
A Review of the Fungi That Degrade Plastic

 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Literature Review
2.2. Phylogenetic Analyses
2.2.1. Taxon Sampling
2.2.2. Initial Phylogenetic Analyses
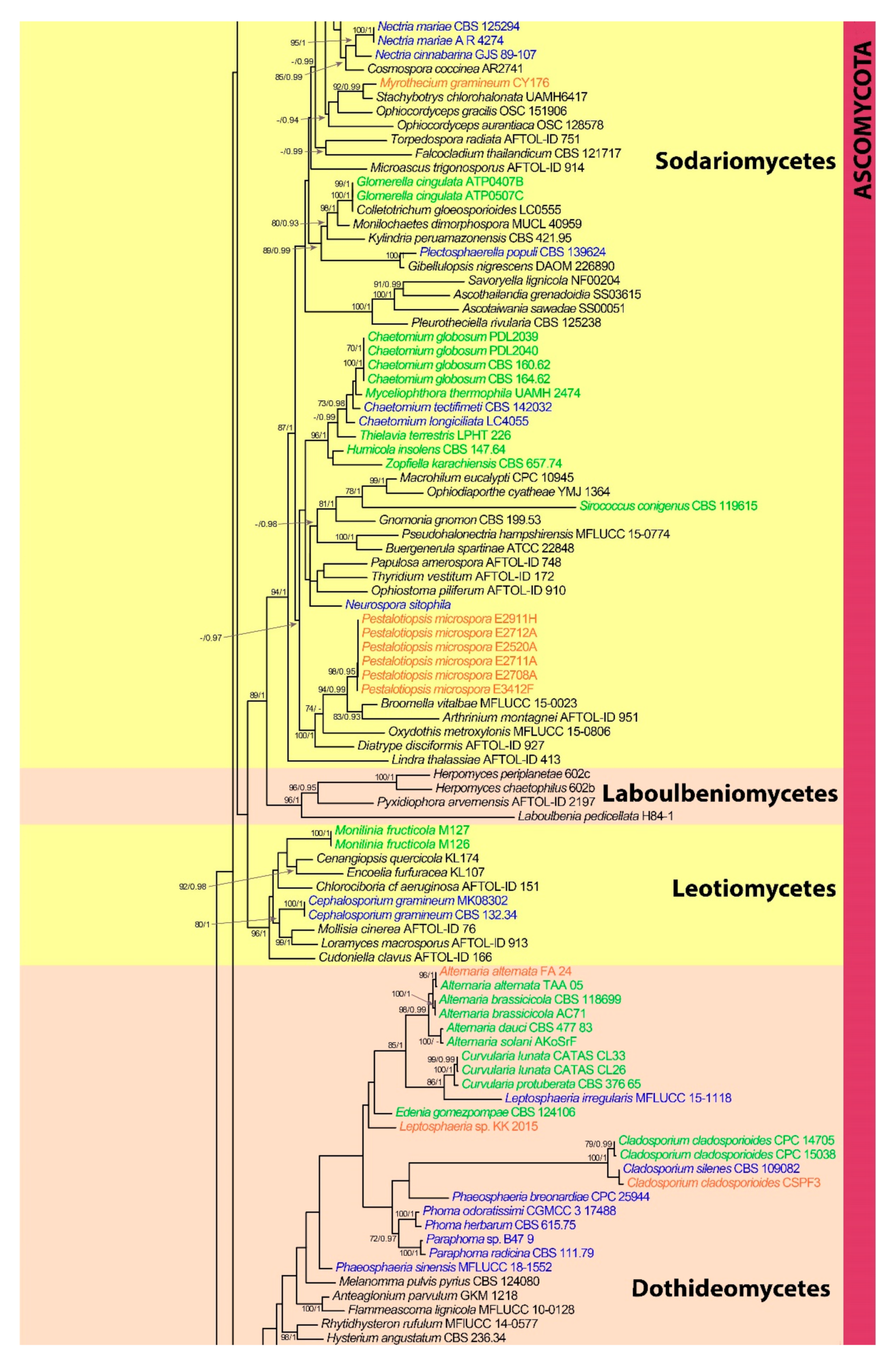
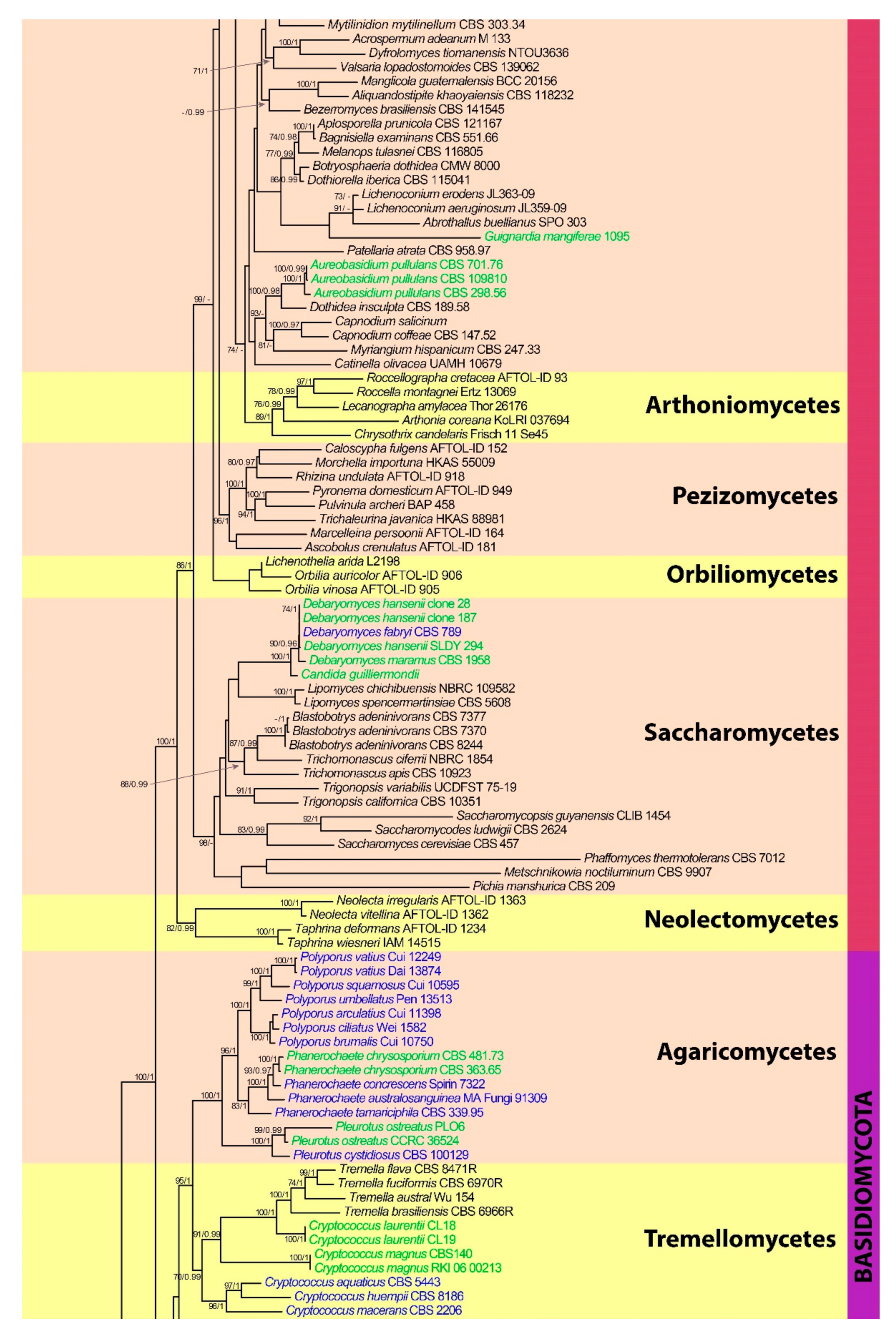
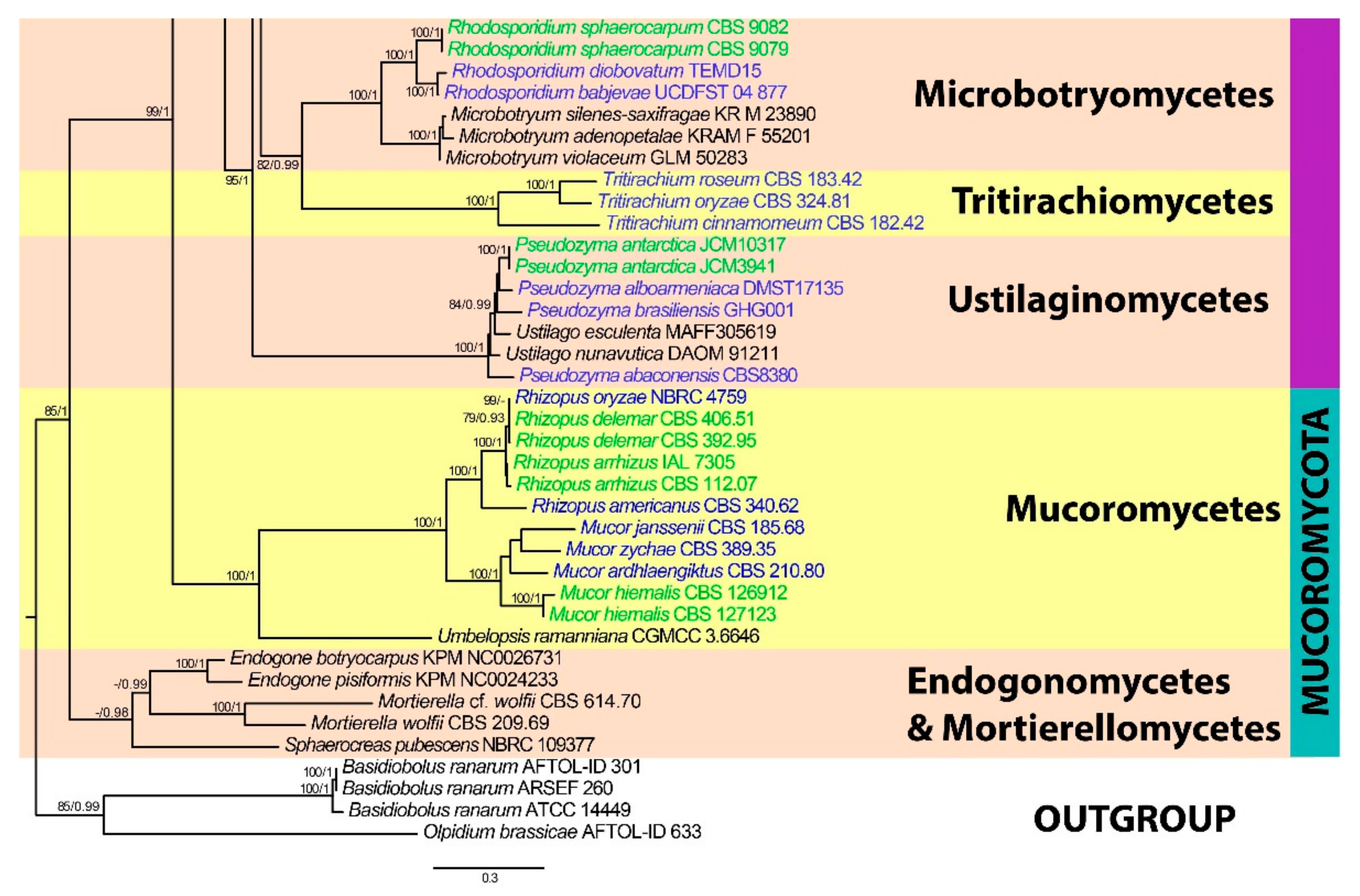
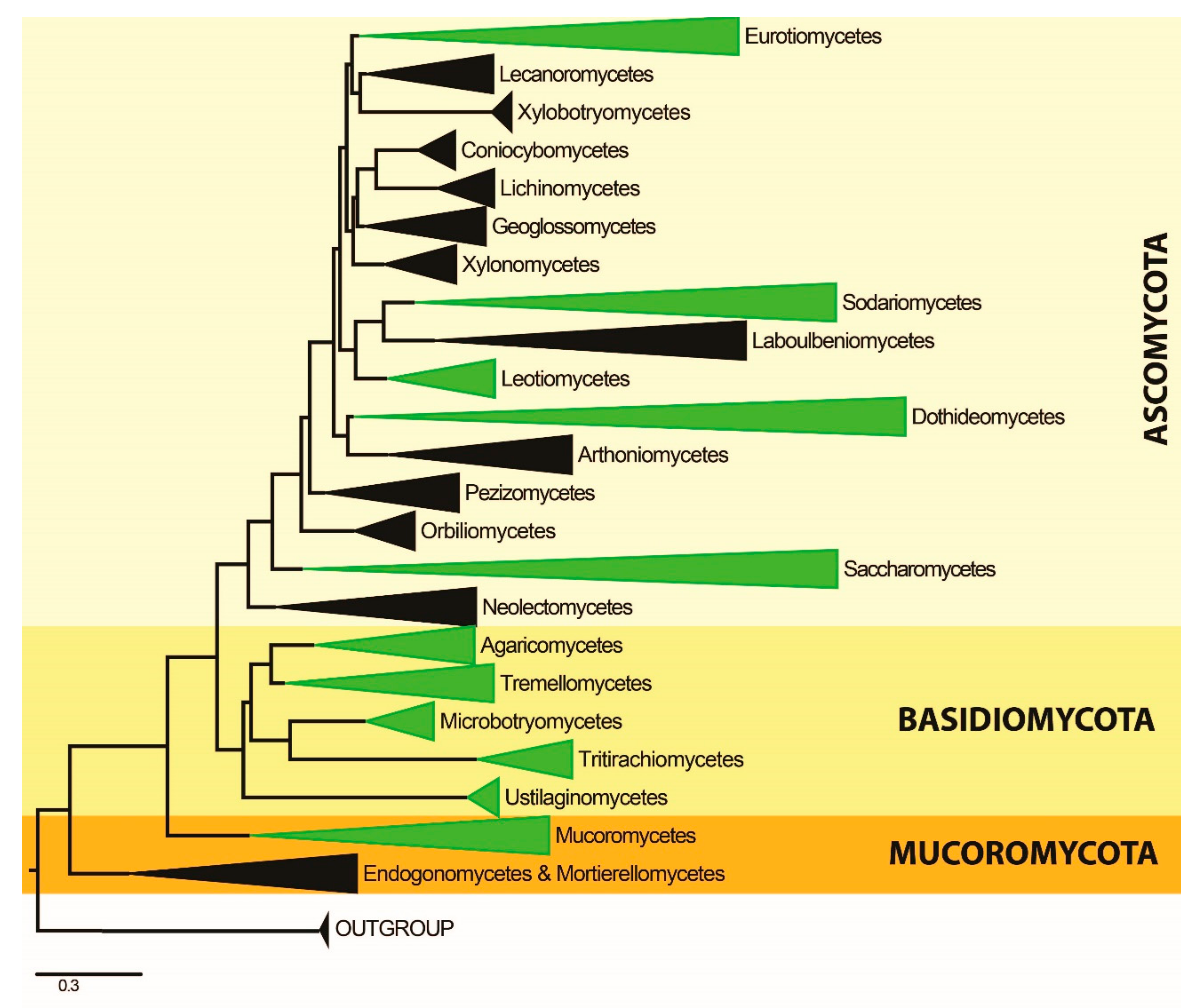
3. Results
3.1. Phylogenetic Analyses
3.2. Phylum Ascomycota
3.2.1. Class Dothideomycetes
3.2.2. Class Eurotiomycetes
3.2.3. Class Leotiomycetes
3.2.4. Class Saccharomycetes
3.2.5. Class Sordariomycetes
3.3. Phylum Basidiomycota
3.3.1. Class Agaricomycetes
3.3.2. Class Microbotryomycetes
3.3.3. Class Tremellomycetes
3.3.4. Class Tritirachiomycetes
3.3.5. Class Ustilaginomycetes
3.4. Phylum Mucoromycota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ogunbayo, A.; Olanipekun, O.; Adamu, I. Preliminary Studies on the Microbial Degradation of Plastic Waste Using Aspergillus niger and Pseudomonas sp. J. Environ. Prot. 2019, 10, 625–631. [Google Scholar] [CrossRef] [Green Version]
- Sangale, M.K.; Shahnawaz, M.; Ade, A.B. Potential of Fungi Isolated from the Dumping Sites Mangrove Rhizosphere Soil to Degrade Polythene. Sci. Rep. 2019, 9, 5390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baheti, P. How Is Plastic Made? Available online: https://www.bpf.co.uk/plastipedia/how-is-plastic-made.aspx (accessed on 15 September 2021).
- Di Bartolo, A.; Infurna, G.; Dintcheva, N.T. A Review of Bioplastics and Their Adoption in the Circular Economy. Polymers 2021, 13, 1229. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, J.E.; Dekle, J.L.; Leads, R.R.; Hunter, R.A. Degradation of Bio-Based and Biodegradable Plastics in a Salt Marsh Habitat: Another Potential Source of Microplastics in Coastal Waters. Mar. Pollut. Bull. 2020, 160, 111518. [Google Scholar] [CrossRef] [PubMed]
- Brunner, I.; Fischer, M.; Rüthi, J.; Stierli, B.; Frey, B. Ability of Fungi Isolated from Plastic Debris Floating in the Shoreline of a Lake to Degrade Plastics. PLoS ONE 2018, 13, e0202047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plastics Europe. How Plastics Are Made. Available online: https://www.plasticseurope.org/en/about-plastics/what-are-plastics/how-plastics-are-made (accessed on 13 September 2021).
- Wayman, C.; Niemann, H. The Fate of Plastic in the Ocean Environment—A Minireview. Environ. Sci. Process. Impacts 2021, 23, 198–212. [Google Scholar] [CrossRef] [PubMed]
- Hardin, T. Plastic: It’s Not All the Same; The Basics on 7 Common Types of Plastic. Available online: https://plasticoceans.org/7-types-of-plastic (accessed on 4 September 2021).
- Plastics Europe. Plastics—The Facts 2020, an Analysis of European Plastics Production, Demand and Waste Data. Available online: https://www.plasticseurope.org/en/resources/publications/4312-plastics-facts-2020 (accessed on 18 September 2021).
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, Use, and Fate of All Plastics Ever Made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [Green Version]
- Rani, A.; Singh, P. Screening of Polyethylene Degrading Fungi from Polyethylene Dump Site. Int. J. Chem. Tech. Res. 2017, 10, 699–704. [Google Scholar]
- United Nations Environment Programme. Fungi Research Lifts Lid on Shy Organisms That Break down Plastic. Available online: https://www.unep.org/news-and-stories/story/fungi-research-lifts-lid-shy-organisms-break-down-plastic (accessed on 12 September 2021).
- Organisation for Economic Co-Operation and Development. Plastic Pollution Is Growing Relentlessly as Waste Management and Recycling Fall Short, Says OECD; Organisation for Economic Co-Operation and Development: Paris, France, 2022. [Google Scholar]
- Jambeck, J.R.; Geyer, R.; Wilcox, C.; Siegler, T.R.; Perryman, M.; Andrady, A.; Narayan, R.; Law, K.L. Plastic Waste Inputs from Land into the Ocean. Science 2015, 347, 768–771. [Google Scholar] [CrossRef]
- da Luz, J.M.R.; da Silva, M.d.C.S.; dos Santos, L.F.; Kasuya, M.C.M. Plastics Polymers Degradation by Fungi. In Microorganisms; IntechOpen: Vienna, Austria, 2019. [Google Scholar]
- Chamas, A.; Moon, H.; Zheng, J.; Qiu, Y.; Tabassum, T.; Jang, J.H.; Abu-Omar, M.; Scott, S.L.; Suh, S. Degradation Rates of Plastics in the Environment. ACS Sustain. Chem. Eng. 2020, 8, 3494–3511. [Google Scholar] [CrossRef] [Green Version]
- Oberbeckmann, S.; Osborn, A.M.; Duhaime, M.B. Microbes on a Bottle: Substrate, Season and Geography Influence Community Composition of Microbes Colonizing Marine Plastic Debris. PLoS ONE 2016, 11, e0159289. [Google Scholar] [CrossRef] [Green Version]
- Ameen, F.; Moslem, M.; Hadi, S.; Al-Sabri, A.E. Biodegradation of Low Density Polyethylene (LDPE) by Mangrove Fungi from the Red Sea Coast. Prog. Rubber Plast. Recycl. Technol. 2015, 31, 125–143. [Google Scholar] [CrossRef]
- Debroas, D.; Mone, A.; Ter Halle, A. Plastics in the North Atlantic Garbage Patch: A Boat-Microbe for Hitchhikers and Plastic Degraders. Sci. Total Environ. 2017, 599, 1222–1232. [Google Scholar] [CrossRef]
- Gascueña, D. Nature’s Allies Against Plastic: Algae, Bacteria, and Fungi. Available online: https://www.bbvaopenmind.com/en/science/environment/natures-allies-against-plastic-algae-bacteria-and-fungi (accessed on 12 September 2021).
- Kale, S.K.; Deshmukh, A.G.; Dudhare, M.S.; Patil, V.B. Microbial Degradation of Plastic: A Review. J. Biochem. Technol. 2015, 6, 952–961. [Google Scholar]
- Webb, H.K.; Arnott, J.; Crawford, R.J.; Ivanova, E.P. Plastic Degradation and Its Environmental Implications with Special Reference to Poly (Ethylene terephthalate). Polymers 2013, 5, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Wilkes, R.A.; Aristilde, L. Degradation and Metabolism of Synthetic Plastics and Associated Products by Pseudomonas sp.: Capabilities and Challenges. J. Appl. Microbiol. 2017, 123, 582–593. [Google Scholar] [CrossRef] [Green Version]
- Pramila, R.; Ramesh, K.V. Biodegradation of Low Density Polyethylene (LDPE) by Fungi Isolated from Municipal Landfill Area. J. Microbiol. Biotechnol. Res. 2011, 1, e136. [Google Scholar]
- Ojha, N.; Pradhan, N.; Singh, S.; Barla, A.; Shrivastava, A.; Khatua, P.; Rai, V.; Bose, S. Evaluation of HDPE and LDPE Degradation by Fungus, Implemented by Statistical Optimization. Sci. Rep. 2017, 7, 39515. [Google Scholar] [CrossRef]
- Soundararajan, N. Pune Researchers Discover Fungi That Can Break up Polythene. Available online: https://researchmatters.in/news/pune-researchers-discover-fungi-can-break-polythene (accessed on 11 September 2021).
- Lear, G.; Kingsbury, J.; Franchini, S.; Gambarini, V.; Maday, S.; Wallbank, J.; Weaver, L.; Pantos, O. Plastics and the Microbiome: Impacts and Solutions. Environ. Microbiome 2021, 16, 2. [Google Scholar] [CrossRef]
- Lacerda, A.L.d.F.; Proietti, M.C.; Secchi, E.R.; Taylor, J.D. Diverse Groups of Fungi Are Associated with Plastics in the Surface Waters of the Western South Atlantic and the Antarctic Peninsula. Mol. Ecol. 2020, 29, 1903–1918. [Google Scholar] [CrossRef]
- Ru, J.; Huo, Y.; Yang, Y. Microbial Degradation and Valorization of Plastic Wastes. Front. Microbiol. 2020, 11, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, C. Fungal Potential for the Degradation of Petroleum-Based Polymers: An Overview of Macro-and Microplastics Biodegradation. Biotechnol. Adv. 2020, 40, 107501. [Google Scholar] [CrossRef] [PubMed]
- Sáenz, M.; Borodulina, T.; Diaz, L.; Banchon, C. Minimal Conditions to Degrade Low Density Polyethylene by Aspergillus terreus and niger. J. Ecol. Eng. 2019, 20, 40–51. [Google Scholar] [CrossRef]
- Iram, D.; Riaz, R.; Iqbal, R.K. Usage of Potential Micro-Organisms for Degradation of Plastics. Open J. Environ. Biol. 2019, 4, 7–15. [Google Scholar]
- Zeghal, E.; Vaksmaa, A.; Vielfaure, H.; Boekhout, T.; Niemann, H. The Potential Role of Marine Fungi in Plastic Degradation—A Review. Front. Mar. Sci. 2021, 8, 738877. [Google Scholar] [CrossRef]
- Statista. Statista: Global No.1 Business Data Platform. 2022. Available online: https://www.statista.com/ (accessed on 22 April 2022).
- Alyamaç-Seydibeyoğlu, E. Impact Modified Poly(Ethylene Terephthalate)-Organoclay Nanocomposites. Master’s Thesis, Middle East Technical University, Ankara, Turkey, 2004. [Google Scholar]
- Karlsson, S.; Ljungquist, O.; Albertsson, A.-C. Biodegradation of Polyethylene and the Influence of Surfactants. Polym. Degrad. Stab. 1988, 21, 237–250. [Google Scholar] [CrossRef]
- Mergaert, J.; Webb, A.; Anderson, C.; Wouters, A.; Swings, J. Microbial Degradation of Poly (3-Hydroxybutyrate) and Poly (3-Hydroxybutyrate-Co-3-Hydroxyvalerate) in Soils. Appl. Environ. Microbiol. 1993, 59, 3233–3238. [Google Scholar] [CrossRef] [Green Version]
- Sowmya, H.; Ramalingappa, B.; Nayanashree, G.; Thippeswamy, B.; Krishnappa, M. Polyethylene Degradation by Fungal Consortium. Int. J. Environ. Res. 2015, 9, 823–830. [Google Scholar]
- Koschorreck, K.; Liu, D.; Kazenwadel, C.; Schmid, R.D.; Hauer, B. Heterologous Expression, Characterization and Site-Directed Mutagenesis of Cutinase CUTAB1 from Alternaria brassicicola. Appl. Microbiol. Biotechnol. 2010, 87, 991–997. [Google Scholar] [CrossRef]
- Russell, J.R.; Huang, J.; Anand, P.; Kucera, K.; Sandoval, A.G.; Dantzler, K.W.; Hickman, D.; Jee, J.; Kimovec, F.M.; Koppstein, D. Biodegradation of Polyester Polyurethane by Endophytic Fungi. Appl. Environ. Microbiol. 2011, 77, 6076–6084. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, I.N.; Maraqa, A.; Hameed, K.M.; Saadoun, I.M.; Maswadeh, H.M. Assessment of Potential Plastic-Degrading Fungi in Jordanian Habitats. Turk. J. Biol. 2011, 35, 551–557. [Google Scholar] [CrossRef]
- Bischoff, F.; Litwińska, K.; Cordes, A.; Baronian, K.; Bode, R.; Schauer, F.; Kunze, G. Three New Cutinases from the Yeast Arxula adeninivorans That Are Suitable for Biotechnological Applications. Appl. Environ. Microbiol. 2015, 81, 5497–5510. [Google Scholar] [CrossRef] [Green Version]
- Benedict, C.V.; Cook, W.J.; Jarrett, P.; Cameron, J.; Huang, S.J.; Bell, J.P. Fungal Degradation of Polycaprolactones. J. Appl. Polym. Sci. 1983, 28, 327–334. [Google Scholar] [CrossRef]
- Nakei, M.D. Isolation and Identification of Plastics—Degrading Microorganisms from Soils of Morogoro, Tanzania. Ph.D. Thesis, Sokoine University of Agriculture, Morogoro, Tanzania, 2015. [Google Scholar]
- Soumya, S.; Nair, B.R. Antifungal Efficacy of Capsicum Frutescens L. Extracts against Some Prevalent Fungal Strains Associated with Groundnut Storage. J. Agric. Technol. 2012, 8, 739–750. [Google Scholar]
- Sakhalkar, S.; Mishra, R. Screening and Identification of Soil Fungi with Potential of Plastic Degrading Ability. Indian J. Appl. Res. 2013, 3, 3. [Google Scholar] [CrossRef]
- Darby, R.T.; Kaplan, A.M. Fungal Susceptibility of Polyurethanes. Appl. Microbiol. 1968, 16, 900–905. [Google Scholar] [CrossRef]
- Zahra, S.; Abbas, S.S.; Mahsa, M.-T.; Mohsen, N. Biodegradation of Low-Density Polyethylene (LDPE) by Isolated Fungi in Solid Waste Medium. Waste Manag. 2010, 30, 396–401. [Google Scholar] [CrossRef]
- Kim, M.-N.; Lee, A.-R.; Yoon, J.-S.; Chin, I.-J. Biodegradation of Poly (3-Hydroxybutyrate), Sky-Green® and Mater-Bi® by Fungi Isolated from Soils. Eur. Polym. J. 2000, 36, 1677–1685. [Google Scholar] [CrossRef]
- Scherer, T.M.; Fuller, R.C.; Lenz, R.W.; Goodwin, S. Production, Purification and Activity of an Extracellular Depolymerase from Aspergillus fumigatus. J. Environ. Polym. Degrad. 1999, 7, 117–125. [Google Scholar] [CrossRef]
- Iyer, S.; Shah, R.; Sharma, A.; Jendrossek, D.; Desai, A. Purification of Aspergillus fumigatus (Pdf1) Poly (β-Hydroxybutyrate)(PHB) Depolymerase Using a New, Single-Step Substrate Affinity Chromatography Method: Characterization of the PHB Depolymerase Exhibiting Novel Self-Aggregation Behavior. J. Polym. Environ. 2000, 8, 197–203. [Google Scholar] [CrossRef]
- Sanchez, J.G.; Tsuchii, A.; Tokiwa, Y. Degradation of Polycaprolactone at 50 °C by a Thermotolerant Aspergillus sp. Biotechnol. Lett. 2000, 22, 849–853. [Google Scholar] [CrossRef]
- Ndahebwa Muhonja, C.; Magoma, G.; Imbuga, M.; Makonde, H.M. Molecular Characterization of Low-Density Polyethene (LDPE) Degrading Bacteria and Fungi from Dandora Dumpsite, Nairobi, Kenya. Int. J. Microbiol. 2018, 2018, 4167845. [Google Scholar] [CrossRef]
- Kathiresan, K. Polythene and Plastics-Degrading Microbes from the Mangrove Soil. Rev. Biol. Trop. 2003, 51, 629–633. [Google Scholar]
- Raaman, N.; Rajitha, N.; Jayshree, A.; Jegadeesh, R. Biodegradation of Plastic by Aspergillus spp. Isolated from Polythene Polluted Sites around Chennai. J. Acad. Indus. Res. 2012, 1, 313–316. [Google Scholar]
- Bermúdez-García, E.; Peña-Montes, C.; Castro-Rodríguez, J.A.; González-Canto, A.; Navarro-Ocaña, A.; Farrés, A. ANCUT2, a Thermo-Alkaline Cutinase from Aspergillus nidulans and Its Potential Applications. Appl. Biochem. Biotechnol. 2017, 182, 1014–1036. [Google Scholar] [CrossRef]
- Raghavan, D.; Torma, A. DSC and FTIR Characterization of Biodegradation of Polyethylene. Polym. Eng. Sci. 1992, 32, 438–442. [Google Scholar] [CrossRef]
- Shabani, F.; Kumar, L.; Esmaeili, A. A Modelling Implementation of Climate Change on Biodegradation of Low-Density Polyethylene (LDPE) by Aspergillus niger in Soil. Glob. Ecol. Conserv. 2015, 4, 388–398. [Google Scholar] [CrossRef] [Green Version]
- Volke-Sepúlveda, T.; Saucedo-Castañeda, G.; Gutiérrez-Rojas, M.; Manzur, A.; Favela-Torres, E. Thermally Treated Low Density Polyethylene Biodegradation by Penicillium pinophilum and Aspergillus niger. J. Appl. Polym. Sci. 2002, 83, 305–314. [Google Scholar] [CrossRef]
- Esmaeili, A.; Pourbabaee, A.A.; Alikhani, H.A.; Shabani, F.; Esmaeili, E. Biodegradation of Low-Density Polyethylene (LDPE) by Mixed Culture of Lysinibacillus xylanilyticus and Aspergillus niger in Soil. PLoS ONE 2013, 8, e71720. [Google Scholar] [CrossRef] [Green Version]
- Nyyssölä, A.; Pihlajaniemi, V.; Järvinen, R.; Mikander, S.; Kontkanen, H.; Kruus, K.; Kallio, H.; Buchert, J. Screening of Microbes for Novel Acidic Cutinases and Cloning and Expression of an Acidic Cutinase from Aspergillus niger CBS 513.88. Enzym. Microb. Technol. 2013, 52, 272–278. [Google Scholar] [CrossRef]
- Laila, M.D. The Potential of Fungi Isolated from Pleurotus ostreatus (Oyster Mushroom) Baglog to Degrade Plastics. Ph.D Thesis, Universitas Andalas, Padang, Indonesia, 2021. [Google Scholar]
- Khruengsai, S.; Sripahco, T.; Pripdeevech, P. Low-Density Polyethylene Film Biodegradation Potential by Fungal Species from Thailand. J. Fungi 2021, 7, 594. [Google Scholar] [CrossRef]
- Munir, E.; Harefa, R.; Priyani, N.; Suryanto, D. Plastic Degrading Fungi Trichoderma viride and Aspergillus nomius Isolated from Local Landfill Soil in Medan; IOP Publishing: Bristol, UK, 2018; Volume 126, p. 012145. [Google Scholar]
- Maeda, H.; Yamagata, Y.; Abe, K.; Hasegawa, F.; Machida, M.; Ishioka, R.; Gomi, K.; Nakajima, T. Purification and Characterization of a Biodegradable Plastic-Degrading Enzyme from Aspergillus oryzae. Appl. Microbiol. Biotechnol. 2005, 67, 778–788. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gosser, Y.; Baker, P.J.; Ravee, Y.; Lu, Z.; Alemu, G.; Li, H.; Butterfoss, G.L.; Kong, X.-P.; Gross, R. Structural and Functional Studies of Aspergillus oryzae Cutinase: Enhanced Thermostability and Hydrolytic Activity of Synthetic Ester and Polyester Degradation. J. Am. Chem. Soc. 2009, 131, 15711–15716. [Google Scholar] [CrossRef] [Green Version]
- Mergaert, J.; Anderson, C.; Wouters, A.; Swings, J.; Kersters, K. Biodegradation of Polyhydroxyalkanoates. FEMS Microbiol. Rev. 1992, 9, 317–321. [Google Scholar] [CrossRef]
- Alshehrei, F. Biodegradation of Low Density Polyethylene by Fungi Isolated from Red Sea Water. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1703–1709. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Nadir, S.; Shah, Z.U.; Shah, A.A.; Karunarathna, S.C.; Xu, J.; Khan, A.; Munir, S.; Hasan, F. Biodegradation of Polyester Polyurethane by Aspergillus tubingensis. Environ. Pollut. 2017, 225, 469–480. [Google Scholar] [CrossRef]
- Gonda, K.; Jendrossek, D.; Molitoris, H.-P. Fungal Degradation of the Thermoplastic Polymer Poly-β-Hydroxybutyric Acid (PHB) under Simulated Deep Sea Pressure. In Life at Interfaces and Under Extreme Conditions; Springer: Berlin/Heidelberg, Germany, 2000; pp. 173–183. [Google Scholar]
- Gu, J.-D.; Mitton, D.; Ford, T.; Mitchell, R. Microbial Degradation of Polymeric Coatings Measured by Electrochemical Impedance Spectroscopy. Biodegradation 1998, 9, 39–45. [Google Scholar] [CrossRef]
- Fields, R.; Rodriguez, F.; Finn, R. Microbial Degradation of Polyesters: Polycaprolactone Degraded by P. Pullulans. J. Appl. Polym. Sci. 1974, 18, 3571–3579. [Google Scholar] [CrossRef]
- Matavulj, M.; Molitoris, H.-P. Fungal Degradation of Polyhydroxyalkanoates and a Semiquantitative Assay for Screening Their Degradation by Terrestrial Fungi. FEMS Microbiol. Rev. 1992, 9, 323–331. [Google Scholar] [CrossRef]
- Kim, H.; Lee, J.W. Effect of Ultrasonic Wave on the Degradation of Polypropylene Melt and Morphology of Its Blend with Polystyrene. Polymer 2002, 43, 2585–2589. [Google Scholar] [CrossRef]
- McLellan, D.W.; Halling, P.J. Acid-Tolerant Poly (3-Hydroxybutyrate) Hydrolases from Moulds. FEMS Microbiol. Lett. 1988, 52, 215–218. [Google Scholar] [CrossRef]
- Owen, S.; Otani, T.; Masaoka, S.; Ohe, T. The Biodegradation of Low-Molecular-Weight Urethane Compounds by a Strain of Exophiala jeanselmei. Biosci. Biotechnol. Biochem. 1996, 60, 244–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, C.A.; Cameron, J.; Huang, S.J.; Vinopal, R.T. Fusarium Polycaprolactone Depolymerase Is Cutinase. Appl. Environ. Microbiol. 1996, 62, 456–460. [Google Scholar] [CrossRef] [Green Version]
- Sang, B.-I.; Hori, K.; Tanji, Y.; Unno, H. Fungal Contribution to in Situ Biodegradation of Poly (3-Hydroxybutyrate-Co-3-Hydroxyvalerate) Film in Soil. Appl. Microbiol. Biotechnol. 2002, 58, 241–247. [Google Scholar] [CrossRef]
- Dimarogona, M.; Nikolaivits, E.; Kanelli, M.; Christakopoulos, P.; Sandgren, M.; Topakas, E. Structural and Functional Studies of a Fusarium oxysporum Cutinase with Polyethylene Terephthalate Modification Potential. Biochim. Biophys. Acta Gen. Subj. 2015, 1850, 2308–2317. [Google Scholar] [CrossRef]
- Nimchua, T.; Punnapayak, H.; Zimmermann, W. Comparison of the Hydrolysis of Polyethylene Terephthalate Fibers by a Hydrolase from Fusarium oxysporum LCH I and Fusarium solani f. sp. pisi. Biotechnol. J. Healthc. Nutr. Technol. 2007, 2, 361–364. [Google Scholar]
- Kwon, M.-A.; Kim, H.S.; Yang, T.H.; Song, B.K.; Song, J.K. High-Level Expression and Characterization of Fusarium solani Cutinase in Pichia Pastoris. Protein Expr. Purif. 2009, 68, 104–109. [Google Scholar] [CrossRef]
- Longhi, S.; Czjzek, M.; Lamzin, V.; Nicolas, A.; Cambillau, C. Atomic Resolution (1.0 Å) Crystal Structure of Fusarium solani Cutinase: Stereochemical Analysis. J. Mol. Biol. 1997, 268, 779–799. [Google Scholar] [CrossRef]
- Prompers, J.J.; Hilbers, C.W.; Pepermans, H.A. Tryptophan Mediated Photoreduction of Disulfide Bond Causes Unusual Fluorescence Behaviour of Fusarium solani pisi Cutinase. FEBS Lett. 1999, 456, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Sowmya, H.; Krishnappa, M.; Thippeswamy, B. Degradation of Polyethylene by Penicillium simplicissimum Isolated from Local Dumpsite of Shivamogga District. Environ. Dev. Sustain. 2015, 17, 731–745. [Google Scholar] [CrossRef]
- Pathirana, R.; Seal, K. Gliocladium Roseum (Bainier), a Potential Biodeteriogen of Polyester Polyurethane Elastomers; John and Wiley and Sons: Hoboken, NJ, USA, 1983. [Google Scholar]
- Seman, W.W.; Bakar, S.; Bukhari, N.; Gaspar, S.; Othman, R.; Nathan, S.; Mahadi, N.; Jahim, J.; Murad, A.; Bakar, F.A. High Level Expression of Glomerella cingulata Cutinase in Dense Cultures of Pichia Pastoris Grown under Fed-Batch Conditions. J. Biotechnol. 2014, 184, 219–228. [Google Scholar] [CrossRef]
- Nyon, M.P.; Rice, D.W.; Berrisford, J.M.; Hounslow, A.M.; Moir, A.J.; Huang, H.; Nathan, S.; Mahadi, N.M.; Bakar, F.D.A.; Craven, C.J. Catalysis by Glomerella cingulata Cutinase Requires Conformational Cycling between the Active and Inactive States of Its Catalytic Triad. J. Mol. Biol. 2009, 385, 226–235. [Google Scholar] [CrossRef]
- Kazenwadel, C.; Eiben, S.; Maurer, S.; Beuttler, H.; Wetzl, D.; Hauer, B.; Koschorreck, K. Thiol-Functionalization of Acrylic Ester Monomers Catalyzed by Immobilized Humicola Insolens Cutinase. Enzym. Microb. Technol. 2012, 51, 9–15. [Google Scholar] [CrossRef]
- Duan, X.; Liu, Y.; You, X.; Jiang, Z.; Yang, S.; Yang, S. High-Level Expression and Characterization of a Novel Cutinase from Malbranchea Cinnamomea Suitable for Butyl Butyrate Production. Biotechnol. Biofuels 2017, 10, 223. [Google Scholar] [CrossRef]
- El-Morsy, E.; Hassan, H.; Ahmed, E. Biodegradative Activities of Fungal Isolates from Plastic Contaminated Soils. Mycosphere 2017, 8, 1071–1087. [Google Scholar] [CrossRef]
- Wang, G.Y.; Michailides, T.J.; Hammock, B.D.; Lee, Y.-M.; Bostock, R.M. Molecular Cloning, Characterization, and Expression of a Redox-Responsive Cutinase from Monilinia fructicola (Wint.) Honey. Fungal Genet. Biol. 2002, 35, 261–276. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Liu, M.; Long, L.; Zhang, R.; Ding, S. Characterization of a Cutinase from Myceliophthora thermophila and Its Application in Polyester Hydrolysis and Deinking Process. Process Biochem. 2018, 66, 106–112. [Google Scholar] [CrossRef]
- Oda, Y.; Asari, H.; Urakami, T.; Tonomura, K. Microbial Degradation of Poly (3-Hydroxybutyrate) and Polycaprolactone by Filamentous Fungi. J. Ferment. Bioeng. 1995, 80, 265–269. [Google Scholar] [CrossRef]
- Suzuki, K.; Noguchi, M.T.; Shinozaki, Y.; Koitabashi, M.; Sameshima-Yamashita, Y.; Yoshida, S.; Fujii, T.; Kitamoto, H.K. Purification, Characterization, and Cloning of the Gene for a Biodegradable Plastic-Degrading Enzyme from Paraphoma-Related Fungal Strain B47-9. Appl. Microbiol. Biotechnol. 2014, 98, 4457–4465. [Google Scholar] [CrossRef]
- Sameshima-Yamashita, Y.; Koitabashi, M.; Tsuchiya, W.; Suzuki, K.; Watanabe, T.; Shinozaki, Y.; Yamamoto-Tamura, K.; Yamazaki, T.; Kitamoto, H. Enhancement of Biodegradable Plastic-Degrading Enzyme Production from Paraphoma-like Fungus, Strain B47-9. J. Oleo Sci. 2016, 65, ess15207. [Google Scholar] [CrossRef] [Green Version]
- Mergaert, J.; Wouters, A.; Swings, J.; Anderson, C. In Situ Biodegradation of Poly (3-Hydroxybutyrate) and Poly (3-Hydroxybutyrate-Co-3-Hydroxyvalerate) in Natural Waters. Can. J. Microbiol. 1995, 41, 154–159. [Google Scholar] [CrossRef]
- Renstad, R.; Karlsson, S.; Albertsson, A.-C. The Influence of Processing Induced Differences in Molecular Structure on the Biological and Non-Biological Degradation of Poly (3-Hydroxybutyrate-Co-3-Hydroxyvalerate), P (3-HB-Co-3-HV). Polym. Degrad. Stab. 1999, 63, 201–211. [Google Scholar] [CrossRef]
- Brucato, C.L.; Wong, S.S. Extracellular Poly (3-Hydroxybutyrate) Depolymerase from Penicillium funiculosum: General Characteristics and Active Site Studies. Arch. Biochem. Biophys. 1991, 290, 497–502. [Google Scholar] [CrossRef]
- Miyazaki, S.; Takahashi, K.; Shiraki, M.; Saito, T.; Tezuka, Y.; Kasuya, K. Properties of a Poly (3-Hydroxybutyrate) Depolymerase from Penicillium funiculosum. J. Polym. Environ. 2000, 8, 175–182. [Google Scholar] [CrossRef]
- Han, J.-S.; Son, Y.-J.; Chang, C.-S.; Kim, M.-N. Purification and Properties of Extracellular Poly (3-Hydroxybutyrate) Depolymerase Produced by Penicillium pinophilum. J. Microbiol. 1998, 36, 67–73. [Google Scholar]
- Torres, A.; Li, S.; Roussos, S.; Vert, M. Screening of Microorganisms for Biodegradation of Poly (Lactic-Acid) and Lactic Acid-Containing Polymers. Appl. Environ. Microbiol. 1996, 62, 2393–2397. [Google Scholar] [CrossRef] [Green Version]
- Yamada-Onodera, K.; Mukumoto, H.; Katsuyaya, Y.; Saiganji, A.; Tani, Y. Degradation of Polyethylene by a Fungus, Penicillium simplicissimum YK. Polym. Degrad. Stab. 2001, 72, 323–327. [Google Scholar] [CrossRef]
- Tokiwa, Y.; Suzuki, T. Purification and Some Properties of Polyethylene Adipate-Degrading Enzyme Produced by Penicillium sp. Strain 14–3. Agric. Biol. Chem. 1977, 41, 265–274. [Google Scholar] [CrossRef] [Green Version]
- Tokiwa, Y. Degradation of Polycaprolactone by a Fungus. J. Appl. Polym. Sci. 1976, 28, 327–334. [Google Scholar]
- Pardo-Rodríguez, M.L.; Zorro-Mateus, P.J.P. Biodegradation of Polyvinyl Chloride by Mucor sp. and Penicillium sp. Isolated from Soil. Rev. Investig. Desarro. Innovación 2021, 11, 387–400. [Google Scholar] [CrossRef]
- Nyyssölä, A.; Pihlajaniemi, V.; Häkkinen, M.; Kontkanen, H.; Saloheimo, M.; Nakari-Setälä, T. Cloning and Characterization of a Novel Acidic Cutinase from Sirococcus conigenus. Appl. Microbiol. Biotechnol. 2014, 98, 3639–3650. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Xu, H.; Yan, Q.; Liu, Y.; Zhou, P.; Jiang, Z. A Low Molecular Mass Cutinase of Thielavia terrestris Efficiently Hydrolyzes Poly (Esters). J. Ind. Microbiol. Biotechnol. 2013, 40, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yan, Q.; Duan, X.; Yang, S.; Jiang, Z. Characterization of an Acidic Cold-Adapted Cutinase from Thielavia terrestris and Its Application in Flavor Ester Synthesis. Food Chem. 2015, 188, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Rubio, M.B.; Cardoza, R.E.; Hermosa, R.; Gutiérrez, S.; Monte, E. Cloning and Characterization of the Thcut1 Gene Encoding a Cutinase of Trichoderma harzianum T34. Curr. Genet. 2008, 54, 301–312. [Google Scholar] [CrossRef]
- Malachová, K.; Novotný, Č.; Adamus, G.; Lotti, N.; Rybková, Z.; Soccio, M.; Šlosarčíková, P.; Verney, V.; Fava, F. Ability of Trichoderma hamatum Isolated from Plastics-Polluted Environments to Attack Petroleum-Based, Synthetic Polymer Films. Processes 2020, 8, 467. [Google Scholar] [CrossRef] [Green Version]
- Roussel, A.; Amara, S.; Nyyssölä, A.; Mateos-Diaz, E.; Blangy, S.; Kontkanen, H.; Westerholm-Parvinen, A.; Carrière, F.; Cambillau, C. A Cutinase from Trichoderma reesei with a Lid-Covered Active Site and Kinetic Properties of True Lipases. J. Mol. Biol. 2014, 426, 3757–3772. [Google Scholar] [CrossRef]
- Thirunavukarasu, K.; Purushothaman, S.; Sridevi, J.; Aarthy, M.; Gowthaman, M.K.; Nakajima-Kambe, T.; Kamini, N.R. Degradation of Poly (Butylene Succinate) and Poly (Butylene Succinate-Co-Butylene Adipate) by a Lipase from Yeast Cryptococcus sp. Grown on Agro-Industrial Residues. Int. Biodeterior. Biodegrad. 2016, 110, 99–107. [Google Scholar] [CrossRef]
- Hung, C.-S.; Barlow, D.E.; Varaljay, V.A.; Drake, C.A.; Crouch, A.L.; Russell, J.N., Jr.; Nadeau, L.J.; Crookes-Goodson, W.J.; Biffinger, J.C. The Biodegradation of Polyester and Polyester Polyurethane Coatings Using Papiliotrema laurentii. Int. Biodeterior. Biodegrad. 2019, 139, 34–43. [Google Scholar] [CrossRef]
- Glenn, J.K.; Gold, M.H. Decolorization of Several Polymeric Dyes by the Lignin-Degrading Basidiomycete Phanerochaete chrysosporium. Appl. Environ. Microbiol. 1983, 45, 1741–1747. [Google Scholar] [CrossRef] [Green Version]
- Tien, M.; Kirk, T.K. Lignin-Degrading Enzyme from the Hymenomycete Phanerochaete chrysosporium Burds. Science 1983, 221, 661–663. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.; Ahmed, S.; Javed, I.; Ali, N.; Atiq, N.; Hameed, A.; Robson, G. Biodegradation of Starch Blended Polyvinyl Chloride Films by Isolated Phanerochaete chrysosporium PV1. Int. J. Environ. Sci. Technol. 2014, 11, 339–348. [Google Scholar] [CrossRef] [Green Version]
- Shinozaki, Y.; Morita, T.; Cao, X.; Yoshida, S.; Koitabashi, M.; Watanabe, T.; Suzuki, K.; Sameshima-Yamashita, Y.; Nakajima-Kambe, T.; Fujii, T. Biodegradable Plastic-Degrading Enzyme from Pseudozyma antarctica: Cloning, Sequencing, and Characterization. Appl. Microbiol. Biotechnol. 2013, 97, 2951–2959. [Google Scholar] [CrossRef]
- Jarerat, A.; Tokiwa, Y. Degradation of Poly (L-lactide) by a Fungus. Macromol. Biosci. 2001, 1, 136–140. [Google Scholar] [CrossRef]
- Tokiwa, H.; Ohnishi, Y.; Rosenkranz, H.S. Mutagenicity and Carcinogenicity of Nitroarenes and Their Sources in the Environment. CRC Crit. Rev. Toxicol. 1986, 17, 23–58. [Google Scholar] [CrossRef]
- Walter, T.; Augusta, J.; Müller, R.-J.; Widdecke, H.; Klein, J. Enzymatic Degradation of a Model Polyester by Lipase from Rhizopus delemar. Enzym. Microb. Technol. 1995, 17, 218–224. [Google Scholar] [CrossRef]
- Nagata, M.; Kiyotsukuri, T.; Takeuchi, S.; Tsutsumi, N.; Sakai, W. Hydrolytic Degradation of Aliphatic Polyesters Copolymerized with Poly (Ethylene Glycol) s. Polym. Int. 1997, 42, 33–38. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. TrimAl: A Tool for Automated Alignment Trimming in Large-Scale Phylogenetic Analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Sánchez, R.; Serra, F.; Tárraga, J.; Medina, I.; Carbonell, J.; Pulido, L.; de María, A.; Capella-Gutíerrez, S.; Huerta-Cepas, J.; Gabaldón, T. Phylemon 2.0: A Suite of Web-Tools for Molecular Evolution, Phylogenetics, Phylogenomics and Hypotheses Testing. Nucleic Acids Res. 2011, 39, W470–W474. [Google Scholar] [CrossRef] [Green Version]
- Hall, T. BioEdit, Version 7.0.9; Ibis Therapeutics: Carlsbad, CA, USA, 2004. [Google Scholar]
- Villesen, P. FaBox: An Online Toolbox for Fasta Sequences. Mol. Ecol. Notes 2007, 7, 965–968. [Google Scholar] [CrossRef]
- Miller, M.A.; Schwartz, T.; Pfeiffer, W. Embedding CIPRES Science Gateway Capabilities in Phylogenetics Software Environments. 2013, p. 1. Available online: https://dl.acm.org/doi/10.1145/2484762.2484806 (accessed on 22 April 2022).
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Nylander, J. MrModeltest, Version 2; Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Rannala, B.; Yang, Z. Probability Distribution of Molecular Evolutionary Trees: A New Method of Phylogenetic Inference. J. Mol. Evol. 1996, 43, 304–311. [Google Scholar] [CrossRef]
- Zhaxybayeva, O.; Gogarten, J.P. Bootstrap, Bayesian Probability and Maximum Likelihood Mapping: Exploring New Tools for Comparative Genome Analyses. BMC Genom. 2002, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.; Ronquist, F. MrBayes: Bayesian Inferences of Phylogeny (Software); University of Rochester: Rochester, NY, USA, 2000. [Google Scholar]
- 135 Rambaut, A.; Drummond, A. Tracer. MCMC Trace Analysis Tool, Version v1.5.0; University of Oxford: Oxford, UK, 2009. [Google Scholar]
- Rambaut, A. FigTree. Tree Figure Drawing Tool, Version 1.3.1; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, UK, 2006. [Google Scholar]
- Hyde, K.D.; Jones, E.G.; Liu, J.-K.; Ariyawansa, H.; Boehm, E.; Boonmee, S.; Braun, U.; Chomnunti, P.; Crous, P.W.; Dai, D.-Q. Families of Dothideomycetes. Fungal Divers. 2013, 63, 1–313. [Google Scholar] [CrossRef]
- Geiser, D.M.; LoBuglio, K.F.; Gueidan, C. 5 Pezizomycotina: Eurotiomycetes. In Systematics and Evolution; Springer: Berlin/Heidelberg, Germany, 2015; pp. 121–141. [Google Scholar]
- Tsang, C.-C.; Tang, J.Y.; Lau, S.K.; Woo, P.C. Taxonomy and Evolution of Aspergillus, Penicillium and Talaromyces in the Omics Era–Past, Present and Future. Comput. Struct. Biotechnol. J. 2018, 16, 197–210. [Google Scholar] [CrossRef]
- Ekanayaka, A.; Hyde, K.; Gentekaki, E.; McKenzie, E.; Zhao, Q.; Bulgakov, T.; Camporesi, E. Preliminary Classification of Leotiomycetes. Mycosphere 2019, 10, 310–489. [Google Scholar] [CrossRef]
- Tanunchai, B.; Juncheed, K.; Wahdan, S.F.M.; Guliyev, V.; Udovenko, M.; Lehnert, A.-S.; Alves, E.G.; Glaser, B.; Noll, M.; Buscot, F. Analysis of Microbial Populations in Plastic–Soil Systems after Exposure to High Poly (Butylene Succinate-Co-Adipate) Load Using High-Resolution Molecular Technique. Environ. Sci. Eur. 2021, 33, 105. [Google Scholar] [CrossRef]
- Suh, S.-O.; Blackwell, M.; Kurtzman, C.P.; Lachance, M.-A. Phylogenetics of Saccharomycetales, the Ascomycete Yeasts. Mycologia 2006, 98, 1006–1017. [Google Scholar] [CrossRef]
- Issa, N. Towards the Biological Degradation of Plastics: Genetic Engineering of Saccharomyces cerevisiae to Secrete Ideonella Sakaiensis Derived PETase. Ph.D. Thesis, University of Kent, Canterbury, UK, 2021. [Google Scholar]
- Maharachchikumbura, S.S.; Hyde, K.D.; Jones, E.G.; McKenzie, E.H.; Huang, S.-K.; Abdel-Wahab, M.A.; Daranagama, D.A.; Dayarathne, M.; D’souza, M.J.; Goonasekara, I.D. Towards a Natural Classification and Backbone Tree for Sordariomycetes. Fungal Divers. 2015, 72, 199–301. [Google Scholar] [CrossRef]
- Sánchez-García, M.; Ryberg, M.; Khan, F.K.; Varga, T.; Nagy, L.G.; Hibbett, D.S. Fruiting Body Form, Not Nutritional Mode, Is the Major Driver of Diversification in Mushroom-Forming Fungi. Proc. Natl. Acad. Sci. USA 2020, 117, 32528–32534. [Google Scholar] [CrossRef]
- de Mattos-Shipley, K.M.; Ford, K.L.; Alberti, F.; Banks, A.; Bailey, A.M.; Foster, G. The Good, the Bad and the Tasty: The Many Roles of Mushrooms. Stud. Mycol. 2016, 85, 125–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oberwinkler, F. Yeasts in Pucciniomycotina. Mycol. Prog. 2017, 16, 831–856. [Google Scholar] [CrossRef]
- Liu, X.-Z.; Wang, Q.-M.; Göker, M.; Groenewald, M.; Kachalkin, A.; Lumbsch, H.T.; Millanes, A.; Wedin, M.; Yurkov, A.; Boekhout, T. Towards an Integrated Phylogenetic Classification of the Tremellomycetes. Stud. Mycol. 2015, 81, 85–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandopadhyay, S.; Liquet y Gonzalez, J.E.; Henderson, K.B.; Anunciado, M.B.; Hayes, D.G.; DeBruyn, J.M. Soil Microbial Communities Associated with Biodegradable Plastic Mulch Films. Front. Microbiol. 2020, 11, 2840. [Google Scholar] [CrossRef] [PubMed]
- Manohar, C.S.; Boekhout, T.; Müller, W.H.; Stoeck, T. Tritirachium candoliense sp. nov., a Novel Basidiomycetous Fungus Isolated from the Anoxic Zone of the Arabian Sea. Fungal Biol. 2014, 118, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Soto, D.; Ortiz-Castellanos, L.; Robledo-Briones, M.; León-Ramírez, C.G. Molecular Mechanisms Involved in the Multicellular Growth of Ustilaginomycetes. Microorganisms 2020, 8, 1072. [Google Scholar] [CrossRef]
- Walther, G.; Wagner, L.; Kurzai, O. Updates on the Taxonomy of Mucorales with an Emphasis on Clinically Important Taxa. J. Fungi 2019, 5, 106. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Stevenson, A. A Feast of Plastic Chewing through the World’s Waste. In Mountain Futures Inspiration and Innovation from the World’s Highlands; World Agroforestry Centre: Nairobi, Kenya, 2018; pp. 209–212. ISBN 978-9966-108-05-0. [Google Scholar]
- Morone, A. Can Tiny Fungi Solve Our Giant Plastic Problem? Available online: https://weather.com/en-IN/india/pollution/news/2019-04-29-can-tiny-fungi-be-a-solution-to-our-giant-plastic-problem (accessed on 10 September 2021).
- Bioplastics Market Development Update Dynamic Growth: Global Production Capacities of Bioplastics 2020–2025; 2020. Available online: https://docs.european-bioplastics.org/conference/Report_Bioplastics_Market_Data_2020_short_version.pdf (accessed on 10 September 2021).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plastic Type and Structure [9] | Main Uses [9] | Annual Production/ Million Metric Tons [35] | Specific Surface Degradation Rate (Min−Max; μm Tear−1) [17] | |
|---|---|---|---|---|
| Land | Marine | |||
| Polyethylene Terephthalate (PET or PETE) [36] | Beverage bottles, Food bottles/jars and polyester clothing or rope | 30.5 | 0 | - |
| High-Density Polyethylene (HDPE) | Milk cartons, detergent bottles, cereal box liners, toys, buckets, park benches, and rigid pipes | 66.96 | 1.0 (0.91−1.1) | 4.3 (0−11) |
| Polyvinyl Chloride (PVC or Vinyl) | Plumbing pipes, credit cards, human and pet toys, rain gutters, teething rings, IV fluid bags and medical tubing, and oxygen masks | 44.3 | 0 | - |
| Low-Density Polyethylene (>LDPE) | Plastic wrap, sandwich and bread bags, bubble wrap, garbage bags, grocery bags and beverage cups | >1500 | 11 | 15 (0−37) |
| Polypropylene (PP) | Straws, bottle caps, prescription bottles, hot food containers, packaging tape, disposable diapers, and DVD/CD boxes | 56 | - | 7.5 |
| Polystyrene (PS or Styrofoam) | Cups, takeout food containers, shipping and product packaging, egg cartons, cutlery and building insulation | 15.61 | 0 | - |
| Other | Eyeglasses, baby and sports bottles, electronics, CD/DVDs, lighting fixtures, and clear plastic cutlery | - | 270 (20−1400) | 16 (7.5−29) |
| Fungus | Polymer Hydrolysed | Class | Family | Environment | References |
|---|---|---|---|---|---|
| Ascomycota | |||||
| Acremonium kiliense | PE | Sordariomycetes | Bionectriaceae | Soils | [37] |
| Acremonium sp. | PHB, Poly[3 HB-co-(10 mol%) 3HV] | Sordariomycetes | Bionectriaceae | Soils | [38] |
| Alternaria alternata | PE, LDPE | Dothideomycetes | Pleosporaceae | Dumpsites, Mangrove stands | [19,39] |
| Alternaria brassicicola | - | Dothideomycetes | Pleosporaceae | Not mentioned | [40] |
| Alternaria dauci | PUR | Dothideomycetes | Pleosporaceae | Rainforest | [41] |
| Alternaria solani | PS-PUR | Dothideomycetes | Pleosporaceae | Soils, Wall paints (Latex), Pieces of plastic debris | [42] |
| Alternaria sp. | PUR | Dothideomycetes | Pleosporaceae | Rainforest | [41] |
| Arxula adeninivorans | - | Saccharomycetes | Trichomonascaceae | Not mentioned | [43] |
| Aspergillus caespitosus | LDPE | Eurotiomycetes | Aspergillaceae | Mangrove stands | [19] |
| Aspergillus fischeri | PCL | Eurotiomycetes | Aspergillaceae | Soils | [44,45] |
| Aspergillus flavus | PE, HDPE, LDPE, PVC, PCL, PS-PUR, PEA, PPA, PBA | Eurotiomycetes | Aspergillaceae | Soils | [12,42,44,46,47,48] |
| Aspergillus fumigatus | PHB, Poly[3HB-co-(10 mol%) 3HV], HDPE, LDPE, PS-PUR, Sky-Green, Poly[3HB-co-(7–77 mol%) 3HV], PHV, Poly[3HB-co-(13–61 mol%) 4HB], PES, PEA, PBA, PCL, PBS | Eurotiomycetes | Aspergillaceae | Soils | [12,38,42,49,50,51,52,53,54] |
| Aspergillus glaucus | PE | Eurotiomycetes | Aspergillaceae | Mangrove Soils | [55] |
| Aspergillus japonicus | LDPE | Eurotiomycetes | Aspergillaceae | Polythene polluted sites | [56] |
| Aspergillus nidulans | LDPE | Eurotiomycetes | Aspergillaceae | Dumpsite | [54,57] |
| Aspergillus Niger | PE, HDPE, LDPE, PVC, Sky-Green, PEA, PPA, PBA | Eurotiomycetes | Aspergillaceae | Soils | [12,47,48,50,56,58,59,60,61,62,63,64] |
| Aspergillus nomius | LDPE | Eurotiomycetes | Aspergillaceae | Landfill soils | [65] |
| Aspergillus ochraceus | LDPE | Eurotiomycetes | Aspergillaceae | Pleurotus ostreatus (Oyster mushroom) baglog | [63] |
| Aspergillus oryzae | LDPE | Eurotiomycetes | Aspergillaceae | Not mentioned | [54,66,67] |
| Aspergillus penicilloides | PHB | Eurotiomycetes | Aspergillaceae | Biological products | [68] |
| Aspergillus sp. | PE | Eurotiomycetes | Aspergillaceae | Sea water | [69] |
| Aspergillus sydowii | PE | Eurotiomycetes | Aspergillaceae | Dumping sites, Mangrove rhizosphere soils | [2] |
| Aspergillus terreus | LDPE, HDPE, PS-PUR, PE | Eurotiomycetes | Aspergillaceae | Soils | [2,12,19,42,45,49] |
| Aspergillus tubingensis | PU | Eurotiomycetes | Aspergillaceae | Soils | [70] |
| Aspergillus ustus | Sky-Green, PHB | Eurotiomycetes | Aspergillaceae | Soils, Deep Sea | [50,71] |
| Aspergillus versicolor | HDPE, LDPE, PVC, PEA, PPA, PBA | Eurotiomycetes | Aspergillaceae | Soils, Degraded polyimides, Marine water | [47,48,71,72] |
| Aureobasidium pullulans | PCL, PEA, PPA, PBA | Dothideomycetes | Saccotheciaceae | Not mentioned | [48,73] |
| Bionectria sp. | PUR | Sordariomycetes | Bionectriaceae | Rainforest | [41] |
| Candida guilliermondii | PHB | Saccharomycetes | Candidaceae | Deep sea | [71] |
| Cephalosporium sp. | PHB | Sordariomycetes | Incertae sedis | Not mentioned | [74] |
| Chaetomium globosum | HDPE, LDPE, PVC, PCL, PEA, PPA, PBA | Sordariomycetes | Chaetomiaceae | Soils | [44,47,48] |
| Chaetomium sp. | PE | Sordariomycetes | Chaetomiaceae | Groundnut | [46] |
| Chrysonilia setophila | HDPE, LDPE, PVC | Sordariomycetes | Sordariaceae | Soils | [47] |
| Cladosporium cladosporioides | PU | Sordariomycetes | Chaetomiaceae | Plastic debris in a shoreline of a lake | [6] |
| Cladosporium sp. | PHB | Sordariomycetes | Chaetomiaceae | Not mentioned | [74] |
| Colletotrichum fructicola | LDPE | Sordariomycetes | Glomerellaceae | Not mentioned | [64] |
| Curvularia lunata | PE | Dothideomycetes | Pleosporaceae | Dumpsites | [39] |
| Curvularia protuberata | Sky-Green | Dothideomycetes | Pleosporaceae | Soils | [50] |
| Debaryomyces hansenii | PHB | Saccharomycetes | Debaryomycetaceae | Deep sea | [71] |
| Diaporthe italiana | LDPE | Sordariomycetes | Diaporthaceae | Not mentioned | [64] |
| Edenia gomezpompae | PUR | Dothideomycetes | Phaeosphaeriaceae | Rainforest | [41] |
| Emericellopsis minima | PHB, Poly[3HB-co-(30 mol%) 3HV] | Sordariomycetes | Incertae sedis | Not mentioned | [75] |
| Eupenicillium hirayamae | - | Eurotiomycetes | Aspergillaceae | Mangrove stand | [19] |
| Eupenicillium rubidurum | - | Eurotiomycetes | Aspergillaceae | Not mentioned | [45] |
| Eupenicillium sp. | PHB | Eurotiomycetes | Aspergillaceae | Soils | [76] |
| Exophiala jeanselmei | Polyether | Eurotiomycetes | Herpotrichiellaceae | Soils | [77] |
| Fusarium moniiforme | PCL | Sordariomycetes | Nectriaceae | Not mentioned | [78] |
| Fusarium oxysporium | Poly[3HB-co-(12 mol%) 3HV], HDPE, LDPE, PVC, PET | Sordariomycetes | Nectriaceae | Soils | [47,79,80,81] |
| Fusarium solani | LDPE, HDPE, PVC, PCL, PS-PUR, PHB, PET | Sordariomycetes | Nectriaceae | Soils | [12,42,47,49,50,78,81,82,83,84] |
| Fusarium sp. | PE, PCL | Sordariomycetes | Nectriaceae | Soils, Dumpsites | [44,85] |
| Gliocladium roseum | PS-PUR | Sordariomycetes | Hypocreaceae | Not mentioned | [86] |
| Gliocladium virens | LDPE | Sordariomycetes | Hypocreaceae | Not mentioned | [60] |
| Glomerella cingulata | - | Sordariomycetes | Glomerellaceae | Not mentioned | [87,88] |
| Guignardia mangiferae | PUR | Dothideomycetes | Phyllostictaceae | Rainforest | [41] |
| Humicola insolens | - | Sordariomycetes | Chaetomiaceae | Not mentioned | [89] |
| Lasiodiplodia sp. | PUR | Dothideomycetes | Botryosphaeriaceae | Rainforest | [41] |
| Leptosphaeria sp. | PU | Dothideomycetes | Botryosphaeriaceae | Plastic debris in a shoreline of a lake | [6] |
| Malbranchea cinnamomea | - | Eurotiomycetes | Onygenaceae | Not mentioned | [90] |
| Monascus sp. | PU | Eurotiomycetes | Monascaceae | Plastic contaminated soils | [91] |
| Monilinia fructicola | - | Leotiomycetes | Sclerotiniaceae | Not mentioned | [92] |
| Myceliophthora thermophila | - | Sordariomycetes | Chaetomiaceae | Not mentioned | [93] |
| Nectria sp. | PUR | Sordariomycetes | Nectriaceae | Rainforest | [41] |
| Paecilomyces farinosus | Poly[3HB-co-(12 mol%) 3HV], PHB, Sky-Green | Eurotiomycetes | Thermoascaceae | Soils | [50,79] |
| Paecilomyces lilacinus | PHB, PCL, Poly[3HB-co-(12 mol%) 3HV] | Eurotiomycetes | Thermoascaceae | Soils | [79,94] |
| Paecilomyces marquandii | PHB | Eurotiomycetes | Thermoascaceae | Biological products | [67] |
| Paecilomyces variotii | - | Eurotiomycetes | Thermoascaceae | Mangrove stands | [19] |
| Paraphoma-like | - | Dothideomycetes | Phaeosphaeriaceae | Barley phylloplane | [95,96] |
| Penicillium adametzii | PHB | Eurotiomycetes | Aspergillaceae | Biological products | [68] |
| Penicillium argillaceum | PCL | Eurotiomycetes | Aspergillaceae | Not mentioned | [53] |
| Penicillium chermisinum | PHB | Eurotiomycetes | Aspergillaceae | Freshwater | [97] |
| Penicillium crysosporium | Poly[3HB-co-(7 mol%) 3HV] | Eurotiomycetes | Aspergillaceae | Not mentioned | [98] |
| Penicillium daleae | PHB | Eurotiomycetes | Aspergillaceae | Biological products | [68] |
| Penicillium dupontii | PCL | Eurotiomycetes | Aspergillaceae | Not mentioned | [53] |
| Penicillium funiculosum | PCL, PHB, PHV, Poly[3HB-co-(7, 14%) 4HB], Poly[3HB-co-(7, 27, 45, 71%) 3HV], PEA, PPA, PBA | Eurotiomycetes | Aspergillaceae | Soils | [44,48,50,94,99,100] |
| Penicillium griseofulvum | PU | Eurotiomycetes | Aspergillaceae | Plastic debris in a shoreline of a lake | [6] |
| Penicillium janthinellum | PHB | Eurotiomycetes | Aspergillaceae | Freshwater | [97] |
| Penicillium minioluteum | PHB | Eurotiomycetes | Aspergillaceae | Soils | [50] |
| Penicillium orchrochloron | PHB | Eurotiomycetes | Aspergillaceae | Biological products | [68] |
| Penicillium oxalicum | HDPE, LDPE | Eurotiomycetes | Aspergillaceae | Soils of a plastic dumping site | [26] |
| Penicillium pinophilium | PHB, LDPE | Eurotiomycetes | Aspergillaceae | Soils | [50,60,101] |
| Penicillium restricum | PHB | Eurotiomycetes | Aspergillaceae | Biological products | [68] |
| Penicillium roqueforti | PLA | Eurotiomycetes | Aspergillaceae | Not mentioned | [102] |
| Penicillium simplicissimum | PE, PHB, Poly[3HB-co-(7 mol%) 3HV], Sky-Green | Eurotiomycetes | Aspergillaceae | Soils | [50,76,85,97,98,103] |
| Penicillium sp. | PHB, HDPE, LDPE, PVC, PEA, PCL, polyalkylene dicarboxylic acids | Eurotiomycetes | Aspergillaceae | Soils | [47,104,105,106] |
| Penicillium verruculosum | Mater-Bi | Eurotiomycetes | Aspergillaceae | Soils | [50] |
| Pestalotiopsis microspora | PUR | Sordariomycetes | Sporocadaceae | Rainforest | [41] |
| Pestalotiopsis sp. | PUR | Sordariomycetes | Sporocadaceae | Rainforest | [41] |
| Phaeosphaeria sp. | PUR | Dothideomycetes | Phaeosphaeriaceae | Rainforest | [41] |
| Phialophora alba | - | Eurotiomycetes | Herpotrichiellaceae | Mangrove stands | [19] |
| Phoma sp. | HDPE, LDPE, PVC | Dothideomycetes | Didymellaceae | Soils | [45,47] |
| Plectosphaerella sp. | PUR | Sordariomycetes | Plectosphaerellaceae | Rainforest | [41] |
| Pleosporales sp. | PUR | Dothideomycetes | Rainforest | [41] | |
| Pullularia pullulans | PEA, PPA, PBA | Dothideomycetes | Saccotheciaceae | Not mentioned | [48] |
| Sirococcus conigenus | - | Sordariomycetes | Gnomoniaceae | Not mentioned | [107] |
| Spicaria spp. | PS-PUR | Incertae sedis | Incertae sedis | Soils, Wall paints (Latex), Plastic debris | [42] |
| Stagonosporopsis citrulli | LDPE | Dothideomycetes | Didymellaceae | Not mentioned | [64] |
| Talaromyces islandicus | - | Eurotiomycetes | Aspergillaceae | Not mentioned | [45] |
| Thermoascus aurantiacus | PHB, PCL, PBS | Eurotiomycetes | Thermoascaceae | Not mentioned | [53] |
| Thielavia terrestris | - | Sordariomycetes | Chaetomiaceae | Soils | [108,109] |
| Thyrostroma jaczewskii | LDPE | Dothideomycetes | Botryosphaeriaceae | Not mentioned | [64] |
| Trichoderma hamatum | LDPE, PS, PVC | Sordariomycetes | Hypocreaceae | Plastic waste material | [110,111] |
| Trichoderma reesei | - | Sordariomycetes | Hypocreaceae | Not mentioned | [112] |
| Trichoderma viride | - | Sordariomycetes | Hypocreaceae | Landfill soils | [65] |
| Verticillium Lecanii | PE | Sordariomycetes | Plectosphaerellaceae | Soils | [37] |
| Verticillium leptobactrum | PHB | Sordariomycetes | Plectosphaerellaceae | Soils | [38] |
| Xepiculopsis gramineae | PU | Sordariomycetes | Incertae sedis | Plastic debris in a shoreline of a lake | [6] |
| Zopfiella karachiensis | PUR | Sordariomycetes | Lasiosphaeriaceae | Rainforest | [41] |
| Basidiomycota | |||||
| Cryptococcus laurentii | PCL | Tremellomycetes | Tremellaceae | Soils | [44] |
| Cryptococcus magnus | - | Tremellomycetes | Tremellaceae | Barley Phylloplane | [95] |
| Cryptococcus sp. | PBS, PBSA | Tremellomycetes | Tremellaceae | - | [113] |
| Papiliotrema laurentii | PBS, PBSA | Tremellomycetes | Rhynchogastremaceae | Part of a microbiome analysis of an aircraft | [114] |
| Phanerochaete chrysosporium | LDPE, Poly[3HB-co-(7 mol%) 3HV], PVC | Agaricomycetes | Phanerochaetaceae | Soils | [60,108,115,116,117] |
| Pleurotus ostreatus | PE, LDPE | Agaricomycetes | Pleurotaceae | - | [16] |
| Polyporus circinatus | PHB | Agaricomycetes | Hymenochaetaceae | - | [74] |
| Pseudozyma antarctica | - | Ustilaginomycetes | Ustilaginaceae | Obtained from the culture collection of the Japan Collection of Microorganisms (JCM) of the Riken BioResource Center in Wako, Japan. | [118] |
| Rhodosporidium sphaerocarpum | PHB | Microbotryomycetes | Sporidiobolaceae | Deep sea | [71] |
| Tritirachium album | PLA | Tritirachiomycetes | Tritirachiaceae | - | [119] |
| Mucoromycota | |||||
| Mucor sp. | PHB, PVC | Mucoromycetes | Mucoraceae | - | [74,106] |
| Mucor hiemalis | HDPE, LDPE, PVC | Mucoromycetes | Mucoraceae | Soils | [47] |
| Rhizopus arrhizus | PCL, polyalkylene dicarboxylic acids | Mucoromycetes | Rhizopodaceae | - | [120] |
| Rhizopus delemar | PPA, PET copolymers with dicarboxylic acids | Mucoromycetes | Rhizopodaceae | - | [121,122] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ekanayaka, A.H.; Tibpromma, S.; Dai, D.; Xu, R.; Suwannarach, N.; Stephenson, S.L.; Dao, C.; Karunarathna, S.C. A Review of the Fungi That Degrade Plastic. J. Fungi 2022, 8, 772. https://doi.org/10.3390/jof8080772
Ekanayaka AH, Tibpromma S, Dai D, Xu R, Suwannarach N, Stephenson SL, Dao C, Karunarathna SC. A Review of the Fungi That Degrade Plastic. Journal of Fungi. 2022; 8(8):772. https://doi.org/10.3390/jof8080772
Chicago/Turabian StyleEkanayaka, Anusha H., Saowaluck Tibpromma, Donqin Dai, Ruifang Xu, Nakarin Suwannarach, Steven L. Stephenson, Chengjiao Dao, and Samantha C. Karunarathna. 2022. "A Review of the Fungi That Degrade Plastic" Journal of Fungi 8, no. 8: 772. https://doi.org/10.3390/jof8080772