
Organization and Unconventional Integration of the Mating-Type Loci in Morchella Species
by

Hongmei Chai
1,2,3, 
Ping Liu
1,2,3,
Yuanhao Ma
1,2,3,
Weimin Chen
1,2,3,
Nan Tao
1,2,3 and
Yongchang Zhao
1,2,3,* 1
Biotechnology and Germplasm Resources Research Institute, Yunnan Academy of Agricultural Sciences, Kunming 650205, China
2
Yunnan Provincial Key Lab of Agricultural Biotechnology, Kunming 650205, China
3
Key Lab of Southwestern Crop Gene Resources and Germplasm Innovation, Ministry of Agriculture, Kunming 650205, China
*
Author to whom correspondence should be addressed.
J. Fungi 2022, 8(7), 746; https://doi.org/10.3390/jof8070746
Submission received: 15 June 2022
/
Revised: 14 July 2022
/
Accepted: 16 July 2022
/
Published: 19 July 2022
(This article belongs to the Special Issue Genomics and Evolution of Macrofungi)
Abstract
:True morels (Morchella spp.) are a group of delicious fungi in high demand worldwide, and some species of morels have been successfully cultivated in recent years. To better understand the sexual reproductive mechanisms of these fungi, we characterized the structure of the mating-type loci from ten morel species, and seven of them were obtained using long-range PCR amplification. Among the studied species, eight were heterothallic, two were homothallic, and four types of composition were observed in the MAT loci. In three of the five black morel species, the MAT1-1-1, MAT1-1-10, and MAT1-1-11 genes were in the MAT1-1 idiomorph, and only the MAT1-2-1 gene was in the MAT1-2 idiomorph, while an integration event occurred in the other two species and resulted in the importation of the MAT1-1-11 gene into the MAT1-2 idiomorph and survival as a truncated fragment in the MAT1-1 idiomorph. However, the MAT1-1-11 gene was not available in the four yellow morels and one blushing morel species. M. rufobrunnea, a representative species of the earliest diverging branch of true morels, along with another yellow morel Mes-15, were confirmed to be homothallic, and the MAT1-1-1, MAT1-1-10, and MAT1-2-1 genes were arranged in a tandem array. Therefore, we hypothesized that homothallism should be the ancestral reproductive state in Morchella. RT-PCR analyses revealed that four mating genes could be constitutively expressed, while the MAT1-1-10 gene underwent alternative splicing to produce different splice variants.
1. Introduction
The sexual life cycle of fungi is not only involved in the production of offspring but also is important in generating variability in the populations, which may facilitate the long-term survival of fungi under unfavorable conditions [1]. In ascomycetes, sexual reproduction is usually controlled by a single mating-type locus, which contains one of two dissimilar DNA sequences occupying the same chromosomal locus [2]. These large DNA sequences were described using the word “idiomorph” to indicate their sharing little homology and not being true alleles [3]. To describe different mating genes from various species, Turgeon and Yoder [4] proposed a standardized nomenclature for idiomorphs. The MAT1-1 idiomorph was defined as containing the MAT1-1-1 gene, which was characterized by an alpha-box protein domain, while the MAT1-2 idiomorph harbored the MAT1-2-1 gene that encoded a protein with a high-mobility group (HMG) domain [4]. Besides the core mating genes MAT1-1-1 and MAT1-2-1, additional genes present at the MAT loci are lineage-specific and conserved at the family and class levels [5]. To date, 10 additional MAT1-1 genes (MAT1-1-2 to MAT1-1-11) and 11 other MAT1-2 genes (MAT1-2-2 to MAT1-2-12) have been identified from the MAT locus of various species [6,7,8].
In the heterothallic ascomycete species, individual isolates usually contain either the MAT1-1 or the MAT1-2 idiomorph in each haploid nucleus, and sexual reproduction requires two haploid strains containing opposite mating types. However, the primary homothallic species carrying two mating types (linked or unlinked) in the haploid genome [2] and pseudohomothallic species possessing nuclei of opposite mating types in multinuclear isolates are all capable of self-fertility and outcrossing [9].
True morels (Morchella spp., Morchellaceae, Pezizomycotina, Ascomycota), a group of highly-prized edible fungi with exceptional taste and nutritional value, are found in most parts of the world and are extensively traded. However, inconspicuous morpho-ecological differences make the classification of morel species confusing [10], and Genealogical Concordance Phylogenetic Species Recognition (GCPSR) based on multi-locus data (ITS, RPB1, RPB2, LSU, and EF1-a) is employed in Morchella systematics [11,12]. Three evolutionary clades were designated, including the basal Rufobrunnea clade (blushing morels), the Esculenta clade (yellow morels), and the Elata clade (black morels), while the phylospecies that had not been described were informally assigned the prefix codes “Mes” (for the Esculenta clade) and “Mel” (for the Elata clade), followed by a unique Arabic number [11]. To date, over 80 species-level lineages in the genus Morchella have been recognized worldwide, although the approaches focus more on molecular phylogenetics than morphology to identify cryptic species [13].
In Tuber and Cordycepes, some studies about the reproductive modes and mating genes have been reported [14,15]. In Morchella, by analyzing the mating-type ratios of single-ascospore populations, 15 species of the Elata clade and 19 species of the Esculenta clade have been confirmed to be heterothallic species [16]. The genome data of two ascospore isolates of M. importuna with opposite mating-type have revealed that the lengths of MAT1-1 and MAT1-2 idiomorphs are 10.5 and 6.7 kb, respectively [17]. On the other hand, the MAT1-2 idiomorph harbors a single MAT1-2-1 gene, while the MAT1-1 idiomorph carries MAT1-1-1 and two more newly-described mating genes, namely MAT1-1-10 and MAT1-1-11, which were first reported and named from Morchella [7,17]. The MAT locus of Morchella sp. Mes-20 was verified using long-range polymerase chain reaction (PCR) amplification. The MAT1-2 idiomorph is 7.5 kb in length, harboring a single MAT1-2-1 gene, whereas the MAT1-1 idiomorph is 7.8 kb in length and carries both MAT1-1-1 and MAT1-1-10 genes [7]. Similarly, the ancestral companions of the MAT locus in euascomycetes, APN2 and SLA2, were located on either side of the mating idiomorph of M. importuna and Mes-20, while SDH2 and MBA1 joined the 5′ and 3′ end of the mating idiomorph, respectively [16].
There is a lack of knowledge on the structures and mating systems of the MAT locus for the other morel species. However, this information may provide better insight into the evolutionary history of Morchella and help identify the confusing species. In this study, we characterized the mating loci of several species from three morel clades, respectively, and sought the probable ancestral state of mating systems in this genus.
2. Materials and Methods
2.1. Species and Strains
Ten single-ascospore strains, a MAT1-1 and a MAT1-2 strain for each of the five morel species: M. eximia, M. purpurascena, M. pulchella, Mes-6, and Mes-19, were isolated in this study (Table 1), while the other strains YAASMHL1 and YAASMHL47 of M. sextelata, M115 of Mes-15, and YAASMVR of M. rufobrunnea were obtained from previous studies [7,17]. All strains were stored at the Mushroom Center of Yunnan Germplasm Bank of Crops at the Yunnan Academy of Agricultural Sciences, Kunming, China.
2.2. PCR Amplification and Sequencing of The Mating Idiomorphs
Improved primer pairs p11-1f/p11-2r, based on the p11-2f/p11-2r reported by Chai et al. [7], were used to amplify the MAT1-1 and MAT1-2 idiomorphs of six morel species, except for Mes-15 and M. rufobrunnea. The appropriate information of Mes-15 was derived from the genome data under accession number GWHBJCX00000000 in the Genome Warehouse in the National Genomics Data Center. For M. rufobrunnea, in which the primer pairs p11-1f/p11-2r were unsuccessful, the full-length sequences of the MAT locus were obtained using a roundabout method. First, two fragments of the following regions SDH2 to MAT1-1-1 (p11-2f/p1-3r) and MAT1-2-1 to MBA1 (p1-4f/p2-3r), were amplified. The primer pairs p1-3f/p1-3r and p1-4f/p1-4r could amplify the complete MAT1-1-1 and MAT1-2-1 genes, respectively, of M. rufobrunnea [7]. The primer p2-3r is located on MBA1 and is used to verify the MAT1-2 idiomorph of M. importuna [17]. Then, these two fragments were sequenced. According to the information of the resultant sequences, the primer pair p11-4f/p11-4r was designed to amplify the entire MAT locus (SDH2 to MBA1) of M. rufobrunnea, and the corresponding long-fragment PCR products were sequenced for re-verification. The above-mentioned primers are listed in Supplementary Table S1.
The strains were maintained on potato dextrose agar medium (PDA) at 23 °C in the dark. Total DNA was isolated from 7-day-old mycelia using the Fungal gDNA Isolation Kit (BW-GD2416) (HangZhou Beiwo Meditech Co., Ltd., Hangzhou, China). All amplifications of mating idiomorphs were performed using long-range PCR, as reported by Chai et al. [7]. A 50 µL PCR reaction mixture contained 5 µL of 10 × LA PCR buffer (TaKaRa Bio, Dalian, China), 8 µL of each dNTP (2.5 mM/L), 2 µL each of forward and reverse primer (10 µM/L), 20 ng of template DNA and 0.5 U of TaKaRa LA Taq. The following cycling conditions were used: denaturation at 94 °C for 1 min, followed by 35 cycles at 98 °C for 10 s and 68 °C for 9 min, and a final extension at 72 °C for 10 min.
Once the PCR amplification was confirmed by electrophoresis, the PCR products were purified using E.Z.N.A.® Gel Extraction Kit (Omega Bio-Tek, Inc., Norcross, GA, USA). The PCR products were sequenced with the PCR primers from both 3′ and 5′ termini, after which the resultant sequences were used to design new internal sequencing primers, and complete sequencing coverage in both directions was eventually achieved after several cycles. In cases where a portion of the sequences could not be sequenced smoothly for its special structure, this segment was amplified using internal primer pairs just near the special structure and the corresponding template of mating idiomorph amplicon, after which it was cloned in Escherichia coli DH5α using the pMDTM19-T vector-cloning Kit (TaKaRa Bio, Dalian, China) and sequenced in both directions using the general primers M13R-48 and M13F-47. Eventually, all sequences were assembled using the software SeqManTMII 5.00 (DNASTAR Inc., Madison, WI, USA). Sequences relevant to our analyses have been deposited at GenBank (Table 2).
2.3. Transcriptional Analyses of The Mating Genes
Using a TaKaRa MiniBEST Plant RNA Extraction Kit, the total RNA was extracted from the mycelia that had been incubated on PDA for 10 d. Then, the first-strand cDNAs were synthesized using an Oligo dT primer (PrimeScript II 1st Strand cDNA Synthesis Kit; TaKaRa Bio, Dalian, China). The primer pairs (Supplementary Table S1) were designed to verify the coding regions of the mating genes according to the corresponding information in the mating idiomorphs, ensuring the amplification of all introns. As the design strategy utilized the conserved regions, several primer pairs could be used in some closely-related species. The PCR products were cloned into Escherichia coli and sequenced using the general primers M13R-48 and M13F-47. Introns were verified manually by comparing the RT-PCR sequences with the corresponding DNA sequences.
All PCR primers were designed using the software Primer Premier 5.0 (Premier Biosoft International, Palo Alto, CA, USA). The synthesis of primers and sequencing of all PCR products were performed at the Beijing Tsingke Biotechnology.
2.4. Comparison of The MAT Loci
3. Results
3.1. PCR Amplifications of The Mating Idiomorphs
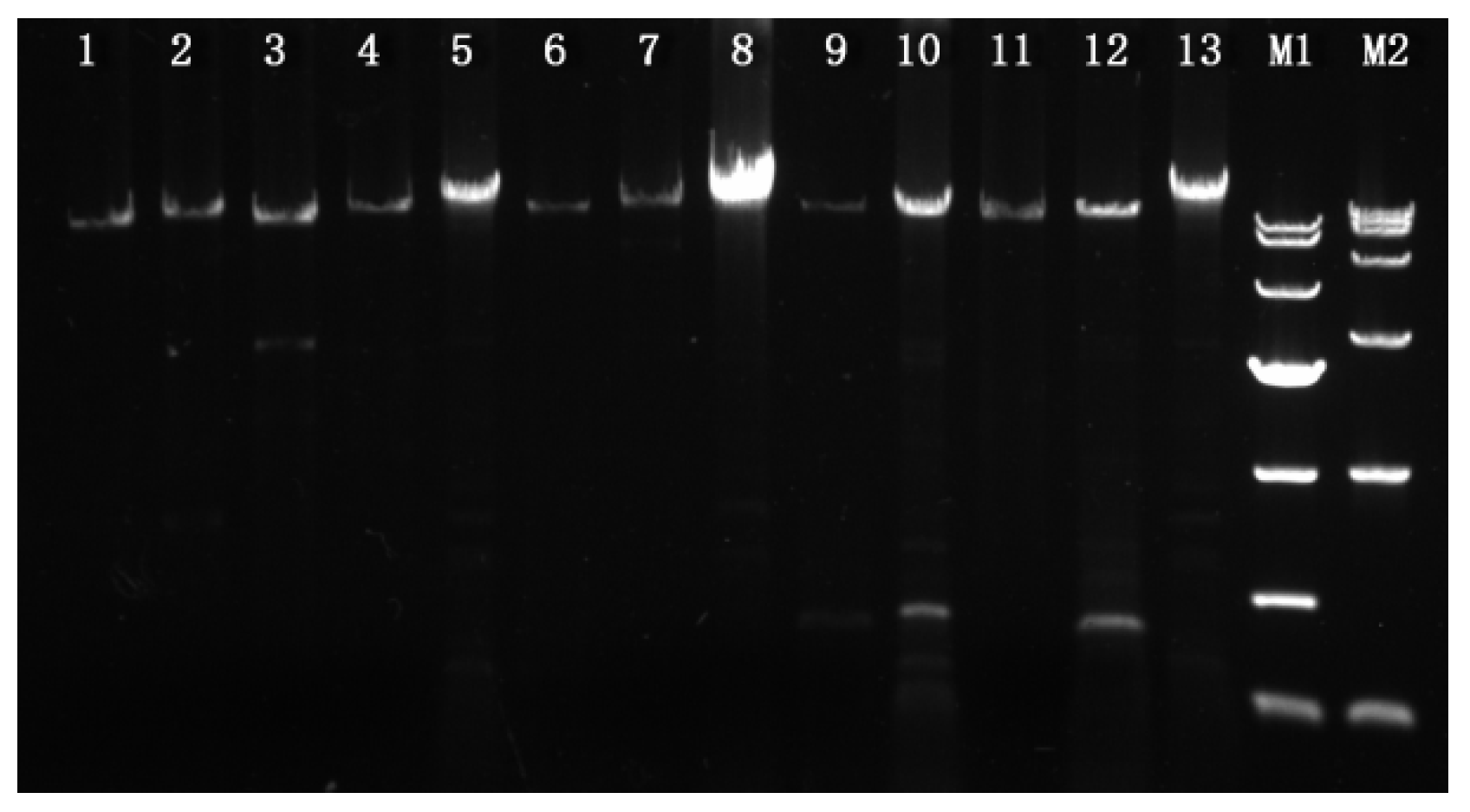
The mating idiomorphs of the 12 single-ascospore strains, a MAT1-1 and a MAT1-2 strain for each of the four black morel species, M. sextelata, M. eximia, M. purpurascena, and M. pulchella, as well as two yellow morel species, Mes-6 and Mes-19, were successfully amplified using the primer pair p11-1f/p11-2r. Primers p11-1f and p11-2r are located in the 5′ and 3′ flanking genes SDH2 and MBA1 of the mating idiomorph, therefore, the ends of the amplicons carried partial sequences of SDH2 and MBA1. Sequence analysis indicated that the lengths of these amplified fragments varied from 8.7 to 18.4 kb. The MAT1-2 amplicons of strains YAASMQM-4 and YAASMHL47 were shortest at 8.7 and 9.0 kb, whereas the similar MAT1-2 fragments of the strains YAASM131-12 and YAASM23-22 were longest at 18 and 18.4 kb, while the other amplified fragments of MAT1-1 or MAT1-2 varied between 9.8 and 13.8 kb (Figure 1).
For strain YAASMVR from the Rufobrunnea clade, the primer pair p11-1f/p11-2r was unsuccessful in amplifying the mating locus, and several attempts were made using the possible primers reported by Chai et al. [7,17]. Eventually, two fragments involving the regions SDH2 to MAT1-1-1 and MAT1-2-1 to MBA1 were obtained and sequenced. Based on this, the primer pair p11-4f and p11-4r, which were located on SDH2 and MAB1, respectively, were designed to amplify the entire mating locus. Using these primers, the lengths of the corresponding amplicons were about 17.8 kb (Figure 1).
3.2. Organization of The Mating Idiomorphs
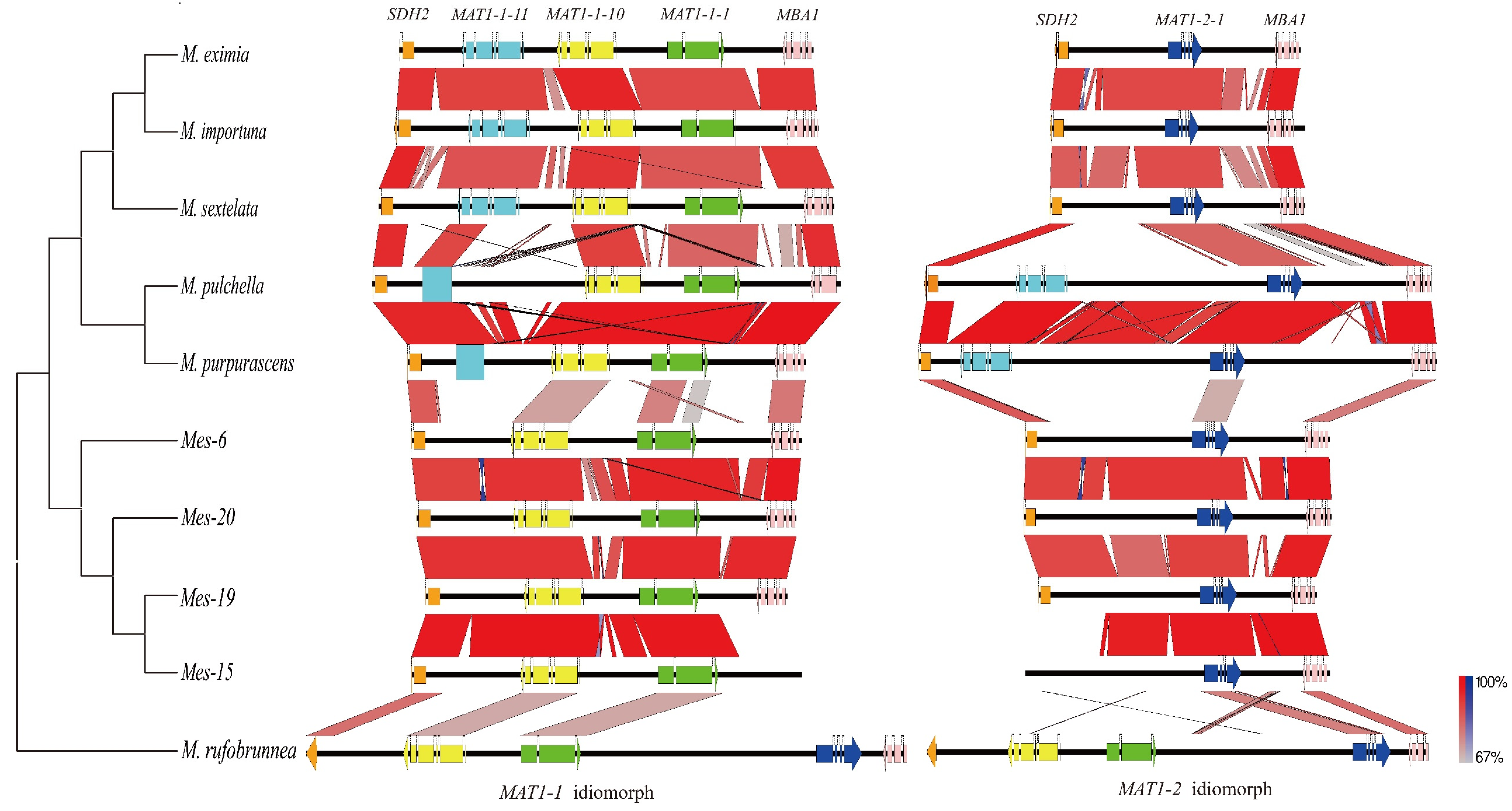
A total of 18 mating idiomorphs from nine morel species, as well as a mating locus from M. rufobrunnea, were analyzed. Of these, the mating idiomorphs from M. importuna and Mes-20 had been characterized previously [7,17], those of Mes-15 were obtained from a draft genome database in our previous study [17], and the others were obtained in this study. The gene arrangements at the mating idiomorphs showed four compositions in these ten species (Figure 2). In the three black morel species M. sextelata, M. eximia, and M. importuna, the MAT1-1 idiomorph contained three complete-mating genes, which were MAT1-1-1, MAT1-1-10, and MAT1-1-11, while the MAT1-2 idiomorph contained the gene MAT1-2-1, which was classified into the M. importuna type. A second organization was observed in two black morel species, M. purpurascena and M. pulchella, and defined as the M. purpurascena type. This type of organization was more special, as the MAT1-2 idiomorph carried two complete-mating genes MAT1-2-1 and MAT1-1-11, whereas a truncated MAT1-1-11 and two complete genes MAT1-1-1 and MAT1-1-10 were present in the MAT1-1 idiomorph. The third Mes-20 type of gene arrangement was observed in four yellow morel species, wherein two genes, MAT1-1-1 and MAT1-1-10, were located on the MAT1-1 idiomorph, while the MAT1-2 idiomorph carried only the MAT1-2-1 gene. For M. rufobrunnea, the mating genes MAT1-1-10, MAT1-1-1, and MAT1-2-1 were arranged in a tandem array and the intergenic length between the MAT1-1-1 and MAT1-2-1 was about 7 kb. Both mating types were linked in the mating locus, a form that was regarded as homothallism.
The size of the mating idiomorph, defined as the region of no detectable homology between opposite mating types, was identified by alignment analyses of two amplicon sequences from the same species, with the identical flanking sequences removed. Interspecific variations in the fragment size ranged from 7.2 to 11.8 kb in the MAT1-1 idiomorphs and from 6.4 to 15.9 kb in the MAT1-2 idiomorphs (Table 2). However, they were similar within the groups regarding the above three mating idiomorphs. For three species from the M. importuna group, the lengths of the MAT1-1 idiomorphs varied from 10.2 to 11.5 kb, which were larger than the 6.6 to 7.1 kb of the MAT1-2 idiomorph. The lengths of the MAT1-1 idiomorph of M. purpurascena and M. pulchella were 9.4 and 11.8 kb, respectively, which were shorter than their MAT1-2 idiomorphs (15.9 kb). For three yellow morel species from the Mes-20 group, the lengths of the MAT1-1 and MAT1-2 idiomorphs were similar and ranged from 6.4 to 8.0 kb. Regarding another yellow morel, Mes-15, the 5′ and 3′ flanking genes were incomplete, and the size of the MAT1-1 and MAT1-2 idiomorphs could not be verified. For the homothallic M. rufobrunnea, the size of the MAT locus, defined as the distance between the stop codon of the 5′ flanking gene (SDH2) and the start codon of the 3′ flanking gene (MBA1), was 16.7 kb in length.
Generally, non-coding regions of the MAT loci showed more inter-specific variability than the coding portions [20]. Between the groups of the above three mating idiomorph types, homology was mainly present in the mating genes and flanking genes, while high sequence divergence was observed in the non-coding regions (Figure 2). This was the same in both M. importuna and M. purpurascena groups, although they were black morels. However, between intra-group species, a high sequence homology in both coding and non-coding regions was observed, which could be between M. sextelata, M. eximia, and M. importuna, as well as between the three yellow morels.
Except for strain M115 of Mes-15, the 5′ and 3′ flanking sequences of the MAT1-1 and MAT1-2 idiomorphs harbored the same genes, i.e., SDH2 and MBA1, which were highly conserved across the nine other species. The BLASTn searchers against the M115 genome were run using the SDH2 and MBA1 genes as the query. These searches indicated that SDH2 was only in the scaffold carrying the MAT1-1 idiomorph, while MBA1 was in the scaffold harboring the MAT1-2 idiomorph.
The orientation of the mating genes was well-conserved in these morel species. When the mating-type locus was oriented toward the direction of the MAT1-2-1 gene, the MAT1-1-1 gene was always arranged in the same direction. However, the MAT1-1-10 and MAT1-1-11 genes, as well as the flanking genes SDH2 and MBA1, were oriented in the opposite direction as the MAT1-2-1 gene.
3.3. Unconventional Integration of The MAT Loci in M. purpurascena and M. pulchella
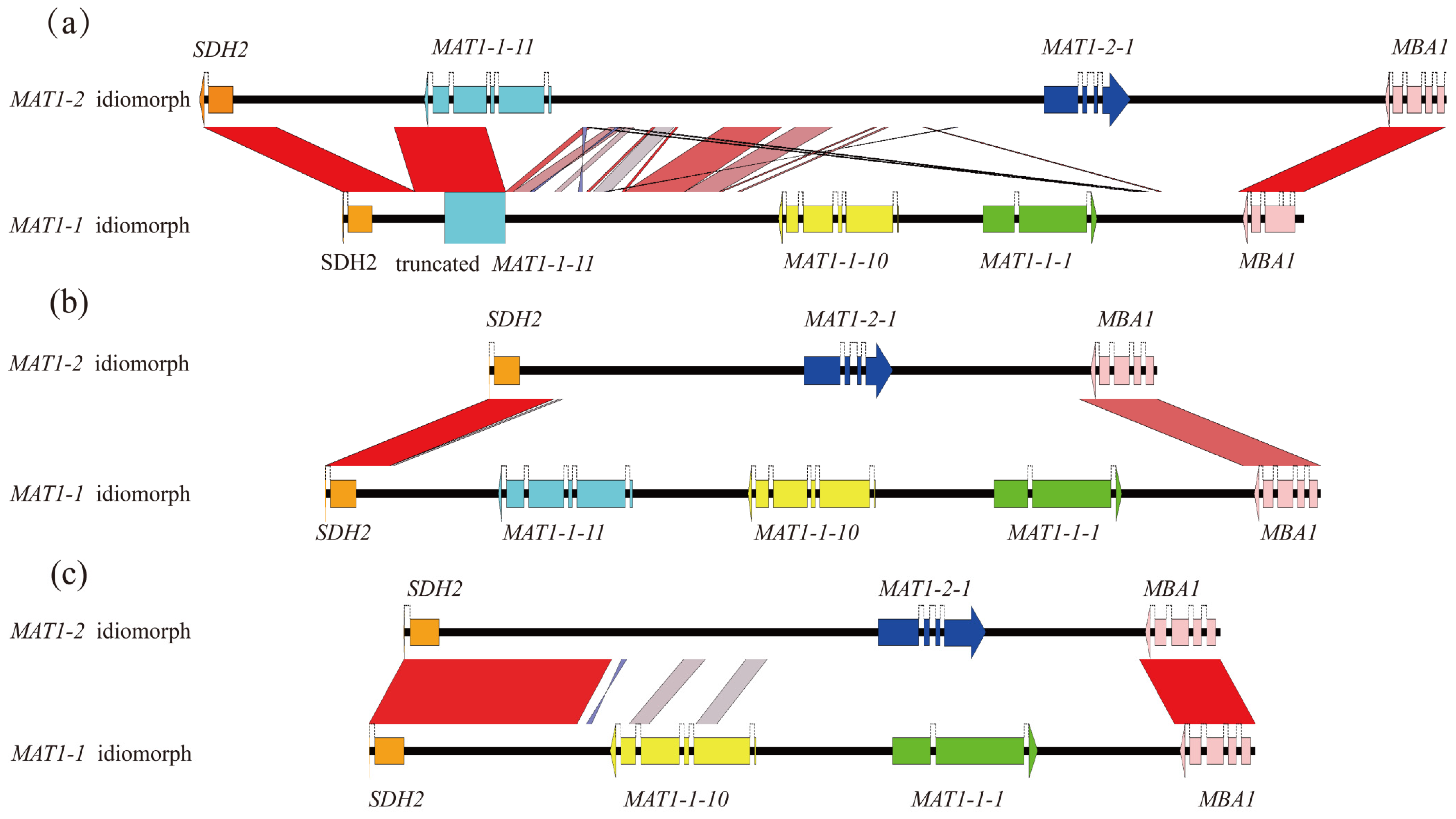
The alignment analyses in M. purpurascena and M. pulchella revealed a complete MAT1-1-11 gene of 1838 and 1828 bp in length, respectively, located in the MAT1-2 idiomorph between the SDH2 and MAT1-2-1 gene. On the other hand, the MAT1-1 idiomorph carried a truncated MAT1-1-11 fragment of a length of around 900 bp. A comparison of the sequences with the MAT1-1-11 of the MAT1-2 idiomorph from the same species indicated that about 900 bp were missing at the 5′ end of the gene in the MAT1-1 idiomorph, although there was 97% sequence similarity between the complete gene and the truncated sequences. Analyses of synteny revealed several homologous regions in the flanking regions of the MAT1-1-11 between the MAT1-1 and MAT1-2 idiomorphs (Figure 3a), unlike that in the M. importuna group, where there was a lack of similar sequences between the opposite-mating idiomorphs (Figure 3b). We hypothesized that unconventional integration occurred at the MAT loci between the opposite-mating type idiomorphs of M. purpurascena and M. pulchella. The complete MAT1-1-11 in the MAT1-2 idiomorphs were probably acquired from the MAT1-1 idiomorph, partial fragments were retained in the MAT1-1 idiomorph, and the sizes of the MAT1-2 idiomorphs of these two species were increased to 15.9 kb. Similar events were also reported in other ascomycetes [21,22]. In seven species in the family Ophiostomatales, an ancient inversion event resulted in the integration of the ancestral MAT1-1-1 gene into the MAT1-2 idiomorph and survival as the truncated MAT1-1-1 gene [21]. For the apple cancer pathogen Valsa mali, two flanking genes, COX13 and APN2, were co-opted into the MAT locus and unconventional recombination occurred between MAT1-1 and MAT1-2 idiomorphs, resulting in reverse insertion in the MAT1-2 idiomorph [22].
Phylogenetic analyses indicated that M. sextelata (Mel-6), M. importuna (Mel-10), and M. exuberans (Mel-9) represented three species lineages of an early evolutionary origin in China, whereas the species lineages of another 27, including M. purpurascena (Mel-20) and M. pulchella (Mel-31), diversified between the middle-Miocene and the present [12]. Therefore, the structure of the mating idiomorphs of the M. importuna type should be of earlier origin in the Elata clade species, and unconventional integration of the MAT locus was observed in M. purpurascena and M. pulchella due to their evolution from species lineages in the Elata clade. However, to confirm this hypothesis, more information on the MAT loci from additional morel species will be necessary.
Some small fragments with sequences that were homologous to the MAT1-1-10 gene were observed in the regions between the SDH2 and MAT1-2-1 in the MAT1-2 idiomorph of four yellow morel species (Figure 3c). The sizes and positions of homologous fragments differed according to the species, while the sequence identities of the MAT1-1-10 gene varied from 60% to 80%. Despite their homology to the MAT1-1-10 gene, these sequences neither coded for any proteins nor contained ORFs, as a start codon was absent.
3.4. Transcription Analyses of The Mating Genes
RT-PCR analyses revealed that four mating genes, MAT1-1-1, MAT1-1-10, MAT1-1-11, and MAT1-2-1, were constitutively expressed in the tested strains incubated on PDA for 10 days. The MAT1-1-11 gene could be successfully amplified with cDNA from the MAT1-1 mating-type strains of M. sextelata, M. eximia, and M. importuna, as well as cDNA from the MAT1-2 mating type strains of M. purpurascena and M. pulchella. However, the truncated MAT1-1-11 fragment could not be detected in the MAT1-1 mating type strains of M. purpurascena and M. pulchella, despite designing more primer pairs to amplify it (Supplementary Table S1).
The coding regions of the mating genes were verified by comparing the RT-PCR sequences with the corresponding DNA sequences, and the encoded protein sequences were obtained (Table 3). For these ten morel species, the MAT1-1-1 gene varied in length from 1645 to 1766 bp in lengths, which had two introns and encoded proteins of 509 to 547 amino acids, respectively. The sizes of the MAT1-2-1 gene varied from 1197 to 1343 bp, while the ORFs carried three introns and the coded proteins contained 329 to 384 amino acids. Both mating genes were longer in four yellow morels and M. rufobrunnea than in the five black morel species, which was also the case with the numbers of encoded amino acids. The MAT1-1-11 gene, harboring five introns, had similar lengths and numbers of encoded amino acids in the five black morel species.
3.5. Alternative Splicing of The MAT1-1-10 Gene
The validation of the coding regions of the MAT1-1-10 gene revealed some interesting phenomena, wherein this gene underwent alternative splicing (AS) to produce different splice variants, and the types of AS varied between species. Among all AS events in the MAT1-1-10 gene, there were two main types, intron retention and alternative 5′ or 3′ splice sites (Figure 4b,c), with the regular splicing mode (Figure 4a). Intron retention is the most common type of AS in fungi [23]. In this study, except for the third intron, the other four introns in the MAT1-1-10 gene were retained (Figure 4b), and the retention of the fifth intron (B4) was the most frequent type of AS in the MAT1-1-10 gene. In some species, for example, in M. sextelata, combinations of four types of intron retention were presented (Table 4), which revealed that the second, fourth, and fifth introns were retained alone or together, while the retention of the second intron resulted in premature termination of translation, so the kinds of combination including this retention could silence the gene.
Another type of AS in the MAT1-1-10 gene was the presence of alternative 5′ or 3′ splice sites, which were present in all except the fourth intron (Figure 4c), and all the splice sites were canonical 5′ GU.....3′AG. In the second intron, alternative 5′ and 3′ splicing was observed whereas, in the first, third, and fifth introns, only one type occurred. In Mes-6, there were two alternative 5′ splice sites in the second intron, and both RNA splice products differed by three amino acids. In some species, alternative 5′ or 3′ splice sites also induced gene silencing (Table 4). Besides, in M. sextelata and M. rufobrunnea, alternative 3′ splice sites were observed in the third intron of the MAT1-2-1 gene, and both RNA splice products resulted in the premature termination of translation.
To date, the MAT1-1-10 gene was only observed in the Morchella species, and its biological function was unknown. During vegetative growth, this gene could normally express and undergo AS to produce different mRNAs that encoded proteins of different sizes. We hypothesize that this mating gene could play an important role during the asexual phase.
4. Discussion
4.1. Mating Systems in Morchella
The characteristics of the mating idiomorphs from five black morel species, M. sextelata, M. eximia, M. importuna, M. purpurascena, and M. pulchella, and three yellow morel species, Mes-6, Mes-19, and Mes-20, indicated heterothallism. The single-ascospore isolates carried either the MAT1-1 or the MAT1-2 idiomorph. Furthermore, the two idiomorphs had the conserved flanking sequences (SDH2 and MAB1), which confirmed that they occupied the same chromosomal locus. Nevertheless, in M. rufobrunnea from the Rufobrunnea clade, the mating genes MAT1-1-10, MAT1-1-1, and MAT1-2-1 were linked and contained in the haploid genome, while the mating locus also had the conserved flanking genes SDH2 and MAB1, which indicated that M. rufobrunnea was homothallic.
For Mes-15, the mating genes MAT1-1-10, MAT1-1-1, and MAT1-2-1 were contained in the single-ascospore strain M115 genome. However, the MAT1-1-10 and MAT1-1-1 genes were in scaffold 677 that carried four genes, viz., CPSF6, TFA1, ATP4, and SDH2 in the 5′ flanking regions like the structures observed in M. importuna and Mes-20 [16,17], except for the absence of MBA1 from the 3′ flanking sequences. Similarly, the MAT1-2-1 and two 3′-flanking genes, MBA1 and SLA2, were located in another scaffold 700, although SDH2 was not found in the 5′ flanking regions. We could not determine whether Mes-15 was a homothallic or pseudohomothallic species, however, the genome data from a yellow morel species, M. peruviana (JGI Project Id 1239452), gave some clues. In this genome, the mating genes MAT1-1-10, MAT1-1-1, and MAT1-2-1 were present in scaffold 12, while the adjacent genes SDH2 and MBA1 were located in the 5′ and 3′ flanking regions, respectively, but the distance between the MAT1-1-1 and MAT1-2-1 was over 38 kb. These features indicated that M. peruviana was homothallic. Analysis of synteny indicated that the scaffold 677 of strain M115 was highly homologous with the MAT1-1-1 region of the scaffold 12 of M. peruviana, while the scaffold 700 shared identity with the MAT1-2-1 region (Supplementary Figure S1). Based on these findings, we hypothesized that Mes-15 was a homothallic species, and the MAT1-1-1 and MAT1-2-1 genes were not tightly linked, like in M. peruviana.
By analyzing the mating-type ratios of the single-spore populations, Chai et al. [17] considered three black morel species, M. sextelata, M. importuna, and Mel-20, to show heterothallism. Du and Yang [16] reported that 15 black morel species and 19 yellow morel species were heterothallic, and among the 38 morel species studied, the MAT1-1-10 gene could be successfully amplified in 34 species except for four black morel species, whereas the MAT1-1-11 gene was found in only seven black morel species. In contrast, recent studies have reported that M. importuna was a pseudohomothallic species [24]. The single-ascospore strain of M. importuna could fructify normally [24,25,26,27]. Furthermore, there was a great deviation in the proportion of two opposite mating-type nuclei in the single-ascospore isolates, and it was sometimes difficult to detect the mating-type with less distribution by PCR amplification. Therefore, for other Morchella species, more detailed research may be required to confirm heterothallism or pseudohomothallism.
Whether homothallism or heterothallism is the ancestral reproductive state in ascomycetes is still a controversial issue [28,29]. In Morchella, the Rufobrunnea clade consisting of M. rufobrunnea and M. Anatolica, demonstrate the earliest diverging branch, followed by the origin of the Esculenta and Elata sister clades from approximately 30 million years later [11,12]. Therefore, the presentation of homothallism in M. rufobrunnea should be the ancestral reproductive state in Morchella. During evolution, some species have retained homothallic modes, whereas others have turned to heterothallism, as heterothallism would offer the advantages of enhanced genetic diversity and adaption to the environment [30]. The MAT1-1-10 gene was the primary mating gene in Morchella, like the MAT1-1-1 and MAT1-2-1 genes, and the MAT1-1-11 gene was integrated into the mating loci during the process of evolution. However, further research is needed to confirm whether this gene was distributed in all species or only in some species from the Elata clade.
4.2. Unconventional Integration of the MAT Locus in Some Morel Species
Generally, the mating-type loci in fungi have often evolved extensive suppression of recombination [31]. However, unconventional recombination or crossover at the MAT locus between the opposite mating types has also been demonstrated in several ascomycetes [21,22]. In Botryosphaeriaceae, some heterothallic species had MAT1-2-5 fragments in the MAT1-1 idiomorph. Furthermore, they showed a truncated MAT1-1-8 gene in the MAT1-2 idiomorph [28]. In some Phyllosticta species, a truncated version of the MAT1-1-8 gene was in the MAT1-2 strains [32]. It has been proposed that these partial gene fragments in the mating-type locus of heterothallic strains can arise either from their heterothallic ancestors that underwent unequal crossing over/recombination events or from their homothallic ancestors that experienced two independent deletion events [28,32].
We hypothesized that unconventional integration occurred at the MAT locus of M. purpurascena and M. pulchella. Phylogenetic analyses indicated that the divergence time of M. sextelata and M. importuna was earlier than that of M. purpurascena and M. pulchella [12]. Therefore, the structure of mating idiomorphs of the M. importuna group-the MAT1-1-1, MAT1-1-10, and MAT1-1-11 in the MAT1-1 idiomorph and only the MAT1-2-1 in the MAT1-2 idiomorph-should be of a relatively older origin in the species from the Elata clade. During evolution, unclear integration events occurred between the opposite idiomorphs in M. purpurascena and M. pulchella, the MAT1-1-11 gene was restructured into the MAT1-2 idiomorph, and the partial fragment was retained in the MAT1-1 idiomorph, giving rise to the mating idiomorph of the M. purpurascena type.
Based on the analyses of the genome data of some morel species submitted to JGI, we observed that the mating idiomorph of the M. importuna type in M. semilibera (Mel-3), M. punctipes (Mel-4), and M. disparilis, but some other species including M. snyderi (Mel-12), M. angusticeps (Mel-15), M. hispaniolensis (Mel-18), M. eohespera (Mel-19), and M. brunnea (Mel-22), had the mating idiomorph of the M. purpurascena type. Therefore, the integration events had not happened by accident in some species but had occurred through evolution from species lineages.
4.3. Alternative Splicing of The MAT1-1-10 Gene
Alternative splicing is a regulated process in eukaryotes, which leads to expansion in the form and function of the proteome [33]. It occurs in 95–100% of human genes [33], 60% of plant genes [34], 8.6% of Basidiomycota, and 6.0% of Ascomycota genes [35]. However, recent studies indicate that AS occurs in fungi at a much higher frequency than reported earlier [36]. For example, rates of AS in Trichoderma longibrachiatum and Cryptococcus neoformans were 48.9% [37] and 59% [38], respectively. AS serves several regulatory functions in fungi, including environmental adaptation, protein localization, meiosis, and gene expression [36]. Genes with regulatory functions, such as transcription factors or signal transducers exhibit high levels of alternative splicing, while basic enzymatic functions show less splice variation [39,40].
The MAT1-1-10 was an ancestral mating gene in Morchella, which was reported in recent years [7], and little was known about its functions. In this study, the alternative splicing phenomenon of this gene was observed in 10 morel species from three clades, leading to different mRNAs that encoded proteins with different amino acid lengths. AS occurred in the mycelia that were incubated in normal growing conditions and were not subjected to environmental pressure, and it is unknown whether these splice variants would exhibit different biological functions during vegetative growth, which has also aroused our curiosity about the roles of the MAT1-1-10 gene.
5. Conclusions
The results from our study showed that homothallism should be the ancestral reproductive state in Morchella. The mating-type loci from ten morel species have been characterized and compared. It confirmed that M. rufobrunnea, a representative species of the earliest diverging branch of true morels, along with another yellow morel Mes-15, were homothallic, and another eight morel species were heterothallic. Three types of composition were observed in the mating idiomorphs of eight heterothallic species, and an unconventional integration event occurred in two black morel species and resulted in the importation of the MAT1-1-11 gene into the MAT1-2 idiomorph and survival as the truncated gene. The primer pairs p11-1f/p11-2r could successfully amplify the mating-type idiomorphs of heterothallic species from the Elata and Esculenta clades, which make it possible to obtain more information on the MAT loci from additional morel species by PCR.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof8070746/s1, Table S1: Primers used in this study. Figure S1: Comparison of the scaffolds including the MAT loci between M. peruviana and Mes-15.
Author Contributions
Conceptualization, Y.Z. and H.C.; methodology, P.L., Y.M. and N.T.; formal analysis, H.C. and W.C.; investigation, Y.M., P.L. and N.T.; resources, Y.Z. and P.L.; writing original draft preparation, H.C. and P.L.; writing—review and editing, H.C. and Y.Z.; project administration, H.C. and Y.Z.; funding acquisition, H.C. and Y.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Nature Science Foundation of China (No. 31960621; No. 31460014), China Agriculture Research System of MOF and MARA (CARS-20).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The MAT1-1 idiomorph sequences of M. sextelata, M. eximia, M. importuna, M. purpurascena, M. pulchella, Mes-6, Mes-15, Mes-19, and Mes-20 can be found at GenBank accession numbers MN589921, ON622485, KY782630, MN589929, ON622483, MN589927, KY782631, MN589925, MN589923. The MAT1-2 idiomorph sequences of each species can be found at GenBank accession numbers MN589922, ON622484, KY782629, MN589930, ON622482, MN589928, KY782632, MN589926, MN589924. The MAT locus sequences of M. rufobrunnea can be found at GenBank accession number MN589931. The genome data of Mes-15 was under accession number GWHBJCX00000000 in the Genome Warehouse in the National Genomics Data Center.
Acknowledgments
Thanks to the US Department of Energy Joint Genome Institute https://www.jgi.doe.gov/ in JGI Project Id 1229104 (accessed on 25 October, 2018); 1229100 (accessed on 25 October, 2018); 1229027 (accessed on 25 October 2018); 1239448(accessed on 12 January 2019); 1229120 (accessed on 25 October 2018); 1239452 (accessed on 12 January 2019); 1229096 (accessed on 25 October 2018); 1229080 (accessed on 25 October 2018); 1133164 (accessed on 3 December 2016) collaboration with the user community, which produced the genome data of some morel species used in the discussion.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chitrampalam, P.; Pryor, B.M. Characterization of mating type (MAT) alleles differentiated by a natural inversion in Sclerotinia minor. Plant Pathol. 2015, 64, 911–920. [Google Scholar] [CrossRef]
- Debuchy, R.; Turgeon, B.G. Mating-type structure, evolution, and function in Euascomycetes. In The Mycota; Springer: Berlin/Heidelberg, Germany, 2006; Volume 1, pp. 293–323. [Google Scholar]
- Metzenberg, R.L.; Glass, N.L. Mating type and mating strategies in Neurospora. BioEssays 1990, 12, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Turgeon, B.G.; Yoder, O.C. Proposed nimenclature for mating type genes of filamentous ascomycetes. Fungal Genet. Biol. 2000, 31, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Debuchy, R.; Berteauxlecellier, V.; Silar, P. Mating systems and sexual morphogenesis in Ascomycetes. In Cellular and Molecular Biology of Filamentous Fungi; Borkovich, K.A., Ebbole, E., Eds.; ASM Press: Washington, DC, USA, 2010; pp. 201–535. [Google Scholar]
- Wilken, P.M.; Steenkamp, E.T.; Wingfield, M.J.; De Beer, Z.W.; Wingfield, B.D. Which MAT gene? Pezizomycotina (Ascomycota) mating-type gene nomenclature reconsidered. Fungal Biol. Rev. 2017, 31, 199–211. [Google Scholar] [CrossRef] [Green Version]
- Chai, H.M.; Chen, W.M.; Zhang, X.L.; Su, K.M.; Zhao, Y.C. Structural variation and phylogenetic analysis of the mating-type locus in the genus Morchella. Mycologia 2019, 111, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Aylward, J.; Havenga, M.; Dreyer, L.L.; Roets, F.; Wingfield, B.D.; Wingfield, M.J. Genomic characterization of mating type loci and mating type distribution in two apparently asexual plantation tree pathogens. Plant Pathol. 2020, 69, 28–37. [Google Scholar] [CrossRef]
- Wilson, A.M.; Wilken, P.M.; van der Nest, M.A.; Steenkamp, E.T.; Wingfield, M.J.; Wingfield, B.D. Homothallism: An umbrella term for describing diverse sexual behaviours. IMA Fungus 2015, 6, 207–214. [Google Scholar] [CrossRef]
- Kuo, M.; Dewsbury, D.R.; O’Donnell, K.; Carter, M.C.; Rehner, S.A.; Moore, J.D.; Moncalvo, J.M.; Canfield, S.A.; Stephenson, S.L.; Methven, A.S.; et al. Taxonomic revision of true morels (Morchella) in Canada and the United States. Mycologia 2012, 104, 1159–1177. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Rooney, A.P.; Mills, G.L.; Kuo, M.; Weber, N.S.; Rehner, S.A. Phylogeny and historical biogeography of true morels (Morchella) reveals an early Cretaceous origin and high continental endemism and provincialism in the Holarctic. Fungal Genet. Biol. 2011, 48, 252–265. [Google Scholar] [CrossRef]
- Du, X.H.; Zhao, Q.; O’Donnell, K.; Rooney, A.P.; Yang, Z.L. Multigene molecular phylogenetics reveals true morels (Morchella) are especially species-rich in China. Fungal Genet. Biol. 2012, 49, 455–469. [Google Scholar] [CrossRef]
- Loizides, M.; Alvarado, P.; Moreau, P.A.; Assyov, B.; Halasu, V.; Stadler, M.; Rinaldi, A.; Guilhermina, M.; Georgios, I.Z.; Borovička, J.; et al. Has taxonomic vandalism gone too far? A case study, the rise of the pay-to-publish model and the pitfalls of Morchella systematics. Mycol. Prog. 2022, 21, 7–38. [Google Scholar] [CrossRef]
- Belfiori, B.; Riccioni, C.; Paolocci, F.; Rubini, A. Characterization of the reproductive mode and life cycle of the whitish truffle T. Borchii. Mycorrhiza 2016, 26, 515–527. [Google Scholar] [CrossRef]
- Lu, Y.H.; Xia, Y.L.; Luo, F.F.; Dong, C.H.; Wang, C.S. Functional convergence and divergence of mating-type genes fulfilling in Cordyceps militaris. Fungal Genet. Biol. 2016, 88, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Du, X.H.; Yang, Z.L. Mating Systems in True Morels (Morchella). Microbiol. Mol. Biol. Rev. 2021, 85, e00220-20. [Google Scholar] [CrossRef]
- Chai, H.M.; Chen, L.J.; Chen, W.M.; Zhao, Q.; Zhang, X.L.; Su, K.M.; Zhao, Y.C. Characterization of mating-type idiomorphs suggests that Morchella importuna, Mel-20 and M. sextelata are heterothallic. Mycol. Prog. 2017, 16, 743–752. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Martin, S.H.; Wingfield, B.D.; Wingfield, M.J.; Steenkamp, E.T. Structure and evolution of the Fusarium mating type locus: New insights from the Gibberellafujikuroi complex. Fungal Genet. Biol. 2011, 48, 734–740. [Google Scholar] [CrossRef] [Green Version]
- Tsui, C.K.M.; DiGuistini, S.; Wang, Y.; Feau, N.; Dhillon, B.; Bohlmann, J.; Hamelin, R.C. Unequal recombination and evolution of the mating-type (MAT) loci in the pathogenic fungus Grosmannia clavigera and relatives. Genes Genomes Genet. 2013, 3, 465–480. [Google Scholar]
- Yin, Z.Y.; Ke, X.W.; Li, Z.P.; Chen, J.L.; Gao, X.L.; Huang, L.L. Unconventional Recombination in the Mating Type Locus of Heterothallic Apple Canker Pathogen Valsa mali. Genes Genomes Genet. 2017, 7, 1259–1265. [Google Scholar] [CrossRef] [Green Version]
- Kempken, F. Alternative splicing in ascomycetes. Appl. Microbiol. Biotechnol. 2013, 97, 4235–4241. [Google Scholar] [CrossRef] [PubMed]
- Chai, H.M.; Ma, Y.H.; Liu, P.; Chen, W.M.; Tao, N.; Zhao, Y.C. The asymmetrical distribution of opposite mating type nuclei in single-ascospore isolates revealed Morchella importuna is a pseudohomothallic fungus. Mycosystema 2022. in-print. Available online: https://kns.cnki.net/kcms/detail/11.5180.Q.20220419.1112.002.html (accessed on 14 June 2022).
- He, X.S.; Zhao, M.; Wang, Y.Z.; Zhang, N.; Tan, R.H.; Wang, Y.; Zhu, W.K.; Chen, B. Study on the hyphal fusion and non-fusion phenomena between morels. Edible Med. Mushrooms 2019, 27, 244–252. [Google Scholar]
- Liu, W.; Cai, Y.L.; He, P.X.; Bian, Y.B. Cultivation of monosporic and hybrid populations and polarity analysis of Morchella importuna. J. Fungal Res. 2019, 17, 43–49. [Google Scholar]
- Tan, F.H. Expounding the development period of morel cultivation technology using exogenous nutrition bag in China. Edible Med. Mushrooms 2019, 27, 257–263. [Google Scholar]
- Nagel, J.H.; Wingfield, M.J.; Slippers, B. Evolution of the mating types and mating strategies in prominent genera in the Botryosphaeriaceae. Fungal Genet. Biol. 2018, 114, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Li, J.Q.; Wingfield, B.D.; Wingfield, M.J.; Barnes, I.; Fourie, A.; Crous, P.W.; Chen, S.F. Mating genes in Calonectria and evidence for a heterothallic ancestral state. Persoonia 2020, 45, 163–176. [Google Scholar] [CrossRef]
- Lumley, A.J.; Michalczyk, L.; Kitson, J.J.; Spurgin, L.G.; Morrison, C.A.; Godwin, J.L.; Dickinson, M.E.; Martin, O.Y.; Emerson, B.C.; Chapman, T.; et al. Sexual selection protects against extinction. Nature 2015, 522, 470–473. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, F.E.; Duhamel, M.; Carpentier, F.; Hood, M.E.; Foulongne-Oriol, M.; Silar, P.; Malagnac, F.; Grognet, P.; Giraud, T. Recombination suppression and evolutionary strata around mating-type loci in fungi: Documenting patterns and understanding evolutionary and mechanistic causes. New Phytol. 2021, 229, 2470–2491. [Google Scholar] [CrossRef]
- Petters-Vandresen, D.A.; Rossi, B.J.; Groenewald, J.Z.; Crous, P.W.; Machado, M.A.; Stukenbrock, E.H.; Glienke, C. Mating-type locus rearrangements and shifts in thallism states in Citrus associated Phyllosticta species. Fungal Genet. Biol. 2020, 144, 103444. [Google Scholar] [CrossRef]
- Lee, Y.; Rio, D.C. Mechanisms and Regulation of Alternative Pre-mRNA Splicing. Annu. Rev. Biochem. 2015, 84, 291–323. [Google Scholar] [CrossRef] [Green Version]
- Sablok, G.; Powell, B.; Braessler, J.; Yu, F.; Min, X.J. Comparative landscape of alternative splicing in fruit plants. Curr. Plant Biol. 2017, 9, 29–36. [Google Scholar] [CrossRef]
- Grutzmann, K.; Szafranski, K.; Pohl, M.; Voigt, K.; Petzold, A.; Schuster, S. Fungal alternative splicing is associated with multicellular complexity and virulence: A genome-wide multi-species study. DNA Res. 2014, 21, 27–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muzafar, S.; Sharma, R.D.; Chauhan, N.; Prasad, R. Intron distribution and emerging role of alternative splicing in fungi. FEMS Microbiol. Lett. 2021, 368, fnab135. [Google Scholar] [CrossRef]
- Xie, B.B.; Li, D.; Shi, W.L.; Qin, Q.L.; Wang, X.W.; Rong, J.C.; Sun, C.Y.; Huang, F.; Zhang, X.Y.; Dong, X.W.; et al. Deep RNA sequencing reveals a high frequency of alternative splicing events in the fungus Trichoderma longibrachiatum. BMC Genom. 2015, 16, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Hilarion, S.; Paulet, D.; Lee, K.T.; Hon, C.C.; Janbon, G. Intron retention-dependent gene regulation in Cryptococcus neoformans. Sci. Rep. 2016, 6, 32252. [Google Scholar] [CrossRef]
- Irimia, M.; Rukov, J.L.; Penny, D.; Roy, S.W. Functional and evolutionary analysis of alternatively spliced genes is consistent with an early eukaryotic origin of alternative splicing. BMC Evol. Biol. 2007, 7, 188. [Google Scholar] [CrossRef] [Green Version]
- Zavolan, M.; Kondo, S.; Schonbach, C.; Adachi, J.; Hume, D.A.; RIKEN GER Group and GSL Members; Hayashizaki, Y.; Gaasterland, T. Impact of alternative initiation, splicing, and termination on the diversity of the mRNA transcripts encoded by the mouse transcriptome. Genome Res. 2003, 13, 1290–1300. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Agarose gel electrophoresis of PCR products of the mating idiomorphs. Lane 1, 3, 5, 7, 9, 11: PCR products from the MAT1-2 type strains YAASMQM-4, YAASMHL47, YAASM23-22, YAASM131-12, YAASM50-09, and YAASM43-08; Lane 2, 4, 6, 8, 10, 12: PCR products from the MAT1-1 type strains YAASMQM-23, YAASMHL1, YAASM23-11, YAASM131-9, YAASM50-38, and YAASM43-24; Lane 13: PCR products from strain YAASMVR; M1: DL10000 DNA Marker (10,000, 7000, 4000, 2000, 1000, 500, 250); M2: DL15000 DNA Marker (15,000, 10,000, 7500, 5000, 2500, 1000, 250).
Figure 1.
Agarose gel electrophoresis of PCR products of the mating idiomorphs. Lane 1, 3, 5, 7, 9, 11: PCR products from the MAT1-2 type strains YAASMQM-4, YAASMHL47, YAASM23-22, YAASM131-12, YAASM50-09, and YAASM43-08; Lane 2, 4, 6, 8, 10, 12: PCR products from the MAT1-1 type strains YAASMQM-23, YAASMHL1, YAASM23-11, YAASM131-9, YAASM50-38, and YAASM43-24; Lane 13: PCR products from strain YAASMVR; M1: DL10000 DNA Marker (10,000, 7000, 4000, 2000, 1000, 500, 250); M2: DL15000 DNA Marker (15,000, 10,000, 7500, 5000, 2500, 1000, 250).

Figure 2.
Pairwise comparison of the mating idiomorphs between species of Morchella. The black horizontal lines represent genomic sequences, and the color-coded arrows represent coding sequences in the forward (right-oriented arrow) or reverse (left-oriented arrow) strand. The red or blue boxes between genomic sequences indicate pairwise similarity based on BLASTn; red boxes indicate that both regions are in the same orientation, while blue boxes indicate that the regions are oriented in the opposite directions. Neighbor-joining (NJ) phylogenetic analysis between Morchella species based on a multi-locus dataset (ITS, RPB1, RPB2, LSU, and EF1-a).
Figure 2.
Pairwise comparison of the mating idiomorphs between species of Morchella. The black horizontal lines represent genomic sequences, and the color-coded arrows represent coding sequences in the forward (right-oriented arrow) or reverse (left-oriented arrow) strand. The red or blue boxes between genomic sequences indicate pairwise similarity based on BLASTn; red boxes indicate that both regions are in the same orientation, while blue boxes indicate that the regions are oriented in the opposite directions. Neighbor-joining (NJ) phylogenetic analysis between Morchella species based on a multi-locus dataset (ITS, RPB1, RPB2, LSU, and EF1-a).

Figure 3.
Comparison of the opposite-mating idiomorphs from the same species. Comparison of the opposite-mating idiomorphs of (a) M. pulchella, (b) M. eximia, (c) Mes-19. Black horizontal lines represent genomic sequences, and color-coded arrows represent coding sequences. Red or blue boxes between genomic sequences indicate pairwise similarity based on BLASTN. Red boxes indicate that both regions are in the same orientation, and blue boxes indicate that the regions are oriented in opposite directions.
Figure 3.
Comparison of the opposite-mating idiomorphs from the same species. Comparison of the opposite-mating idiomorphs of (a) M. pulchella, (b) M. eximia, (c) Mes-19. Black horizontal lines represent genomic sequences, and color-coded arrows represent coding sequences. Red or blue boxes between genomic sequences indicate pairwise similarity based on BLASTN. Red boxes indicate that both regions are in the same orientation, and blue boxes indicate that the regions are oriented in opposite directions.

Figure 4.
A schematic diagram showing the different types of AS events in the MAT1-1-10 gene. (a) Normal splicing. (b) Intron retention: B1, Retention of the first intron; B2, Retention of the second intron; B3, Retention of the fourth intron; B4, Retention of the fifth intron. (c) Alternative 5′ and 3′ splicing: C1, Alternative 3′ splicing in the first intron; C2, Alternative 5′ splicing in the second intron; C3, Alternative 3′ splicing in the second intron; C4, Alternative 3′ splicing in the third intron; C5, Alternative 5′ splicing in the fifth intron.
Figure 4.
A schematic diagram showing the different types of AS events in the MAT1-1-10 gene. (a) Normal splicing. (b) Intron retention: B1, Retention of the first intron; B2, Retention of the second intron; B3, Retention of the fourth intron; B4, Retention of the fifth intron. (c) Alternative 5′ and 3′ splicing: C1, Alternative 3′ splicing in the first intron; C2, Alternative 5′ splicing in the second intron; C3, Alternative 3′ splicing in the second intron; C4, Alternative 3′ splicing in the third intron; C5, Alternative 5′ splicing in the fifth intron.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The morel strains used in this study.
| Strain | Species | Code | Ascocarp | Origin | Mating-Type Idiomorph |
|---|---|---|---|---|---|
| YAASMHL1 | M. sextelata | Mel-6 | YAASMHL | Sichuan, China | MAT1-1 |
| YAASMHL47 | MAT1-2 | ||||
| YAASMQM-23 | M. eximia | Mel-7 | YAASMQM | Tibet, China | MAT1-1 |
| YAASMQM-4 | MAT1-2 | ||||
| YAASM23-11 | M. purpurascena | Mel-20 | YAASM23 | Yunnan, China | MAT1-1 |
| YAASM23-22 | MAT1-2 | ||||
| YAASM131-9 | M. pulchella | Mel-31 | YAASM131 | Yunnan, China | MAT1-1 |
| YAASM131-12 | MAT1-2 | ||||
| YAASM50-38 | Mes-6 | YAASM50 | Shanxi, China | MAT1-1 | |
| YAASM50-09 | MAT1-2 | ||||
| M115 | Mes-15 | YAASM2689 | Yunnan, China | MAT1-1, MAT1-2 | |
| YAASM43-24 | Mes-19 | YAASM43 | Sichuan, China | MAT1-1 | |
| YAASM43-08 | MAT1-2 | ||||
| YAASMVR | M. rufobrunnea | American | MAT1-1, MAT1-2 |
Table 2.
Organization of the mating idiomorphs.
| Species | Code | MAT1-1 Idiomorph | MAT1-2 Idiomorph | ||||
|---|---|---|---|---|---|---|---|
| Gene Arrangements | Lengths (kb) | GenBank Accession | Gene Arrangements | Lengths (kb) | GenBank Accession | ||
| M. sextelata | Mel-6 | MAT1-1-1, MAT1-1-10, MAT1-1-11 | 11.5 | MN589921 | MAT1-2-1 | 7.1 | MN589922 |
| M. eximia | Mel-7 | MAT1-1-1, MAT1-1-10, MAT1-1-11 | 10.2 | ON622485 | MAT1-2-1 | 6.6 | ON622484 |
| M. importuna | Mel-10 | MAT1-1-1, MAT1-1-10, MAT1-1-11 | 10.5 | KY782630 | MAT1-2-1 | 6.7 | KY782629 |
| M. purpurascena | Mel-20 | MAT1-1-1, MAT1-1-10, truncated MAT1-1-11 | 9.4 | MN589929 | MAT1-2-1, MAT1-1-11 | 15.9 | MN589930 |
| M. pulchella | Mel-31 | MAT1-1-1, MAT1-1-10, truncated MAT1-1-11 | 11.8 | ON622483 | MAT1-2-1, MAT1-1-11 | 15.9 | ON622482 |
| Mes-6 | MAT1-1-1, MAT1-1-10 | 8.0 | MN589927 | MAT1-2-1 | 7.3 | MN589928 | |
| Mes-15 | MAT1-1-1, MAT1-1-10 | - | KY782631 | MAT1-2-1 | - | KY782632 | |
| Mes-19 | MAT1-1-1, MAT1-1-10 | 7.2 | MN589925 | MAT1-2-1 | 6.4 | MN589926 | |
| Mes-20 | MAT1-1-1, MAT1-1-10 | 7.8 | MN589923 | MAT1-2-1 | 7.5 | MN589924 | |
| M. rufobrunnea | MAT1-1-1, MAT1-1-10, MAT1-2-1 | 16.7 | MN589931 | ||||
Table 3.
The variance of the mating genes in ten morel species.
| Species | Code | MAT1-1-1 | MAT1-2-1 | MAT1-1-11 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| DNA (bp) | Intron Numbers | Amino Acids (aa) | DNA (bp) | Intron Numbers | Amino Acids (aa) | DNA (bp) | Intron Numbers | Amino Acids (aa) | ||
| M. sextelata | Mel-6 | 1730 | 2 | 537 | 1199 | 3 | 329 | 1823 | 5 | 511 |
| M. eximia | Mel-7 | 1676 | 2 | 519 | 1203 | 3 | 330 | 1825 | 5 | 511 |
| M. importuna | Mel-10 | 1694 | 2 | 525 | 1197 | 3 | 329 | 1825 | 5 | 512 |
| M. purpurascena | Mel-20 | 1651 | 2 | 511 | 1247 | 3 | 344 | 1838 | 5 | 511 |
| M. pulchella | Mel-31 | 1645 | 2 | 509 | 1247 | 3 | 344 | 1828 | 5 | 511 |
| Mes-6 | 1757 | 2 | 547 | 1328 | 3 | 385 | ||||
| Mes-15 | 1766 | 2 | 543 | 1304 | 3 | 377 | ||||
| Mes-19 | 1751 | 2 | 545 | 1301 | 3 | 376 | ||||
| Mes-20 | 1751 | 2 | 545 | 1286 | 3 | 371 | ||||
| M. rufobrunnea | 1760 | 2 | 546 | 1343 | 3 | 384 | ||||
Table 4.
The types of alternative splicing in the MAT1-1-10 gene.
| Species | Code | Total Numbers of Clones | Types of Splicing a | Numbers of Clones | Intron Numbers | DNA (bp) | Amino Acids |
|---|---|---|---|---|---|---|---|
| M. sextelata | Mel-6 | 21 | A | 11 | 5 | 1723 | 482 |
| B4 | 3 | 4 | 1689 | 490 | |||
| B3 + B4 | 3 | 3 | 1689 | 510 | |||
| B2 + B4 | 1 | 3 | - b | - | |||
| B2 + B3 + B4 | 3 | 2 | - | - | |||
| M. eximia | Mel-7 | 10 | B4 | 10 | 4 | 1661 | 479 |
| M. importuna | Mel-10 | 10 | B4 | 10 | 4 | 1688 | 488 |
| M. purpurascena | Mel-20 | 10 | A | 7 | 5 | 1737 | 482 |
| C3 | 3 | 5 | 1737 | 486 | |||
| M. pulchella | Mel-31 | 10 | C3 | 6 | 5 | 1737 | 478 |
| C4 | 2 | 5 | 1737 | 482 | |||
| C3 + C4 | 2 | 5 | 1737 | 486 | |||
| Mes-6 | 10 | A | 2 | 5 | 1762 | 493 | |
| C1 | 1 | 5 | - | - | |||
| C1 + B4 | 1 | 4 | - | - | |||
| B4 | 2 | 4 | 1654 | 478 | |||
| C5 | 3 | 5 | 1704 | 473 | |||
| C5-2 c | 1 | 5 | 1733 | 476 | |||
| Mes-15 | 10 | A | 8 | 5 | 1760 | 491 | |
| C5 | 1 | 5 | - | - | |||
| C1 + C5 | 1 | 5 | - | - | |||
| Mes-19 | 14 | A | 11 | 5 | 1760 | 491 | |
| B4 | 3 | 4 | 1760 | 512 | |||
| Mes-20 | 10 | A | 1 | 5 | 1757 | 491 | |
| B4 | 6 | 4 | 1648 | 476 | |||
| C5 | 1 | 5 | 1699 | 420 | |||
| B1 + C5 | 1 | 4 | 1699 | 489 | |||
| B1 + B4 + C2 | 1 | 3 | 1649 | 474 | |||
| M. rufobrunnea | Mruf | 11 | B4 | 4 | 4 | 1698 | 485 |
| C3 | 3 | 5 | 1850 | 515 | |||
| C3 + B4 | 3 | 4 | 1698 | 489 | |||
| B1 + C3 | 1 | 4 | - | - |
a: Represented by the codes in Figure 4; b: Gene silencing; c: Another alternative 5′ splice site in the fifth intron.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chai, H.; Liu, P.; Ma, Y.; Chen, W.; Tao, N.; Zhao, Y. Organization and Unconventional Integration of the Mating-Type Loci in Morchella Species. J. Fungi 2022, 8, 746. https://doi.org/10.3390/jof8070746
AMA Style
Chai H, Liu P, Ma Y, Chen W, Tao N, Zhao Y. Organization and Unconventional Integration of the Mating-Type Loci in Morchella Species. Journal of Fungi. 2022; 8(7):746. https://doi.org/10.3390/jof8070746
Chicago/Turabian StyleChai, Hongmei, Ping Liu, Yuanhao Ma, Weimin Chen, Nan Tao, and Yongchang Zhao. 2022. "Organization and Unconventional Integration of the Mating-Type Loci in Morchella Species" Journal of Fungi 8, no. 7: 746. https://doi.org/10.3390/jof8070746
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.