
Plant Beneficial Bacteria as Bioprotectants against Wheat and Barley Diseases



Université de Reims Champagne-Ardenne, INRAE, RIBP USC 1488, SFR Condorcet, 51100 Reims, France
*
Author to whom correspondence should be addressed.
J. Fungi 2022, 8(6), 632; https://doi.org/10.3390/jof8060632
Submission received: 19 May 2022
/
Revised: 7 June 2022
/
Accepted: 9 June 2022
/
Published: 14 June 2022
(This article belongs to the Special Issue Plant Fungal Pathogens: Isolation, Characterization and Control Strategies)
Abstract
:Wheat and barley are the main cereal crops cultivated worldwide and serve as staple food for a third of the world’s population. However, due to enormous biotic stresses, the annual production has significantly reduced by 30–70%. Recently, the accelerated use of beneficial bacteria in the control of wheat and barley pathogens has gained prominence. In this review, we synthesized information about beneficial bacteria with demonstrated protection capacity against major barley and wheat pathogens including Fusarium graminearum, Zymoseptoria tritici and Pyrenophora teres. By summarizing the general insights into molecular factors involved in plant-pathogen interactions, we show to an extent, the means by which beneficial bacteria are implicated in plant defense against wheat and barley diseases. On wheat, many Bacillus strains predominantly reduced the disease incidence of F. graminearum and Z. tritici. In contrast, on barley, the efficacy of a few Pseudomonas, Bacillus and Paraburkholderia spp. has been established against P. teres. Although several modes of action were described for these strains, we have highlighted the role of Bacillus and Pseudomonas secondary metabolites in mediating direct antagonism and induced resistance against these pathogens. Furthermore, we advance a need to ascertain the mode of action of beneficial bacteria/molecules to enhance a solution-based crop protection strategy. Moreover, an apparent disjoint exists between numerous experiments that have demonstrated disease-suppressive effects and the translation of these successes to commercial products and applications. Clearly, the field of cereal disease protection leaves a lot to be explored and uncovered.
1. Wheat and Barley: Duo Cereals at the Foundation of Global Food Stability
Wheat and barley are among the main cereals cultivated worldwide with an estimated production of 765 million tons and 140 million tons, respectively, in 2020 [1]. The production of these cereals is crucial in view of their global use as both food and animal feed. Thus, both crops are the staple food for a third of the world’s population. The world’s cultivated area remains generally stable, with increasing yields mainly due to genetic selection [2,3].
Wheat is a monocotyledonous crop belonging to the kingdom Plantae, the class Liliopsida, the family Poaceae and the genus Triticum. The genus Triticum contains approximately 300 species occurring worldwide. Their genome plasticity has allowed them to occur across the globe with more than 25,000 different wheat varieties adapted to a large number of environments [4,5]. Triticum has two major species, namely common wheat, Triticum aestivum (2n = 42; AABBDD), and durum wheat, Triticum durum (2n = 28; AABB). Common wheat is mainly grown in northern and eastern Europe, while durum wheat is more suitable for warm regions. Moreover, based on their composition, they are not used for the same purposes. Common wheat, with a high content of gluten and protein, is used to make flour for bread and biscuit products. In contrast, durum wheat is richer in protein and amino acids with harder albumen, which can be transformed into semolina or pasta [6,7].
Barley, also a monocot, belongs to the same class and family as wheat but is affiliated to the genus Hordeum. This genus is categorized into three large species groups of which the most commonly cultivated belongs to the species Hordeum vulgare (2n = 14) [8]. Based on the fertility of their spikes, barleys can be classified into two groups comprising 2-row barleys and 6-row barleys [9,10]. Another method of classification is based on crop conditions namely spring barley, winter barley and escourgeon, which includes six-row cyclic-spiked varieties [11]. Barley is mainly used for animal feed but is also processed for the manufacture of alcohol (beer and whisky) or as food, particularly in several regions of the world such as North Africa, the Middle East and Asia, where average and annual consumption varies between 2 and 36 kg per person [8].
Like any cereal, the development cycle of these plants is influenced by seasonal and interannual climate variations and can be divided into three main stages: (i) the vegetative phase including germination with root, leaves and tillers growth until the stem elongation phase; (ii) the reproductive phase corresponding to the period between the tiller and the fertilization characterized in particular by the development of flowers and ears; and (iii) the maturation phase of the grains commencing with the fertilization up to the filling and maturation of the grains [12,13,14].
Since 1970, France has been the leading European producer and exporter of wheat ahead of Germany with five million hectares. The main production basins are Bassin Parisien, Nord-Pas-de-Calais, Centre, Poitou-Charentes and Bourgogne [1]. Common wheat accounts for about 54% of the agricultural area dedicated to cereals compared to only 4% for durum wheat. In addition, France ranks second in Europe for barley production, covering 1.77 million hectares for 11 million tons produced. In France, 1.7 million tons of malting barley were transformed into 1.4 million tons of malt in 2019, allowing the country to become the biggest producer in Europe. The main production areas are the Centre, Bourgogne and Champagne-Ardenne regions [1].
2. Main Diseases Affecting Barley and Wheat
During their development cycle, cereals such as wheat and barley are subject to different biotic and abiotic stresses. Among them are fungal diseases that can be extremely deleterious to the plant by attacking different organs including the roots, leaves and ears. The main fungal pathogens can be characterized according to their trophic biology as being biotrophic, necrotrophic and hemibiotrophic [15]. Biotrophic agents establish a long-term association with the host within living plant cells. They are highly specialized and absorb the nutrients present in plant cells without inducing cell death. In wheat and barley, biotrophic diseases are yellow and brown rust (caused by Puccinia striiformis f. sp. tritici and Puccinia recondita f. sp. tritici, respectively), powdery mildew (caused by Blumeria graminis f. sp. tritici.) and covered smut and loose smut (caused by Ustilago hordei and Ustilago nuda respectively). On the other hand, the necrotrophic agents develop by inducing cell death in tissues allowing them to proliferate for example, the pathogen eyespot (caused by Tapesia yallundae and Tapesia acuformis). Finally, hemibiotrophs are pathogens having an initial biotrophic cycle followed by a necrotrophic phase. Rhynchosporium (caused by Rhynchosporium secalis) and ramularia (caused by Ramularia collo-cygni) are specifically barley-associated pathogens. Ultimately, Fusarium head blight, Septoria tritici blotch and Net blotch can be considered to be the most harmful diseases on the cultivation of barley and wheat and are also caused by hemibiotrophic fungi [16,17].
2.1. Fusarium Diseases
Fusarium is an important disease that affects all plant organs and causes 30–70% yield losses depending on the severity of the attacks [18]. Besides wheat, Fusarium also affects other plants such as oats (Avena sativa L.), barley (Hordeum vulgare L.), rice (Zizania palustris L.), corn (Zea mays L.), and sorghum (Sorghum bicolor L.). Infection by the Fusarium pathogen may give rise to three distinct types of symptoms: Fusarium seedling blight, resulting in germination losses, Fusarium foot rot causing necrosis of crown tissues, and Fusarium head blight (FHB) attacking the ears [19,20]. The FHB is regarded as the main disease of cereals in Europe because of its significant presence worldwide and represents the fourth major global phytopathogenic fungus [21,22]. The disease is characterized by wilting of the ears and premature senescence. If the infection is early, the kernels are hollow and small, wrinkled and have a whitish color. If the contamination is later, the grains may have continued to fill but are pink with high concentrations of mycotoxins.
Fusarium diseases are associated with two genera of phytopathogenic fungi (Fusarium and Microdochium) that contain 19 species [23]. Under unfavorable conditions, as in winter, fungi survive as chlamydospores or mycelium in the soil. The fungus may then be present in the form of ascospores (anamorph form), which are mainly primary inoculum, and in the form of macroconidia (the teleomorph phase). Ascospores are released and disseminated by wind while the conidiospores present are spread by the splashing effect of water at the upper stages up to the ear. If the conditions are favorable at the level of the ear, meaning that the humidity is near 100% and the temperature is around 20 °C for a period of 48 to 72 h, the spores can germinate and initiate infection. In addition to environmental conditions, the stage of plant development is a key factor. The period of anthesis corresponds to the critical stage of infection due to the high level of expression of stimulants such as choline and betaine which stimulate the growth of F. graminearum and contribute to the infection of the ears by the pathogen [19,24,25,26].
Except for the genus Microdochium, the majority of species involved in the incidence of Fusarium head blight are capable of producing a wide range of mycotoxins namely Nivalenol (VIN), Deoxynivalenol (DON), Zearalenone (ZEA), Fumonisines (WUF) [27]. These mycotoxins present a major economic and health dilemma for the cereal industry as numerous studies have demonstrated the capacity of these fusariotoxins to inhibit protein synthesis in eukaryotic cells thereby altering the liver and nervous system of humans and animals [27]. Zearalenone causes hormonal disruption due to its structure, which is similar to estrogen [28]. In addition, DON is a virulence factor in wheat and barley that plays a crucial role in the development of the fungus in the ear. Naturally, a non-mycotoxin-producing Fusarium isolate is incapable of infecting other spikelets [29]. It has also been observed that at high concentrations, DON elicits different defense mechanisms including the synthesis of hydrogen peroxide (H2O2) whose main function is to induce cell death [30].
2.2. Septoria tritici Blotch
Septoria tritici blotch (STB) is one the most devasting disease of wheat and barley worldwide. Septoria tritici blotch, caused by the fungus ascomycete Mycosphaerella graminicola (teleomorph—sexual form) and Zymoseptoria tritici (anamorph—asexual form) is the most widespread disease in Europe, affecting crops particularly in humid climates such as north of France, United Kingdom or Germany. The impact on yield can cause up to 40% of yield loss and depends on both the intensity of the infection and the stage of plant growth at the time of contamination, as seed filling is primarily dependent on photosynthesis by the leaves [31]. The initial symptoms are small chlorotic spots on the leaves. Then, in the mature leaves, lesions develop and are mostly long, narrow, delimited by the leaf veins and contain black or brown fruiting structure named pycnidia. Zymoseptoria tritici, belongs to the class Dothideomycetes, of order Botryosphaeriales and family Sphaerioidaceae [32]. The fungus is highly dependent on environmental conditions but possess a high genetic diversity that gives them a high adaptive capacity [33]. The infection of Z. tritici usually begins during the fall period as soon as the plant is emerged. Sexually propagated ascospores are considered the primary form of primary inoculum. These spores are produced in the crop residues of the previous crop and contaminate the next crop of the following year. Secondary infection is caused by pycnidiospores carried over a shorter distance by splashing (vertical progression) [34]. Contamination begins with spores adhering to leaves that secrete an extracellular matrix to remain attached to the leaf surface until germination. When environmental conditions become favorable with 85% humidity and temperatures around 20 °C, spore germination commences as early as 2 h post-contamination. Infection begins with the penetration of germ tubes into plant tissues and occurs approximately 24–48 h after inoculation. Zymoseptoria tritici is a hemibiotrophic pathogen, which develops slowly in the apoplast without penetrating directly into the host cells and without forming specialized structures such as haustorium. Consequently, during this period of about 9 to 16 days, Z. tritici does not induce visible symptoms and seems to survive by assimilating nutrients in solution. The transition from the biotrophic phase to the necrotrophic phase results in the appearance of chlorotic lesions developing into necrotic spots [34,35,36,37].
Wheat can also be affected by the Septoria glume blotch (SGB) which attacks leaves and spikes and is caused by Phaeosphaeria nodorum (Muller) (teleomorph) and Stagonospora nodorum (Berk.) (anamorph). This pathogen is less prevalent in France but can also have important economic impacts with losses up to 50% and a lower grain quality [38]. The disease can affect all plant parts and results in small dark-brown lesions in the leaves and grayish-white center with a dark-brown periphery.
2.3. Net Blotch
Net blotch is the most damaging disease for both winter and spring barley and is present worldwide, especially in humid area. This disease can infect leaves, stems, and kernels. Leaf necrosis causes significant economic losses of up to nearly 50% of final yield loss and reduces seed quality in the absence of control over susceptible varieties [39]. Symptoms on leaves are brown necrotic spots increasing in size to form elliptical or fusiform lesions with sometimes chlorotic lesions [40].
The net blotch disease is caused by the ascomycete fungus Pyrenophora teres Drechsler, anamorph Drechslera teres (Sacc.) Shoemaker, which exists in two forms: Pyrenophora teres f. teres (Ptt) and P. teres f. maculata (Ptm). Pyrenophora teres belongs to the class of Dothideomycetes and order Pleosporales. Morphologically, Ptt and Ptm are indistinguishable, so their identification is based on the visible symptoms of the plant. The disease caused by Ptt has elongated light brown lesions with dark brown necrotic reticulations while Ptm causes ovoid black lesions that are surrounded by a distinct chlorotic zone [41]. Pyrenophora teres f. maculata may negatively impact the quality of the grain thereby reducing its ability to be used in malting [42,43].
Pyrenophora teres Drechs. is a hemibiotrophic fungus that is mainly in the necrotrophic form rather than biotrophic [44]. The fungus survives as ascospores in seed or residue from previous cultivation. The pseudothecia are formed after harvest and the ascospores during the winter period until spring according to the climatic conditions. Mature ascospores are dispersed by wind and serve as primary inoculum. Similarly, conidia in previously infected soils can be a source of primary contamination. Infection is most effective if temperatures are around 10–15 °C. After the first infection, the fungus produces a large number of conidia used as secondary inoculum. Sporulation occurs when the humidity reaches 100% and the temperature is between 15 and 25 °C for 10 to 30 h [41,45]. Once implanted, the fungus produces hyphae of greater or lesser length before forming a structure called the appressorium allowing the penetration of the pathogen. A study by Ronen et al. (2019) [39] demonstrated that Ptm is 70% more virulent and showed 20% more necrosis compared to Ptt. In addition, the study also confirmed that, depending on location and environmental conditions, one of the two forms of P. teres predominates, with Ptm more common in warm areas (from July to September). This finding would also explain the proliferation of Ptm in new regions whose average temperatures are increasing due to climate change [42,46].
Symptoms caused by P. teres are mainly caused by several toxins. Four toxins were successively identified in both Ptt and Ptm namely N-(2-amino-2-carboxyethyl)-aspartic acid, anhydroaspergillomarasmine A, aspergillomarasmine A and aspergillomarasmine B. These toxins belong to the class of marasmins whose main function is chelation of iron ions. They are very sensitive to abiotic stress including light and temperature. In addition to these toxins, four metabolites called pyrenoids A, B, C and D were extracted from P. teres without showing any phytotoxicity unlike the isoquinoline, pyrenoline A and B toxins, which are known to be toxic to monocotyledons. These toxins are involved in the development of necrosis (pyrenoid A) and chlorosis (pyrenoids B and C) [44,47,48].
3. Current Control Strategies against Pathogens of Barley and Wheat
Besides this strong pathogen pressure, farmers and the various participants in the agricultural sector must also face the emergence of new challenges and expectations from consumers, resulting in changes in consumption patterns and regulations. In order to meet the ever-increasing demand, the stakeholders need to create different control strategies that are mainly based on: chemical, biological, agronomic and genetic factors (Figure 1).
3.1. Chemical Control, Prophylactic Strategies and Genetics Selection
Chemical control is the most widespread method worldwide because of its effectiveness and its almost systematic use for several generations. In 2018, more than five million tons of pesticides were used globally to control diseases and pests [1]. At the national level, France is the 3rd largest consumer of pesticides in the world and the 1st in Europe [1]. A wide variety of molecules are already used in marketed products, but their efficiencies depend on the target pathogen. For example, the pathogens Fusarium and Microdochium do not have the same sensitivity to fungicides. Fusarium graminearum is particularly sensitive to triazoles while F. avenaceum is more sensitive to strobilurins. Tetraconazole has interesting effects on F. graminearum reduction and DON production [49]. Two triazoles, prothioconazole and tebuconazole, show significant efficacy on Fusarium with the advantage that prothioconazole is effective on both Fusarium and Microdochium. The addition of strobilurine, can disrupt the competition between fungi within the ear and cause a reduction of DON. Against Septoria tritici blotch, there are many fungicides such as quinone outside inhibitors (QoI or strobilurins), sterol biosynthesis inhibitors and succinate dehydrogenase inhibitors (SDHI) [50]. The best efficacy is achieved with the combination of a triazole, such as prothioconazole, with one or two active ingredients of the SDHI family such as bixafen in the last leaf stage. However, due to the plasticity of their genome, increasing numbers of species have developed resistance, which can potentially limit the effectiveness of plant protection products. For Net blotch, a study of the in vitro action of several fungicides on P. teres shows that the Ptt form is more sensitive than the Ptm form. The strobilurins have been largely used in recent years and contact fungicides are most effective in limiting conidia germination, whereas systemic fungicides show better efficacy on mycelial growth. On the other hand, seed treatments are not very effective on pathogens, particularly in the case of Ptm [51].
Although great progress has been made and limitations were created, the use of a so-called “conventional” agricultural system remains the most immediate and easiest way to prevent disease development. However, intensive use of plant protection products generates several risks: pathogen resistance development, groundwater and soil contamination, wildlife toxicity, trace elements in consumer products, etc. With the awareness of these issues, several international initiatives with agreements and regulatory frameworks to control the use of plant protection products and fertilizers have been established. Among these alternative solutions, the farmer can act on different levers such as direct control (physical struggle, use of biocontrol, trap plants) and indirect control (adaptation of cultural practices, use of resistant varieties, auxiliaries, and microbial ecology). All these control levers must consider different climatic, physiological and agronomic factors. The environmental conditions necessary for the survival and proliferation of pathogens can be partially controlled in the field by the farmer through different cropping practices [52,53]. At the soil preparation stage, tillage management allows the burial of residues and thus limits the risks of disease appearance [52,54,55]. Certainly, primary inoculum of pathogens such as F. graminearum, Z. tritici or P. teres are present in crop residues. The pathogens can survive for several years in the soil but can only develop in the first centimeters of the residue. To confirm this approach, Pfender and Wootke (1988), demonstrated that P. teres has a better chance of survival in upper mulch than in buried straw [54].
Secondly, crop rotation involves alternation of crops and lengthening of rotations to break the disease cycle. For instance, the risk of Fusarium Head Blight on wheat is reduced if the previous crop was not wheat, barley or corn but instead another family such as a Fabaceae (soybean) because the main Fusarium species of soybeans is F. sporotrichioides whereas in wheat it is F. graminearum [56]. In some cases, farmers also grow intermediate crops that can trap ascospores in residues and thus prevent them from being spread by wind to other fields [55,57]. A third prophylactic strategy is the management of seed sowing dates. If the date of sowing and thus of flowering coincides with the release of the spores, then the infection is more frequent and more severe. More so, early varieties are generally more resistant to diseases than other slow-growing varieties and are therefore more susceptible to pathogens over a longer time [57,58]. Furthermore, irrigation alters the microclimate of the plot by increasing moisture, resulting in favorable conditions for the pathogen. Depending on the type of irrigation (cannon, sprinkler, ramp), a massive supply of water over a short period increases the risk of leaching of contact chemical products. Besides, depending on the source of water, water may be a vector for some pathogens such as Fusarium. Lastly, the concentration of minerals in the soil can have a strong impact on pathogen development. This impact is very dependent on the stage of the plant and the balance between the different elements present in the soil. For example, excess nitrogen with a potassium deficiency causes the activation of enzymes such as amylase, protease and glucosidases in pathogens that promote their development [26]. In addition, the composition of fertilizers can have a profound effect on microbial communities. Organic fertilizers, produced naturally, promote soil processes, and improve soil microbial biomass. This higher microbial biomass supports microbial competition and prevents the proliferation of a small number of micro-organisms including pathogens [59]. However, other studies have shown that pathogens survive in organic fertilizers increasing the risk of contamination [60].
As a complement to the different farming practices, genetic control through using resistant varieties is a widely used method to effectively control wheat and barley diseases. A plant’s resistance is its ability to prevent the infection or growth of a pathogen in its tissues. This resistance is acquired through two main types of resistance: qualitative, with the «gene-for-gene» model of Flor (1971) [61] and quantitative constituting a polygenic resistance. Several genes of specific resistance have been well characterized to describe the wheat-Z. tritici interaction and have been called stb1 to stb18 [62]. This kind of resistance is very effective because it enables the activation of the plant’s defense mechanisms but is very often circumvented by the pathogen. Currently, TE 9111 is the most resistant strain in Europe because it has several qualitative resistance genes, namely Stb11, Stb6 and Stb7 [63,64]. In barley, several major effect QTLs were identified for resistance to P. teres on chromosomes 2H [65], 4H [66], 5H [67], 6H [66], and 7H [65]. Clare et al. (2019) [47] characterized a large number of resistance QTLs based on the pathogen Ptt or Ptm. Several genes have a particular effect on the Rpt5 locus which is considered as an important locus in the Ptt-barley interaction. Another gene involved in the P. teres-barley interaction is the HvS40 gene whose expression is induced by jasmonic acid and salicylic acid. Krupinska et al. (2002) [68] observed high expression levels only in leaf tissue with necrosis and chlorosis after infection suggesting that this gene has an important role during infection.
3.2. Biological Control
Biocontrol has long been considered as a less efficient, more expensive, and more burdensome method, and has remained mainly used for more specialized crops such as horticulture or market gardening. However, in recent years, with the increase of regulations, biological control including the use of plant growth promoting bacteria (PGPB) or their natural compounds have been extensively studied due to their potential benefits to reduce the use of chemical plant protection products. Subsequently, more companies embrace the potentiality of these products and are actively developing microbe-based product portfolios to suppress cereal diseases (Table 1).
Thus, combined with optimization of agronomic levers, the use of biocontrol is beginning to find its place in culture systems. The PGPB can directly benefit host plants by improving the absorption of plant nutrients and/or by modulating growth and phytohormones related to stress thereby conferring an evolutionary advantage to the plant [69]. In a field experiment, Pseudomonas chlororaphis MA32 was demonstrated to reduce the incidence of several wheat and barley pathogens via promotion of root and shoot growth. The two commercialized products based on this bacterium, Cedomon® and Cerall®, are currently largely used in ecological farms in Europe [70].
Indirectly, beneficial bacteria can improve plant health via competition for ecological niches, antibiotic production, lytic enzymes or volatile compounds, or induction of resistance mechanisms in the plant (Table 1) [69,71,72]. These modes of action are particularly studied for the research of new potential marketed bacteria. As demonstrated in Table 1, most of the existing products are composed of bacteria directly impacting the integrity of the pathogens. The InateqTM Active manufactured by Corteva is based on the properties of the Streptomyces sp. 517-02. This strain shows good crop safety and disease control on Z. tritici by inhibiting spore germination and also inhibiting mycelial growth post-germination [73]. The production of metabolites and the competition for space and nutrients are also a major part of the mechanisms for biocontrol products as it is the case with the Cerall product [70] or with the Streptomyces sp. K61 derivative Mycostop® [74]
4. The Context of Plant Defense
In any season, plants are confronted with permanent attacks by pathogens, possessing diverse life strategies. Some of these pathogens proliferate outside of the plant tissues, while others can directly penetrate plant cells. The apoplast is the place of the first interaction, between the plants and the pathogens, which is mediated by the recognition of microbial elicitors known as pathogen-associated molecular patterns (PAMPs) and that are identified in plants by membrane-localized pattern recognition receptors (PRRs). To disrupt cellular functions throughout the entire process of infection, pathogens secrete several effectors. In contrast to PAMPs, effectors are various and can involve proteins, chemicals, toxins, or hormones, which elevate pathogen infectiveness by either profiting the pathogen or by suppressing host defenses. Intracellular receptors, termed nucleotide-binding domain, leucine-rich repeat-containing proteins (NLRs, also identified as NB-LRRs), perceive specific effectors carried within the plant cell to activate effector-triggered immunity (ETI). The subsequent recognition of microbial derived PAMPs by PRRs of the plants, which are receptor-like kinases will trigger the first line of defense, called PAMP-triggered immunity (PTI). Among the prompt responses is an extracellular Ca2+ influx into the cytosol, which is then succeeded by the induction of cell oxidative burst with the production of the reactive oxygen species (ROS) and the activation of the mitogen-activated protein kinase, and additional signaling molecules, such as reactive nitrogen species, callose, n-hydroxypipecolic acid, salicylic acid, jasmonic acid, ethylene, and cytokinin (Figure 2) [50,75].
The synthesis of salicylic acid (SA) follows two different pathways, one involving the phenylalanine (Phe) ammonia-lyase (PAL) and the other, isochorismate synthase 1 (ICS1) [76]. Salicylic acid plays a crucial role in the long-distance signaling mechanism for systemic acquired resistance (SAR) induction, which leads to the localized programmed cell death and the activation of the pathogenesis-related (PR) genes especially the PR1, PR2 and PR5 [77,78]. The signaling of SAR involves lipid transfer protein DIR1 (Defective in Induced Resistance 1) but also different metabolites such as MeSA (Methyl Ester of SA), G3P (Glycerol-3-Phosphate), DA (DiterpenoiddehydroAbietinal), pipecolic acid (Pip) and azelaic acid (AzA) [79,80]. One of the key components for the mediation of the SAR pathway is the protein NPR1 which is a redox-mediated protein used as a transcriptional co-activator of PR genes. The NPR1 gene is a receptor for SA inducing a modification of the protein structure which is essential for the activation of the PR genes [81,82,83]. Furthermore, previous studies have highlighted the existence of two to six NPR1-like genes and especially two paralogs of NPR1, NPR3 and NPR4 that have a different affinity with SA [84]. They regulate the activity and the stability of NPR1 in different ways: at high SA concentration, NPR3 supposedly regulates the degradation of NPR1 during the effector-triggered immunity (ETI) phase resulting in localized programmed cell death whereas NPR4 is activated with lower SA concentration and causes the activation of PR gene expression. In non-stress conditions, NPR1 is present in quantity as a cytoplasmic oligomer. After an oxidative burst induced by the SA, NPR1 is monomerized and translocated to the nucleus via a bipartite nuclear localization signal (NLS) to indirectly switch on the PR gene expression via the activation of the TGA family of basic domain/leucine zipper (bZIP) transcription factors. In addition, some WRKY transcription factor genes have been identified to be SA-dependent and potentially involved in the regulation of the expression of NPR1 [84].
5. First Steps of Interaction between Plants and Beneficial Bacteria
5.1. Bacteria Perception
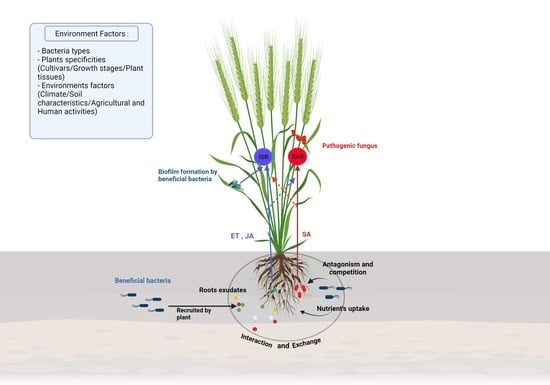
Plants are in constant interaction with other organisms like bacteria, which can interact directly with the surface of the plant and, in some cases, penetrate the tissues and colonize the interspatial region between plant cells. In order to be of benefit to the plant, it is necessary to set up a communication route in the bacteria-plant and bacteria-bacteria directions [71,85] (Figure 3).
For this purpose, specific communication is established between the plant and these bacteria via exudates secretion and the recognition of these compounds through a phenomenon named chemotaxis. These exudates are secreted by the plant root in the rhizosphere and their composition is dependent on plant specificity and environmental factors [86]. The root exudation is mainly passive through diffusion, ion channels or vesicle transport [69]. Plants secrete carbohydrates such as sugar, via an anion channel, metals through metal transporters, whereas water and uncharged molecules are secreted through aquaporins. Additionally, low molecular weight compounds and high molecular weight compounds, are secreted through diffusion and vesicles, respectively. When bacteria perceive the concentration gradient of root exudates, they become motile by means of flagellum or their pili in the specific direction [87,88].
Quorum sensing (QS) has also been studied for its role as a target for host recognition and its implication in communication and recognition during bacteria interactions. Bacteria can synthesize low molecular weight molecules which can be released extracellularly and recognized by autoinducers of other bacteria. Once the extracellular level of the autoinducers attains a critical level, the autoinducers bind to a cellular receptor and trigger a signal transduction cascade. The direct consequence is a change in bacterial gene expression facilitating interaction between cells and enabling the coordination of bacteria through synchronized gene expression [69,89,90]. The most studied signal molecule produced by Gram-negative bacteria is the acyl-homoserine lactone (AHL) which can bind the receptor LuxR-like protein to form a complex which in turn affects gene expression and activate transcription of QS-target genes [90,91,92,93]. This recognition is a key component in the communication between the beneficial bacteria. Indeed, N-AHLs mutants of Paraburkholderia phytofirmans PsJN could not colonize efficiently Arabidopsis thaliana and promote plant growth [94]. In Gram-positive bacteria, the autoinducers are generally peptides that interact with cognate regulators, phosphatases, or transcriptional regulators. In some PGPR, QS can regulate gene expression in the plant by the induction of plant systemic resistance and facilitation of plant growth. Indeed, some AHLs trigger the formation of adventitious roots due to hydrogen peroxide (H2O2) and nitric oxide (NO) dependent cyclic GMP signaling in mung bean. These two reactive oxygen are known for their effects of induce systemic acquired resistance [95,96].
5.2. Colonization and By-Passing of Plant Defense
Bacterial colonization depends on two distinct processes; bacterial adhesion and biofilm formation in the root or on the leaves [97]. The attachment of bacteria is an essential step for efficient colonization and is bacteria dependent. Obviously, the components of the bacterial cell surface play a significant role in the early stage of adhesion and colonization [98]. First, by means of its flagella, pili, or substances such as the exopolysaccharide (EPS) the bacteria cell moves to the beneficial position and overcome the energy barrier and bind to the plant surface. Thus, an important step in the colonization of endophytic bacteria is the formation of a biofilm that acts both as a boundary and a protective physical barrier. The biofilm formation process has been largely studied in the Gram-positive Bacillus subtilis. It is dependent on a large change in gene expression and particularly the transcriptional factor Spo0A, sigma-H and AbrB [99].
After establishing themselves in the interface with the plant, bacteria must move to find the best point of penetration via twitching which is under the control of two loci pilT and pilA essential for the formation and the retraction of the pilus [72,100,101]. Then, bacteria can migrate into the plant by employing different mechanisms that can be categorized into two strategies: passive and active colonization. In the rhizosphere, bacteria can passively colonize the plant interior through root wounds but also through primary and lateral root cracks [72]. They can also penetrate through natural openings on leaves and young stems, the cell wall junction like stomata, but also in hydathodes and stomatal pits or lenticels, which usually are present in the periderm of stems and germinating radicles [102]. Plant-associated bacteria such as Burkholeria sp., Azoarcus sp., Bacillus sp. or Streptomyces sp. are capable of secreting cell-wall degrading enzymes (CWDEs) that are active against the main plant cell wall components namely cellulose, hemicellulose and pectins [103,104,105]. These CWDEs allow bacteria to colonize the plant via an active form, or when they have colonized passively, continue to survive and proliferate into the host. Pectinase was demonstrated for example to be an essential enzyme for the Azoarcus sp. strain BH72 colonization inside rice roots [106]. Once they are situated in the host, bacterial colonization can be restricted to the specific tissue level or systematically throughout the plant. The movement of endophytes within the plant is supported by bacterial flagella, pili, and the plant transpiration stream. Furthermore, the perforated plates in the xylem vessel allow bacteria to pass through large pores without requiring specific CWDEs. Conversely, migration along intercellular spaces required the secretion of active CWDEs [100,107,108].
As they reside inside the plant, endophytic bacteria may have an advantage over rhizosphere-colonized bacteria. They are protected from environmental variations, can have access to a continuous supply of nutrients and have protection against competitive microorganisms [109]. Beneficial bacteria secrete and transport effectors proteins to the host via various kinds of transporter systems. Generally, the type I and II secretions systems are present in several beneficial endophytes. Two other types of secretion systems have been identified namely type V and VI. Type V is an autotransporter of endophytes whereas type VI (T6SSs) is a determinant in plant-microbe interactions [110]. These secretion systems allow bacteria to establish a prolonged and intimate mutualistic interaction with their host. Furthermore, to pass through the first layer of plant defense, bacteria secrete different molecules known as MAMPs which activate defensive reactions. However, some bacteria can protect themselves from these attacks by producing enzymes. In case of oxidative burst, endophytes can also produce dehydrogenases, synthases and hydratases such as superoxide dismutases (SOD), catalases (CatA), peroxidases (POD), or glutathione-S-transferases (GSTs) which prevent these beneficial bacteria from the ROS [111,112,113,114]. Another mechanism employed by bacteria to minimize the stimulation of the host’s immune system is the production of lower levels of cell-wall degrading enzymes. This pathway can act as a signal to specify to the plant that the bacteria present are beneficial [100]. Endophytic bacteria can induce phenotypic variation strategy as an adaptive process in which bacteria exhibit frequent and often reversible genetic modifications resulting in a reversible switch between colonies with different morphology [115]. Some bacteria have also co-evolved with the plant resulting in an abolishment of immune activation of the plant [116]. Thus, P. putida is supposed to be able to secrete AprA, which degrades flagellin monomers preventing immune recognition of flagellin in plant. This mechanism was revealed in the colonization process of this bacterium in barley roots [116]. Finally, similar to pathogens, some beneficial endophytes can modulate the expression and the concentration of the main phytohormones to promote host invasion like the PIIN_08944 effector of bacterium P. indica, which suppresses the expression of flg22-induced SA in Arabidopsis thaliana [117,118,119].
6. Beneficial Bacteria Implicated in Plant Defense
Beneficial bacteria present at the interface with the plant improve plant health through several mechanisms which may be direct or indirect [114,120]. In a direct approach, bacteria can promote plant growth by facilitating nutrients uptake and by reducing the effects of pathogens through competition for space and nutrients or by enzyme secretions to mediate antibiosis. On the other hand, the Induced Systemic Resistance (ISR), is an indirect and vital mechanism that explains a part of the disease suppression. Induced systemic resistance enhances the plant defense against many pathogens without directly activating important defenses while limiting the use of chemical fungicides [82,114]. These mechanisms will be explained in the context of wheat and barley defense against their pathogens (Table 2 and Table 3).
6.1. Induction of Plant Defense Mechanisms
6.1.1. SAR and ISR Pathways
Numerous studies have demonstrated the capacity of PGPRs to improve plant health by enhancing defense against a broad range of pathogens. When PGPRs encounter the pathogen, bacteria can induce a systemic resistance known as systemic acquired resistance (SAR), which has been associated with an enhanced level of endogenous SA. Beneficial bacteria can also induce a plant reaction named ISR which is regulated by jasmonic acid/ethylene (JA/ET)-dependent signaling pathways [114,167]. The role of JA and ET in the regulation of the ISR pathway was extensively studied in Arabidopsis with Pseudomonas fluorescens, Pseudomonas protegens, Serratia marcescens and Paenibacillus polymyxa but also in with some agronomical crops such as tomato and cereals [114,167,168,169,170,171,172]. This mechanism is based on the activation of the potentiation of plant genes defense resulting in a faster or stronger response upon pathogen attack. During this event, a quick H2O2 accumulation and callose deposition have been observed suggesting that ISR activates the first steps of plant protection. Jasmonic acid is synthesized from α-linolenic acid (LnA) originating from chloroplast galactolipids. Jasmonic acid is under the control of a positive-feedback regulatory system which includes the SCFCOI1 complex and JAZ repressor proteins as key components in the signal transduction [76,81]. Ethylene synthesis begins with the transformation of methionine to s-adenosylmethionine (SAM). Then, it is converted into 1-aminocyclopropane-1-carboxylic acid (ACC) by ACC synthase (ACS) and 5′-methylthioadenosine (MTA) which will be recycled to L-methionine (the immediate precursor of SAM) allowing the stabilization of L-methionine levels during fairly high rates of ethylene production [69]. Finally, ET is synthesized from aminocyclopropane-1-carboxylate (ACC) via the catalysis of a second enzyme, ACC oxidase (ACO) [69,78]. Jasmonic acid is converted into the active form JA-lIe (JA-isoleucine) by JARI. Then, JA-Ile is recognized and degrades a domain of the JAZ (Jasmonate ZIM) protein, which is a transcriptional suppressor of genes dependent on JA, through the 26s proteasome. Thus, the JAZ degradation allows the activation of JA TF as ERF1 and MYC2 that induces JA-dependent gene expression such as vsp2 and pdf1.2 [114,167,173]. Furthermore, the role of MYC2 in the interaction between H. parasitica and Pseudomonas fluorescens WCS417r highlighted the important role of MYC2 as a regulator of ABA-dependent defenses which regulate a part of the primed defense, particularly the deposition of callose-rich papillae at the sites of attempted spore penetration. This shows that the activation of MYC2 modulates the expression of JA-responsive transcription factor genes, and thus indirectly affecting the expression of numerous JA-responsive genes [174,175,176,177]. Some studies have succeeded in showing a direct relationship between the reduction of the impact of certain pathogens and the stimulation of plant ISR by beneficial bacteria. For example, Paenibacillus sp. strain B2 was demonstrated to significantly reduce the symptoms caused by S. tritici by inducing an overexpression of different plant genes involved in defense and cell rescue [151]. In the same way, Petti et al. (2010) [178] carried out one of the first transcriptomic analyses on barley to understand the impact of the Pseudomonas fluorescens strain MKB158 to protect from Fusarium head blight. They discovered a regulation on the JA/SA and ET pathways and an induction of long-distance signaling during SAR.
The SAR and ISR pathways are two distinct modes of defense with one particularly efficient against biotrophic and hemibiotrophic pathogens and the other against necrotrophic pathogens respectively. Generally, the JA/ET and SAR pathways act antagonistically; at a high level, SAR often antagonizes the ISR reaction by inhibiting JA signaling. Furthermore, ET acts positively on the expression of the ERF branch of the JA pathway whereas it suppresses the MYC branch which results in a prioritization of the ISR reaction. However, because of the constant pressure on plants by different pathogen lifestyles, there exists a SA-JA/ET cross talk which can be a potent mechanism to prioritize one pathway depending on the pathogen. Mur et al. (2006) [179] observed that the concentration of applied JA and SA can directly affect the response of the plant. In Arabidopsis thaliana, at low concentrations, the three main phytohormones act synergically on the JA and SA-responsive genes PDF1.2 and PR-1, respectively, whereas, at high concentrations, SA suppresses the JA pathway. NPR1 is an essential component in the SA-JA/ET crosstalk, although its role appears to be different. In SAR signaling, NPR1 is connected to a function in the nucleus while in the JA/ET signaling, it is associated with cytosolic function. Some studies indicate that SAR and ISR do not compete for the same pool of NPR1 [114,167]. Nie et al. (2017) [180] demonstrated that B. cereus AR165 induces plant resistance against Pseudomonas syringae DC3000 in Arabidopsis thaliana. The ISR pathway is dependent on SA and NPR1 but not JA/ET. They also showed that AR156 is effective against B. cinerea but in this case, via the JA-ET pathway and NPR1. Thus, this study highlighted that NPR1 is necessary in both cases and the same bacteria can induce two types of reactions.
6.1.2. Priming Effect
Following the perception of stimulus, plants can improve their resistance capacity by using a long-term sensitization which could allow them to react faster and more strongly. This phenomenon called priming is an adaptative strategy that includes the SAR pathway, induced by necrotizing pathogens, but it also includes ISR, activated by beneficial bacteria [114]. Priming is defined as “an induced, triggered or activated state in which the plant is initially treated with the prime inducing agent that enhances its resistance against secondary stresses” [174]. This supposed that the enhanced capacity of plants defense is not linked with direct activation of defensive genes but a stronger activation of basal defense and an accumulation of dormant protein kinases, which would require a secondary post-translational modification [181]. Cantoro et al. (2020) demonstrated an induced strengthening of epidermal cell wall after inoculation of B. velezensis RC 218 in the wheat tissues surrounding the F. graminearum infection site. The same strain was also studied for its capacity to induce plant defense in susceptible wheat cultivars by preventing an increase of pathogen-related hormones caused by the infection of F. graminearum on wheat spikes [126]. These studies highlighted the protective effect of the bacterium to limit the proliferation of the pathogen with activation of basal defense. Hypotheses regarding priming mechanisms have been demonstrated. The process can be divided into three main phases: (i) stimulus perception or priming phase, (ii) secondary stimulus challenges or primed state after challenge, and (iii) trans-generational phase or inherited state from primed parents [182]. During the first step of the perception, the cytosolic calcium increases trigger ion fluxes across the membrane to lead membrane depolarization. Depolarization is followed by a production of reactive oxygen species (ROS) which is crucial for the induction of defense. Furthermore, structural modifications occur during priming with callose deposition and infiltration of phenolic compounds to create an impermeable barrier to pathogen penetration [183]. In Arabidopsis thaliana, the SAR activator benzo (1,2,3) thiadiazole-7-carbothioic acid S-methyl ester was correlated with an accumulation of mRNA transcripts and inactive proteins of mitogen-activated protein kinase 3 (MAPK3) and MAPK6 and not a direct activation of PR [184]. The MAPK proteins are three-tiered signaling kinase modules that are receptors implicated in cellular signal amplification. The increase of MAPK level is associated with an enhancement of PR1 and PAL1 defense genes [181]. Thus, in the dormant phase, these proteins are a part of the mechanism explaining the prolonged defense priming. Furthermore, priming is also driven by the recognition of the bacteria by a plant receptor. Among these receptors, FLS2 is a leucine-rich repeat receptor kinase (LRR-RK) of the conserved N-terminal amino acid epitope flg22 in the bacterial MAMP flagellin which interacts with its signaling partner, BRI1-ASSOCIATED RECEPTOR KINASE1 (BAK1). Their interaction induces phosphorylation of both but also an accumulation of ROS and activation of MAPK cascade composed of MEKK1, MKK4/5, and MAPK3/6 [185]. The authors demonstrated that after treatment with benzothiadiazole in Arabidopsis thaliana, the levels of FLS2 and BAK1 increased with an enhancement of plant response to flg22. This finding suggests that priming increases the biosynthesis and the secretion of PRRs to the plasma membrane allowing the plant to respond more specifically and more quickly to MAMPs and PAMPs even at low doses.
6.2. Direct Antagonism
6.2.1. Space and Nutrients Competition and Plant Health
While beneficial bacteria are situated in the rhizosphere, they can directly interact with other microorganisms and compete with plant pathogens for space and nutrients. Competition for nutrients and space is considered as a means to limit the development of pathogens via the reduction of the number of habitable sites and thus, the inhibition of the germination of fungal spores in the soil [136]. When the antagonist is present in sufficient quantity at the right place and at the right time, it can reduce the fungal pressure. Besides, space competition has been demonstrated in the soil but also at the plant level [186]. Beneficial bacteria can colonize the plant root through the formation of biofilms and can also penetrate directly into the plant, hence limiting the pathogen. For example, Bacillus cereus is supposed to protect wheat crops against Septoria tritici by excluding the pathogen from the stomata and substomatal chambers [121]. Concerning the competition for nutrients like carbon, nitrogen or iron, this method can be considered as a common way to limit the growth of other microorganisms even if demonstrating its actual effect at field scale is difficult. A previous study showed that Pseudomonas sp. AS 64.4 reduced the Fusarium head blight (FHB) disease incidence by metabolizing chlorine, which is an essential nutrient found in wheat flower tissues, because chlorine is used as a source of carbon for the pathogen [154]. When iron becomes limiting, bacteria can produce a range of iron-chelating compounds known as siderophores which have a strong affinity with iron [187,188]. This strategy to take up available iron is a competitive advantage for the development of beneficial bacteria as it reduces the availability of the element for pathogens. Thus, similar to Streptomyces spp. and Pseudomonas mediterranea can inhibit different wheat pathogens including F. oxysporum, F. moniliforme, P. oryzae, and M. phaseolina [189]. The inhibition of pathogen growth or the inhibition of metabolic activity were also reported in different crop systems with Kosakonia radicincitans against Penicillium expansum, Botrytis cinerea, Rhizopus sp., Alternaria sp. and Cladosporium cladosporioides or with Pseudomonas strains GRP3A against Colletotrichum dematium, Rhizoctonia solani and Sclerotium rolfsi [190,191].
Furthermore, beneficial bacteria may also protect the plant by affecting the plant growth and development, and by facilitating nutrients acquisition. Certainly, plants need necessary elements like iron, nitrogen, and phosphorous to grow properly and optimize their yield and, at the same time, resist biotic stresses. However, these elements are rarely available in the environment because of their presence in an unassimilable form [120]. Among the essential nutrients, iron is essential for the growth, metabolism, and survival of the plant although the free form is rare in the environment because of the oxidation converting much of soluble iron into insoluble ferric oxides and oxyhydroxide. Wheat and barley plants have different strategies to acquire iron: the first one is the acidification of the rhizosphere followed by reduction of Fe3+ ions by membrane-bound Fe(III)− chelate reductase and then the uptake of Fe2+ into the cell. The second involved the secretion of phytosiderophore to solubilize the bound iron which is then transported into the cell in membrane protein [192]. Microbial siderophores are mainly used by the plant for iron uptake from the soil considering the fact that the plant can produce siderophores and other molecules negatively charged that can bind iron, but their affinity is must lower than microbial siderophore. Siderophores chelate ferric iron to form ferric siderophore complexes that are transported into the bacteria cell where ferric iron is released from the siderophore and reduced to Fe2+ to be utilized for metabolic processes [193,194,195]. Several bacteria have already been identified for their biocontrol efficacity against wheat pathogens. An in vitro essay from Lounaci et al. to analyze wheat protection from F. graminearum suggested that the production of siderophore by the bacterium Paenibacillus polymyxa SGK2 was one of the main mechanisms involved in the fungal inhibition [149]. Likewise, an assay realized with siderophore Pseudomonas putida strain BK8661 mutants revealed that antibiotic and siderophores production have a suppressive effect on Z. tritici. Furthermore, nitrogen is the most important element because it is the main constituent of amino acids, proteins and nucleic acids, constituting DNA and RNA but it must be reduced to ammonia before it can be metabolized by plants [196]. Some strains like Bradyrhizobium japonicum can convert H2 from the atmosphere into H+ via hydrogenase and produce ATP that can be used to fix more nitrogen. However, this particularity is not commonly found in the natural microbiome. Even if the fixation of the nitrogen costs energy to the bacteria, some studies revealed a direct impact of these bacteria on plant growth, especially during plant abiotic stress [197]. Thus, even if the promotion of plants can be efficient under abiotic stress, they can also be beneficial to control the cereals pathogens by making plants more vigorous and more resistant.
6.2.2. Secretion of Metabolites
Enzymes
Another mode of action employed by beneficial bacteria to inhibit pathogen proliferation is the direct confrontation and the secretion of antifungal compounds. Mycoparasitism is one of the most important antagonism involving direct physical contact with bacterial enzymes produced against the fungus thereby causing digestion of the host cell [198,199]. However, even if this mechanism is usual for some fungus such as Trichoderma spp., bacteria from Bacillus spp. [22,121,131,134], Pseudomonas spp. [154,155], Streptomyces spp. [126,137,141,142], Burkholderia spp. [200,201], and Lactobacillus spp. [148], routinely secrete antifungal enzymes, secondary metabolites or cell wall degrading enzymes. Among these antifungal enzymes, bacteria secrete β-1,3 and β-1,4 glucanases, chitinases, lipases and proteases. Moreover, these enzymes degrade the main structural component of pathogenic fungal cell walls namely chitin, β-1,3-glucan and protein. The β-1,3 glucanases hydrolyze the O-glycosidic linkages of β-1,4 glucan chains by sequentially cleaving glucose residues from the non-reducing end and by cleaving β-linkages at random sites oligosaccharides [202]. The action of enzymes against F. graminearum was demonstrated by He et al. (2009) [150] with Paenibacillus polymyxa W1-14-3 and C1-8-b which produce cell-wall degrading enzymes to disrupt the integrity of the pathogen. Bacillus subtilis SG6 could also induce the cell wall degradation enzyme of F. graminearum by chitinase secretion [203]. The compound 2,4-diacetylphloroglucinol produced by P. fluorescens PFM2 may also be implicated in the natural antagonism between the beneficial strain and Z. tritici. In addition to their effect on the integrity of the cell wall, bacteria can also affect the virulence of the pathogen as it was the case with Bacillus sp. 240B1 which is capable of enzymatic inactivation of autoinducers Ais (N-acylhomoserine lactones) by releasing AiiA enzyme. The AiiA enzyme is a lactonase that inactivates acyl-homoserine-lactone molecules which regulate the expression of virulence genes of Pseudomonas solanacearum [204,205].
Volatiles Compounds
Furthermore, beneficial bacteria can produce a wide variety of antibiotics and secondary metabolites which are differentially bioactive. Among metabolites, volatile compounds (VOCs) can inhibit the development or the germination of pathogens or activate plant defense mechanisms via induction of ISR. They are small odorous compounds (<C15) with low molecular mass (<300 Da), high vapor pressure, low boiling point, and a lipophilic moiety [206]. They are divided into different chemical classes including alcohols, alkenes, benzenoids, ketones, pyrazines, sulfides, and terpenes. The VOC production is dependent on the environmental conditions (temperature, oxygen availability, pH among others) and the growth stage of the bacteria and follows the GacS/GacA two-component regulatory system for many bacteria. The VOCs are considered to be a major key regulator system in the induction of ISR and induce the priming effect but also affect the plant’s secondary metabolite production and influence plant development via the modification of the hormone pathway. Thus, the VOC produced by B. subtilis GB03, named 2,3-butanediol, is known to be one of the major VOCs produced by Bacillus, and plays a critical role in the induction of systemic resistance against P. carotovorum. Bacillus secretes alcohols, such as 3-methyl-1-butanol, 2-methyl-1-butanol and butane-1-methoxy-3-methyl, which share a similar functional motif to that of 2,3-butanediol [207]. Li et al., (2015) [208] showed that emitted VOCs (mostly ketones and alcohols) obtained from eight Bacillus strains demonstrated 56–82% inhibition against the mycelial growth of Fusarium solani. The VOCs also inhibit pathogen growth by direct confrontation. Interestingly, a direct confrontation of Streptomyces salmonis PSRDC-09 with C. gloeosporioides PSU-03 led to the production of l-linalool that induced irregular distortions in the fungal hyphae [209]. A study of wheat protection against Z. tritici also demonstrated due to a dual plate culture assay the effect of VOC production by B. megaterium MKB135 on the pathogen germination [135]. Likewise, Streptomyces albulus NJZJSA2 produced 4-methoxystyrene, 2- pentylfuran, and anisole and successfully suppressed Sclerotinia sclerotiorum and Fusarium oxysporum [210].
Secondary Metabolites
Beneficial bacteria belonging to the Bacillus, Pseudomonas, Burkholderia, and Streptomyces genera produce a broad arsenal of secondary metabolites which are involved in direct antibiosis and induction of systemic resistance against plant-pathogens [211,212,213]. Pseudomonas strains produce several molecules involved in plant pathogen interactions including phenazines, pyoluteorins, pyrrolnitrins, 2,4-diacetylphloroglucinol and lipopeptides. On the other hand, Bacillus strains directly suppress plant pathogens by producing lipopeptides (iturin, surfactin and fengycin), siderophores, polyketides and oligopeptides among others [214].
Secondary metabolites of Pseudomonas have been implicated in the biocontrol of several plant pathogens [215,216]. Pseudomonas strains are interesting for application because of the enormous diversity of metabolites produced and for their good adaption to multiple environments. Towards the biocontrol of cereal pathogens, phenazine-1-carboxylic acid, produced by Pseudomonas fluorescens and P. aureofaciens suppressed Gaeumannomyces graminis var. tritici on wheat [217,218]. In another study, 2,4-diacetylphloroglucinol was shown to suppress the same pathogen on wheat [219,220]. Furthermore, pyocyanin derived from P. aeruginosa was effective against the wheat pathogen Septoria tritici [155,221,222]. In multiple studies, the pyrrolnitrin antibiotic from P. fluorescens effectively controlled Pyrenophora tritici-repens on wheat [223,224]. Several lipopeptide-producing pseudomonads have been isolated from the wheat rhizosphere [225] and show antagonism against soil-borne plant pathogens. However, to the best of our knowledge, none of these strains or their corresponding lipopeptides have been tested for efficacy against the main foliar pathogens of wheat and barley.
Bacillus strains are largely studied because of their capacities to suppress and inhibit plant pathogens, particularly through metabolites production. Bacillus strains can devote 4–5% of their genome to synthesize secondary metabolites [123]. The antimicrobial peptides are synthesized ribosomally or non-ribosomally by non-ribosomal peptide synthetases (NRPSs) and act directly in the suppression of pathogens. Bacteriocins are an example of ribosomally synthesized peptides including lantibiotics such as amylolysin, mersacidin, subtilin among several others [226]. Non-ribosomal peptides correspond to the unusual peptides dipeptides and oligopeptides, the cyclic lipopeptides (CLPs) and the polyketides (Pks). The dipeptides and oligopeptides include bacilysin, rhizocticin, chlorotetain, and bacitracin. Bacillaene, difficidin and macrolactin are the three main types of PKs [227]. The antifungal proprieties of the cyclo-lipopeptides (CLPs) have been widely demonstrated for many diseases. Thus, in contrast with Pseudomonas metabolites, the Bacillus LPs have been well studied against a wide range of cereal pathogens.
Lipopeptides (LPs) are described for their antagonism function, mobility and attachment to surface [228]. In general, LPs have been shown to have a strong protective effect against several pathogens on host plants. They are highly resistant to the environmental conditions and the attack by peptidases, proteinases, or oxidases. Their biosurfactant proprieties enable them to interfere with the cell wall membrane permeability in a dose-dependent manner which causes inhibition of the cell wall or the formation of spores to the pathogen. The LPs are known to penetrate the fungi cell membranes to form ion channels and cause the membrane osmotic imbalance causing cell death [212,228].
Although Bacillus produce several antibiotics according to strains, the main CLPs excreted by the bacterial strains can be classified into three main families, surfactins, iturins and fengycines, according to their amino acid sequence, the structure within the peptide ring, and the type and/or attachment of the fatty acyl chain [229]. They are synthesized by modular mega-enzymes called non-ribosomal peptide synthetases (NRPSs) [211]. The peptide part is hydrophilic while the lipid part is lipophilic thus resulting in amphiphilic molecules. This particularity has a direct consequence on the physico-chemical properties of these molecules and their biological activity as is the case for surfactins which are potent surfactants. Within each family, there are isoforms, differentiated by the nature of amino acids in the cycle, and homologs, which vary according to length and lipid chain isomeric [230]. The iturin family includes bacillomycin, iturin and mycosubtilin. They are cyclic lipoheptapeptides bound by a residue of ß-amino acid. It is known to disturb the cytoplasmic membrane causing leaching of K+ ions and other constituents involving cell death.
The annotation of the complete genome of Bacillus amyloliquefaciens AS 43.3 revealed nine gene clusters encoding secondary metabolites associated with the control of Fusarium graminearum [231]. Five of these clusters were identified as non-ribosomal peptide synthetases encoding three lipopeptides (surfactin, iturin, and fengycin), one siderophore (bacillibactin), and one antibiotic (bacilysin). Bacillus amyloliquefaciens S76-3 displayed a strong antifungal activity against Fusarium graminearum by producing iturin A and plipastatin A which caused leakage of fungi cellular contents and vacuolation [131]. The most active form of iturin, mycosubtilin, interacts with the phospholipid and sterols, especially with the acyl chains of the phospholipid and alcohol group of cholesterol in the membranes [123,131]. Fengycins are the other cyclic-lipopeptides family with antifungal activity like the fengycin produced by Bacillus subtilis strain IB which exhibit strong activity against F. graminearum due to the production of this metabolite [128,232]. Fengycin has a broad spectrum of action but is particularly effective against filamentous fungi. Like iturins, fengycins interact with the membrane by inserting the fatty acyl chain with the cyclic peptide region from the membrane leading to an increased thickness of the bilayer [229]. At high fengycin concentration, the cyclic peptide sections of the lipopeptides can increase the positive curvature of the membrane due to the shape of the compound leading to the formation of disk-like micelles of membrane phospholipids supported around the edges by the amphiphilic fengycins. When the concentrations are high, the pore formation in the membrane can lead to complete solubilization of the membrane into micelles as it was demonstrated by Deleu et al. (2005) [232]. Lastly, the surfactin family is composed of variants with heptapeptides interlinked with a β-hydroxy fatty acid to form a cyclic lactone ring structure [233]. Like fengycin, in membrane model, surfactin can insert the fatty acyl chain into the membrane but, in this specific case, the insertion is independent of the phase of the fatty acyl chain within the membrane and can be divided into two steps: peptide insertion into the membrane and electrostatic repulsion between charges borne by lipid headgroups and the negatively charged SF amino acid [234]. However, surfactin has no direct effect on fungi but seems to act synergically with iturin A activity [235]. The study of Cawoy et al. (2014) [236] revealed that surfactin and fengycin also induce the ISR mechanism in the tobacco plant. A strong correlation was found between the plant defense mechanism and the concentration of surfactin secreted by the bacteria. Cyclic lipopeptides were shown to be a non-volatile elicitor by stimulating the oxylipin pathway. Moreover, the coproduction of chitinases, fengycin and surfactins significantly reduced the development of Fusarium graminearum and DON production on wheat [203]. Surfactin is also involved in the attachment and aggregation of microcolonies in the formation of biofilm. This property is a key determinant in the control of some pathogens. The surfactin produced by B. subtilis 6051 participates in the formation of a stable biofilm on Arabidopsis thaliana roots. The deletion of the surfactin synthase gene in the strain decreases the rate of colonization of the bacteria and the biocontrol activity against P. syringae [237]. In this way, surfactin can be a good inhibitor of pathogen adhesion and biofilm formation to prevent them from infecting the host. Surfactin and iturin can also modulate the mobility of bacteria facilitating the colonization of the bacteria [238,239].
6.2.3. Limitation of Mycotoxins Production
The contamination of plants and thus food by mycotoxins is a significant cereal production problem. Mycotoxins are considered as major food safety issues due to their highly toxicity and their carcinogenic properties on animals and humans. Their regulation is traced by the International Agency for Research on Cancer and is subject to regulations with maximum residue levels. Aflatoxins, ochratoxin A and Fusarium toxins are the main mycotoxins that contaminate wheat and barley grains. Aflatoxins are mainly produced by Aspergillus flavus and Aspergillus parasiticus, while ochratoxins are a group produced by Aspergillus ochraceus, Penicillium verrucosum, and other Penicillium species. Both can contaminate processed products and animal-derived products which can lead to health problems that can be lethal in extreme cases [240,241].
Among Fusarium toxins, the zearalenone (ZEA) and deoxynivalenol (DON) produced by Fusarium species especially F. culmorum and F. graminearum are also considered as metabolites of particular concern to food and regulation agencies [150]. During the first step of the infection when the pathogen survives biotrophically, DON is assumed to be unimportant. However, at high concentration, DON in the plant cell promotes the synthesis of H2O2, resulting in the inhibition of defense-related responses and cell death promoting pathogen necrotrophic growth [30,242]. Currently, several methods are used to limit the risk of contamination of these mycotoxins but they are costly for a limiting efficacity [243]. Thereby, in recent years, the study of microorganisms and especially beneficial bacteria able to degrade mycotoxins to non or less toxic compounds appear promising [244]. In addition to the direct effects of the beneficial bacteria, their ability to reduce or to alter the production of mycotoxins by the pathogen can be an essential step in the reduction of the pathogen severity. Among these bacteria, several Rhodococcus, Pseudomonas or Bacillus species are instrumental in the degradation of ZEA. Altalhi et al. (2007) [245] demonstrated that Pseudomonas putida ZEA-1 was able to use ZEA as a carbon source by transforming the toxin into products with less or no toxicity. Moreover, B. licheniformis CK1 [246], B. subtilis [247], B. subtilis ANSB01G showed potential to degrade ZEA. In addition, the use of DON-degrading bacterium has been the subject of several studies [137,160,203,248,249,250]. Shima et al. [250] showed that the bacterial isolate E3-39, obtained from a mixed culture, had the capability to oxidize the 3-OH group of DON to generate 3-keto-4-deoxynivalenol (3-keto-DON) leading to less than one-tenth immunosuppressive toxicity relative to deoxynivalenol. Similarly, the bacteria Devosia mutants 17-2-E-8 were able to convert DON into a major and minor product, named 3-epi-DON and 3-keto-DON, respectively [249]. In a separate study, 13 isolates belonging to the genus Nocardioides and Devosia were obtained from field soils and wheat leaves and were all able to degrade 100 μg mL−1 DON [160,248].
7. Conclusions and Perspectives
In this review, we describe the main diseases affecting wheat and barley production and their causative pathogens. Furthermore, we discussed the current control measures being applied while highlighting the emerging potentials of biological control. By summarizing the general insights into molecular actors involved in plant-pathogen interactions, we show to an extent, how beneficial bacteria are implicated in plant defense against wheat and barley diseases.
For the control of wheat and barley diseases, previous research has focused predominantly on chemical methods, prophylactic strategies, and genetic selection. More recently, there has been a surge in the exploration of biological control methods albeit, with limited in-depth studies. Biological control is especially worthy of exploration in view of current trends to limit the use of environmental pollution due to pesticides use. Thus, we have summarized information on beneficial bacteria with demonstrated antifungal activity against pathogens of wheat and barley, including known modes of action while identifying potential areas for in-depth research. First, it appears that the origin of beneficial microbes could count towards their selective mode of action. Specifically, Pseudomonas strains derived from the wheat/barley phyllosphere or kernels/anthers suppressed target wheat and barley pathogens via production of hydrolytic enzymes or antibiotics such as DAPG, HCN, siderophores and phenazines (pyocyanin, phenazine-l-carboxylic acid and phenazine-1-carboxamide) [155,158,160]. Thus, disease suppression by these metabolites was efficient both through direct contact with the pathogen and via induced systemic resistance. To the best of our knowledge, most of the beneficial bacteria with efficacy against barley pathogens originated from the soil or maize/sorghum/crawberry rhizosphere. Exceptionally, seeds treatment with the crawberry rhizosphere isolate P. chlororaphis MA 342 (Cedomon, Cerall) has shown good disease control effects against P. teres in barley and S. nodorum in greenhouse experiments [157,163].
In contrast, numerous foliar- and soil-derived Bacillus species have been well studied in the control of wheat pathogens. This genus mediates direct antagonism or ISR via competition for space and nutrients or mainly through production of secondary metabolites which are either synthesized (non-) ribosomally. Such non-ribosomal peptides include mycosubtilin, surfactin, fengycin, iturin, plipastatin and bacillomycin among others [122,130,131,138]. It appears that for most of the Bacillus strains tested, the mode of action is suggested and not shown experimentally, which should be the focus for future studies [121,136,141]. In a very recent study, rhamnolipids (synthesized by species in Pseudomonas and Burkholderia genera), were shown to protect wheat against Z. tritici mainly through direct antifungal activity and without major impact on leaf physiology [251]. Such in-depth studies that explore the use of antimicrobial molecules accompanied by gene expression and metabolic profiling studies will expound our understanding of cereal-bacteria-metabolites-based research thereby contributing to increased barley and wheat production.
During the past decade, beneficial bacteria have emerged as promising alternatives compared to the use of chemical pesticides. However, the commercialization of strains efficient in greenhouses and fields is still a long way off for obvious reasons. Bacteria are very sensitive to their environment such as the temperature and humidity fluctuation, competition with other species, UV and the plant host, among others [252]. In this review, we have showed that the biocontrol commercial products available against cereal pathogens are not the main research topic of companies which focus especially on Trichoderma, Pseudomonas or Streptomyces. Noticeably, no Bacillus-based products are available. Clearly, a lot more needs to be done to translate in vitro and greenhouse-based research to practical solutions towards sustainable protection of wheat and barley pathogens. Factors to be considered include specific plant and pathogen lifestyle, the timing of biocontrol agent application, the persistence on the plant and host-specific responses to the biocontrol agent.
Author Contributions
Conceptualization, E.D., F.E.O. and Q.E.; methodology, E.D., F.E.O. and Q.E.; software, E.D., E.A.B.; validation, E.D., F.E.O., Q.E. and E.A.B.; formal analysis, E.D., F.E.O., Q.E. and E.A.B.; investigation, E.D., F.E.O., Q.E. and E.A.B.; resources; Writing—Original draft preparation, E.D. Writing—Review and editing, E.D., F.E.O., Q.E, C.C. and E.A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the University of Reims Champagne-Ardenne. We gratefully acknowledge the financial support provided by the Bio4Grain project, ADEME (Agence de la Transition Ecologique), France.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Food and Agriculture Organization of the United Nations FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/RP (accessed on 19 April 2022).
- Sharma, I.; Tyagi, B.; Singh, G. Enhancing Wheat Production—A Global Perspective. Indian J. Agric. Sci. 2015, 85, 3–13. [Google Scholar]
- Watson, A.; Ghosh, S.; Williams, M.J.; Cuddy, W.S.; Simmonds, J.; Rey, M.-D.; Hatta, M.A.M.; Hinchliffe, A.; Steed, A.; Reynolds, D.; et al. Speed breeding is a powerful tool to accelerate crop research and breeding. Nat. Plants 2018, 4, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siou, D. Développement épidémique de la Fusariose des épis de blé et Conséquences des Intéractions Entre Espèces du Complexe Fusarien. Ph.D. Thesis, Université Paris Sud, Paris, France, 2013. Available online: https://tel.archives-ouvertes.fr/tel-00849969 (accessed on 15 December 2021).
- Feldman, M. Origin of Cultivated Wheat. In The World Wheat Book: A History of Wheat Breeding; Bonjean, A.P., Angus, W.J., Eds.; Intercept Ltd.: London, UK, 2001; pp. 3–56. [Google Scholar]
- Matsuoka, Y. Evolution of Polyploid Triticum Wheats under Cultivation: The Role of Domestication, Natural Hybridization and Allopolyploid Speciation in their Diversification. Plant Cell Physiol. 2011, 52, 750–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shewry, P.R. Wheat. J. Exp. Bot. 2009, 60, 1537–1553. [Google Scholar] [CrossRef] [PubMed]
- El-Haramein, F.J.; Grando, S. Determination of iron and zinc content in food barley. In Proceedings of the 10th International Barley Genetics Symposium, Alexandria, Egypt, 5–10 April 2008. [Google Scholar]
- Baum, B.B. Classification of cultivated barley (Hordeum vulgare). 1. Historical aspects, and phenetic character analysis of some characters by information theory and by spatial autocorrelation. Can. J. Bot. 1986, 64, 2769–2773. [Google Scholar] [CrossRef]
- Wiggans, R.-G.; Meunissier, A. Classification des variétés d’Orges cultivées. J. D’agriculture Tradit. Bot. Appliquée 1992, 14, 568–571. [Google Scholar] [CrossRef]
- Harwood, W.A. An Introduction to Barley: The Crop and the Model. In Barley: Methods and Protocols; Harwood, W.A., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2019; pp. 1–5. ISBN 978-1-4939-8944-7. [Google Scholar]
- Soltner, D. Les grandes Productions Végétales: Céréales, Plantes Sarclées, Prairies. In Sciences et Techniques Agricoles; Université de Lorraine: Nancy, France, 1995; Volume 18, p. 471. ISBN 2-907710-02-8. [Google Scholar]
- Meier, U. Stades phénologiques des mono-et dicotylédones cultivées. In BBCH Monographie; Open Agrar Repositorium: Greifswald, Germany, 2018; Volume 18. [Google Scholar] [CrossRef]
- Bingham, I.J.; Hoad, S.P.; Spink, J.H.; Blake, J.; Foulkes, J. The Barley Growth Guide; Agriculture and Horticulture Development Board: Kenilworth, UK, 2006; pp. 3–27.
- Vleeshouwers, V.G.A.A.; Oliver, R.P. Effectors as Tools in Disease Resistance Breeding Against Biotrophic, Hemibiotrophic, and Necrotrophic Plant Pathogens. Mol. Plant Microbe Interact. 2014, 27, 196–206. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, M.; Hammond-Kosack, K.E.; Solomon, P.S. A review of wheat diseases-A field perspective. Mol. Plant Pathol. 2018, 19, 1523–1536. [Google Scholar] [CrossRef]
- BASF Maladies du Blé. Available online: https://www.agro.basf.fr/fr/cultures/ble/maladies_du_ble/ (accessed on 7 December 2021).
- Ludovic, B.; Giles, C.; Philippe, C.; Denis, G.; Lise, G.V.; Eric, M.; Jean-Yves, M.; Claude, M.; Pierre, T.; Romain, V.; et al. Céréales à Paille: Interventions de Printemps; Arvalis—Institut du vegetal: Paris, France, 2019; pp. 1–172. [Google Scholar]
- Karlsson, I.; Persson, P.; Friberg, H. Fusarium Head Blight from a Microbiome Perspective. Front. Microbiol. 2021, 12, 371. [Google Scholar] [CrossRef]
- Parry, D.W.; Jenkinson, P.; McLeod, L. Fusarium ear blight (scab) in small grain cereals? a review. Plant Pathol. 1995, 44, 207–238. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comby, M.; Gacoin, M.; Robineau, M.; Rabenoelina, F.; Ptas, S.; Dupont, J.; Profizi, C.; Baillieul, F. Screening of wheat endophytes as biological control agents against Fusarium head blight using two different in vitro tests. Microbiol. Res. 2017, 202, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Nicholson, P. Community Ecology of Fungal Pathogens Causing Wheat Head Blight. Annu. Rev. Phytopathol. 2009, 47, 83–103. [Google Scholar] [CrossRef] [PubMed]
- Bal, G. Scab of Wheat: Prospects For Control. Plant Dis. 1994, 78, 760–766. [Google Scholar] [CrossRef]
- Kazan, K.; Gardiner, D.M.; Manners, J.M. On the trail of a cereal killer: Recent advances in Fusarium graminearum pathogenomics and host resistance. Mol. Plant Pathol. 2012, 13, 399–413. [Google Scholar] [CrossRef]
- Champeil, A.; Doré, T.; Fourbet, J. Fusarium head blight: Epidemiological origin of the effects of cultural practices on head blight attacks and the production of mycotoxins by Fusarium in wheat grains. Plant Sci. 2004, 166, 1389–1415. [Google Scholar] [CrossRef]
- Trail, F. For Blighted Waves of Grain: Fusarium graminearum in the Postgenomics Era. Plant Physiol. 2009, 149, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, K.; Habrowska-Górczyńska, D.E.; Piastowska-Ciesielska, A.W. Zearalenone as an endocrine disruptor in humans. Environ. Toxicol. Pharmacol. 2016, 48, 141–149. [Google Scholar] [CrossRef]
- Miller, S.S.; Watson, E.M.; Lazebnik, J. Characterization of an Alien Source of Resistance to Fusarium Head Blight Transferred to Chinese Spring Wheat; Canadian Science Publishing: Ottawa, ON, Canada, 2011; pp. 301–311. [Google Scholar]
- Desmond, O.J.; Manners, J.M.; Stephens, A.E.; Maclean, D.J.; Schenk, P.M.; Gardiner, D.M.; Munn, A.L.; Kazan, K. The Fusarium mycotoxin deoxynivalenol elicits hydrogen peroxide production, programmed cell death and defence responses in wheat. Mol. Plant Pathol. 2008, 9, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Shaw, M.W.; Royle, D.J. Airborne inoculum as a major source of Septoria tritici (Mycosphaerella graminicola) infections in winter wheat crops in the UK. Plant Pathol. 1989, 38, 35–43. [Google Scholar] [CrossRef]
- Nasraoui, B. Les Champignons Parasites des Plantes Cultivées (Biologie, Systématique, Pathologie, Maladies); Centre de Publication Universitaire: Manouba, Tunisia, 2006; Volume 1, ISBN 978-9973-37-302-1. [Google Scholar]
- El Chartouni, L.; Tisserant, B.; Siah, A.; Duyme, F.; Leducq, J.-B.; Deweer, C.; Fichter-Roisin, C.; Sanssené, J.; Durand, R.; Halama, P.; et al. Genetic diversity and population structure in French populations of Mycosphaerella graminicola. Mycologia 2011, 103, 764–774. [Google Scholar] [CrossRef] [PubMed]
- Brokenshire, T. Wheat Debris as an Inoculum Source for Seedling Infection by Septoria tritici. Plant Pathol. 1975, 24, 202–207. [Google Scholar] [CrossRef]
- Suffert, F.; Sache, I.; Lannou, C. Early stages of septoria tritici blotch epidemics of winter wheat: Build-up, overseasoning, and release of primary inoculum. Plant Pathol. 2010, 60, 166–177. [Google Scholar] [CrossRef]
- Palma-Guerrero, J.; Ma, X.; Torriani, S.F.F.; Zala, M.; Francisco, C.S.; Hartmann, F.E.; Croll, D.; McDonald, B.A. Comparative Transcriptome Analyses in Zymoseptoria tritici Reveal Significant Differences in Gene Expression Among Strains during Plant Infection. Mol. Plant Microbe Interact. 2017, 30, 231–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morais, D. Les Déterminants des Phases épidémiques Précoces de la Septoriose du Blé (Zymoseptoria tritici): Quantité, Efficacité et Origine de L’inoculum Primaire. Ph.D. Thesis, AgroParisTech, Paris, France, 2015. Available online: hal.archives-ouvertes.fr/tel-01142864 (accessed on 15 December 2021).
- Downie, R.C.; Lin, M.; Corsi, B.; Ficke, A.; Lillemo, M.; Oliver, R.P.; Phan, H.T.T.; Tan, K.-C.; Cockram, J. Septoria Nodorum Blotch of Wheat: Disease Management and Resistance Breeding in the Face of Shifting Disease Dynamics and a Changing Environment. Phytopathology 2021, 111, 906–920. [Google Scholar] [CrossRef]
- Ronen, M.; Sela, H.; Fridman, E.; Perl-Treves, R.; Kopahnke, D.; Moreau, A.; Ben-David, R.; Harel, A. Characterization of the Barley Net Blotch Pathosystem at the Center of Origin of Host and Pathogen. Pathogens 2019, 8, 275. [Google Scholar] [CrossRef] [Green Version]
- Backes, A.; Guerriero, G.; Barka, E.A.; Jacquard, C. Pyrenophora teres: Taxonomy, Morphology, Interaction with Barley, and Mode of Control. Front. Plant Sci. 2021, 12, 614951. [Google Scholar] [CrossRef]
- Moolhuijzen, P.; Lawrence, J.A.; Ellwood, S.R. Potentiators of Disease during Barley Infection by Pyrenophora teres f. teres in a Susceptible Interaction. Mol. Plant Microbe Interact. 2021, 34, 779–792. [Google Scholar] [CrossRef]
- Rau, D.; Attene, G.; Brown, A.H.D.; Nanni, L.; Maier, F.J.; Balmas, V.; Saba, E.; Schäfer, W.; Papa, R. Phylogeny and evolution of mating-type genes from Pyrenophora teres, the causal agent of barley “net blotch” disease. Curr. Genet. 2007, 51, 377–392. [Google Scholar] [CrossRef]
- McLean, M.S.; Martin, A.; Gupta, S.; Sutherland, M.W.; Hollaway, G.J.; Platz, G.J. Validation of a new spot form of net blotch differential set and evidence for hybridisation between the spot and net forms of net blotch in Australia. Australas. Plant Pathol. 2014, 43, 223–233. [Google Scholar] [CrossRef]
- Sarpeleh, A.; Wallwork, H.; Catcheside, D.E.A.; Tate, M.E.; Able, A.J. Proteinaceous Metabolites from Pyrenophora teres Contribute to Symptom Development of Barley Net Blotch. Phytopathology 2007, 97, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Jalli, M. The Virulence of Finnish Pyrenophora Teres f.Teres Isolates and Its Implications for Resistance Breeding. Ph.D. Thesis, Faculty of Agriculture and Forestry of the University of Helsinki, Helsinki, Finland, 2010. Available online: http://www.mtt.fi/mtttiede/pdf/mtttiede9.pdf (accessed on 15 December 2021).
- Rau, D.; Maier, F.J.; Papa, R.; Brown, A.H.; Balmas, V.; Saba, E.; Schaefer, W.; Attene, G. Isolation and characterization of the mating-type locus of the barley pathogen Pyrenophora teres and frequencies of mating-type idiomorphs within and among fungal populations collected from barley landraces. Genome 2005, 48, 855–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clare, S.J.; Wyatt, N.A.; Brueggeman, R.S.; Friesen, T.L. Research advances in the Pyrenophora teres–barley interaction. Mol. Plant Pathol. 2019, 21, 272–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muria-Gonzalez, M.J.; Zulak, K.G.; Allegaert, E.; Oliver, R.P.; Ellwood, S.R. Profile of the in vitro secretome of the barley net blotch fungus, Pyrenophora teres f. teres. Physiol. Mol. Plant Pathol. 2019, 109, 101451. [Google Scholar] [CrossRef]
- Haidukowski, M.; Pascale, M.; Perrone, G.; Pancaldi, D.; Campagna, C.; Visconti, A. Effect of fungicides on the development ofFusarium head blight, yield and deoxynivalenol accumulation in wheat inoculated under field conditions with Fusarium graminearum and Fusarium culmorum. J. Sci. Food Agric. 2004, 85, 191–198. [Google Scholar] [CrossRef]
- Mejri, S. Efficacités et Modes D’action de Nouveaux Simulateurs de Défenses des Plantes sur le Pathosystème Blé—Zymoseptoria tritici; Sciences Agricoles; Université du Littoral Côte d’Opale: Dunkerque, France, 2018. [Google Scholar]
- Bendahmane, B.; Barrault, G.; Albertini, L.; Toubia-Rahme, H. Etude de l’action in Vitro de Divers Fongicides Sur Le Développement de Drechslera Teres f. Teres et f. Maculata. Phytopathol. Mediterr. 1992, 31, 77–84. [Google Scholar]
- Jørgensen, L.; Olsen, L. Control of tan spot (Drechslera tritici-repentis) using cultivar resistance, tillage methods and fungicides. Crop Prot. 2007, 26, 1606–1616. [Google Scholar] [CrossRef]
- Bayer les Méthodes Agronomiques Pour Limiter la Pression Maladies: Bayer-Agri, Conseils Phyto. Available online: https://www.bayer-agri.fr/cultures/les-methodes-agronomiques-pour-limiter-la-pression-maladies_1498/ (accessed on 5 December 2021).
- Pfender, W.F.; Wootke, S.L. Microbial communities ofPyrenophora-infested wheat straw as examined by multivariate analysis. Microb. Ecol. 1988, 15, 95–113. [Google Scholar] [CrossRef]
- Bankina, B.; Bimšteine, G.; Arhipova, I.; Kaņeps, J.; Stanka, T. Importance of Agronomic Practice on the Control of Wheat Leaf Diseases. Agriculture 2018, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Dill-Macky, R.; Jones, R.K. The Effect of Previous Crop Residues and Tillage on Fusarium Head Blight of Wheat. Plant Dis. 2000, 84, 71–76. [Google Scholar] [CrossRef] [Green Version]
- McDonald, B.A.; Mundt, C.C. How Knowledge of Pathogen Population Biology Informs Management of Septoria Tritici Blotch. Phytopathology 2016, 106, 948–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, E.M.; Boareto, C.; Danelli, A.L.D.; Zoldan, S.M. Anthesis, the infectious process and disease progress curves for fusarium head blight in wheat. Summa Phytopathol. 2016, 42, 134–139. [Google Scholar] [CrossRef]
- Liu, X.; Herbert, S.; Hashemi, A.; Zhang, X.; Ding, G. Effects of agricultural management on soil organic matter and carbon transformation—A review. Plant Soil Environ. 2011, 52, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Soonvald, L.; Loit, K.; Runno-Paurson, E.; Astover, A.; Tedersoo, L. The role of long-term mineral and organic fertilisation treatment in changing pathogen and symbiont community composition in soil. Appl. Soil Ecol. 2019, 141, 45–53. [Google Scholar] [CrossRef]
- Flor, H.H. Current Status of the Gene-For-Gene Concept. Annu. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Zhong, Z.; Marcel, T.C.; Hartmann, F.E.; Ma, X.; Plissonneau, C.; Zala, M.; Ducasse, A.; Confais, J.; Compain, J.; Lapalu, N.; et al. A small secreted protein in Zymoseptoria tritici is responsible for avirulence on wheat cultivars carrying the Stb6 resistance gene. New Phytol. 2017, 214, 619–631. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.K.; Chartrain, L.; Lasserre-Zuber, P.; Saintenac, C. Genetics of resistance to Zymoseptoria tritici and applications to wheat breeding. Fungal Genet. Biol. 2015, 79, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Chartrain, L.; Joaquim, P.; Berry, S.T.; Arraiano, L.S.; Azanza, F.; Brown, J.K.M. Genetics of resistance to septoria tritici blotch in the Portuguese wheat breeding line TE 9111. Theor. Appl. Genet. 2005, 110, 1138–1144. [Google Scholar] [CrossRef]
- Williams, K.J.; Lichon, A.; Gianquitto, P.; Kretschmer, J.M.; Karakousis, A.; Manning, S.; Langridge, P.; Wallwork, H. Identification and mapping of a gene conferring resistance to the spot form of net blotch (Pyrenophora teres f maculata) in barley. Theor. Appl. Genet. 1999, 99, 323–327. [Google Scholar] [CrossRef]
- Grewal, T.S.; Rossnagel, B.G.; Pozniak, C.J.; Scoles, G.J. Mapping quantitative trait loci associated with barley net blotch resistance. Theor. Appl. Genet. 2007, 116, 529–539. [Google Scholar] [CrossRef]
- Manninen, O.; Jalli, M.; Kalendar, R.; Schulman, A.; Afanasenko, O.; Robinson, J. Mapping of major spot-type and net-type net-blotch resistance genes in the Ethiopian barley line CI 9819. Genome 2006, 49, 1564–1571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krupinska, K.; Haussühl, K.; Schäfer, A.; van der Kooij, T.A.; Leckband, G.; Lorz, H.; Falk, J. A Novel Nucleus-Targeted Protein Is Expressed in Barley Leaves during Senescence and Pathogen Infection. Plant Physiol. 2002, 130, 1172–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glick, B.R. Beneficial Plant-Bacterial Interactions; Springer International Publishing: Cham, Switzerland, 2015; ISBN 978-3-319-13920-3. [Google Scholar]
- Agence Française de Sécurité Sanitaire des Aliments. Avis de l’Agence Française de Sécurité Sanitaire des Aliments Relatif à une Demande D’autorisation de Mise sur le Marché de la Préparation CERALL à Base de Pseudomonas chlororaphis souche MA 342. In Produite par la Société BELCHIM CROP PROTECTION NV/SA; AFSSA: Maisons-Alfort, France, 2008; pp. 1–9. [Google Scholar]
- Nascimento, F.X.; Rossi, M.J.; Glick, B.R.; Nascimento, F.X.; Rossi, M.J.; Glick, B.R. Ethylene and 1-Aminocyclopropane-1-carboxylate (ACC) in Plant–Bacterial Interactions. Front. Plant Sci. 2018, 9, 114. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, G.; Moreno-Hagelsieb, G.; del Carmen Orozco-Mosqueda, M.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
- Corteva Agriscience Inatreq Active. Technical Bulletin. 2018, pp. 4–15. Available online: https://www.corteva.com/content/dam/dpagco/corteva/global/corporate/general/files/active-ingredients/INA.045.Corteva.Inatreq.Global.Technical.Bulletin.cereals.pdf (accessed on 5 December 2021).
- Bubici, G.N. Streptomyces spp. as biocontrol agents against Fusarium species. CAB Rev. Perspect. Agric. Veter. Sci. Nutr. Nat. Resour. 2018, 13, 50. [Google Scholar] [CrossRef]
- Jemmali, L. Stimulateurs des Défenses Naturelles du blé dur en Tunisie et du blé Tendre en France Contre la Septoriose Causée par Zymoseptoria tritici. Ph.D. Thesis, Université du Littoral Côte d’Opale, Dunkerque, France, 2015. Available online: https://tel.archives-ouvertes.fr/tel-01626008 (accessed on 3 December 2021).
- Knogge, W.; Lee, J.; Rosahl, S.; Scheel, D. Signal Perception and Transduction in Plants. In The Mycota; Deising, H.B., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 5, pp. 337–361. ISBN 978-3-540-87406-5. [Google Scholar]
- Villena, J.; Kitazawa, H.; Van Wees, S.; Pieterse, C.M.J.; Takahashi, H. Receptors and Signaling Pathways for Recognition of Bacteria in Livestock and Crops: Prospects for Beneficial Microbes in Healthy Growth Strategies. Front. Immunol. 2018, 9, 2223. [Google Scholar] [CrossRef]
- Tyagi, S.; Mulla, S.I.; Lee, K.-J.; Chae, J.-C.; Shukla, P. VOCs-mediated hormonal signaling and crosstalk with plant growth promoting microbes. Crit. Rev. Biotechnol. 2018, 38, 1277–1296. [Google Scholar] [CrossRef]
- Van Oosten, V.R.; Bodenhausen, N.; Reymond, P.; Van Pelt, J.A.; Van Loon, L.C.; Dicke, M.; Pieterse, C.M.J. Differential Effectiveness of Microbially Induced Resistance Against Herbivorous Insects in Arabidopsis. Mol. Plant Microbe Interact. 2008, 21, 919–930. [Google Scholar] [CrossRef] [Green Version]
- Champigny, M.J.; Shearer, H.; Mohammad, A.; Haines, K.; Neumann, M.; Thilmony, R.; He, S.Y.; Fobert, P.; Dengler, N.; Cameron, R.K. Localization of DIR1 at the tissue, cellular and subcellular levels during Systemic Acquired Resistance in Arabidopsisusing DIR1:GUS and DIR1:EGFP reporters. BMC Plant Biol. 2011, 11, 125. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal Modulation of Plant Immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [Green Version]
- Bakker, P.A.; Doornbos, R.F.; Zamioudis, C.; Berendsen, R.; Pieterse, C.M. Induced Systemic Resistance and the Rhizosphere Microbiome. Plant Pathol. J. 2013, 29, 136–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankiewicz, U.; Kołtonowicz, M. The involvement of Pseudomonas bacteria in induced systemic resistance in plants (Review). Appl. Biochem. Microbiol. 2012, 48, 244–249. [Google Scholar] [CrossRef]
- Backer, R.; Naidoo, S.; van den Berg, N. The nonexpressor of pathogenesis-related genes 1 (NPR1) and related family: Mechanistic insights in plant disease resistance. Front. Plant Sci. 2019, 10, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orozco-Mosqueda, M.D.C.; Santoyo, G. Plant-microbial endophytes interactions: Scrutinizing their beneficial mechanisms from genomic explorations. Curr. Plant Biol. 2020, 25, 100189. [Google Scholar] [CrossRef]
- Bren, A.; Eisenbach, M. How Signals Are Heard during Bacterial Chemotaxis: Protein-Protein Interactions in Sensory Signal Propagation. J. Bacteriol. 2000, 182, 6865–6873. [Google Scholar] [CrossRef] [Green Version]
- Porter, S.L.; Wadhams, G.H.; Armitage, J.P. Signal processing in complex chemotaxis pathways. Nat. Rev. Microbiol. 2011, 9, 153–165. [Google Scholar] [CrossRef]
- Mark, G.L.; Dow, J.M.; Kiely, P.D.; Higgins, H.; Haynes, J.; Baysse, C.; Abbas, A.; Foley, T.; Franks, A.; Morrissey, J.; et al. Transcriptome profiling of bacterial responses to root exudates identifies genes involved in microbe-plant interactions. Proc. Natl. Acad. Sci. USA 2005, 102, 17454–17459. [Google Scholar] [CrossRef] [Green Version]
- Elasri, M.; Delorme, S.; Lemanceau, P.; Stewart, G.; Laue, B.; Glickmann, E.; Oger, P.M.; Dessaux, Y. Acyl-Homoserine Lactone Production Is More Common among Plant-Associated Pseudomonas spp. than among Soilborne Pseudomonas spp. Appl. Environ. Microbiol. 2001, 67, 1198–1209. [Google Scholar] [CrossRef] [Green Version]
- Whiteley, M.; Diggle, S.P.; Greenberg, E.P. Progress in and promise of bacterial quorum sensing research. Nature 2017, 551, 313–320. [Google Scholar] [CrossRef]
- Bassler, B.L. Small Talk: Cell-to-Cell Communication in Bacteria. Cell 2002, 109, 421–424. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Shaaban, M.I.; Bar, F.A.; El-Mahdy, A.; Shokralla, S. Quorum Sensing Inhibiting Activity of Streptomyces coelicoflavus Isolated from Soil. Front. Microbiol. 2016, 7, 659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esmaeel, Q.; Sanchez, L.; Robineau, M.; Dorey, S.; Clément, C.; Jacquard, C.; Ait Barka, E. Paraburkholderia phytofirmans PsJN-Plants Interaction: From Perception to the Induced Mechanisms. Front. Microbiol. 2018, 9, 2093. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Ayesha; Hameed, S.; Imran, A.; Iqbal, M.; Iqbal, J.; Oresnik, I.J. Functional characterization of a soybean growth stimulator Bradyrhizobium sp. strain SR-6 showing acylhomoserine lactone production. FEMS Microbiol. Ecol. 2016, 92, fiw115. [Google Scholar] [CrossRef] [Green Version]
- Jung, B.K.; Khan, A.R.; Hong, S.-J.; Park, G.-S.; Park, Y.-J.; Kim, H.-J.; Jeon, H.-J.; Khan, M.A.; Waqas, M.; Lee, I.-J.; et al. Quorum sensing activity of the plant growth-promoting rhizobacterium Serratia glossinae GS2 isolated from the sesame (Sesamum indicum L.) rhizosphere. Ann. Microbiol. 2017, 67, 623–632. [Google Scholar] [CrossRef]
- Hori, K.; Matsumoto, S. Bacterial adhesion: From mechanism to control. Biochem. Eng. J. 2010, 48, 424–434. [Google Scholar] [CrossRef]
- Mukherjee, S.; Bassler, B.L. Bacterial quorum sensing in complex and dynamically changing environments. Nat. Rev. Microbiol. 2019, 17, 371–382. [Google Scholar] [CrossRef]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef]
- Böhm, M.; Hurek, T.; Reinhold-Hurek, B. Twitching Motility Is Essential for Endophytic Rice Colonization by the N2-Fixing Endophyte Azoarcus sp. Strain BH72. Mol. Plant Microbe Interact. 2007, 20, 526–533. [Google Scholar] [CrossRef] [Green Version]
- Sivakumar, N.; Sathishkumar, R.; Selvakumar, G.; Shyamkumar, R.; Arjunekumar, K. Phyllospheric Microbiomes: Diversity, Ecological Significance, and Biotechnological Applications. Plant Microbiomes Sustain. Agric. 2020, 25, 113–172. [Google Scholar] [CrossRef]
- Saranraj, P. Screening of Pectinase Producing Bacteria and Fungi for Its Pectinolytic Activity Using Fruit Wastes. Int. J. Biochem. Biotechnol. Sci. 2012, 1, 30–40. [Google Scholar]
- Beldman, G.; Leeuwen, M.F.S.-V.; Rombouts, F.M.; Voragen, F.G.J. The cellulase of Trichoderma viride. JBIC J. Biol. Inorg. Chem. 1985, 146, 301–308. [Google Scholar] [CrossRef]
- Zhang, X.-Z.; Zhang, Y.-H.P. Cellulases: Characteristics, Sources, Production, and Applications. In Bioprocessing Technologies in Biorefinery for Sustainable Production of Fuels, Chemicals, and Polymers; Yang, S.-T., El-Enshasy, H.A., Thongchul, N., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; Volume 8, pp. 131–146. ISBN 978-1-118-64204-7. [Google Scholar]
- Reinhold-Hurek, B.; Maes, T.; Gemmer, S.; Van Montagu, M.; Hurek, T. An Endoglucanase Is Involved in Infection of Rice Roots by the Not-Cellulose-Metabolizing Endophyte Azoarcus sp. Strain BH72. Mol. Plant Microbe Interact. 2006, 19, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Truyens, S.; Weyens, N.; Cuypers, A.; Vangronsveld, J. Bacterial seed endophytes: Genera, vertical transmission and interaction with plants. Environ. Microbiol. Rep. 2014, 7, 40–50. [Google Scholar] [CrossRef]
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo- and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Turner, T.R.; James, E.K.; Poole, P.S. The plant microbiome. Genome Biol. 2013, 14, 209. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; Duan, J.; Charles, T.; Glick, B.R. A bioinformatics approach to the determination of genes involved in endophytic behavior in Burkholderia spp. J. Theor. Biol. 2014, 343, 193–198. [Google Scholar] [CrossRef]
- Khare, E.; Mishra, J.; Arora, N.K. Multifaceted Interactions Between Endophytes and Plant: Developments and Prospects. Front. Microbiol. 2018, 9, 2732. [Google Scholar] [CrossRef]
- Zeidler, D.; Zähringer, U.; Gerber, I.; Dubery, I.; Hartung, T.; Bors, W.; Hutzler, P.; Durner, J. Innate immunity in Arabidopsis thaliana: Lipopolysaccharides activate nitric oxide synthase (NOS) and induce defense genes. Proc. Natl. Acad. Sci. USA 2004, 101, 15811–15816. [Google Scholar] [CrossRef] [Green Version]
- Taghavi, S.; van der Lelie, D.; Hoffman, A.; Zhang, Y.-B.; Walla, M.D.; Vangronsveld, J.; Newman, L.; Monchy, S. Genome Sequence of the Plant Growth Promoting Endophytic Bacterium Enterobacter sp. 638. PLoS Genet. 2010, 6, e1000943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallet, B. Playing Dr Jekyll and Mr Hyde: Combined mechanisms of phase variation in bacteria. Curr. Opin. Microbiol. 2001, 4, 570–581. [Google Scholar] [CrossRef] [Green Version]
- Yu, K.; Yu, K.; Pieterse, C.M.; Pieterse, C.M.; Bakker, P.A.; Bakker, P.A.; Berendsen, R.L.; Berendsen, R.L.; Yu, K.; Yu, K.; et al. Beneficial microbes going underground of root immunity. Plant Cell Environ. 2019, 42, 2860–2870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clay, N.K.; Adio, A.M.; Denoux, C.; Jander, G.; Ausubel, F.M. Glucosinolate Metabolites Required for an Arabidopsis Innate Immune Response. Science 2009, 323, 95–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millet, Y.A.; Danna, C.H.; Clay, N.K.; Songnuan, W.; Simon, M.D.; Werck-Reichhart, D.; Ausubel, F.M. Innate Immune Responses Activated in Arabidopsis Roots by Microbe-Associated Molecular Patterns. Plant Cell 2010, 22, 973–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akum, F.N.; Esteinbrenner, J.; Ebiedenkopf, D.; Eimani, J.; Kogel, K.-H. The Piriformospora indica effector PIIN_08944 promotes the mutualistic Sebacinalean symbiosis. Front. Plant Sci. 2015, 6, 906. [Google Scholar] [CrossRef] [Green Version]
- Davidescu, D.; Davidescu, V. Evaluation of Fertility by Plant and Soil Analysis; Taylor & Francis: Abingdon-on-Thames, UK; Abacus Press: London, UK, 1982; Volume 1, ISBN 978-0-85626-123-7. [Google Scholar]
- Alippi, A.; Perelló, A.; Sisterna; Greco, N.; Cordo, C. Potential of Spore-Forming Bacteria as Biocontrol Agents of Wheat Foliar Diseases under Laboratory and Greenhouse Conditions. J. Plant Dis. Prot. 2000, 107, 155–169. [Google Scholar]
- Mejri, S.; Siah, A.; Coutte, F.; Magnin-Robert, M.; Randoux, B.; Tisserant, B.; Krier, F.; Jacques, P.; Reignault, P.; Halama, P. Biocontrol of the wheat pathogen Zymoseptoria tritici using cyclic lipopeptides from Bacillus subtilis. Environ. Sci. Pollut. Res. 2017, 25, 29822–29833. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef]
- Cantoro, R.; Palazzini, J.M.; Yerkovich, N.; Miralles, D.J.; Chulze, S.N. Bacillus velezensis RC 218 as a biocontrol agent against Fusarium graminearum: Effect on penetration, growth and TRI5 expression in wheat spikes. BioControl 2020, 66, 259–270. [Google Scholar] [CrossRef]
- Palazzini, J.M.; Ramirez, M.; Alberione, E.; Torres, A.; Chulze, S. Osmotic stress adaptation, compatible solutes accumulation and biocontrol efficacy of two potential biocontrol agents on Fusarium head blight in wheat. Biol. Control 2009, 51, 370–376. [Google Scholar] [CrossRef]
- Palazzini, J.; Roncallo, P.; Cantoro, R.; Chiotta, M.; Yerkovich, N.; Palacios, S.; Echenique, V.; Torres, A.; Ramirez, M.; Karlovsky, P.; et al. Biocontrol of Fusarium graminearum sensu stricto, Reduction of Deoxynivalenol Accumulation and Phytohormone Induction by Two Selected Antagonists. Toxins 2018, 10, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Li, R.; Qin, S.; Huang, L.; Wei, L.; Brian, K. Screening of Antagonistic Strain against Fusarium Head Blight and Its Inhibition Efficiency. Available online: https://en.cnki.com.cn/Article_en/CJFDTotal-ZBJS201705002.htm (accessed on 6 June 2021).
- Wang, J.; Liu, J.; Chen, H.; Yao, J. Characterization of Fusarium graminearum inhibitory lipopeptide from Bacillus subtilis IB. Appl. Microbiol. Biotechnol. 2007, 76, 889–894. [Google Scholar] [CrossRef]
- Pan, D.; Mionetto, A.; Tiscornia, S.; Bettucci, L. Endophytic bacteria from wheat grain as biocontrol agents of Fusarium graminearum and deoxynivalenol production in wheat. Mycotoxin Res. 2015, 31, 137–143. [Google Scholar] [CrossRef]
- Crane, J.M.; Gibson, D.M.; Vaughan, R.H.; Bergstrom, G.C. Iturin Levels on Wheat Spikes Linked to Biological Control of Fusarium Head Blight by Bacillus amyloliquefaciens. Phytopathology 2013, 103, 146–155. [Google Scholar] [CrossRef] [Green Version]
- Gong, A.-D.; Li, H.-P.; Yuan, Q.-S.; Song, X.-S.; Yao, W.; He, W.-J.; Zhang, J.-B.; Liao, Y.-C. Antagonistic Mechanism of Iturin A and Plipastatin A from Bacillus amyloliquefaciens S76-3 from Wheat Spikes against Fusarium graminearum. PLoS ONE 2015, 10, e0116871. [Google Scholar] [CrossRef] [Green Version]
- Jamal, Q.; Cho, J.-Y.; Moon, J.-H.; Kim, K.Y. Purification and antifungal characterization of Cyclo (D-Pro-L- Val) from Bacillus amyloliquefaciens Y1 against Fusarium graminearum to control head blight in wheat. Biocatal. Agric. Biotechnol. 2017, 10, 141–147. [Google Scholar] [CrossRef]
- Grosu, A.I.; Sicuia, O.-A.; Dobre, A.; Voaideş, C.; Cornea, C.P. Evaluation of Some Bacillus spp. Strains for the Biocontrol of Fusarium graminearum and F. culmorum in Wheat. Agric. Agric. Sci. Procedia 2015, 6, 559–566. [Google Scholar] [CrossRef] [Green Version]
- Zalila-Kolsi, I.; Ben Mahmoud, A.; Ali, H.; Sellami, S.; Nasfi, Z.; Tounsi, S.; Jamoussi, K. Antagonist effects of Bacillus spp. strains against Fusarium graminearum for protection of durum wheat (Triticum turgidum L. subsp. durum). Microbiol. Res. 2016, 192, 148–158. [Google Scholar] [CrossRef]
- Kildea, S.; Ransbotyn, V.; Khan, M.R.; Fagan, B.; Leonard, G.; Mullins, E.; Doohan, F.M. Bacillus megaterium shows potential for the biocontrol of septoria tritici blotch of wheat. Biol. Control 2008, 47, 37–45. [Google Scholar] [CrossRef]
- Nourozian, J.; Etebarian, H.R.; Khodakaramian, G. Biological Control of Fusarium graminearum on Wheat by Antagonistic Bacteria. Songklanakarin J. Sci. Technol. 2006, 28, 29–38. [Google Scholar]
- Palazzini, J.M.; Ramirez, M.L.; Torres, A.M.; Chulze, S.N. Potential biocontrol agents for Fusarium head blight and deoxynivalenol production in wheat. Crop Prot. 2007, 26, 1702–1710. [Google Scholar] [CrossRef]
- Palazzini, J.M.; Alberione, E.; Torres, A.; Donat, C.; Köhl, J.; Chulze, S. Biological control of Fusarium graminearum sensu stricto, causal agent of Fusarium head blight of wheat, using formulated antagonists under field conditions in Argentina. Biol. Control 2016, 94, 56–61. [Google Scholar] [CrossRef]
- Palazzini, J.M.; Dunlap, C.; Bowman, M.J.; Chulze, S.N. Bacillus velezensis RC 218 as a biocontrol agent to reduce Fusarium head blight and deoxynivalenol accumulation: Genome sequencing and secondary metabolite cluster profiles. Microbiol. Res. 2016, 192, 30–36. [Google Scholar] [CrossRef]
- Palazzini, J.; Torres, A.; Chulze, S. Tolerance of triazole-based fungicides by biocontrol agents used to control Fusarium head blight in wheat in Argentina. Lett. Appl. Microbiol. 2018, 66, 434–438. [Google Scholar] [CrossRef]
- Colombo, E.M.; Kunova, A.; Pizzatti, C.; Saracchi, M.; Cortesi, P.; Pasquali, M. Selection of an Endophytic Streptomyces sp. Strain DEF09 from Wheat Roots as a Biocontrol Agent Against Fusarium graminearum. Front. Microbiol. 2019, 10, 2356. [Google Scholar] [CrossRef] [Green Version]
- Jungkwan, L.; Park, S.-Y.; Lee, Y.-W.; Lee, J. Biological Efficacy of Streptomyces sp. Strain BN1 against the Cereal Head Blight Pathogen Fusarium graminearum. Plant Pathol. J. 2013, 29, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Bello, G.D.; Mónaco, C.; Simon, M.R. Biological control of seedling blight of wheat caused by Fusarium graminearum with beneficial rhizosphere microorganisms. World J. Microbiol. Biotechnol. 2002, 18, 627–636. [Google Scholar] [CrossRef]
- Khan, N.I.; Schisler, D.A.; Boehm, M.J.; Slininger, P.J.; Bothast, R.J. Selection and Evaluation of Microorganisms for Biocontrol of Fusarium Head Blight of Wheat Incited by Gibberella zeae. Plant Dis. 2001, 85, 1253–1258. [Google Scholar] [CrossRef] [Green Version]
- Schisler, D.A.; Khan, N.I.; Boehm, M.J.; Slininger, P.J. Greenhouse and Field Evaluation of Biological Control of Fusarium Head Blight on Durum Wheat. Plant Dis. 2002, 86, 1350–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunlap, C.A.; Schisler, D.A.; Bowman, M.J.; Rooney, A.P. Genomic analysis of Bacillus subtilis OH 131.1 and co-culturing with Cryptococcus flavescens for control of Fusarium head blight. Plant Gene 2015, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lynch, K.; Zannini, E.; Guo, J.; Axel, C.; Arendt, E.K.; Kildea, S.; Coffey, A. Control of Zymoseptoria tritici cause of septoria tritici blotch of wheat using antifungal Lactobacillus strains. J. Appl. Microbiol. 2016, 121, 485–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavermicocca, P.; Valerio, F.; Evidente, A.; Lazzaroni, S.; Corsetti, A.; Gobbetti, M. Purification and Characterization of Novel Antifungal Compounds from the Sourdough Lactobacillus plantarum Strain 21B. Appl. Environ. Microbiol. 2000, 66, 4084–4090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lounaci, L.; Guemouri-Athmani, S.; Boureghda, H.; Achouak, W.; Heulin, T. Suppression of crown and root rot of wheat by the rhizobacterium Paenibacillus polymyxa. Phytopathol. Mediterr. 2017, 55, 355–365. [Google Scholar]
- He, J.; Boland, G.; Zhou, T. Concurrent Selection for Microbial Suppression of Fusarium graminearum, Fusarium Head Blight and Deoxynivalenol in Wheat. J. Appl. Microbiol. 2009, 106, 1805–1817. [Google Scholar] [CrossRef]
- Samain, E.; Aussenac, T.; Selim, S. The Effect of Plant Genotype, Growth Stage, and Mycosphaerella graminicola Strains on the Efficiency and Durability of Wheat-Induced Resistance by Paenibacillus sp. Strain B2. Front. Plant Sci. 2019, 10, 587. [Google Scholar] [CrossRef] [Green Version]
- Levy, E.; Eyal, Z.; Carmely, S.; Kashman, Y.; Chet, I. Suppression of Septoria tritici and Puccinia recondita of wheat by an antibiotic-producing fluorescent pseudomonad. Plant Pathol. 1989, 38, 564–570. [Google Scholar] [CrossRef]
- Levy, E.; Gough, F.J.; Berlin, K.D.; Guiana, P.W.; Smith, J.T. Inhibition of Septoria tritici and other phytopathogenic fungi and bacteria by Pseudomonas fluorescens and its antibiotics. Plant Pathol. 1992, 41, 335–341. [Google Scholar] [CrossRef]
- Schisler, D.; Khan, N.; Boehm, M.; Lipps, P.; Slininger, P.; Zhang, S. Selection and evaluation of the potential of choline-metabolizing microbial strains to reduce Fusarium head blight. Biol. Control 2006, 39, 497–506. [Google Scholar] [CrossRef]
- Flaishman, M.A.; Eyal, Z.; Voisard, C.; Haas, D. Suppression of Septoria Tritici Blotch and Leaf Rust of Wheat by Recombiant Cyanide-Producing Strains of Pseudomonas putida. Mol. Plant-Microbe Interact. 1996, 9, 642. [Google Scholar] [CrossRef]
- Flaishman, M.; Eyal, Z.; Voisard, C.; Haas, D. Suppression of Septoria tritici by Phenazine- or Siderophore-deficient mutants ofPseudomonas. Curr. Microbiol. 1990, 20, 121–124. [Google Scholar] [CrossRef]
- Johnsson, L.; Hökeberg, M.; Gerhardson, B. Performance of the Pseudomonas chlororaphis biocontrol agent MA 342 against cereal seed-borne diseases in field experiments. Eur. J. Plant Pathol. 1998, 104, 701–711. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, J.; Yang, N.; Wen, Z.; Sun, X.; Chai, Y.; Ma, Z. Wheat microbiome bacteria can reduce virulence of a plant pathogenic fungus by altering histone acetylation. Nat. Commun. 2018, 9, 3429. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.-Y.; Xie, Y.-S.; Cui, Y.-Y.; Xu, J.; He, W.; Chen, H.-G.; Guo, J.-H. Conjunctively screening of biocontrol agents (BCAs) against fusarium root rot and fusarium head blight caused by Fusarium graminearum. Microbiol. Res. 2015, 177, 34–42. [Google Scholar] [CrossRef]
- Morimura, H.; Ito, M.; Yoshida, S.; Koitabashi, M.; Tsushima, S.; Camagna, M.; Chiba, S.; Takemoto, D.; Kawakita, K.; Sato, I. In Vitro Assessment of Biocontrol Effects on Fusarium Head Blight and Deoxynivalenol (DON) Accumulation by DON-Degrading Bacteria. Toxins 2020, 12, 399. [Google Scholar] [CrossRef]
- Khan, M.R.; Brien, E.O.; Carney, B.F.; Doohan, F.M. A fluorescent pseudomonad shows potential for the control of net blotch disease of barley. Biol. Control 2010, 54, 41–45. [Google Scholar] [CrossRef]
- Hökeberg, M.; Gerhardson, B.; Johnsson, L. Biological control of cereal seed-borne diseases by seed bacterization with greenhouse-selected bacteria. Eur. J. Plant Pathol. 1997, 103, 25–33. [Google Scholar] [CrossRef]
- Tombolini, R.; van der Gaag, D.J.; Gerhardson, B.; Jansson, J.K. Colonization Pattern of the Biocontrol Strain Pseudomonas chlororaphis MA 342 on Barley Seeds Visualized by Using Green Fluorescent Protein. Appl. Environ. Microbiol. 1999, 65, 3674–3680. [Google Scholar] [CrossRef] [Green Version]
- Chemitei, K.; Amendi, M.B.; Mwamburi, L.A.; Ochuodho, J.O. Bio-Control of Net-Blotch and Scald Pathogens of Barley Using Paenibacillus Polymyxa KAI245 Isolated from Sorghum Rhizosphere in Western Kenya. Am. J. Microbiol. Res. 2019, 7, 28–36. [Google Scholar] [CrossRef]
- Backes, A.; Vaillant-Gaveau, N.; Esmaeel, Q.; Barka, E.A.; Jacquard, C. A biological agent modulates the physiology of barley infected with Drechslera teres. Sci. Rep. 2021, 11, 8330. [Google Scholar] [CrossRef]
- Backes, A.; Charton, S.; Planchon, S.; Esmaeel, Q.; Sergeant, K.; Hausman, J.-F.; Renaut, J.; Barka, E.A.; Jacquard, C.; Guerriero, G. Gene expression and metabolite analysis in barley inoculated with net blotch fungus and plant growth-promoting rhizobacteria. Plant Physiol. Biochem. 2021, 168, 488–500. [Google Scholar] [CrossRef]
- Romera, F.J.; García, M.J.; Lucena, C.; Martinez-Medina, A.; Aparicio, M.A.; Ramos, J.; Alcántara, E.; Angulo, M.; Pérez-Vicente, R. Induced Systemic Resistance (ISR) and Fe Deficiency Responses in Dicot Plants. Front. Plant Sci. 2019, 10, 287. [Google Scholar] [CrossRef]
- Ahn, I.-P.; Lee, S.-W.; Suh, S.-C. Rhizobacteria-Induced Priming in Arabidopsis Is Dependent on Ethylene, Jasmonic Acid, and NPR1. Mol. Plant Microbe Interact. 2007, 20, 759–768. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.; Sultana, F.; Kubota, M.; Hyakumachi, M. Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp. GP16-2 and its cell free filtrate. Plant Soil 2008, 304, 227–239. [Google Scholar] [CrossRef]
- Iavicoli, A.; Boutet, E.; Buchala, A.; Métraux, J.-P. Induced Systemic Resistance in Arabidopsis thaliana in Response to Root Inoculation with Pseudomonas fluorescens CHA0. Mol. Plant Microbe Interact. 2003, 16, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Ryu, C.-M.; Murphy, J.F.; Mysore, K.S.; Kloepper, J.W. Plant growth-promoting rhizobacteria systemically protect Arabidopsis thaliana against Cucumber mosaic virus by a salicylic acid and NPR1-independent and jasmonic acid-dependent signaling pathway. Plant J. 2004, 39, 381–392. [Google Scholar] [CrossRef]
- Weller, D.M.; Mavrodi, D.V.; van Pelt, J.A.; Pieterse, C.M.J.; van Loon, L.C.; Bakker, P.A.H.M. Induced Systemic Resistance in Arabidopsis thaliana Against Pseudomonas syringae pv. tomato by 2,4-Diacetylphloroglucinol-Producing Pseudomonas fluorescens. Phytopathology 2012, 102, 403–412. [Google Scholar] [CrossRef] [Green Version]
- Katsir, L.; Chung, H.S.; Koo, A.J.; Howe, G.A. Jasmonate signaling: A conserved mechanism of hormone sensing. Curr. Opin. Plant Biol. 2008, 11, 428–435. [Google Scholar] [CrossRef] [Green Version]
- Pozo, M.J.; Van Der Ent, S.; Van Loon, L.C.; Pieterse, C.M.J. Transcription factor MYC2 is involved in priming for enhanced defense during rhizobacteria-induced systemic resistance in Arabidopsis thaliana. New Phytol. 2008, 180, 511–523. [Google Scholar] [CrossRef]
- Galletti, R.; Ferrari, S.; De Lorenzo, G. Arabidopsis MPK3 and MPK6 Play Different Roles in Basal and Oligogalacturonide- or Flagellin-Induced Resistance against Botrytis cinerea. Plant Physiol. 2011, 157, 804–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stringlis, I.A.; Yu, K.; Feussner, K.; de Jonge, R.; Van Bentum, S.; Van Verk, M.C.; Berendsen, R.L.; Bakker, P.A.H.M.; Feussner, I.; Pieterse, C.M.J. MYB72-dependent coumarin exudation shapes root microbiome assembly to promote plant health. Proc. Natl. Acad. Sci. USA 2018, 115, E5213–E5222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oleńska, E.; Małek, W.; Wójcik, M.; Swiecicka, I.; Thijs, S.; Vangronsveld, J. Beneficial features of plant growth-promoting rhizobacteria for improving plant growth and health in challenging conditions: A methodical review. Sci. Total Environ. 2020, 743, 140682. [Google Scholar] [CrossRef] [PubMed]
- Petti, C.; Khan, M.; Doohan, F. Lipid transfer proteins and protease inhibitors as key factors in the priming of barley responses to Fusarium head blight disease by a biocontrol strain of Pseudomonas fluorescens. Funct. Integr. Genom. 2010, 10, 619–627. [Google Scholar] [CrossRef]
- Mur, L.A.; Kenton, P.; Atzorn, R.; Miersch, O.; Wasternack, C. The Outcomes of Concentration-Specific Interactions between Salicylate and Jasmonate Signaling Include Synergy, Antagonism, and Oxidative Stress Leading to Cell Death. Plant Physiol. 2005, 140, 249–262. [Google Scholar] [CrossRef] [Green Version]
- Nie, P.; Li, X.; Wang, S.; Guo, J.; Zhao, H.; Niu, D. Induced Systemic Resistance against Botrytis cinerea by Bacillus cereus AR156 through a JA/ET- and NPR1-Dependent Signaling Pathway and Activates PAMP-Triggered Immunity in Arabidopsis. Front. Plant Sci. 2017, 8, 238. [Google Scholar] [CrossRef]
- Pastor, V.; Luna, E.; Mauch-Mani, B.; Ton, J.; Flors, V. Primed plants do not forget. Environ. Exp. Bot. 2013, 94, 46–56. [Google Scholar] [CrossRef]
- Iriti, M. Plant Innate Immunity; Editorial Office; MDPI: Basel, Switzerland, 2019; Volume 2, ISBN 978-3-03897-580-9. [Google Scholar]
- Bhattacharyya, D.; Garladinne, M.; Lee, Y.H. Volatile Indole Produced by Rhizobacterium Proteus vulgaris JBLS202 Stimulates Growth of Arabidopsis thaliana Through Auxin, Cytokinin, and Brassinosteroid Pathways. J. Plant Growth Regul. 2014, 34, 158–168. [Google Scholar] [CrossRef]
- Beckers, G.J.; Jaskiewicz, M.; Liu, Y.; Underwood, W.R.; He, S.Y.; Zhang, S.; Conrath, U. Mitogen-Activated Protein Kinases 3 and 6 Are Required for Full Priming of Stress Responses in Arabidopsis thaliana. Plant Cell 2009, 21, 944–953. [Google Scholar] [CrossRef] [Green Version]
- Tateda, C.; Zhang, Z.; Shrestha, J.; Jelenska, J.; Chinchilla, D.; Greenberg, J.T. Salicylic Acid Regulates Arabidopsis Microbial Pattern Receptor Kinase Levels and Signaling. Plant Cell 2014, 26, 4171–4187. [Google Scholar] [CrossRef] [Green Version]
- Bonanomi, G.; D’Ascoli, R.; Scotti, R.; Gaglione, S.; Caceres, M.G.; Sultana, S.; Scelza, R.; Rao, M.A.; Zoina, A. Soil quality recovery and crop yield enhancement by combined application of compost and wood to vegetables grown under plastic tunnels. Agric. Ecosyst. Environ. 2014, 192, 1–7. [Google Scholar] [CrossRef]
- Haas, D.; Défago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Genet. 2005, 3, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Legrand, F.; Picot, A.; Díaz, J.F.C.; Chen, W.; Le Floch, G. Challenges facing the biological control strategies for the management of Fusarium Head Blight of cereals caused by F. graminearum. Biol. Control 2017, 113, 26–38. [Google Scholar] [CrossRef]
- Ullah, H.; Yasmin, H.; Mumtaz, S.; Jabeen, Z.; Naz, R.; Nosheen, A.; Hassan, M.N. Multitrait Pseudomonas spp. Isolated from Monocropped Wheat (Triticum aestivum) Suppress Fusarium Root and Crown Rot. Phytopathology 2020, 110, 582–592. [Google Scholar] [CrossRef]
- Sharma, A.; Johri, B. Growth promoting influence of siderophore-producing Pseudomonas strains GRP3A and PRS9 in maize (Zea mays L.) under iron limiting conditions. Microbiol. Res. 2003, 158, 243–248. [Google Scholar] [CrossRef]
- Lambrese, Y.; Guiñez, M.; Calvente, V.; Sansone, G.; Cerutti, S.; Raba, J.; Sanz, M.I. Production of siderophores by the bacterium Kosakonia radicincitans and its application to control of phytopathogenic fungi. Bioresour. Technol. Rep. 2018, 3, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Thakur, S.; Dhingra, G.; Singh, A.; Pal, M.K.; Harshvardhan, K.; Dubey, R.; Maheshwari, D. Inoculation of siderophore producing rhizobacteria and their consortium for growth enhancement of wheat plant. Biocatal. Agric. Biotechnol. 2018, 15, 264–269. [Google Scholar] [CrossRef]
- Sayyed, R.Z.; Chincholkar, S.B.; Reddy, M.S.; Gangurde, N.S.; Patel, P.R. Siderophore Producing PGPR for Crop Nutrition and Phytopathogen Suppression. In Bacteria in Agrobiology: Disease Management; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 449–471. ISBN 978-3-642-33638-6. [Google Scholar]
- Trapet, P.; Avoscan, L.; Klinguer, A.; Pateyron, S.; Citerne, S.; Chervin, C.; Mazurier, S.; Lemanceau, P.; Wendehenne, D.; Besson-Bard, A. The Pseudomonas fluorescens Siderophore Pyoverdine Weakens Arabidopsis thaliana Defense in Favor of Growth in Iron-Deficient Conditions. Plant Physiol. 2016, 171, 675–693. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, M.J.; Silva, H.; Cunha, A. Siderophore-Producing Rhizobacteria as a Promising Tool for Empowering Plants to Cope with Iron Limitation in Saline Soils: A Review. Pedosphere 2019, 29, 409–420. [Google Scholar] [CrossRef]
- Morot-Gaudry, J.F. Nitrogen Assimilation by Plants: Physiological, Biochemical, and Molecular Aspects; CRC Press: Boca Raton, FL, USA, 2001; ISBN 978-1-4822-7984-9. [Google Scholar]
- Söğüt, S.; Çiğ, F. Determination of the effect of plant growth promoting bacteria on wheat (Triticum aestivum L.) Development under salinity stress conditions. Appl. Ecol. Environ. Res. 2019, 17, 1129–1141. [Google Scholar] [CrossRef]
- Barh, D.; Azevedo, V. Omics Technologies and Bio-Engineering: Towards Improving Quality of Life; Barh, D., Azevedo, V., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 1, ISBN 978-0-12-815899-9. [Google Scholar]
- Takenaka, S.; Nishio, Z.; Nakamura, Y. Induction of Defense Reactions in Sugar Beet and Wheat by Treatment with Cell Wall Protein Fractions from the Mycoparasite Pythium oligandrum. Phytopathology 2003, 93, 1228–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esmaeel, Q.; Pupin, M.; Jacques, P.; Leclère, V. Nonribosomal peptides and polyketides of Burkholderia: New compounds potentially implicated in biocontrol and pharmaceuticals. Environ. Sci. Pollut. Res. 2017, 25, 29794–29807. [Google Scholar] [CrossRef] [PubMed]
- Elshafie, H.; Camele, I. An Overview of Metabolic Activity, Beneficial and Pathogenic Aspects of Burkholderia spp. Metabolites 2021, 11, 321. [Google Scholar] [CrossRef] [PubMed]
- Saad, M. Destruction of Rhizoctonia Solani and Phytophthora Capsici Causing Tomato Root-Rot by Pseudomonas fluorescences Lytic Enzymes. Res. J. Agric. Biol. Sci. 2006, 2, 274–281. [Google Scholar]
- Zhao, Y.; Selvaraj, J.N.; Xing, F.; Zhou, L.; Wang, Y.; Song, H.; Tan, X.; Sun, L.; Sangare, L.; Folly, Y.M.E.; et al. Antagonistic Action of Bacillus subtilis Strain SG6 on Fusarium graminearum. PLoS ONE 2014, 9, e92486. [Google Scholar] [CrossRef]
- Beauregard, P.B.; Chai, Y.; Vlamakis, H.; Losick, R.; Kolter, R. Bacillus subtilis biofilm induction by plant polysaccharides. Proc. Natl. Acad. Sci. USA 2013, 110, E1621–E1630. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.-H.; Xu, J.-L.; Li, X.-Z.; Zhang, L.-H. AiiA, an enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Proc. Natl. Acad. Sci. USA 2000, 97, 3526–3531. [Google Scholar] [CrossRef]
- Bohm, K.; Martín-Sánchez, L.; Garbeva, P. Microbial Volatiles: Small Molecules with an Important Role in Intra- and Inter-Kingdom Interactions. Front. Microbiol. 2017, 8, 2484. [Google Scholar] [CrossRef]
- Farag, M.A.; Ryu, C.-M.; Sumner, L.W.; Paré, P.W. GC–MS SPME profiling of rhizobacterial volatiles reveals prospective inducers of growth promotion and induced systemic resistance in plants. Phytochemistry 2006, 67, 2262–2268. [Google Scholar] [CrossRef]
- Li, X.-Y.; Mao, Z.-C.; Wu, Y.-X.; Ho, H.-H.; He, Y.-Q. Comprehensive volatile organic compounds profiling of Bacillus species with biocontrol properties by head space solid phase microextraction with gas chromatography-mass spectrometry. Biocontrol Sci. Technol. 2014, 25, 132–143. [Google Scholar] [CrossRef]
- Boukaew, S.; Cheirsilp, B.; Prasertsan, P.; Yossan, S. Antifungal effect of volatile organic compounds produced by Streptomyces salmonis PSRDC-09 against anthracnose pathogen Colletotrichum gloeosporioides PSU-03 in postharvest chili fruit. J. Appl. Microbiol. 2021, 131, 1452–1463. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yuan, J.; E, Y.; Raza, W.; Shen, Q.; Huang, Q. Effects of volatile organic compounds from Streptomyces albulus NJZJSA2 on growth of two fungal pathogens. J. Basic Microbiol. 2015, 55, 1104–1117. [Google Scholar] [CrossRef] [PubMed]
- Esmaeel, Q.; Pupin, M.; Kieu, N.P.; Chataigné, G.; Béchet, M.; Deravel, J.; Krier, F.; Höfte, M.; Jacques, P.; Leclère, V. Burkholderia genome mining for nonribosomal peptide synthetases reveals a great potential for novel siderophores and lipopeptides synthesis. MicrobiologyOpen 2016, 5, 512–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimkić, I.; Janakiev, T.; Petrović, M.; Degrassi, G.; Fira, D. Plant-associated Bacillus and Pseudomonas antimicrobial activities in plant disease suppression via biological control mechanisms—A review. Physiol. Mol. Plant Pathol. 2021, 117, 101754. [Google Scholar] [CrossRef]
- Hoskisson, P.A.; van Wezel, G.P. Streptomyces coelicolor. Trends Microbiol. 2019, 27, 468–469. [Google Scholar] [CrossRef]
- Andrić, S.; Meyer, T.; Ongena, M. Bacillus Responses to Plant-Associated Fungal and Bacterial Communities. Front. Microbiol. 2020, 11, 1350. [Google Scholar] [CrossRef]
- Oni, F.E.; Esmaeel, Q.; Onyeka, J.T.; Adeleke, R.; Jacquard, C.; Clement, C.; Gross, H.; Barka, E.A.; Höfte, M. Pseudomonas Lipopeptide-Mediated Biocontrol: Chemotaxonomy and Biological Activity. Molecules 2022, 27, 372. [Google Scholar] [CrossRef]
- Höfte, M. The Use of Pseudomonas spp. as Bacterial Biocontrol Agents to Control Plant Diseases. In Burleigh Dodds Series in Agricultural Science; Köhl, J., Ed.; Wageningen University & Research: Wageningen, The Netherlands; Burleigh Dodds Science Publishing: Cambridge, UK, 2021; pp. 301–374. ISBN 978-1-78676-813-1. [Google Scholar]
- Pierson, I.L.S.; Pierson, E.A. Phenazine antibiotic production in Pseudomonas aureofaciens: Role in rhizosphere ecology and pathogen suppression. FEMS Microbiol. Lett. 1996, 136, 101–108. [Google Scholar] [CrossRef]
- Thomashow, L.S.; Weller, D.M. Role of a phenazine antibiotic from Pseudomonas fluorescens in biological control of Gaeumannomyces graminis var. tritici. J. Bacteriol. 1988, 170, 3499–3508. [Google Scholar] [CrossRef] [Green Version]
- Keel, C.; Schnider, U.; Maurhofer, M.; Voisard, C.; Laville, J.; Burger, U.; Wirthner, P.J.; Haas, D.; Défago, G. Suppression of Root Diseases by Pseudomonas fluorescens CHA0: Importance of the Bacterial Secondary Metabolite 2,4-Diacetylphloroglucinol. Mol. Plant Microbe Interact. 1992, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Keel, C.; Oberhansli, T.; Haas, D.; Defag, G.; Wirthner, P. Pseudomonads as Antagonists of Plant Pathogens in the Rhizosphere: Role of the Antibiotic 2,4-Diacetylphloroglucinol in the Suppression of Black Root Rot of Tobacco. Symbiosis 1990, 9, 327–341. [Google Scholar]
- Baron, S.S.; Terranova, G.; Rowe, J.J. Molecular mechanism of the antimicrobial action of pyocyanin. Curr. Microbiol. 1989, 18, 223–230. [Google Scholar] [CrossRef]
- Baron, S.S.; Rowe, J.J. Antibiotic action of pyocyanin. Antimicrob. Agents Chemother. 1981, 20, 814–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLoughlin, A.J. Plasmid stability and ecological competence in recombinant cultures. Biotechnol. Adv. 1994, 12, 279–324. [Google Scholar] [CrossRef]
- Chin-A-Woeng, T.F.C.; Bloemberg, G.V.; Lugtenberg, B.J.J. Phenazines and their role in biocontrol by Pseudomonas bacteria. New Phytol. 2003, 157, 503–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, T.H.; Sørensen, D.; Tobiasen, C.; Andersen, J.B.; Christophersen, C.; Givskov, M.; Sørensen, J. Antibiotic and Biosurfactant Properties of Cyclic Lipopeptides Produced by Fluorescent Pseudomonas spp. from the Sugar Beet Rhizosphere. Appl. Environ. Microbiol. 2002, 68, 3416–3423. [Google Scholar] [CrossRef] [Green Version]
- Fira, D.; Dimkić, I.; Berić, T.; Lozo, J.; Stanković, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef]
- Anckaert, A.; Arias, A.A.; Hoff, G.; Calonne-Salmon, M.; Declerck, S.; Ongena, M. The Use of Bacillus spp. as Bacterial Biocontrol Agents to Control Plant Diseases. In Burleigh Dodds Series in Agricultural Science; Köhl, J., Ed.; Wageningen University & Research: Wageningen, The Netherlands; Burleigh Dodds Science Publishing: Cambridge, UK, 2021; pp. 247–300. ISBN 978-1-78676-813-1. [Google Scholar]
- Raaijmakers, J.M.; De Bruijn, I.; Nybroe, O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef] [Green Version]
- Falardeau, J.; Wise, C.; Novitsky, L.; Avis, T.J. Ecological and Mechanistic Insights Into the Direct and Indirect Antimicrobial Properties of Bacillus subtilis Lipopeptides on Plant Pathogens. J. Chem. Ecol. 2013, 39, 869–878. [Google Scholar] [CrossRef]
- Parent, Z.K. Potentiel de Bacillus Amyloliquefaciens Pour Lutter Contre les Maladies Fongiques Endémiques du Maïs au Sud Kivu: Efficacité et Mode d’Action. Ph.D. Thesis, Liège University Gembloux Agro-Bio Tech, Gembloux, Belgium, 2017. Available online: https://hdl.handle.net/2268/213753 (accessed on 4 January 2022).
- Dunlap, C.; Bowman, M.J.; Schisler, D.A. Genomic analysis and secondary metabolite production in Bacillus amyloliquefaciens AS 43.3: A biocontrol antagonist of Fusarium head blight. Biol. Control 2013, 64, 166–175. [Google Scholar] [CrossRef]
- Deleu, M.; Paquot, M.; Nylander, T. Fengycin interaction with lipid monolayers at the air–aqueous interface—Implications for the effect of fengycin on biological membranes. J. Colloid Interface Sci. 2004, 283, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Peypoux, F.; Bonmatin, J.M.; Wallach, J. Recent trends in the biochemistry of surfactin. Appl. Microbiol. Biotechnol. 1999, 51, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Buchoux, S.; Him, J.L.K.; Garnier, M.; Tsan, P.; Besson, F.; Brisson, A.; Dufourc, E.J. Surfactin-Triggered Small Vesicle Formation of Negatively Charged Membranes: A Novel Membrane-Lysis Mechanism. Biophys. J. 2008, 95, 3840–3849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maget-Dana, R.; Thimon, L.; Peypoux, F.; Ptak, M. Surfactin/iturin A interactions may explain the synergistic effect of surfactin on the biological properties of iturin A. Biochimie 1992, 74, 1047–1051. [Google Scholar] [CrossRef]
- Cawoy, H.; Mariutto, M.; Henry, G.; Fisher, C.; Vasilyeva, N.; Thonart, P.; Dommes, J.; Ongena, M. Plant Defense Stimulation by Natural Isolates of Bacillus Depends on Efficient Surfactin Production. Mol. Plant Microbe Interact. 2014, 27, 87–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bais, H.P.; Fall, R.; Vivanco, J.M. Biocontrol of Bacillus subtilis against Infection of Arabidopsis Roots by Pseudomonas syringae Is Facilitated by Biofilm Formation and Surfactin Production. Plant Physiol. 2004, 134, 307–319. [Google Scholar] [CrossRef] [Green Version]
- Kinsinger, R.F.; Shirk, M.C.; Fall, R. Rapid Surface Motility in Bacillus subtilis Is Dependent on Extracellular Surfactin and Potassium Ion. J. Bacteriol. 2003, 185, 5627–5631. [Google Scholar] [CrossRef] [Green Version]
- Leclère, V.; Marti, R.; Béchet, M.; Fickers, P.; Jacques, P. The lipopeptides mycosubtilin and surfactin enhance spreading of Bacillus subtilis strains by their surface-active properties. Arch. Microbiol. 2006, 186, 475–483. [Google Scholar] [CrossRef]
- Alshannaq, A.; Yu, J.-H. Occurrence, Toxicity, and Analysis of Major Mycotoxins in Food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef] [Green Version]
- Hathout, A.S.; Abel-Fattah, S.M.; Abou-Sree, Y.H.; Fouzy, A.S. Incidence and exposure assessment of aflatoxins and ochratoxin A in Egyptian wheat. Toxicol. Rep. 2020, 7, 867–873. [Google Scholar] [CrossRef]
- Audenaert, K.; Vanheule, A.; Höfte, M.; Haesaert, G. Deoxynivalenol: A Major Player in the Multifaceted Response of Fusarium to Its Environment. Toxins 2014, 6, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoev, S.D. Food Safety and Increasing Hazard of Mycotoxin Occurrence in Foods and Feeds. Crit. Rev. Food Sci. Nutr. 2013, 53, 887–901. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Fan, Y.; Zhao, L. Review on biological degradation of mycotoxins. Anim. Nutr. 2016, 2, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Altalhi, A.D. Plasmid-mediated Detoxification of Mycotoxin Zearalenone in Pseudomonas sp. ZEA-1. Am. J. Biochem. Biotechnol. 2007, 3, 150–158. [Google Scholar] [CrossRef]
- Yi, P.-J.; Pai, C.-K.; Liu, J.-R. Isolation and characterization of a Bacillus licheniformis strain capable of degrading zearalenone. World J. Microbiol. Biotechnol. 2010, 27, 1035–1043. [Google Scholar] [CrossRef]
- Cho, K.J.; Kang, J.S.; Cho, W.T.; Lee, C.H.; Ha, J.K.; Bin Song, K. In vitro degradation of zearalenone by Bacillus subtilis. Biotechnol. Lett. 2010, 32, 1921–1924. [Google Scholar] [CrossRef]
- Sato, I.; Ito, M.; Ishizaka, M.; Ikunaga, Y.; Sato, Y.; Yoshida, S.; Koitabashi, M.; Tsushima, S. Thirteen novel deoxynivalenol-degrading bacteria are classified within two genera with distinct degradation mechanisms. FEMS Microbiol. Lett. 2011, 327, 110–117. [Google Scholar] [CrossRef] [Green Version]
- He, J.W.; Bondy, G.S.; Zhou, T.; Caldwell, D.; Boland, G.J.; Scott, P.M. Toxicology of 3-epi-deoxynivalenol, a deoxynivalenol-transformation product by Devosia mutans 17-2-E-8. Food Chem. Toxicol. 2015, 84, 250–259. [Google Scholar] [CrossRef]
- Shima, J.; Takase, S.; Takahashi, Y.; Iwai, Y.; Fujimoto, H.; Yamazaki, M.; Ochi, K. Novel detoxification of the trichothecene mycotoxin deoxynivalenol by a soil bacterium isolated by enrichment culture. Appl. Environ. Microbiol. 1997, 63, 3825–3830. [Google Scholar] [CrossRef] [Green Version]
- Platel, R.; Lucau-Danila, A.; Baltenweck, R.; Maia-Grondard, A.; Chaveriat, L.; Magnin-Robert, M.; Randoux, B.; Trapet, P.; Halama, P.; Martin, P.; et al. Bioinspired Rhamnolipid Protects Wheat Against Zymoseptoria tritici Through Mainly Direct Antifungal Activity and without Major Impact on Leaf Physiology. Front. Plant Sci. 2022, 13, 1530. [Google Scholar] [CrossRef]
- Riseh, R.S.; Skorik, Y.A.; Thakur, V.K.; Pour, M.M.; Tamanadar, E.; Noghabi, S.S. Encapsulation of Plant Biocontrol Bacteria with Alginate as a Main Polymer Material. Int. J. Mol. Sci. 2021, 22, 11165. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Current strategies of plant protection used in the field. To fight the various biotic stresses and reduce fungal pressure, several strategies are used by the agricultural sector. Chemical control is the most widespread method worldwide because of its effectiveness and its almost systematic use over several generations. However, due to various risks to the environment and human health, other alternatives are increasingly used. Among these alternative solutions, the farmer can rely on various levers such as direct struggles (physical struggle, use of biocontrol, trap plants), indirect control (adaptation of cultivation practices, use of resistant varieties, auxiliaries, microbial ecology).
Figure 1.
Current strategies of plant protection used in the field. To fight the various biotic stresses and reduce fungal pressure, several strategies are used by the agricultural sector. Chemical control is the most widespread method worldwide because of its effectiveness and its almost systematic use over several generations. However, due to various risks to the environment and human health, other alternatives are increasingly used. Among these alternative solutions, the farmer can rely on various levers such as direct struggles (physical struggle, use of biocontrol, trap plants), indirect control (adaptation of cultivation practices, use of resistant varieties, auxiliaries, microbial ecology).

Figure 2.
Insights into some molecular factors in plant-pathogen interactions that may be involved in plant immunity. The perception of host damaged-associated molecular patterns (DAMPs), resulting from the interaction between endo-polygalacturonases (ePGs) secreted by some fungi and the PG-inhibiting proteins (PGIPs) secreted by the plant, through DAMP receptors (such as, DAMP receptor wall-associated receptor kinase 1 (WAK1)) triggers plant defense responses. Additionally, microbe-associated molecular patterns (MAMPs) are delivered from microbes to the apoplast (apoplastic effectors) or delivered inside host cells (cytoplasmic effectors) to perturb plant cell physiology. The MAMPs may be perceived by the cell surface pattern-recognition receptors (PRRs; receptor-like kinases (RLKs) or receptor-like proteins (RLPs)) and triggers downstream phosphorylation cascades and provoke an enhancement of [Ca2+] and reactive oxygen species (ROS). The activation of pathogen-responsive MAPK cascades is one of the earliest signaling events in PTI and ETI. The pathogen effectors are recognized by intracellular receptors, nucleotide binding site-leucine-rich-repeat (NLRs also known as NB-LRRs), triggering therefore downstream responses including Salicylic acid (SA) accumulation. The results of defense signaling involve modulation of gene expression, the synthesis of (PR) proteins, and biosynthesis of antimicrobial metabolites. More details in the main text. Ub, Ubiquitin. P, phosphate group. S, Small ubiquitin-like modifiers; ABA, Abscisic acid; BR, Brassinosteroid; CAM, Calmodulin; CPK, Calcium-dependent Protein Kinases; CY, Cytokinins; GA, Gibberellic acid; HR, Hypersensitive response; IAA, Indol acetic acid; JA-Ile, Jasmonoyl–isoleucine; JAZ, Jasmonate-zim-domain protein; LysM-RLK, Lysin motif receptor-like kinases; MAPKs, Mitogen-activated protein kinase; NPR, Nonexpressor of pathogenesis-related genes; PUB, Plant U-box; T3SS, type III secretion system.
Figure 2.
Insights into some molecular factors in plant-pathogen interactions that may be involved in plant immunity. The perception of host damaged-associated molecular patterns (DAMPs), resulting from the interaction between endo-polygalacturonases (ePGs) secreted by some fungi and the PG-inhibiting proteins (PGIPs) secreted by the plant, through DAMP receptors (such as, DAMP receptor wall-associated receptor kinase 1 (WAK1)) triggers plant defense responses. Additionally, microbe-associated molecular patterns (MAMPs) are delivered from microbes to the apoplast (apoplastic effectors) or delivered inside host cells (cytoplasmic effectors) to perturb plant cell physiology. The MAMPs may be perceived by the cell surface pattern-recognition receptors (PRRs; receptor-like kinases (RLKs) or receptor-like proteins (RLPs)) and triggers downstream phosphorylation cascades and provoke an enhancement of [Ca2+] and reactive oxygen species (ROS). The activation of pathogen-responsive MAPK cascades is one of the earliest signaling events in PTI and ETI. The pathogen effectors are recognized by intracellular receptors, nucleotide binding site-leucine-rich-repeat (NLRs also known as NB-LRRs), triggering therefore downstream responses including Salicylic acid (SA) accumulation. The results of defense signaling involve modulation of gene expression, the synthesis of (PR) proteins, and biosynthesis of antimicrobial metabolites. More details in the main text. Ub, Ubiquitin. P, phosphate group. S, Small ubiquitin-like modifiers; ABA, Abscisic acid; BR, Brassinosteroid; CAM, Calmodulin; CPK, Calcium-dependent Protein Kinases; CY, Cytokinins; GA, Gibberellic acid; HR, Hypersensitive response; IAA, Indol acetic acid; JA-Ile, Jasmonoyl–isoleucine; JAZ, Jasmonate-zim-domain protein; LysM-RLK, Lysin motif receptor-like kinases; MAPKs, Mitogen-activated protein kinase; NPR, Nonexpressor of pathogenesis-related genes; PUB, Plant U-box; T3SS, type III secretion system.

Figure 3.
Schematic of plant-bacteria interactions. Abbreviations used in the figure: reactive oxygen species (ROS), oxide anion (O2), hydrogen peroxide (H2O2), type III secretion system (T3SS), type VI secretion system (T6SS), microbe-associated molecular patterns (MAMPs), jasmonic acid (JA), ethylene (ET), salicylic acid (SA), pathogenesis-related proteins (PRs), the nonexpressor of pathogenesis-related gene 1 (NPR1), induced systemic resistance (ISR) and systemic acquired resistance (SAR).
Figure 3.
Schematic of plant-bacteria interactions. Abbreviations used in the figure: reactive oxygen species (ROS), oxide anion (O2), hydrogen peroxide (H2O2), type III secretion system (T3SS), type VI secretion system (T6SS), microbe-associated molecular patterns (MAMPs), jasmonic acid (JA), ethylene (ET), salicylic acid (SA), pathogenesis-related proteins (PRs), the nonexpressor of pathogenesis-related gene 1 (NPR1), induced systemic resistance (ISR) and systemic acquired resistance (SAR).


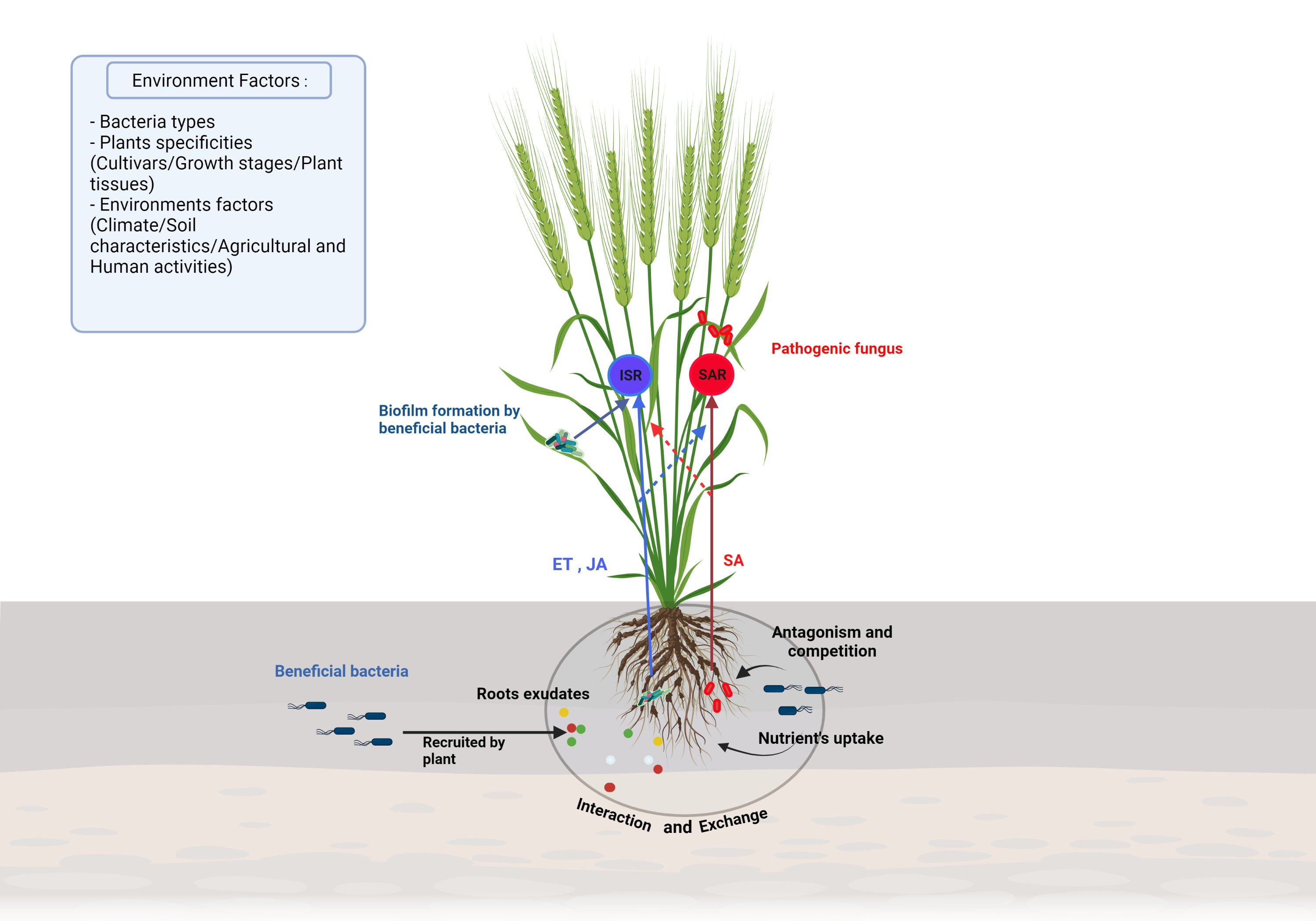{kind=link}
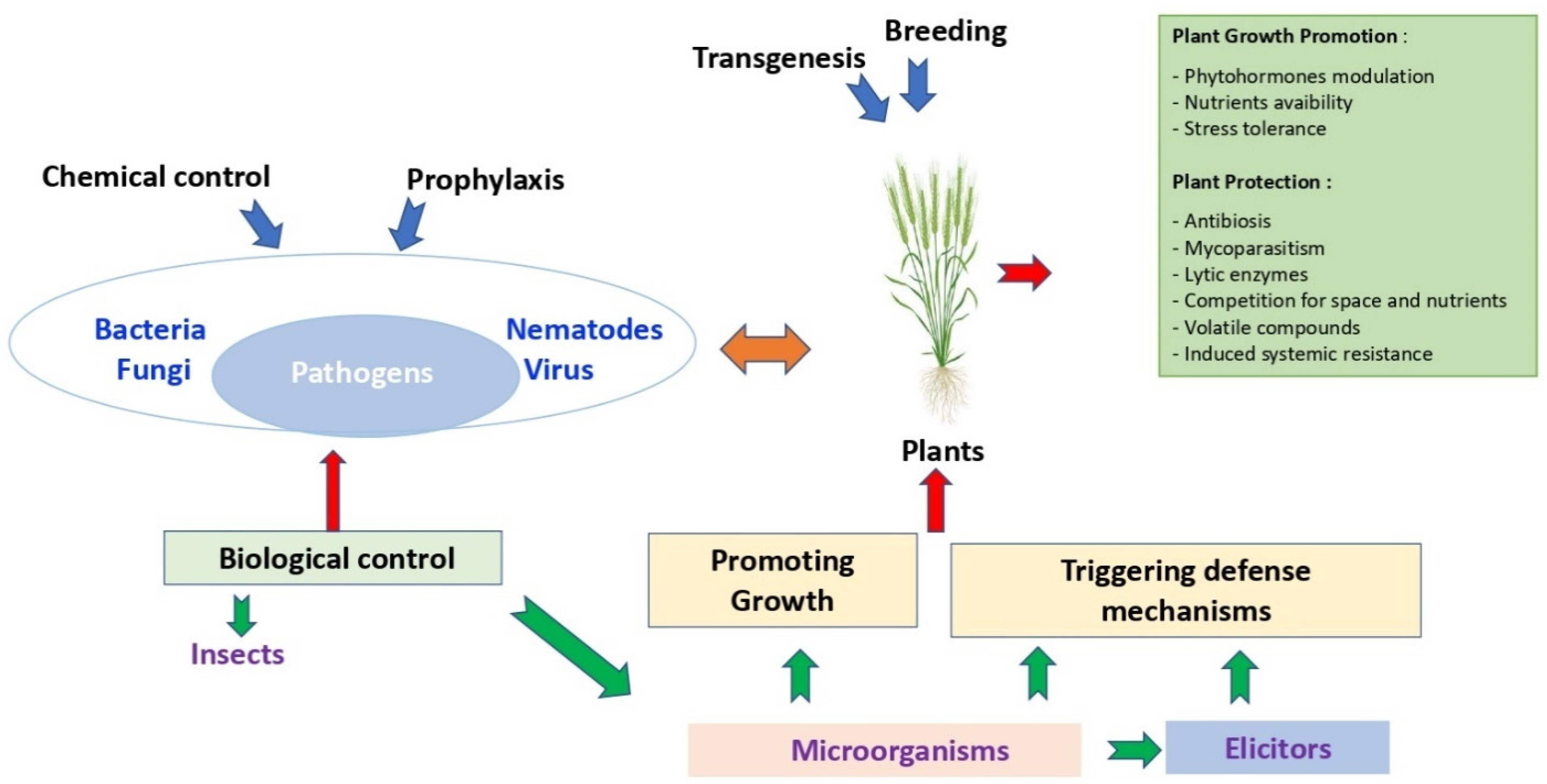{kind=link}
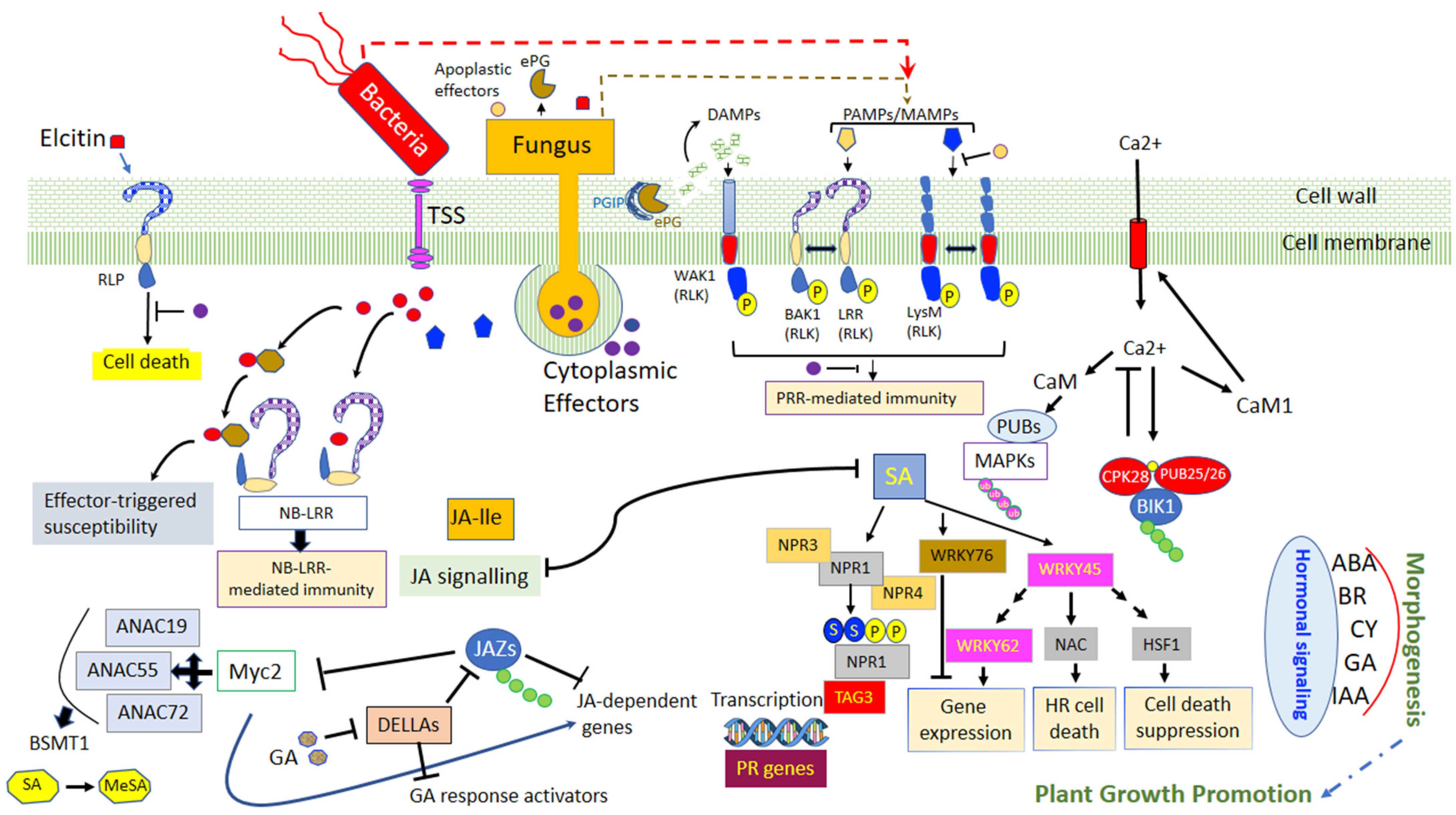{kind=link}
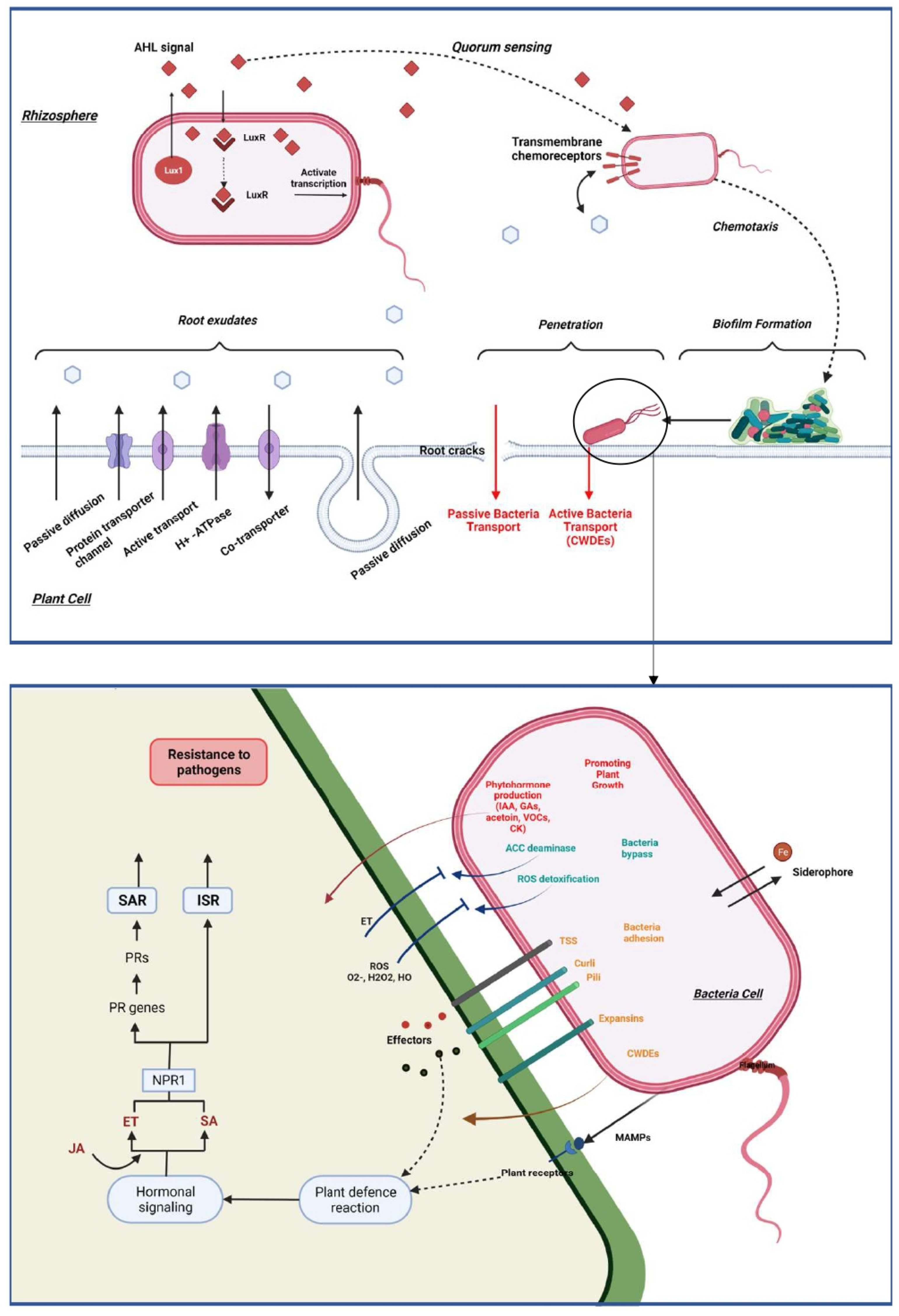{kind=link}
Table 1.
Commercial bacterium-based products against barley and wheat pathogens.
| Products Name | Company | Beneficial Microbes | Pathogen | Targeted Crop | Mode of Action for Biocontrol |
|---|---|---|---|---|---|
| Polyversum® | De Sangosse | Pythium oligandrum M1 | Fusarium and Sclerotina | Wheat, barley, and colza |
|
| Mycostop® | Lallemend | Streptomyces sp. K61 | Fusarium | Wheat, corn, barley, sugar-beet, and tomato |
|
| Inatreq™ Active® | Corteva | Fenpicoxamid from fermentation broths of the Streptomyces sp. 517–02 | Septoria | Wheat |
|
| Cerall, Cedomon® | Koppert | Pseudomonas chlororaphis MA342 | Fusarium spp., Septoria, wheat bunt and Drechslera teres | Wheat, barley, triticale, and rye |
|
Table 2.
Beneficial bacteria which demonstrated protection against main wheat pathogens. n/a: not applicable.
Table 2.
Beneficial bacteria which demonstrated protection against main wheat pathogens. n/a: not applicable.
| Strains | Origin | Pathogen | Biostimulation | Mode of Action for Biocontrol | Methodology | References |
|---|---|---|---|---|---|---|
| Bacillus subtilis ATCC 10783; B. cereus ATCC 11778; B. licheniformis NRRLB-510; B. pumilus ATCC 7061; Brevibacillus laterosporus BLA170; Paenibacillus polymyxa NA | Soil | Zymoseptoria tritici, Pyrenophora tritici-repentis, Cochliobolus sativus, Alternaria triticimaculans | n/a | n/a | In vitro and greenhouse (leaves) | [121] |
| B. subtilis BBG125, BBG131, Bs2504 | ProBioGEM, Centre Wallon de Biologie Industrielle | Z. tritici | n/a | Lipopeptides (LPs): mycosubtilin, surfactin, fengycin | In vitro and greenhouse (leaves) | [122] |
| B. velezensis RC 218 | Wheat anthers | Fusarium graminearum | n/a | Ericin lantiobiotic Plant phytohormone modulation (jasmonic and salicylic acid) ISR cell wall thickening preventing cell plasmolysis and collapse | Field and greenhouse (spikes) | [123,124,125,126] |
| B. velezensis LM2303 | Wild yak | F. graminearum | n/a | n/a | In silico and field (spikes) | [127] |
| B. subtilis IB | Soil | F. graminearum | n/a | Fengycin | In vitro | [128] |
| B. megaterium BM1 and B. subtilis BS43, BSM0, BSM2 | Wheat spikes | F. graminearum | n/a | Degradation of DON Metabolites | In vitro and field (spikes) | [129] |
| B. subtilis BaSu1/BaSu3, B. amyloliquefaciens BaAm and Chaetomium globosum CG1/CG2, Phoma glomerata PG1, Aureobasidium proteae AP5 and Sarocladium kiliense SK1/SK2 | Wheat endosphere | F. graminearum and F. culmorum | n/a | Antibiosis | In vitro and greenhouse (detached wheat spikelets) | [22] |
| B. amyloliquefaciens TrigoCor | Wheat rhizosphere | F. graminearum | n/a | Iturin | Field and greenhouse (spikes) | [130] |
| B. amyloliquefaciens S76-3 | Wheat spikes | F. graminearum | n/a | Iturin A and plipastatin | In vitro | [131] |
| B. amyloliquefaciens Y1 | Soil | F. graminearum | n/a | Metabolites Cyclo D-PRO-L- VAL | In vitro | [132] |
| B. amyloliquefaciens B8 and B3 | Soil | F. graminearum and culmorum | Yes | Phytohormones | In vitro and greenhouse | [133] |
| B. amyloliquefaciens BLB369, B. subtilis BLB277, Pae. polymyxa BLB267 | Soil | F. graminearum | Yes | Supernatant (iturin and surfactin, fengycin, fusaricidin and polymyxin) | In vitro and greenhouse | [134] |
| B. megaterium MKB135 Pseudomonas fluorescens MKB21 and MKB91 | Cereal rhizospheres, leaves, grain and weeds | Z. tritici | Yes | Cell free surpernantant and VOC | Field and greenhouse (leaves) | [135] |
| B. subtilis strains 53 and 71, P. fluorescens biov1 strain 32 and Streptomyces sp. strain 3 | Wheat kernels | F. graminearum | Yes | Volatiles metabolites | In vitro and greenhouse (spikes) | [136] |
| Antibiotics tubercidin, phosphlactomycin and candicidin, 2,4-diacetylphloroglucinol, phenasin, fengymcine, bacillomycin | ||||||
| Phytohormone regulation | ||||||
| B. subtilis RC 218, S. sp. BRC87B. and Brevibacillus sp. BRC263 | Wheat anthers | F. graminearum | Yes | Space and nutrients competition | Field, greenhouse and in vitro | [125,126,137,138,139,140] |
| Metabolites | ||||||
| Streptomyces. sp. DEF09 | Wheat root | F. graminearum | Yes | Metabolites | Field, greenhouse (spikes) and in vitro | [141] |
| IAA | ||||||
| Streptomyces. sp. BN1 | Rice kernels | F. graminearum | n/a | n/a | In vitro and greenhouse (seeds and spikes) | [142] |
| B. cereus | Soil from wheat fields | F. graminearum | Yes | Dose and cultivar dependent | In vitro and greenhouse (seeds) | [143] |
| B. subtilis AS43.3/AS43.4, Cryptococcus sp. OH71.4 and Cryptococcus nodaensis OH182.9 | Wheat anthers | F. graminearum | n/a | n/a | Field, greenhouse (spikes) and in vitro | [144,145] |
| Co-cultures of B. subtilis OH 131.1 and Cryptococcus flavescens OH 182.9 | ARS NRRL | F. graminearum | n/a | Plipastatin and subtilomycin | Greenhouse (spikes) | [146] |
| Lactobacillus brevis JJ2P; Lactobacillus reuteri R2 | Porcine gut, cheese | Z. tritici | n/a | Cell free supernatant (phenyllactic acid and hydroxyphenyllactic acid) | In vitro and greenhouse (leaves) | [147] |
| Lactobacillus plantarum strain 21B | Sourdough breads | F. graminearum | n/a | Antifungal phenyllactic acid and 4-hydroxyphenyllactic acid | In vitro | [148] |
| Paenibacillus polymyxa SGK2 | Wheat rhizosphere | F. graminearum, F. culmorum, F. verticillioides | Yes | Competition for nutrients (iron) | In vitro and greenhouse (seeds) | [149] |
| Pae. polymyxa W1-14-3 and C1-8-b | Rhizosphere | F. graminearum | Yes | Inhibition of fungal germination | In vitro and greenhouse (spikes) | [150] |
| glucanolytic enzyme, cellulase, mannanase xylase, chitinase and protease | ||||||
| supernatant activity (enzymatic or antibiotic activities: polymyxins, benzoic acid, fusaricidin A and antibiotic peptides) | ||||||
| Pae. sp. B2 | INRAE Dijon | Z. tritici | n/a | ISR | Field and greenhouse (leaves) | [151] |
| P. fluorescens LEC1 | Wheat phyllosphere | Z. tritici | n/a | Antibiotics 1- hydroxyphenazine and chlororaphin | In vitro and greenhouse (leaves) | [152] |
| P. fluorescens PFM2 | Wheat phyllosphere | M. graminicola | n/a | Antibiotics 2-4-diacetylphoroglucinol and phenazine-l-carboxylic acid | In vitro | [153] |
| P sp. AS 64.4 | Wheat anthers | F. graminearum | n/a | Nutrients competition (choline metabolizing strain) | Field, greenhouse (spikes) and in vitro | [154] |
| P. putida BK8661 | Wheat phyllosphere | Z. tritici | Yes | HCN, siderophore, antibiotics | In vitro and greenhouse (leaves) | [155] |
| P. aeruginosa LEC1 | Soil | Z. tritici | n/a | Antibiotic (Pyocyanine) and Siderophore (pyoverdine) | In vitro and field (leaves) | [156] |
| P. chlororaphis MA 342 | Craw berry rhizosphere | Septoria nodorum | n/a | n/a | Field (seeds) | [157] |
| P. piscium ZJU60 | Wheat anthers | F. graminearum | n/a | Phenazine-1-carboxamide | Field and greenhouse (spikes) | [158] |
| P. fluorescens LY1-8 | Wheat tissues | F. graminearum | n/a | Extracellular hydrolytic enzymes (protease, chitinase, cellulose, glucanase and siderophore) and antagonistic activity | Field and greenhouse (spikes) | [159] |
| Devosia sp. strain NKJ1 and Nocardioides spp. strains SS3 or SS4 | Wheat field soil | F. graminearum | Yes | Degradation of DON | In vitro | [160] |
Rows highlighted in grey depict the authors hypothesis for the mode of action of beneficial bacteria based on their preliminary studies.
Table 3.
Beneficial bacteria which demonstrated protection against main barley pathogens.
| Strains | Origin | Pathogen | Biostimulation | Mode of Action of Biocontrol | M&M | Source |
|---|---|---|---|---|---|---|
| Pseudomonas fluorescens MKB100 and MKB156 | Cereal rhizosphere | P. teres | n/a | ISR | Field, greenhouse (leaves and drenching) and in vitro | [161] |
| Production of antifungal compounds (2,4-DAPG and HCN) | ||||||
| Pseudomonas chlororaphis MA 342 | Craw berry rhizosphere | D. teres | n/a | n/a | Field and greenhouse (seeds) | [157,162,163] |
| D. graminea | ||||||
| U. hordei | ||||||
| Paenibacillus polymyxa KaI245 | Sorghum rhizosphere | Drechsclera teres f. sp. teres and Rhynchosporium commune | Yes | Cell free supernatant | In vitro and greenhouse (leaves) | [164] |
| Burkholderia sp. strain BE25 | Maize rhizosphere | P. teres | Yes | Induction plant genes defense | In vitro and greenhouse (leaves) | [165,166] |
| Limitation of the fungus on photosynthetic and respiratory parameters |
Rows highlighted in grey depict the authors hypothesis for the mode of action of beneficial bacteria based on their preliminary studies.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dutilloy, E.; Oni, F.E.; Esmaeel, Q.; Clément, C.; Barka, E.A. Plant Beneficial Bacteria as Bioprotectants against Wheat and Barley Diseases. J. Fungi 2022, 8, 632. https://doi.org/10.3390/jof8060632
AMA Style
Dutilloy E, Oni FE, Esmaeel Q, Clément C, Barka EA. Plant Beneficial Bacteria as Bioprotectants against Wheat and Barley Diseases. Journal of Fungi. 2022; 8(6):632. https://doi.org/10.3390/jof8060632
Chicago/Turabian StyleDutilloy, Emma, Feyisara Eyiwumi Oni, Qassim Esmaeel, Christophe Clément, and Essaid Ait Barka. 2022. "Plant Beneficial Bacteria as Bioprotectants against Wheat and Barley Diseases" Journal of Fungi 8, no. 6: 632. https://doi.org/10.3390/jof8060632
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.