
Dynamic Genome-Wide Transcription Profiling and Direct Target Genes of CmWC-1 Reveal Hierarchical Light Signal Transduction in Cordyceps militaris



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Fungal Mycelia after Light Exposure for RNA-Seq
2.2. RNA Extraction, RNA Quality Examination, Library Preparation for RNA Sequencing and Library Examination
2.3. Library Clustering and Sequencing
2.4. Differential Expression Analysis
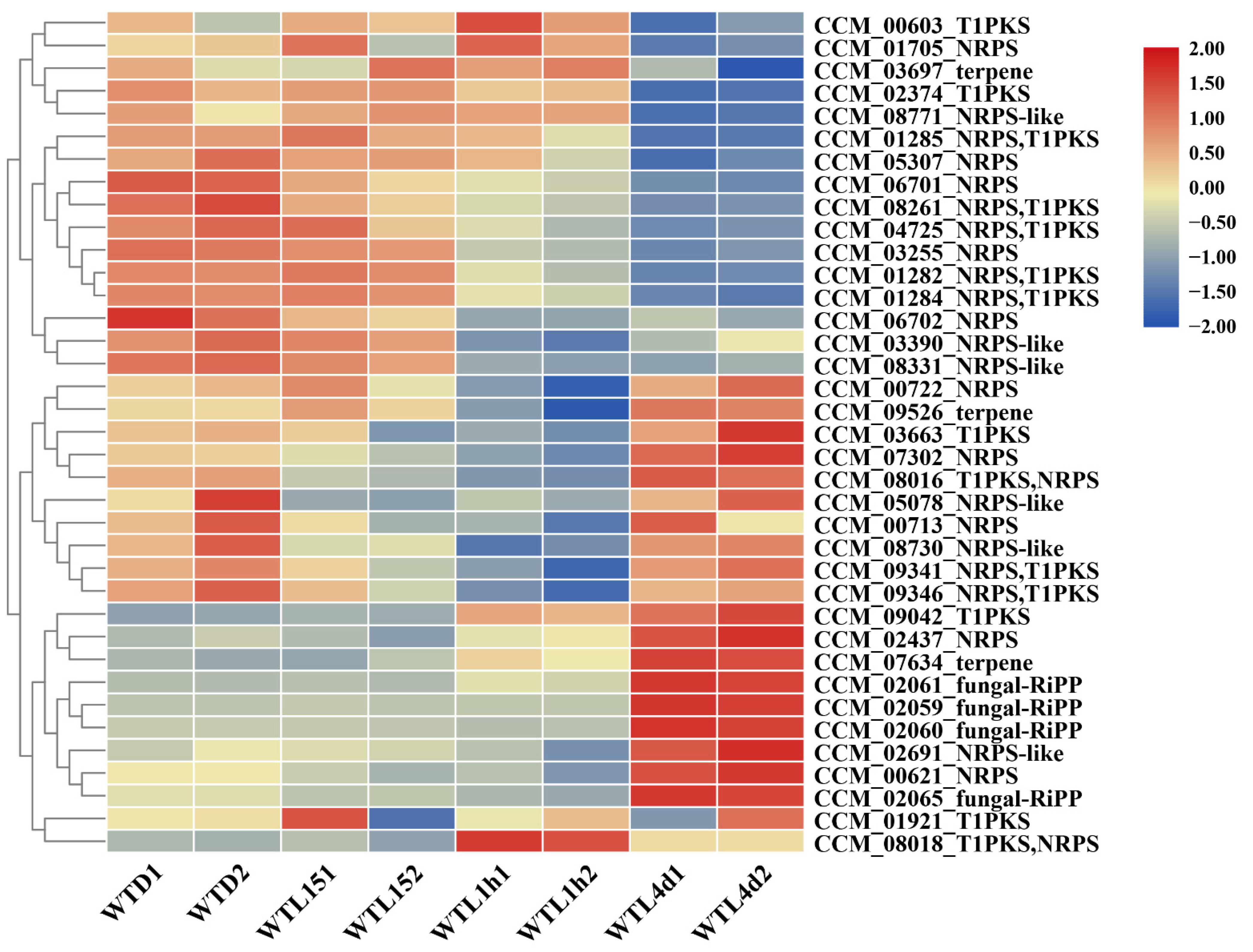
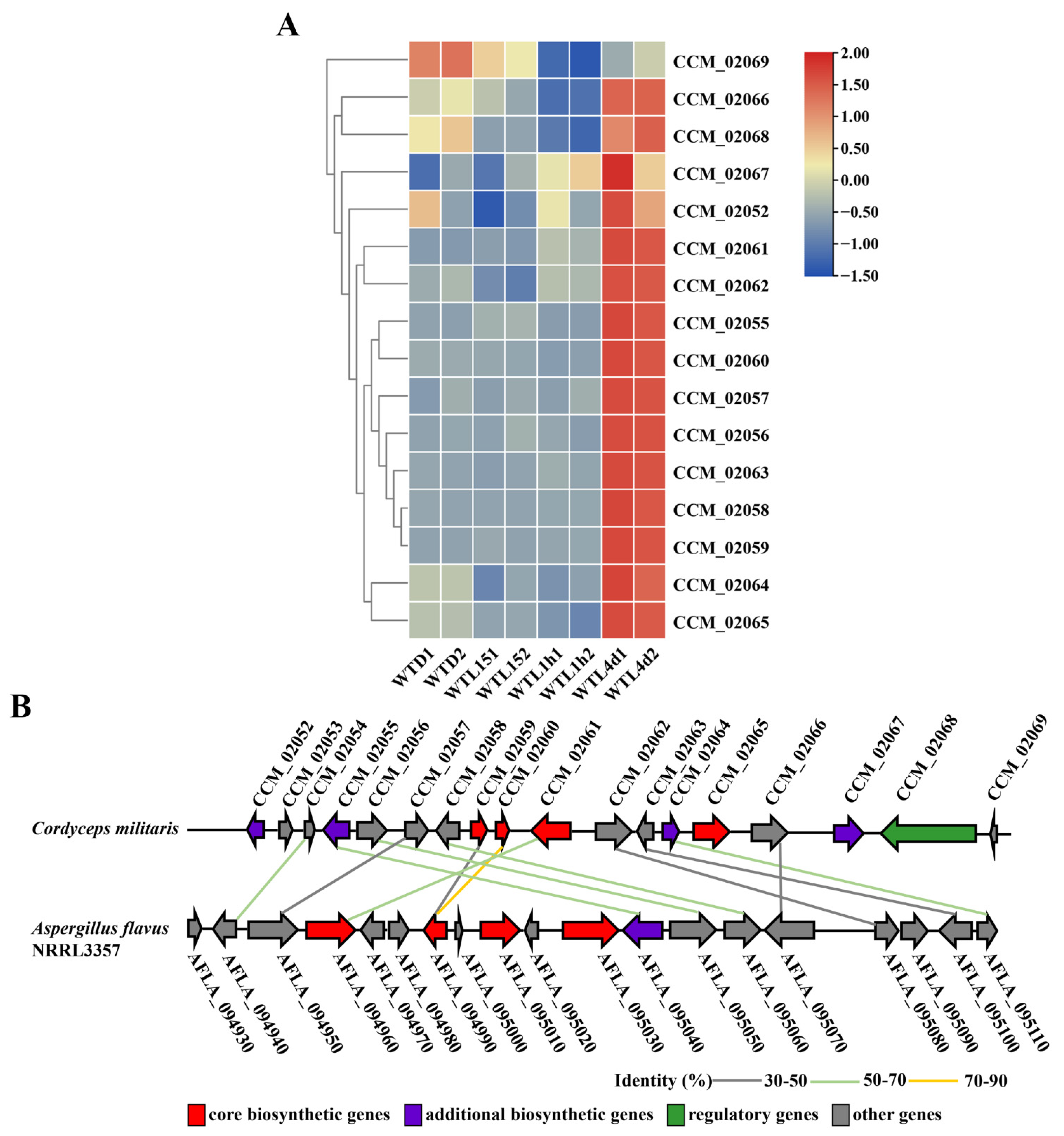
2.5. Secondary Metabolite Biosynthetic Gene Clusters
2.6. Transcription Factor
2.7. Chromatin Immunoprecipitation (ChIP)
2.7.1. Growth Conditions
2.7.2. Cross-Linking
2.7.3. DNA Sonication
2.7.4. Chromatin Immunoprecipitation
2.7.5. DNA Purification for ChIP Samples
2.7.6. ChIP Sequencing, Alignment, Peak Finding
2.7.7. Motif Discovery and Enrichment in CmWC-1 Regulated Genes
2.8. Quantitative Real-Time PCR
3. Results
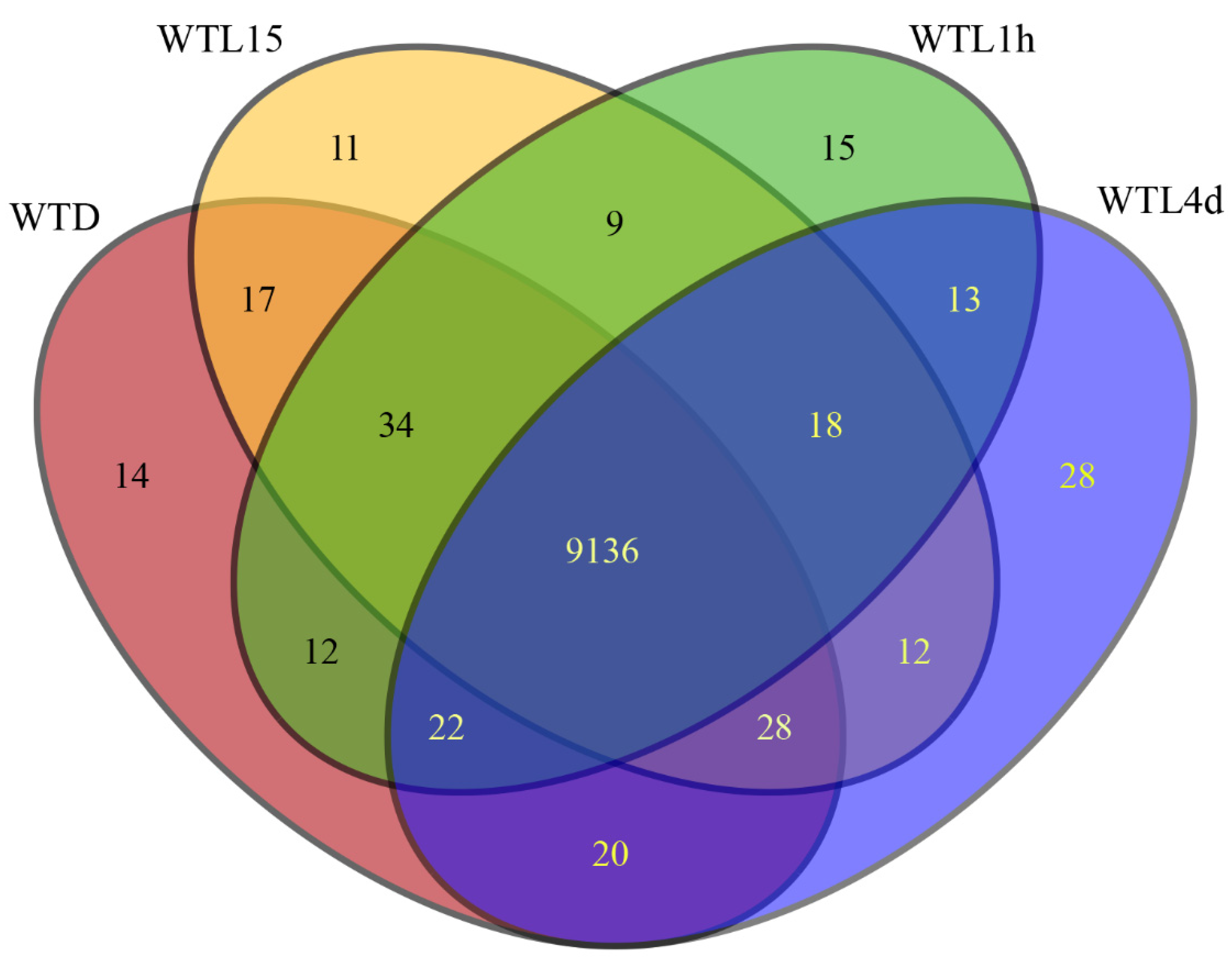
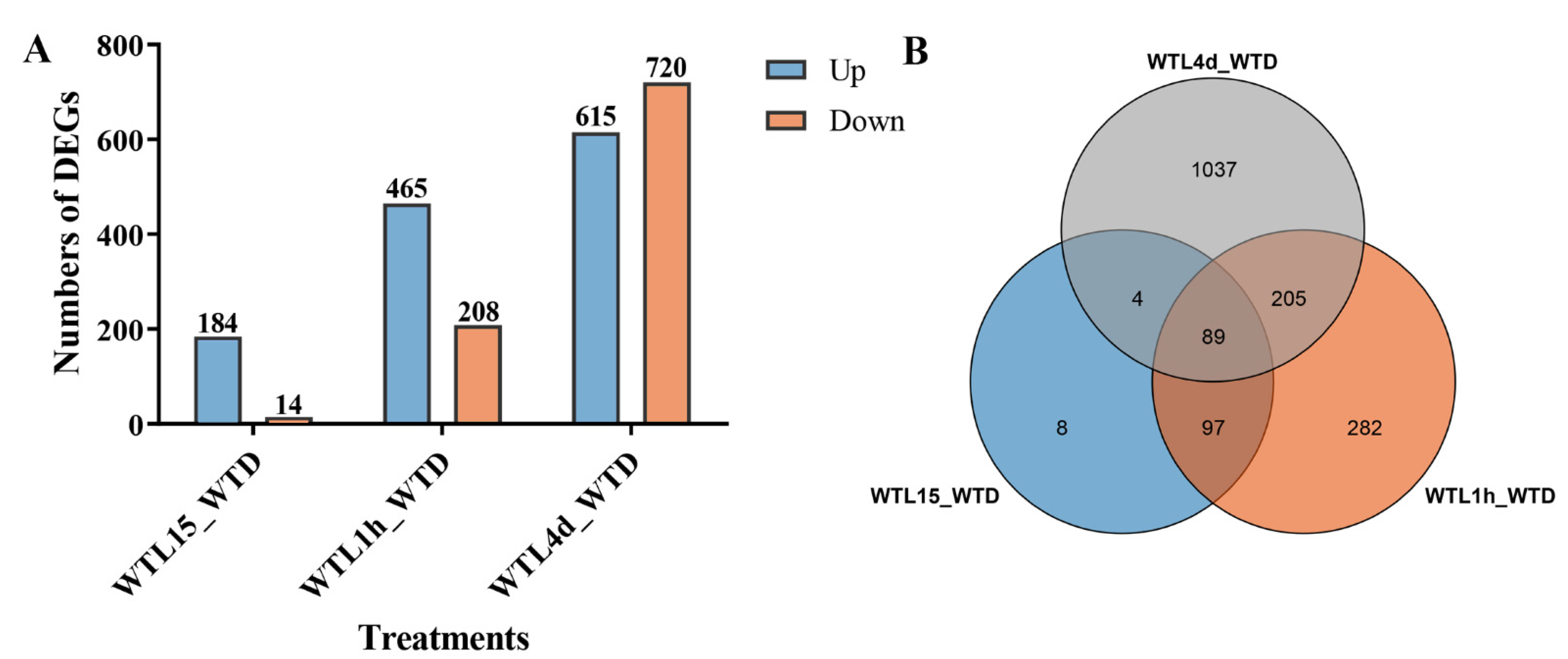
3.1. Gene Transcript Profiles after Light Exposure in Cordyceps militaris
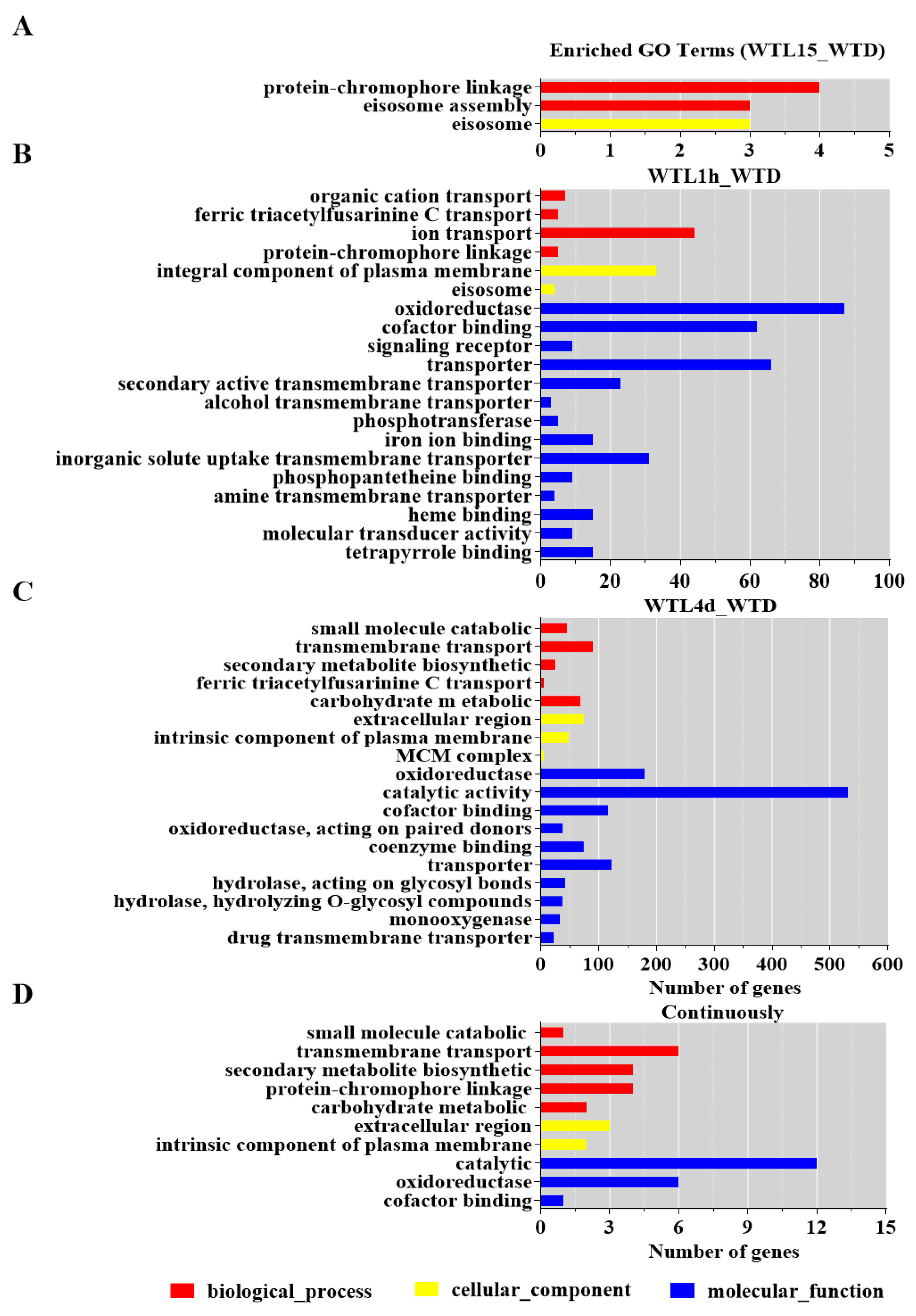
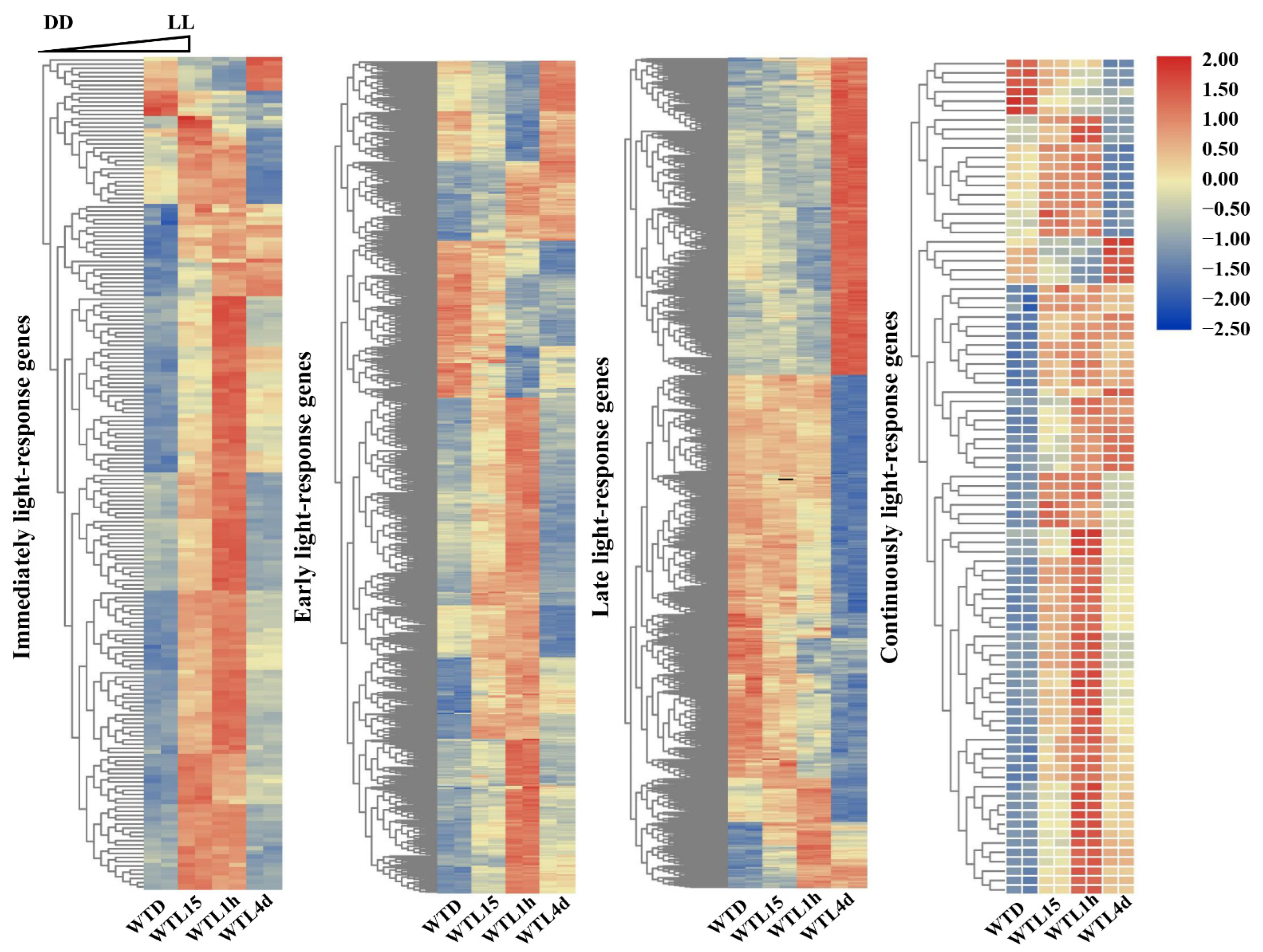
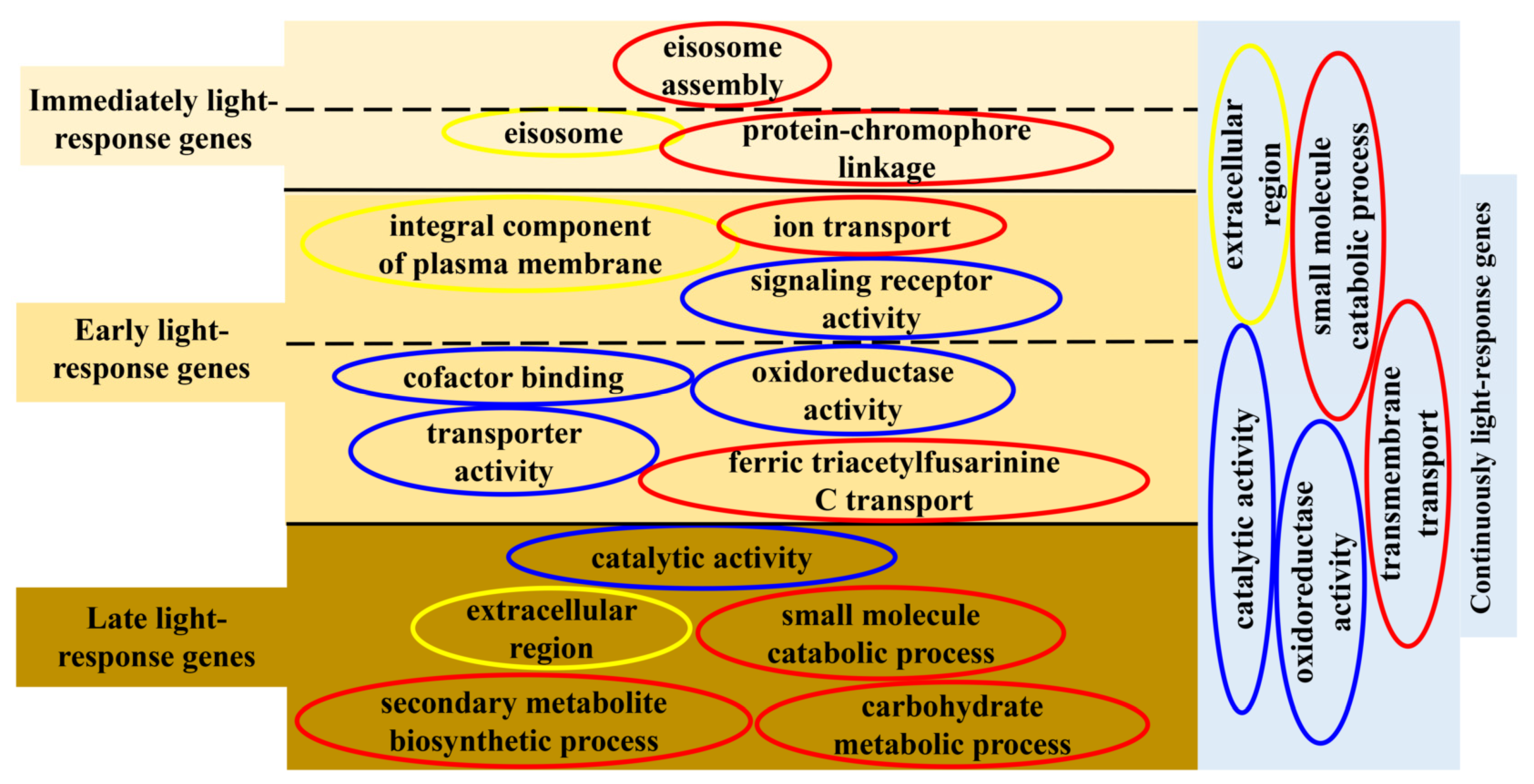
3.2. Hierarchical Analysis of Light-Responsive Genes Identified in Cordyceps militaris
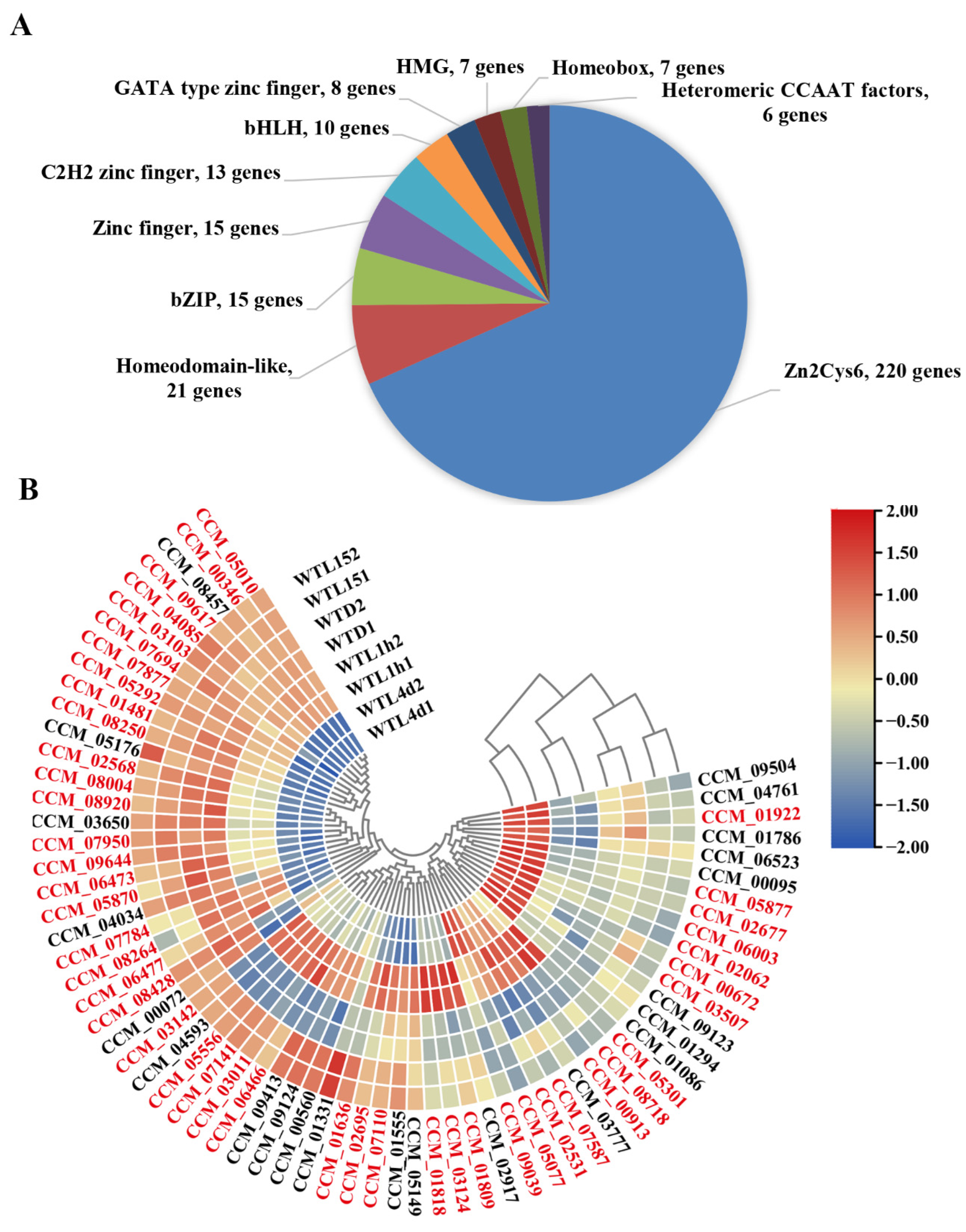
3.3. Transcription Factors Responding to Light in Cordyceps militaris Were Mainly Zn2Cys6-Type and the Majority Responded to Light after Prolonged Exposure
3.4. Metabolite Biosynthesis Cluster Regulated by Light Signal in Cordyceps militaris
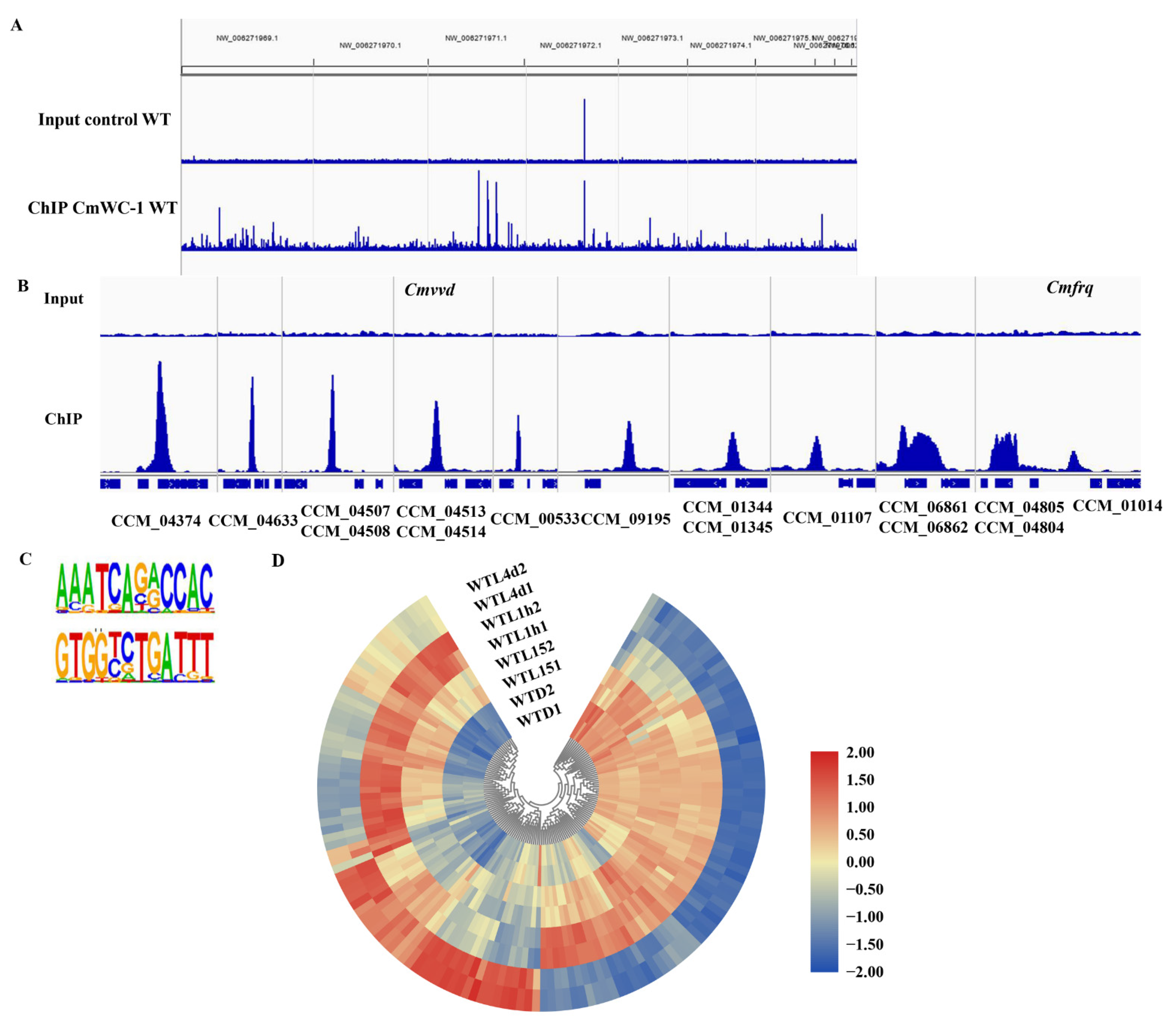
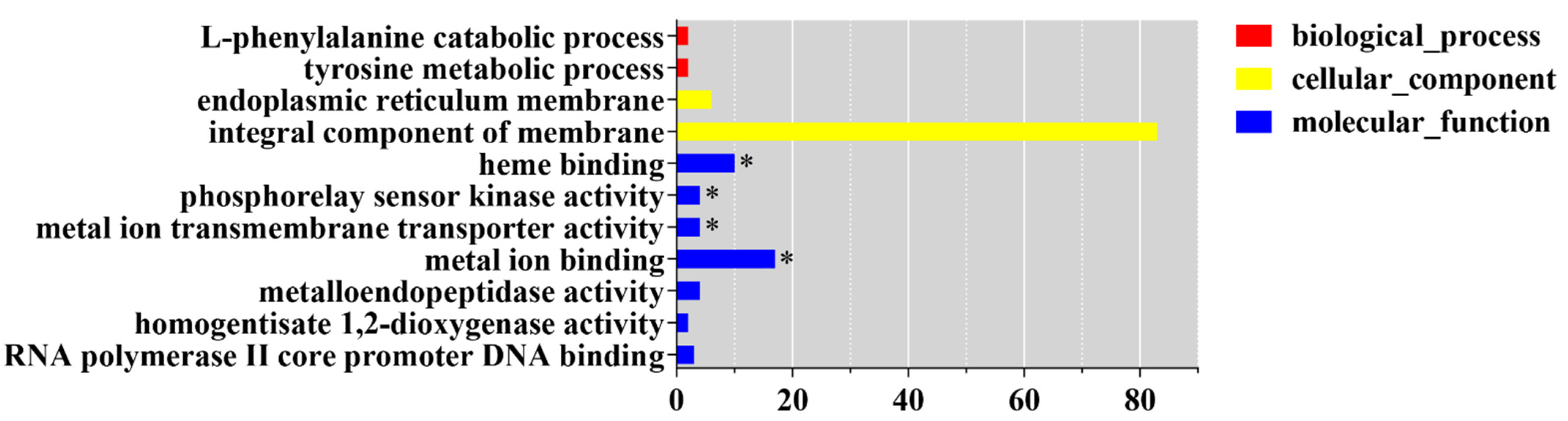
3.5. Light-Responsive Genes Regulated by CmWC-1 in Cordyceps militaris
3.6. ChIP-Seq Analysis Identifies a Core Set of CmWC-1 Target Genes in Response to Light
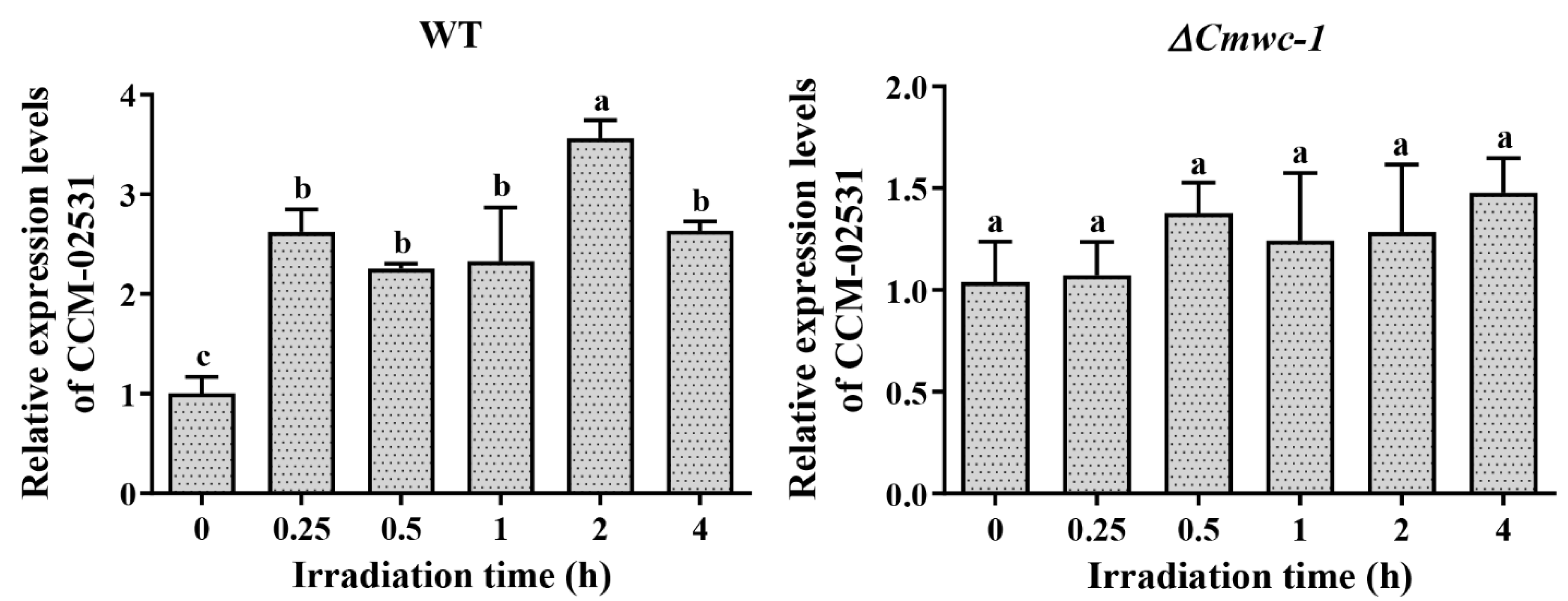
3.7. Light Response of Target Genes of CmWC-1 in Cordyceps militaris
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ambra, R.; Grimaldi, B.; Zamboni, S.; Filetici, P.; Macino, G.; Ballario, P. Photomorphogenesis in the hypogeous fungus Tuber borchii: Isolation and characterization of Tbwc-1, the homologue of the blue-light photoreceptor of Neurospora crassa. Fungal Genet. Biol. 2004, 41, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Kamada, T.; Sano, H.; Nakazawa, T.; Nakahori, K. Regulation of fruiting body photomorphogenesis in Coprinopsiscinerea. Fungal Genet. Biol. 2010, 47, 917–921. [Google Scholar] [CrossRef] [PubMed]
- Ohm, R.A.; Aerts, D.; Wösten, H.A.B.; Lugones, L.G. The blue light receptor complex WC-1/2 of Schizophyllum commune is involved in mushroom formation and protection against phototoxicity. Environ. Microbiol. 2012, 15, 943–955. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Ma, L.; Ying, Z.; Jiang, X.; Lin, Y. Sequence Analysis and Expression of a Blue-light Photoreceptor Gene, Slwc-1 from the Cauliflower Mushroom Sparassis latifolia. Curr. Microbiol. 2017, 74, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.-Y.; Huang, E.-W.; Lin, C.-P. Compositional Differences of the Winter Culinary-Medicinal Mushroom, Flammulina velutipes (Agaricomycetes), under Three Types of Light Conditions. Int. J. Med. Mushrooms 2017, 19, 267–276. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, D.Y.; Park, Y.-J.; Jang, M.-J. Transcriptome analysis of the edible mushroom Lentinula edodes in response to blue light. PLoS ONE 2020, 15, e0230680. [Google Scholar] [CrossRef]
- Tang, L.-H.; Jian, H.-H.; Song, C.-Y.; Bao, D.-P.; Shang, X.-D.; Wu, D.-Q.; Tan, Q.; Zhang, X.-H. Transcriptome analysis of candidate genes and signaling pathways associated with light-induced brown film formation in Lentinula edodes. Appl. Microbiol. Biotechnol. 2013, 97, 4977–4989. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, R.; Qiu, Y.; Wu, H.; Xiang, Q.; Yu, X.; Zhao, K.; Zhang, X.; Chen, Q.; Penttinen, P.; et al. RNA-seq Profiling Showed Divergent Carbohydrate-Active Enzymes (CAZymes) Expression Patterns in Lentinula edodes at Brown Film Formation Stage Under Blue Light Induction. Front. Microbiol. 2020, 11, 1044. [Google Scholar] [CrossRef]
- Terashima, K.; Yuki, K.; Muraguchi, H.; Akiyama, M.; Kamada, T. The dst1 Gene Involved in Mushroom Photomorphogenesis of Coprinus cinereus Encodes a Putative Photoreceptor for Blue Light. Genetics 2005, 171, 101–108. [Google Scholar] [CrossRef]
- Kuratani, M.; Tanaka, K.; Terashima, K.; Muraguchi, H.; Nakazawa, T.; Nakahori, K.; Kamada, T. The dst2 gene essential for photomorphogenesis of Coprinopsis cinerea encodes a protein with a putative FAD-binding-4 domain. Fungal Genet. Biol. 2010, 47, 152–158. [Google Scholar] [CrossRef]
- Sano, H.; Narikiyo, T.; Kaneko, S.; Yamazaki, T.; Shishido, K. Sequence Analysis and Expression of a Blue-Light Photoreceptor Gene, Le.phrA from the Basidiomycetous Mushroom Lentinula edodes. Biosci. Biotechnol. Biochem. 2007, 71, 2206–2213. [Google Scholar] [CrossRef] [PubMed]
- Sano, H.; Kaneko, S.; Sakamoto, Y.; Sato, T.; Shishido, K. The basidiomycetous mushroom Lentinula edodes white collar-2 homolog PHRB, a partner of putative blue-light photoreceptor PHRA, binds to a specific site in the promoter region of the L. edodes tyrosinase gene. Fungal Genet. Biol. 2009, 46, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Tong, X.; Tian, F.; Jia, C.; Li, C.; Li, Y. Transcriptomic profiling sheds light on the blue-light and red-light response of oyster mushroom (Pleurotus ostreatus). AMB Express 2020, 10, 10. [Google Scholar] [CrossRef]
- Zhang, C.; Huang, H.; Deng, W.; Li, T. Genome-Wide Analysis of the Zn(II)2Cys6 Zinc Cluster-Encoding Gene Family in Tolypocladium guangdongense and Its Light-Induced Expression. Genes 2019, 10, 179. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, G.; Deng, W.; Li, T. Distribution, evolution and expression of GATA-TFs provide new insights into their functions in light response and fruiting body development of Tolypocladium guangdongense. PeerJ 2020, 8, e9784. [Google Scholar] [CrossRef]
- Degli Innocenti, F.; A Chambers, J.; E Russo, V. Conidia induce the formation of protoperithecia in Neurospora crassa: Further characterization of white collar mutants. J. Bacteriol. 1984, 159, 808–810. [Google Scholar] [CrossRef] [PubMed]
- Harding, R.W.; Melles, S. Genetic Analysis of Phototropism of Neurospora crassa Perithecial Beaks Using White Collar and Albino Mutants. Plant Physiol. 1983, 72, 996–1000. [Google Scholar] [CrossRef]
- Harding, R.W.; Turner, R.V. Photoregulation of the Carotenoid Biosynthetic Pathway in Albino and White Collar Mutants of Neurospora crassa. Plant Physiol. 1981, 68, 745–749. [Google Scholar] [CrossRef]
- Sargent, M.L.; Briggs, W.R. The Effects of Light on a Circadian Rhythm of Conidiation in Neurospora. Plant Physiol. 1967, 42, 1504–1510. [Google Scholar] [CrossRef]
- Baker, C.L.; Loros, J.J.; Dunlap, J.C. The circadian clock of Neurospora crassa. FEMS Microbiol. Rev. 2012, 36, 95–110. [Google Scholar] [CrossRef]
- Schwerdtfeger, C. VIVID is a flavoprotein and serves as a fungal blue light photoreceptor for photoadaptation. EMBO J. 2003, 22, 4846–4855. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, A.; Fuller, K.K.; Dunlap, J.C.; Loros, J.J. Seeing the world differently: Variability in the photosensory mechanisms of two model fungi. Environ. Microbiol. 2016, 18, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-H.; Ringelberg, C.S.; Gross, R.H.; Dunlap, J.C.; Loros, J.J. Genome-wide analysis of light-inducible responses reveals hierarchical light signalling in Neurospora. EMBO J. 2009, 28, 1029–1042. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Yang, F.; Smith, K.M.; Peterson, M.; Dekhang, R.; Zhang, Y.; Zucker, J.; Bredeweg, E.L.; Mallappa, C.; Zhou, X.; et al. Genome-Wide Characterization of Light-Regulated Genes in Neurospora crass. G3 Genes Genomes Genet. 2014, 4, 1731–1745. [Google Scholar] [CrossRef]
- Carrillo, A.J.; Schacht, P.; Cabrera, I.E.; Blahut, J.; Prudhomme, L.; Dietrich, S.; Bekman, T.; Mei, J.; Carrera, C.; Chen, V.; et al. Functional Profiling of Transcription Factor Genes in Neurospora crassa. G3 Genes Genomes Genet. 2017, 7, 2945–2956. [Google Scholar] [CrossRef]
- Smith, K.M.; Sancar, G.; Dekhang, R.; Sullivan, C.M.; Li, S.; Tag, A.G.; Sancar, C.; Bredeweg, E.L.; Priest, H.D.; McCormick, R.F.; et al. Transcription Factors in Light and Circadian Clock Signaling Networks Revealed by Genomewide Mapping of Direct Targets for Neurospora White Collar Complex. Eukaryot. Cell 2010, 9, 1549–1556. [Google Scholar] [CrossRef]
- Mooney, J.L.; Yager, L.N. Light is required for conidiation in Aspergillus nidulans. Genes Dev. 1990, 4, 1473–1482. [Google Scholar] [CrossRef]
- Bayram, O.; Braus, G.H.; Fischer, R.; Rodriguez-Romero, J. Spotlight on Aspergillus nidulans photosensory systems. Fungal Genet. Biol. 2010, 47, 900–908. [Google Scholar] [CrossRef]
- Bayram, O.; Feussner, K.; Dumkow, M.; Herrfurth, C.; Feussner, I.; Braus, G.H. Changes of global gene expression and secondary metabolite accumulation during light-dependent Aspergillus nidulans development. Fungal Genet. Biol. 2016, 87, 30–53. [Google Scholar] [CrossRef]
- Yue, K.; Ye, M.; Zhou, Z.; Sun, W.; Lin, X. The genus Cordyceps: A chemical and pharmacological review. J. Pharm. Pharmacol. 2012, 65, 474–493. [Google Scholar] [CrossRef]
- Shashidhar, M.; Giridhar, P.; Sankar, K.U.; Manohar, B. Bioactive principles from Cordyceps sinensis: A potent food supplement—A review. J. Funct. Foods 2013, 5, 1013–1030. [Google Scholar] [CrossRef] [PubMed]
- Tuli, H.S.; Sharma, A.K.; Sandhu, S.S.; Kashyap, D. Cordycepin: A bioactive metabolite with therapeutic potential. Life Sci. 2013, 93, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Sun, J.; Lian, T.; Wang, W.; Dong, C.-H. Process Optimization for Extraction of Carotenoids from Medicinal Caterpillar Fungus, Cordyceps militaris (Ascomycetes). Int. J. Med. Mushrooms 2014, 16, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Nakaya, S.; Mizuno, S.; Ishigami, H.; Yamakawa, Y.; Kawagishi, H.; Ushimaru, T. New Rapid Screening Method for Anti-Aging Compounds Using Budding Yeast and Identification of Beauveriolide I as a Potent Active Compound. Biosci. Biotechnol. Biochem. 2012, 76, 1226–1228. [Google Scholar] [CrossRef]
- Wang, X.; Gao, Y.-L.; Zhang, M.-L.; Zhang, H.-D.; Huang, J.-Z.; Li, L. Genome mining and biosynthesis of the Acyl-CoA:cholesterol acyltransferase inhibitor beauveriolide I and III in Cordyceps militaris. J. Biotechnol. 2020, 309, 85–91. [Google Scholar] [CrossRef]
- Xia, Y.; Luo, F.; Shang, Y.; Chen, P.; Lu, Y.; Wang, C. Fungal Cordycepin Biosynthesis Is Coupled with the Production of the Safeguard Molecule Pentostatin. Cell Chem. Biol. 2017, 24, 1479–1489.e4. [Google Scholar] [CrossRef]
- Wu, W.-C.; Hsiao, J.-R.; Lian, Y.-Y.; Lin, C.-Y.; Huang, B.-M. The apoptotic effect of cordycepin on human OEC-M1 oral cancer cell line. Cancer Chemother. Pharmacol. 2006, 60, 103–111. [Google Scholar] [CrossRef]
- Noh, E.-M.; Kim, J.-S.; Hur, H.; Park, B.-H.; Song, E.-K.; Han, M.-K.; Kwon, K.-B.; Yoo, W.-H.; Shim, I.-K.; Lee, S.J.; et al. Cordycepin inhibits IL-1 -induced MMP-1 and MMP-3 expression in rheumatoid arthritis synovial fibroblasts. Rheumatology 2008, 48, 45–48. [Google Scholar] [CrossRef]
- Taofiq, O.; González-Paramás, A.M.; Martins, A.; Barreiro, M.F.; Ferreira, I.C. Mushrooms extracts and compounds in cosmetics, cosmeceuticals and nutricosmetics—A review. Ind. Crop. Prod. 2016, 90, 38–48. [Google Scholar] [CrossRef]
- Kim, H.G.; Shrestha, B.; Lim, S.Y.; Yoon, D.H.; Chang, W.C.; Shin, D.-J.; Han, S.K.; Park, S.M.; Park, J.H.; Park, H.I.; et al. Cordycepin inhibits lipopolysaccharide-induced inflammation by the suppression of NF-κB through Akt and p38 inhibition in RAW 264.7 macrophage cells. Eur. J. Pharmacol. 2006, 545, 192–199. [Google Scholar] [CrossRef]
- Zhou, X.; Cai, G.; He, Y.; Tong, G. Separation of cordycepin from Cordyceps militaris fermentation supernatant using preparative HPLC and evaluation of its antibacterial activity as an NAD+-dependent DNA ligase inhibitor. Exp. Ther. Med. 2016, 12, 1812–1816. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Dong, C. Photo morphogenesis and photo response of the blue-light receptor gene Cmwc-1in different strains of Cordyceps militaris. FEMS Microbiol. Lett. 2014, 352, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.Z.; Lei, C.; Ai, X.R.; Wang, Y. Selenium Enrichment on Cordyceps militaris Link and Analysis on Its Main Active Components. Appl. Biochem. Biotechnol. 2012, 166, 1215–1224. [Google Scholar] [CrossRef] [PubMed]
- Lian, T.T.; Dong, C.H.; Yang, T.; Sun, J.D. Effects of blue light on the growth and bioactive compound production of Cordyceps militaris. Mycosystema 2014, 33, 838–846. [Google Scholar]
- Yang, T.; Guo, M.; Yang, H.; Guo, S.; Dong, C. The blue-light receptor CmWC-1 mediates fruit body development and secondary metabolism in Cordyceps militaris. Appl. Microbiol. Biotechnol. 2015, 100, 743–755. [Google Scholar] [CrossRef]
- Wang, F.; Song, X.; Dong, X.; Zhang, J.; Dong, C. DASH-type cryptochromes regulate fruiting body development and secondary metabolism differently than CmWC-1 in the fungus Cordyceps militaris. Appl. Microbiol. Biotechnol. 2017, 101, 4645–4657. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, F.; Yang, Y.; Wang, Y.; Dong, C. CmVVD is involved in fruiting body development and carotenoid production and the transcriptional linkage among three blue-light receptors in edible fungus Cordyceps militaris. Environ. Microbiol. 2019, 22, 466–482. [Google Scholar] [CrossRef]
- Zheng, P.; Xia, Y.; Xiao, G.; Xiong, C.; Hu, X.; Zhang, S.; Zheng, H.; Huang, Y.; Zhou, Y.; Wang, S.; et al. Genome sequence of the insect pathogenic fungus Cordyceps militaris, a valued traditional chinese medicine. Genome Biol. 2011, 12, R116. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Supek, F.; Škunca, N.; Repar, J.; Vlahoviček, K.; Šmuc, T. Translational Selection Is Ubiquitous in Prokaryotes. PLoS Genet. 2010, 6, e1001004. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Jang, S.; Kim, S.; Kong, S.; Choi, J.; Ahn, K.; Kim, J.; Lee, S.; Park, B.; Jung, K.; et al. FTFD: An informatics pipeline supporting phylogenomic analysis of fungal transcription factors. Bioinformatics 2008, 24, 1024–1025. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆C(T) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Umemura, M.; Nagano, N.; Koike, H.; Kawano, J.; Ishii, T.; Miyamura, Y.; Kikuchi, M.; Tamano, K.; Yu, J.; Shin-Ya, K.; et al. Characterization of the biosynthetic gene cluster for the ribosomally synthesized cyclic peptide ustiloxin B in Aspergillus flavus. Fungal Genet. Biol. 2014, 68, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Heinz, S.; Benner, C.; Spann, N.; Bertolino, E.; Lin, Y.C.; Laslo, P.; Cheng, J.X.; Murre, C.; Singh, H.; Glass, C.K. Simple Combinations of Lineage-Determining Transcription Factors Prime cis-Regulatory Elements Required for Macrophage and B Cell Identities. Mol. Cell 2010, 38, 576–589. [Google Scholar] [CrossRef]
- Li, Y.; Sun, T.; Guo, D.; Gao, J.; Zhang, J.; Cai, F.; Fischer, R.; Shen, Q.; Yu, Z. Comprehensive analysis of the regulatory network of blue-light-regulated conidiation and hydrophobin production in Trichoderma guizhouense. Environ. Microbiol. 2021, 23, 6241–6256. [Google Scholar] [CrossRef]
- Foderaro, J.E.; Douglas, L.M.; Konopka, J.B. MCC/Eisosomes Regulate Cell Wall Synthesis and Stress Responses in Fungi. J. Fungi 2017, 3, 61. [Google Scholar] [CrossRef]
- Zhang, L.-B.; Tang, L.; Ying, S.-H.; Feng, M.-G. Two eisosome proteins play opposite roles in autophagic control and sustain cell integrity, function and pathogenicity in Beauveria bassiana. Environ. Microbiol. 2017, 19, 2037–2052. [Google Scholar] [CrossRef]
- Zhang, L.-B.; Tang, L.; Guan, Y.; Feng, M.-G. Subcellular localization of Sur7 and its pleiotropic effect on cell wall integrity, multiple stress responses, and virulence of Beauveria bassiana. Appl. Microbiol. Biotechnol. 2020, 104, 6669–6678. [Google Scholar] [CrossRef]
- UniProt-GOA. Gene Ontology Annotation Based on UniProtKB/Swiss-Prot Keyword Mapping, Accompanied by Conservative Changes to GO Terms Applied by UniProt. 2012. Available online: https://www.yeastgenome.org/reference/S000250402 (accessed on 3 January 2022).
- Nakamura, K.; Izumiyama, N.; Ohtsubo, K.; Koiso, Y.; Iwasaki, S. Apoptosis induced in the liver, kidney and urinary bladder of mice by the fungal toxin produced by Ustilaginoidea virens. JSM Mycotoxins 1993, 1993, 25–30. [Google Scholar] [CrossRef]
- Hu, Z.; Dang, Y.; Liu, C.; Zhou, L.; Liu, H. Acute exposure to ustiloxin A affects growth and development of early life zebrafish, Danio rerio. Chemosphere 2019, 226, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Koiso, Y.; Li, Y.; Iwasaki, S.; Hanaka, K.; Kobayashi, T.; Sonoda, R.; Fujita, Y.; Yaegashi, H.; Sato, Z. Ustiloxins, antimitotic cydic peptides from false smut balls on rice panicles caused by Ustilaginoidea virens. J. Antibiot. 1994, 47, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Ludueña, R.F.; Roach, M.C.; Prasad, V.; Banerjee, M.; Koiso, Y.; Li, Y.; Iwasaki, S. Interaction of ustiloxin a with bovine brain tubulin. Biochem. Pharmacol. 1994, 47, 1593–1599. [Google Scholar] [CrossRef]
- Ranaivoson, F.M.; Gigant, B.; Berritt, S.; Joullié, M.; Knossow, M. Structural plasticity of tubulin assembly probed by vinca-domain ligands. Acta Crystallogr. Sect. D Biol. Crystallogr. 2012, 68, 927–934. [Google Scholar] [CrossRef]
- Chung, D.; Barker, B.M.; Carey, C.C.; Merriman, B.; Werner, E.R.; Lechner, B.E.; Dhingra, S.; Cheng, C.; Xu, W.J.; Blosser, S.J.; et al. ChIP-seq and In Vivo Transcriptome Analyses of the Aspergillus fumigatus SREBP SrbA Reveals a New Regulator of the Fungal Hypoxia Response and Virulence. PLoS Pathog. 2014, 10, e1004576. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Wang, F.; Liu, M.; Fu, M.; Dong, C. Dynamic Genome-Wide Transcription Profiling and Direct Target Genes of CmWC-1 Reveal Hierarchical Light Signal Transduction in Cordyceps militaris. J. Fungi 2022, 8, 624. https://doi.org/10.3390/jof8060624
Zhang J, Wang F, Liu M, Fu M, Dong C. Dynamic Genome-Wide Transcription Profiling and Direct Target Genes of CmWC-1 Reveal Hierarchical Light Signal Transduction in Cordyceps militaris. Journal of Fungi. 2022; 8(6):624. https://doi.org/10.3390/jof8060624
Chicago/Turabian StyleZhang, Jiaojiao, Fen Wang, Mengqian Liu, Mingjia Fu, and Caihong Dong. 2022. "Dynamic Genome-Wide Transcription Profiling and Direct Target Genes of CmWC-1 Reveal Hierarchical Light Signal Transduction in Cordyceps militaris" Journal of Fungi 8, no. 6: 624. https://doi.org/10.3390/jof8060624