Q.; Massapust, P.A.; Rozental, S.; Alviano, C.S.; Santos, A.L.S.; Kneipp, L.F. 1,10-phenanthroline inhibits the metallopeptidase secreted by Phialophora verrucosa and modulates its growth, morphology and differentiation. Mycopathologia 2015, 179, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Granato, M.Q.; Sousa, I.S.; Rosa, T.L.S.A.; Gonçalves, D.S.; Seabra, S.H.; Alviano, D.S.; Pessolani, M.C.V.; Santos, A.L.S.; Kneipp, L.F. Aspartic peptidase of Phialophora verrucosa as target of HIV peptidase inhibitors: Blockage of its enzymatic activity and interference with fungal growth and macrophage interaction. J. Enzyme Inhib. Med. Chem. 2020, 35, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Mendes-Giannini, M.J.S.; Soares, C.P.; da Silva, J.L.M.; Andreotti, P.F. Interaction of pathogenic fungi with host cells: Molecular and cellular approaches. FEMS Immunol. Med. Microbiol. 2005, 45, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, D.C.; Filler, S.G. Host cell invasion by medically important fungi. Cold Spring Harb. Perspect. Med. 2015, 5, a019687. [Google Scholar] [CrossRef]
- Harding, M.W.; Marques, L.L.; Howard, R.J.; Olson, M.E. Can filamentous fungi form biofilms? Trends Microbiol. 2009, 17, 475–480. [Google Scholar] [CrossRef]
- Vila, T.V.M.; Rozental, S. Biofilm formation as a pathogenicity factor of medically important fungi. In Fungal Pathogenicity; Sultan, S., Ed.; InTech: London, UK, 2016; pp. 953–978. [Google Scholar]
- Roudbary, M.; Vahedi-Shahandashti, R.; Santos, A.L.S.; Mohammadi, S.R.; Aslani, P.; Lass-florl, C.; Rodrigues, C.F. Biofilm formation in clinically relevant filamentous fungi: A therapeutic challenge. Crit. Rev. Microbiol. 2021, 48, 197–221. [Google Scholar] [CrossRef]
- Hawser, S.P.; Douglas, L.J. Biofilm formation by Candida species on the surface of catheter materials in vitro. Infect. Immun. 1994, 62, 915–921. [Google Scholar] [CrossRef]
- Lattif, A.A.; Mukherjee, P.K.; Chandra, J.; Swindell, K.; Lockhart, S.R.; Diekema, D.J.; Pfaller, M.A.; Ghannoum, M.A. Characterization of biofilms formed by Candida parapsilosis, C. metapsilosis, and C. orthopsilosis. Int. J. Med. Microbiol. 2010, 300, 265–270. [Google Scholar] [CrossRef]
- Silva, S.; Negri, M.; Henriques, M.; Oliveira, R.; Williams, D.W.; Azeredo, J. Adherence and biofilm formation of non-Candida albicans Candida species. Trends Microbiol. 2011, 19, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Ramos, L.S.; Mello, T.; Branquinha, M.H.; Santos, A.L.S. Biofilm formed by Candida haemulonii species complex: Structural analysis and extracellular matrix composition. J. Fungi 2020, 6, 46. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.L.S.; Galdino, A.C.M.; Mello, T.; Ramos, L.S.; Branquinha, M.H.; Bolognese, A.M.; Columbano Neto, J.; Roudbary, M. What are the advantages of living in a community? A microbial biofilm perspective! Mem. Inst. Oswaldo Cruz 2018, 113, e180212. [Google Scholar] [CrossRef] [PubMed]
- Mowat, E.; Butcher, J.; Lang, S.; Williams, C.; Ramage, G. Development of a simple model for studying the effects of antifungal agents on multicellular communities of Aspergillus fumigatus. J. Med. Microbiol. 2007, 56, 1205–1212. [Google Scholar] [CrossRef] [PubMed]
- Costa-Orlandi, C.B.; Sardi, J.C.O.; Santos, C.T.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. In vitro characterization of Trichophyton rubrum and T. mentagrophytes biofilms. Biofouling 2014, 30, 719–727. [Google Scholar] [CrossRef]
- Peiqian, L.; Xiaoming, P.; Huifang, S.; Jingxin, Z.; Ning, H.; Birun, L. Biofilm formation by Fusarium oxysporum f. sp. cucumerinum and susceptibility to environmental stress. FEMS Microbiol. Lett. 2014, 350, 138–145. [Google Scholar] [CrossRef]
- Iturrieta-Gonzalez, I.A.; Padovan, A.C.; Bizerra, F.C.; Hahn, R.C.; Colombo, A.L. Multiple species of Trichosporon produce biofilms highly resistant to triazoles and amphotericin B. PLoS ONE 2014, 9, e109553. [Google Scholar] [CrossRef] [PubMed]
- Davis, L.E.; Cook, G.; Costerton, J.W. Biofilm on ventriculo-peritoneal shunt tubing as a cause of treatment failure in coccidioidal meningitis. Emerg. Infect. Dis. 2002, 8, 376–379. [Google Scholar] [CrossRef]
- Percival, S.L.; Suleman, L.; Vuotto, C.; Donelli, G. Healthcare-associated infections, medical devices and biofilms: Risk, tolerance and control. J. Med. Microbiol. 2015, 64, 323–334. [Google Scholar] [CrossRef]
- Zhang, K.; Li, X.; Yu, C.; Wang, Y. Promising therapeutic strategies against microbial biofilm challenges. Front. Cell Infect. Microbiol. 2020, 10, 359. [Google Scholar] [CrossRef]
- Tits, J.; Cammue, B.P.A.; Thevissen, K. Combination therapy to treat fungal biofilm-based infections. Int. J. Mol. Sci. 2020, 21, 8873. [Google Scholar] [CrossRef]
- Ramage, G.; Rajendran, R.; Sherry, L.; Williams, C. Fungal biofilm resistance. Int. J. Microbiol. 2012, 2012, 528521. [Google Scholar] [CrossRef] [PubMed]
- Desai, J.V.; Mitchell, A.P.; Andes, D.R. Fungal biofilms, drug resistance, and recurrent infection. Cold Spring Harb Perspect Med 2014, 4, a019729. [Google Scholar] [CrossRef] [PubMed]
- Wiederhold, N.P. Antifungal resistance: Current trends and future strategies to combat. Infect. Drug Resist. 2017, 10, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, K.F.; Zarnowski, R.; Andes, D.R. The extracellular matrix of fungal biofilms. Adv. Exp. Med. Biol. 2016, 931, 21–35. [Google Scholar]
- Kovács, R.; Majoros, L. Fungal quorum-sensing molecules: A review of their antifungal effect against Candida biofilms. J. Fungi 2020, 6, 99. [Google Scholar] [CrossRef]
- Prasad, R.; Rawal, M.K. Efflux pump proteins in antifungal resistance. Front. Pharmacol. 2014, 5, 202. [Google Scholar] [CrossRef]
- Holmes, A.R.; Cardno, T.S.; Strouse, J.J.; Ivnitski-Steele, I.; Keniya, M.V.; Lackovic, K.; Monk, B.C.; Sklar, L.A.; Cannon, R.D. Targeting efflux pumps to overcome antifungal drug resistance. Future Med. Chem. 2016, 8, 1485–1501. [Google Scholar] [CrossRef]
- Eboigbodin, K.E.; Seth, A.; Biggs, C.A. A review of biofilms in domestic plumbing. J. Am. Water Works Assoc. 2008, 100, 131–138. [Google Scholar] [CrossRef]
- Heinrichs, G.; Hubner, I.; Schmidt, C.K.; de Hoog, G.S.; Haase, G. Analysis of black fungal biofilms occurring at domestic water taps (I): Compositional analysis using tag-encoded FLX amplicon pyrosequencing. Mycopathologia 2013, 175, 387–397. [Google Scholar] [CrossRef]
- Siqueira, V.M.; Lima, N. Biofilm formation by filamentous fungi recovered from a water system. J. Mycol. 2013, 2013, 152941. [Google Scholar] [CrossRef]
- Alonso, M.; Gow, N.; Erwig, L.; Bain, J. Macrophage migration is impaired within Candida albicans biofilms. J. Fungi 2017, 3, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seneviratne, C.J.; Fong, P.H.L.; Wong, S.S.W.; Lee, V.H.F. Antifungal susceptibility and phenotypic characterization of oral isolates of a black fungus from a nasopharyngeal carcinoma patient under radiotherapy. BMC Oral Health 2015, 15, 39. [Google Scholar] [CrossRef] [PubMed]
- Sav, H.; Ozakkas, F.; Altınbas, R.; Kiraz, N.; Tümgör, A.; Gümral, R.; Döğen, A.; Ilkit, M.; de Hoog, G.S. Virulence markers of opportunistic black yeast in Exophiala. Mycoses 2016, 59, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Kirchhoff, L.; Olsowski, M.; Zilmans, K.; Dittmer, S.; Haase, G.; Sedlacek, L.; Steinmann, E.; Buer, J.; Rath, P.-M.; Steinmann, J. Biofilm formation of the black yeast-like fungus Exophiala dermatitidis and its susceptibility to antiinfective agents. Sci. Rep. 2017, 7, 42886. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Sun, X.; Wang, Z.; Zhang, Y.; Hou, W. Keratitis-associated fungi form biofilms with reduced antifungal drug susceptibility. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7774–7778. [Google Scholar] [CrossRef] [PubMed]
- Mello, T.P.; Aor, A.C.; Gonçalves, D.S.; Seabra, S.H.; Branquinha, M.H.; Santos, A.L.S. Assessment of biofilm formation by Scedosporium apiospermum, S. aurantiacum, S. minutisporum and Lomentospora prolificans. Biofouling 2016, 32, 737–749. [Google Scholar] [CrossRef]
- Rollin-Pinheiro, R.; de Meirelles, J.V.; Vila, T.V.M.; Fonseca, B.B.; Alves, V.; Frases, S.; Rozental, S.; Barreto-Bergter, E. Biofilm formation by Pseudallescheria/Scedosporium species: A comparative study. Front. Microbiol. 2017, 8, 1568. [Google Scholar] [CrossRef]
- Kvich, L.; Burmølle, M.; Bjarnsholt, T.; Lichtenberg, M. Do mixed-species biofilms dominate in chronic infections?—Need for in situ visualization of bacterial organization. Front. Cell Infect. Microbiol. 2020, 10, 396. [Google Scholar] [CrossRef]
- Costa-Orlandi, C.B.; Sardi, J.C.O.; Pitangui, N.S.; Oliveira, H.C.; Scorzoni, L.; Galeane, M.C.; Medina-Alarcón, K.P.; Melo, W.C.M.A.; Marcelino, M.Y.; Braz, J.D.; et al. Fungal biofilms and polymicrobial diseases. J. Fungi 2017, 3, 22. [Google Scholar] [CrossRef]
- Bertesteanu, S.; Triaridis, S.; Stankovic, M.; Lazar, V.; Chifiriuc, M.C.; Vlad, M.; Grigore, R. Polymicrobial wound infections: Pathophysiology and current therapeutic approaches. Int. J. Pharm. 2014, 463, 119–126. [Google Scholar] [CrossRef]
- Kalan, L.; Loesche, M.; Hodkinson, B.P.; Heilmann, K.; Ruthel, G.; Gardner, S.E.; Grice, E.A. Redefining the chronic-wound microbiome: Fungal communities are prevalent, dynamic, and associated with delayed healing. mBio 2016, 7, e01058-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, S.G.; Bomfim, M.R.Q.; Azevedo, C.M.P.S.; Martins, C.V.B.; Marques, A.C.G.; Gonçalves, A.G.; Vicente, V.A.; Santos, A.M.; Costa, M.C.; Freitas, G.J.C.; et al. Mixed secondary bacterial infection is associated with severe lesions of chromoblastomycosis in a neglected population from Brazil. Diagn. Microbiol. Infect. Dis. 2019, 95, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Palmeira, V.F.; Kneipp, L.F.; Alviano, C.S.; Santos, A.L.S. Phospholipase and esterase production by clinical strains of Fonsecaea pedrosoi and their interactions with epithelial cells. Mycopathologia 2010, 170, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Peeters, E.; Nelis, H.J.; Coenye, T. Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 2008, 72, 157–165. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, 2nd ed.; Approved Standard M38-A2; Clinical Laboratory Standards Institute CLSI: Wayne, PA, USA, 2008. [Google Scholar]
- Silva, L.N.; Ramos, L.S.; Oliveira, S.S.C.; Magalhães, L.B.; Squizani, E.D.; Kmetzsch, L.; Vainstein, M.H.; Branquinha, M.H.; Santos, A.L.S. Insights into the multi-azole resistance profile in Candida haemulonii species complex. J. Fungi 2020, 6, 215. [Google Scholar] [CrossRef]
- Donlan, R.M. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef]
- Lipke, P.N. What we do not know about fungal cell adhesion molecules. J. Fungi 2018, 4, 59. [Google Scholar] [CrossRef]
- Guzmán-Soto, I.; McTiernan, C.; Gonzalez-Gomez, M.; Ross, A.; Gupta, K.; Suuronen, E.J.; Mah, T.-F.; Griffith, M.; Alarcon, E.I. Mimicking biofilm formation and development: Recent progress in in vitro and in vivo biofilm models. iScience 2021, 24, 102443. [Google Scholar] [CrossRef]
- Ramage, G.; VandeWalle, K.; López-Ribot, J.L.; Wickes, B.L. The filamentation pathway controlled by the Efg1 regulator protein is required for normal biofilm formation and development in Candida albicans. FEMS Microbiol. Lett. 2002, 214, 95–100. [Google Scholar] [CrossRef]
- Tan, X.; Fuchs, B.B.; Wang, Y.; Chen, W.; Yuen, G.J.; Chen, R.B.; Jayamani, E.; Anastassopoulou, C.; Pukkila-Worley, R.; Coleman, J.J.; et al. The role of Candida albicans SPT20 in filamentation, biofilm formation and pathogenesis. PLoS ONE 2014, 9, e94468. [Google Scholar] [CrossRef]
- Kaur, S.; Singh, S. Biofilm formation by Aspergillus fumigatus. Med. Mycol. 2014, 52, 2–9. [Google Scholar] [PubMed] [Green Version]
- Mello, T.; Oliveira, S.S.C.; Frases, S.; Branquinha, M.H.; Santos, A.L.S. Surface properties, adhesion and biofilm formation on different surfaces by Scedosporium spp. and Lomentospora prolificans. Biofouling 2018, 34, 800–814. [Google Scholar] [CrossRef] [PubMed]
- Alnuaimi, A.D.; O’Brien-Simpson, N.M.; Reynolds, E.C.; McCullough, M.J. Clinical isolates and laboratory reference Candida species and strains have varying abilities to form biofilm. FEMS Yeast Res. 2013, 13, 689–699. [Google Scholar] [CrossRef]
- Kean, R.; Delaney, C.; Rajendran, R.; Sherry, L.; Thomas, R.; Williams, C.; Ramage, G.; Metcalfe, R.; McLean, W. Gaining insights from Candida biofilm heterogeneity: One size does not fit all. J. Fungi 2018, 4, 12. [Google Scholar] [CrossRef]
- Azeredo, J.; Azevedo, N.F.; Briandet, R.; Cerca, N.; Coenye, T.; Costa, A.R.; Desvaux, M.; Bonaventura, G.D.; Hébraud, M.; Jaglic, Z.; et al. Critical review on biofilm methods. Crit. Rev. Microbiol. 2017, 43, 313–351. [Google Scholar] [CrossRef]
- Montelongo-Jauregui, D.; Srinivasan, A.; Ramasubramanian, A.K.; Lopez-Ribot, J.L. An in vitro model for oral mixed biofilms of Candida albicans and Streptococcus gordonii in synthetic saliva. Front. Microbiol. 2016, 7, 686. [Google Scholar] [CrossRef] [PubMed]
- Cavalheiro, M.; Teixeira, M.C. Candida biofilms: Threats, challenges, and promising strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef]
- Rajendran, R.; Sherry, L.; Lappin, D.F.; Nile, C.J.; Smith, K.; Williams, C.; Munro, C.A.; Ramage, G. Extracellular DNA release confers heterogeneity in Candida albicans biofilm formation. BMC Microbiol. 2014, 14, 303. [Google Scholar] [CrossRef]
- Rajendran, R.; Williams, C.; Lappin, D.F.; Millington, O.; Martins, M.; Ramage, G. Extracellular DNA release acts as an antifungal resistance mechanism in mature Aspergillus fumigatus biofilms. Eukaryot Cell 2013, 12, 420–429. [Google Scholar] [CrossRef]
- Martins, M.; Uppuluri, P.; Thomas, D.P.; Cleary, I.A.; Henriques, M.; Lopez-Ribot, J.L.; Oliveira, R. Presence of extracellular DNA in the Candida albicans biofilm matrix and its contribution to biofilms. Mycopathologia 2010, 169, 323–331. [Google Scholar] [CrossRef]
- Chandra, J.; Mukherjee, P.K.; Leidich, S.D.; Faddoul, F.F.; Hoyer, L.L.; Douglas, L.J.; Ghannoum, M.A. Antifungal resistance of candidal biofilms formed on denture acrylic in vitro. J. Dent. Res. 2001, 80, 903–908. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Henriques, M.; Martins, A.; Oliveira, R.; Williams, D.; Azeredo, J. Biofilms of non-Candida albicans Candida species: Quantification, structure and matrix composition. Med. Mycol. 2009, 47, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Fanning, S.; Mitchell, A.P. Fungal biofilms. PLoS Pathog. 2012, 8, e1002585. [Google Scholar] [CrossRef] [PubMed]
- Sardi, J.C.O.; Pitangui, N.S.; Voltan, A.R.; Braz, J.D.; Machado, M.P.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. In vitro Paracoccidioides brasiliensis biofilm and gene expression of adhesins and hydrolytic enzymes. Virulence 2015, 6, 642–651. [Google Scholar] [CrossRef]
- Martinez, L.R.; Casadevall, A. Cryptococcus neoformans biofilm formation depends on surface support and carbon source and reduces fungal cell susceptibility to heat, cold, and UV light. Appl. Environ. Microbiol. 2007, 73, 4592–4601. [Google Scholar] [CrossRef]
- Kirchhoff, L.; Weisner, A.-K.; Schrepffer, M.; Hain, A.; Scharmann, U.; Buer, J.; Rath, P.-M.; Steinmann, J. Phenotypical characteristics of the black yeast Exophiala dermatitidis are affected by Pseudomonas aeruginosa in an artificial sputum medium mimicking cystic fibrosis–like conditions. Front. Microbiol. 2020, 11, 471. [Google Scholar] [CrossRef]
- Martinez, L.R.; Fries, B.C. Fungal Biofilms: Relevance in the setting of human disease. Curr. Fungal Infect. Rep. 2010, 4, 266–275. [Google Scholar] [CrossRef]
- Relucenti, M.; Familiari, G.; Donfrancesco, O.; Taurino, M.; Li, X.; Chen, R.; Artini, M.; Papa, R.; Selan, L. Microscopy methods for biofilm imaging: Focus on SEM and VP-SEM pros and cons. Biology 2021, 10, 51. [Google Scholar] [CrossRef]
- Gonzalez-Ramírez, A.I.; Ramírez-Granillo, A.; Medina-Canales, M.G.; Rodríguez-Tovar, A.V.; Martínez-Rivera, M.A. Analysis and description of the stages of Aspergillus fumigatus biofilm formation using scanning electron microscopy. BMC Microbiol. 2016, 16, 243. [Google Scholar] [CrossRef]
- Yin, W.; Wang, Y.; Liu, l.; He, J. Biofilms: The microbial “protective clothing” in extreme environments. Int. J. Mol. Sci. 2019, 20, 3423. [Google Scholar] [CrossRef]
- Katragkou, A.; Kruhlak, M.J.; Simitsopoulou, M.; Chatzimoschou, A.; Taparkou, A.; Cotten, C.J.; Paliogianni, F.; Diza-Mataftsi, E.; Tsantali, C.; Walsh, T.J.; et al. Interactions between human phagocytes and Candida albicans biofilms alone and in combination with antifungal agents. J. Infect. Dis. 2010, 201, 1941–1949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidler, M.J.; Salvenmoser, S.; Muller, F.M. Aspergillus fumigatus forms biofilms with reduced antifungal drug susceptibility on bronchial epithelial cells. Antimicrob. Agents Chemother. 2008, 52, 4130–4136. [Google Scholar] [CrossRef] [PubMed]
- Kernien, J.F.; Snarr, B.D.; Sheppard, D.C.; Nett, J.E. The Interface between fungal biofilm and innate immunity. Front. Immunol. 2018, 8, 1968. [Google Scholar] [CrossRef] [PubMed]
- Arce Miranda, J.E.; Baronetti, J.L.; Sotomayor, C.E.; Paraje, M.G. Oxidative and nitrosative stress responses during macrophage-Candida albicans biofilm interaction. Med. Mycol. 2019, 57, 101–113. [Google Scholar] [CrossRef]
- Mello, T.; Ramos, L.S.; Braga-silva, L.A.; Branquinha, M.H.; Santos, A.L.S. Fungal biofilm—A real obstacle against an efficient therapy: Lessons from Candida. Curr. Top. Med. Chem. 2017, 17, 1987–2004. [Google Scholar] [CrossRef]
- Morelli, K.A.; Kerkaert, J.D.; Cramer, R.A. Aspergillus fumigatus biofilms: Toward understanding how growth as a multicellular network increases antifungal resistance and disease progression. PLoS Pathog. 2021, 17, e1009794. [Google Scholar] [CrossRef]
- Córdova-Alcántara, I.M.; Venegas-Cortés, D.L.; Martínez-Rivera, M.Á.; Pérez, N.O.; Rodriguez-Tovar, A.V. Biofilm characterization of Fusarium solani keratitis isolate: Increased resistance to antifungals and UV light. J. Microbiol. 2019, 57, 485–497. [Google Scholar] [CrossRef]
- Sun, Y.; Gao, L.; He, C.; Li, M.; Zeng, T. In vitro interactions between IAP antagonist AT406 and azoles against planktonic cells and biofilms of pathogenic fungi Candida albicans and Exophiala dermatitidis. Med. Mycol. 2018, 56, 1045–1049. [Google Scholar] [CrossRef]
- Silva, S.; Rodrigues, C.F.; Araújo, D.; Rodrigues, M.E.; Henriques, M. Candida species biofilms’antifungal resistance. J. Fungi 2017, 3, 8. [Google Scholar] [CrossRef]
- Granato, M.Q.; Mello, T.; Nascimento, R.S.; Pereira, M.D.; Rosa, T.L.S.A.; Pessolani, M.C.V.; Mccann, M.; Devereux, M.; Branquinha, M.H.; Santos, A.L.S.; et al. Silver(I) and copper(II) complexes of 1,10-phenanthroline-5,6-dione against Phialophora verrucosa: A focus on the interaction with human macrophages and Galleria mellonella larvae. Front. Microbiol. 2021, 12, 641258. [Google Scholar] [CrossRef]
- Lara, H.H.; Romero-Urbina, D.G.; Pierce, C.; Lopez-Ribot, J.L.; Arellano-Jiménez, M.J.; Jose-Yacaman, M. Effect of silver nanoparticles on Candida albicans biofilms: An ultrastructural study. J. Nanobiotechnol. 2015, 13, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padmavathi, A.R.; Murthy, S.; Das, A.; Priya, A.; Sushmitha, T.J.; Pandian, S.K.; Toleti, S.R. Impediment to growth and yeast-to-hyphae transition in Candida albicans by copper oxide nanoparticles. Biofouling 2020, 36, 56–72. [Google Scholar] [CrossRef] [PubMed]
- Atriwal, T.; Azeem, K.; Husain, F.M.; Hussain, A.; Khan, M.N.; Alajmi, M.F.; Abid, M. Mechanistic understanding of Candida albicans biofilm formation and approaches for its inhibition. Front. Microbiol. 2021, 12, 638609. [Google Scholar] [CrossRef]
- Mello, T.; Silva, L.N.; Ramos, L.S.; Frota, H.F.; Branquinha, M.H.; Santos, A.L.S. Drug repurposing strategy against fungal biofilms. Curr. Top. Med. Chem. 2020, 20, 509–516. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Patterson, T.F. Multidrug-resistant Candida: Epidemiology, molecular mechanisms, and treatment. J. Infect. Dis. 2017, 216, S445–S451. [Google Scholar] [CrossRef] [PubMed]
- Rangel, L.P.; Moreira, O.C.; Livramento, G.N.; Britto, C.; Alviano, D.S.; Alviano, C.S.; Ferreira-Pereira, A. Putative role of an ABC transporter in Fonsecaea pedrosoi multidrug resistance. Int. J. Antimicrob. Agents 2012, 40, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Ponde, N.O.; Lortal, L.; Ramage, G.; Naglik, J.R.; Richardson, J.P. Candida albicans biofilms and polymicrobial interactions. Crit. Rev. Microbiol. 2021, 47, 91–111. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F. pedrosoi MIC (µM/mg/L) | P. verrucosa MIC (µM/mg/L) | |||||
|---|---|---|---|---|---|---|
| Antifungal Agents | Planktonic | Biofilm | bMIC/MIC | Planktonic | Biofilm | bMIC/MIC |
| Amphotericin B | 6.25/5.78 | 200/185 | ↑ 32× | 3.12/2.88 | 25/23 | ↑ 8× |
| Ketoconazole | 0.39/0.21 | 800/425 | ↑ 2051× | 50/27 | 400/213 | ↑ 8× |
| Itraconazole | 0.19/0.13 | >800/>560 | ↑ >4200× | 0.19/0.13 | >800/>560 | ↑ >4200× |
| Posaconazole | 0.04/0.03 | >800/>600 | ↑ >20.000× | 0.01/0.007 | 400/280 | ↑ 40.000× |
| Terbinafine | 0.78/0.23 | >800/>230 | ↑ >1000× | 0.04/0.01 | >800/>230 | ↑ >20.000× |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa, I.S.; Mello, T.P.; Pereira, E.P.; Granato, M.Q.; Alviano, C.S.; Santos, A.L.S.; Kneipp, L.F. Biofilm Formation by Chromoblastomycosis Fungi Fonsecaea pedrosoi and Phialophora verrucosa: Involvement with Antifungal Resistance. J. Fungi 2022, 8, 963. https://doi.org/10.3390/jof8090963
Sousa IS, Mello TP, Pereira EP, Granato MQ, Alviano CS, Santos ALS, Kneipp LF. Biofilm Formation by Chromoblastomycosis Fungi Fonsecaea pedrosoi and Phialophora verrucosa: Involvement with Antifungal Resistance. Journal of Fungi. 2022; 8(9):963. https://doi.org/10.3390/jof8090963
Chicago/Turabian StyleSousa, Ingrid S., Thaís P. Mello, Elaine P. Pereira, Marcela Q. Granato, Celuta S. Alviano, André L. S. Santos, and Lucimar F. Kneipp. 2022. "Biofilm Formation by Chromoblastomycosis Fungi Fonsecaea pedrosoi and Phialophora verrucosa: Involvement with Antifungal Resistance" Journal of Fungi 8, no. 9: 963. https://doi.org/10.3390/jof8090963