
Resistance to Crayfish Plague: Assessing the Response of Native Iberian Populations of the White-Clawed Freshwater Crayfish

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
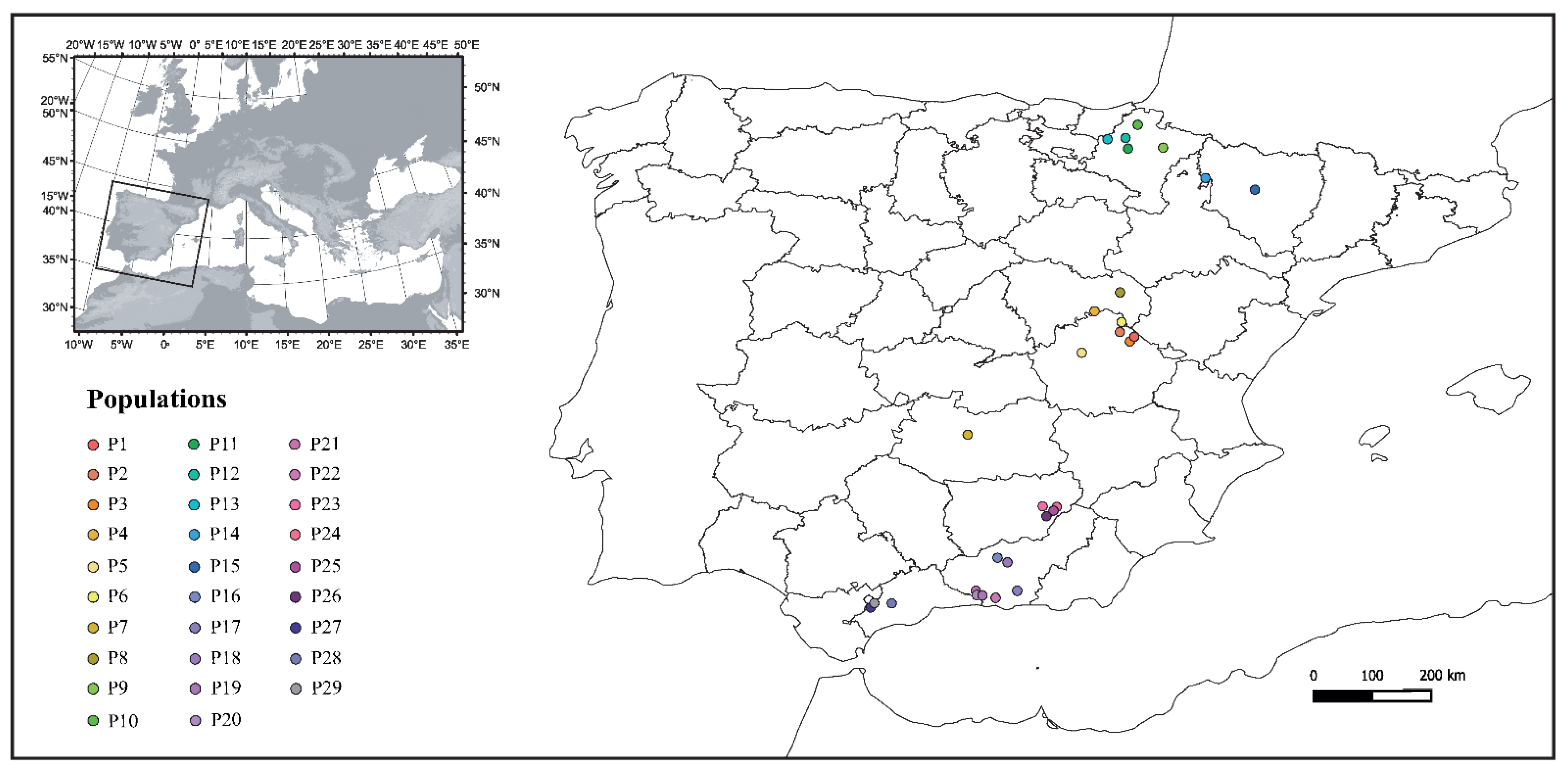
2.1. Crayfish Sampling
2.2. Challenge Experiments with Aphanomyces astaci
2.3. Mean Survival Time and Cumulative Mortality
2.4. Macroscopic and Microscopic Examination
2.5. Molecular Analysis: Aphanomyces astaci-Specific PCR Test and rnnL and rnnS mtDNA Loci Sequencing
3. Results
3.1. Aphanomyces astaci-Challenge Experiments
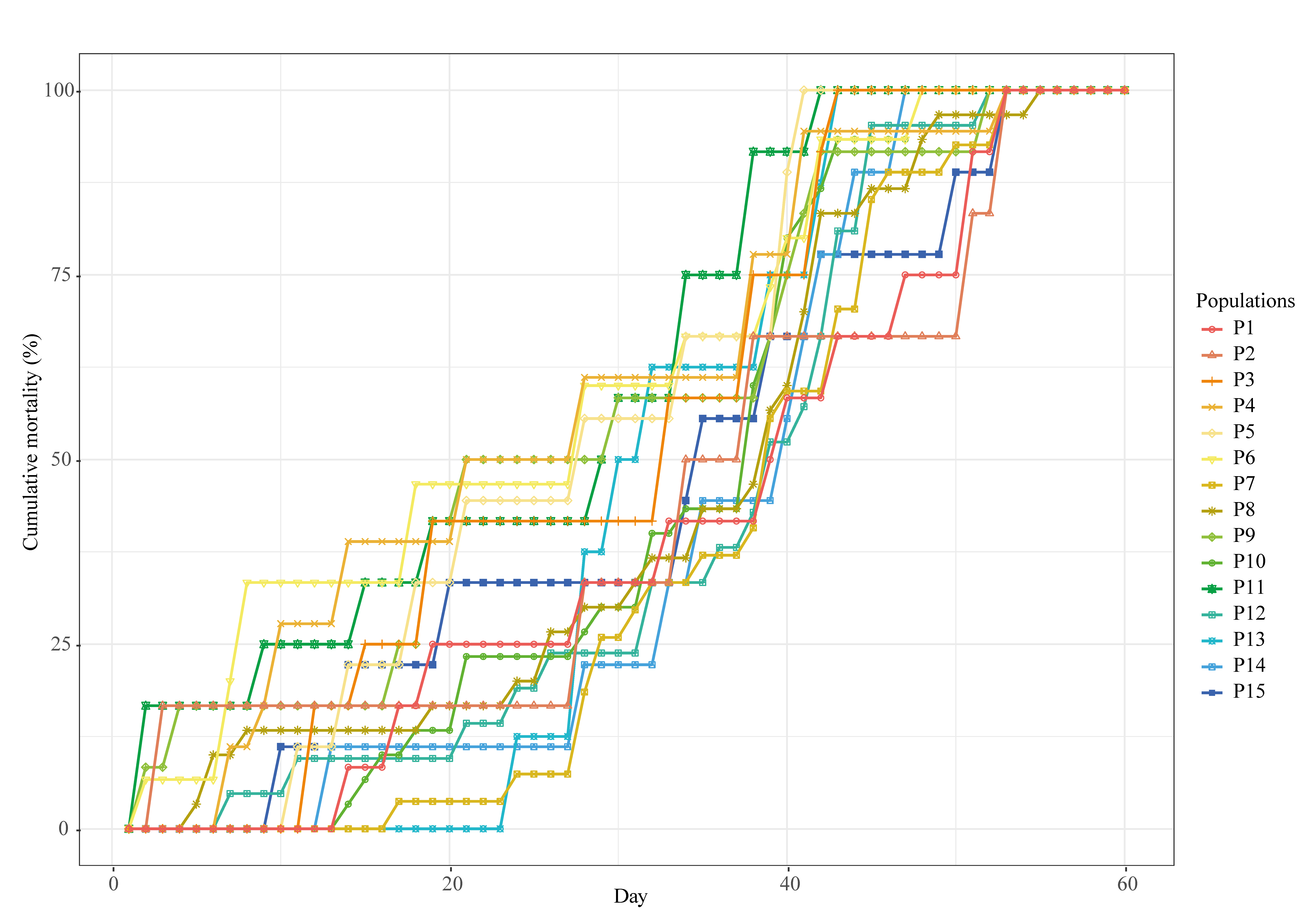
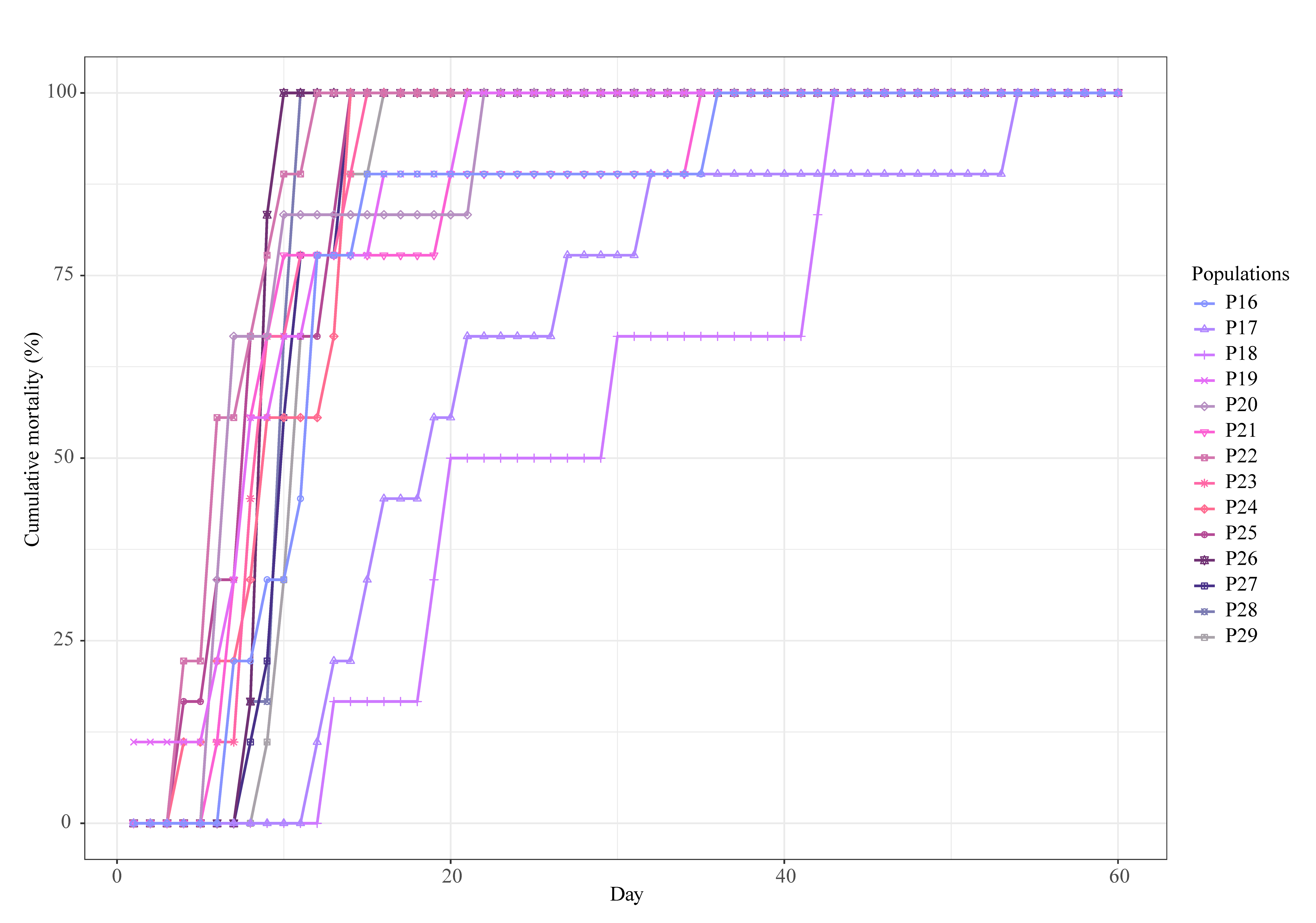
3.2. Mean Survival Time and Cumulative Mortality
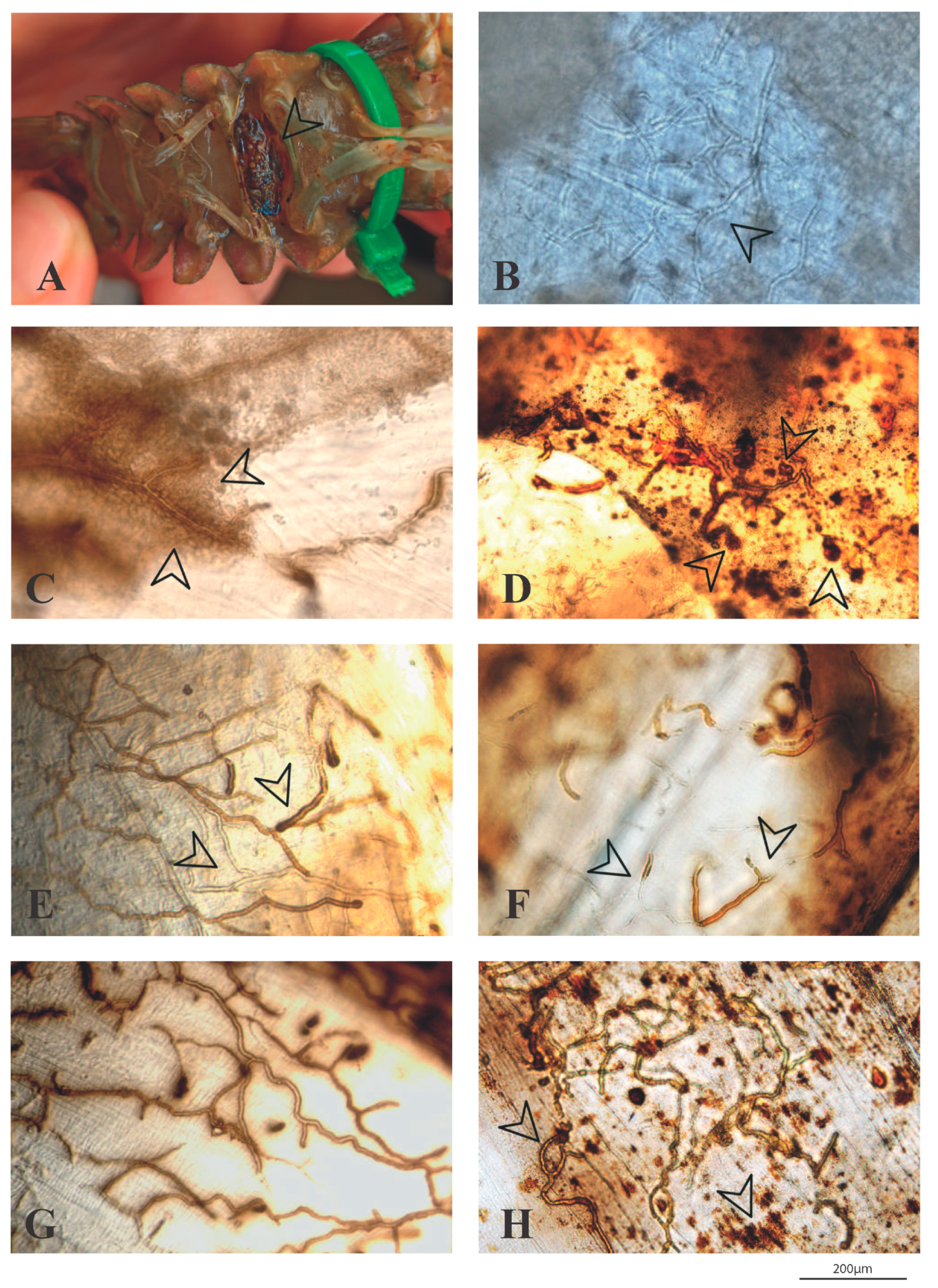
3.3. Macroscopic and Microscopic Examinations
3.4. Molecular Analyses
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethical Statement
References
- Jussila, J.; Edsman, L.; Maguire, I.; Diéguez-Uribeondo, J.; Theissinger, K. Money kills native ecosystems: European crayfish as an example. Front. Ecol. Evol. 2021, 9, 476. [Google Scholar] [CrossRef]
- Unestam, T. On the host range and origin of the crayfish plague fungus. Rep. Inst. Freshw. Res. Drottningholm 1972, 52, 192–198. [Google Scholar]
- Martín-Torrijos, L.; Martínez-Ríos, M.; Casabella-Herrero, G.; Adams, S.B.; Jackson, C.R.; Diéguez-Uribeondo, J. Tracing the origin of the crayfish plague pathogen, Aphanomyces astaci, to the Southeastern United States. Sci. Rep. 2021, 11, 9332. [Google Scholar] [CrossRef] [PubMed]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; The Invasice Species Specialist Group (ISSG): Auckland, New Zeland, 2000; p. 6. [Google Scholar]
- Unestam, T. Defence reactions in and susceptibility of Australian and New Guinean freshwater crayfish to European-crayfish-plague fungus. Aust. J. Exp. Biol. Med. Sci. 1975, 53, 349–359. [Google Scholar] [CrossRef]
- Persson, M.; Söderhäll, K. Pacifastacus leniusculus Dana and its resistance to the parasitic fungus Aphanomyces astaci Schikora. Freshw. Crayfish 1983, 5, 292–298. [Google Scholar]
- Persson, M.; Cerenius, L.; Söderhäll, K. The influence of haemocyte number on the resistance of the freshwater crayfish, Pacifastacus leniusculus Dana, to the parasitic fungus Aphanomyces astaci. J. Fish Dis. 1987, 10, 471–477. [Google Scholar] [CrossRef]
- Diéguez-Uribeondo, J.; Söderhäll, K. Procambarus clarkii Girard as a vector for the crayfish plague fungus, Aphanomyces astaci Schikora. Aquac. Res. 1993, 24, 761–765. [Google Scholar] [CrossRef]
- Söderhäll, K.; Cerenius, L. Crustacean immunity. Annu. Rev. Fish Dis. 1992, 2, 3–23. [Google Scholar] [CrossRef]
- Thörnqvist, P.O.; Söderhäll, K. Psorospermium haeckeli and its interaction with the crayfish defence system. Aquaculture 1993, 117, 205–213. [Google Scholar] [CrossRef]
- Aydin, H.; Kokko, H.; Makkonen, J.; Kortet, R.; Kukkonen, H.; Jussila, J. The signal crayfish is vulnerable to both the as and the PsI-isolates of the crayfish plague. Knowl. Manag. Aquat. Ecosyst. 2014, 413, 3. [Google Scholar] [CrossRef] [Green Version]
- Cerenius, L.; Bangyeekhun, E.; Keyser, P.; Söderhäll, I.; Söderhäll, K. Host prophenoloxidase expression in freshwater crayfish is linked to increased resistance to the crayfish plague fungus, Aphanomyces astaci. Cell. Microbiol. 2003, 5, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Söderhäll, K.; Cerenius, L. Role of the prophenoloxidase-activating system in invertebrate immunity. Curr. Opin. Immunol. 1998, 10, 23–28. [Google Scholar] [CrossRef]
- Yu, X.Q.; Jiang, H.; Wang, Y.; Kanost, M.R. Nonproteolytic serine proteinase homologs are involved in prophenoloxidase activation in the tobacco hornworm, Manduca sexta. Insect Biochem. Mol. Biol. 2003, 33, 197–208. [Google Scholar] [CrossRef]
- Cerenius, L.; Liang, Z.; Duvic, B.; Keyser, P.; Hellman, U.; Palva, E.T.; Iwanaga, S.; Söderhäll, K. Structure and biological activity of a 1,3-beta-D-glucan-binding protein in crustacean blood. J. Biol. Chem. 1994, 269, 29462–29467. [Google Scholar] [CrossRef]
- Ma, C.; Kanost, M.R. A β1,3-glucan recognition protein from an insect, Manduca sexta, agglutinates microorganisms and activates the phenoloxidase cascade. J. Biol. Chem. 2000, 275, 7505–7514. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Wang, R.; Söderhäll, K. A lipopolysaccharide-and β-1,3-glucan-binding protein from hemocytes of the freshwater crayfish Pacifastacus leniusculus: Purification, characterization, and cDNA cloning. J. Biol. Chem. 2000, 275, 1337–1343. [Google Scholar] [CrossRef] [Green Version]
- Kang, D.; Liu, G.; Lundström, A.; Gelius, E.; Steiner, H. A peptidoglycan recognition protein in innate immunity conserved from insects to humans. Proc. Natl. Acad. Sci. USA 1998, 95, 10078–10082. [Google Scholar] [CrossRef] [Green Version]
- Ochiai, M.; Ashida, M. A pattern recognition protein for peptidoglycan: Cloning the cDNA and the gene of the silkworm, Bombyx mori. J. Biol. Chem. 1999, 274, 11854–11858. [Google Scholar] [CrossRef] [Green Version]
- Söderhäll, K.; Ajaxon, R. Effect of quinones and melanin on mycelial growth of Aphanomyces spp. and extracellular protease of Aphanomyces astaci, a parasite on crayfish. J. Invertebr. Pathol. 1982, 39, 105–109. [Google Scholar] [CrossRef]
- Unestam, T. Resistance to the crayfish plague in some American, Japanese and European crayfishes. Rep. Inst. Freshw. Res. Drottningholm 1969, 49, 202–209. [Google Scholar]
- Alderman, D.J.; Polglase, J.L.; Frayling, M. Aphanomyces astaci pathogenicity under laboratory and field conditions. J. Fish Dis. 1987, 10, 385–393. [Google Scholar] [CrossRef]
- Rezinciuc, S.; Sandoval-Sierra, J.V.; Oidtmann, B.; Diéguez-Uribeondo, J. The biology of crayfish plague pathogen Aphanomyces astaci: Current answers to most frequent questions. In Current Answers to Most Frequent Questions in Freshwater Crayfish—A Global Overview, 1st ed.; Kawai, T., Faulkes, Z., Scholtz, G., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 182–204. [Google Scholar]
- Cerenius, L.; Rufelt, S.; Söderhäll, K. Effects of Ampropylfos (RS)-1 aminopropylphosphonic acid) on zoospore formation, repeated zoospore emergence and oospore formation in Aphanomyces spp. Pestic. Sci. 1992, 36, 189–194. [Google Scholar] [CrossRef]
- Huang, T.; Cerenius, L.; Söderhäll, K. Analysis of genetic diversity in the crayfish plague fungus, Aphanomyces astaci, by random amplification of polymorphic DNA. Aquaculture 1994, 126, 1–9. [Google Scholar] [CrossRef]
- Diéguez-Uribeondo, J.; Temiño, C.; Múzquiz, J.L. The crayfish plague fungus (Aphanomyces astaci) in Spain. Bull. Fr. Pêche Piscic. 1997, 347, 753–763. [Google Scholar] [CrossRef] [Green Version]
- Lilley, J.H.; Cerenius, L.; Söderhäll, K. RAPD evidence for the origin of crayfish plague outbreaks in Britain. Aquaculture 1997, 157, 181–185. [Google Scholar] [CrossRef]
- Vennerström, P.; Söderhäll, K.; Cerenius, L. The origin of two crayfish plague (Aphanomyces astaci) epizootics in Finland on noble crayfish, Astacus astacus. Ann. Zool. Fenn. 1998, 35, 43–46. [Google Scholar]
- Diéguez-Uribeondo, J.; Söderhäll, K. RAPD evidence for the origin of an outbreak of crayfish plague in Spain. Freshw. Crayfish 1999, 12, 313–318. [Google Scholar]
- Oidtmann, B.; Cerenius, L.; Schmid, I.; Hoffmann, R.; Söderhäll, K. Crayfish plague epizootics in Germany—Classification of two German isolates of the crayfish plague fungus Aphanomyces astaci by random amplification of polymorphic DNA. Dis. Aquat. Org. 1999, 35, 235–238. [Google Scholar] [CrossRef] [Green Version]
- Alonso, F.; Temiño, C.; Diéguez-Uribeondo, J. Status of the white-clawed crayfish, Austropotamobius pallipes (Lereboullet, 1858), in Spain: Distribution and legislation. Bull. Fr. Pêche Piscic. 2000, 356, 31–53. [Google Scholar] [CrossRef] [Green Version]
- Holdich, D.M.; Reynolds, J.D.; Souty-Grosset, C.; Sibley, P.J. A review of the ever increasing threat to European crayfish from non-indigenous crayfish species. Knowl. Manag. Aquat. Ecosyst. 2009, 394–395, 11. [Google Scholar] [CrossRef] [Green Version]
- Makkonen, J.; Jussila, J.; Kortet, R.; Vainikka, A.; Kokko, H. Differing virulence of Aphanomyces astaci isolates and elevated resistance of noble crayfish Astacus astacus against crayfish plague. Dis. Aquat. Org. 2012, 102, 129–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makkonen, J.; Kokko, H.; Vainikka, A.; Kortet, R.; Jussila, J. Dose-dependent mortality of the noble crayfish (Astacus astacus) to different strains of the crayfish plague (Aphanomyces astaci). J. Invertebr. Pathol. 2014, 115, 86–91. [Google Scholar] [CrossRef]
- Francesconi, C.; Makkonen, J.; Schrimpf, A.; Jussila, J.; Kokko, H.; Theissinger, K. Controlled infection experiment with Aphanomyces astaci provides additional evidence for latent infections and resistance in freshwater crayfish. Front. Ecol. Evol. 2021, 9, 647037. [Google Scholar] [CrossRef]
- Martín-Torrijos, L.; Campos Llach, M.; Pou-Rovira, Q.; Diéguez-Uribeondo, J. Resistance to the crayfish plague, Aphanomyces astaci (Oomycota) in the endangered freshwater crayfish species, Austropotamobius pallipes. PLoS ONE 2017, 12, e0181226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diéguez-Uribeondo, J.; Huang, T.S.; Cerenius, L.; Söderhäll, K. Physiological adaptation of an Aphanomyces astaci strain isolated from the freshwater crayfish Procambarus clarkii. Mycol. Res. 1995, 99, 574–578. [Google Scholar] [CrossRef]
- Martín-Torrijos, L.; Kokko, H.; Makkonen, J.; Jussila, J.; Diéguez-Uribeondo, J. Mapping 15 years of crayfish plague in the Iberian Peninsula: The impact of two invasive species on the endangered native crayfish. PLoS ONE 2019, 14, e0219223. [Google Scholar] [CrossRef] [Green Version]
- Diéguez-Uribeondo, J.; Rueda, A.; Castien, E.; Bascones, J.C. A plan of restauration in Navarra for the native freshwater crayfish species of Spain, Austropotamobius pallipes. Bull. Fr. Pêche Piscic. 1997, 347, 625–637. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Version 2021-3. 2021. Available online: https://www.iucnredlist.org/species/2430/9438817 (accessed on 23 February 2021).
- Rantamäki, J.; Cerenius, L.; Söderhäll, K. Prevention of transmission of the crayfish plague fungus (Aphanomyces astaci) to the freshwater crayfish Astacus astacus by treatment with MgCl2. Aquaculture 1992, 104, 11–18. [Google Scholar] [CrossRef]
- Unestam, T. On the adaptation of Aphanomyces astaci as a parasite. Physiol. Plant. 1969, 22, 221–235. [Google Scholar] [CrossRef]
- Royo, F.; Gironés, O.; de Blas, I.; Muzquiz, J.L. Influencia de la edad en la susceptibilidad a la afanomicosis del cangrejo señal, Pacifastacus leniusculus. Rev. AquaTIC 2016, 13, 1–7. [Google Scholar]
- Alonso, F. Austropotamobius pallipes. In Bases Ecológicas Preliminares para la Conservación de las Especies de Interés Comunitario en España: Invertebrados; Lázaro Varas, E.M., Ed.; Ministerio de Agricultura, Alimentación y Medio Ambiente: Madrid, Spain, 2012; 69p. [Google Scholar]
- Cayuela Delgado, L.; de la Cruz Rot, M. Análisis de Datos Ecológicos en R; Mundi-Prensa: Madrid, Spain, 2022. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 2 February 2022).
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models, R Package Version 0.4.5; R Foundation: Vienna, Austria, 2022. Available online: https://CRAN.R-project.org/package=DHARMa(accessed on 2 February 2022).
- Lesnoff, M.; Lancelot, R. aods3: Analysis of Overdispersed Data Using S3 Methods, aods3 Package Version 0.4-1.1; R Foundation: Vienna, Austria, 2018.
- Barton, K. MuMIn: Multi-Model Inference, R Package Version 1.43.17; R Foundation: Vienna, Austria, 2020. Available online: https://CRAN.R-project.org/package=MuMIn(accessed on 2 February 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 2 February 2022).
- Diéguez-Uribeondo, J.; Förster, H.; Adaskaveg, J.E. Digital image analysis of internal light spots of appressoria of Colletotrichum acutatum. Phytopathology 2003, 93, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oidtmann, B.; Geiger, S.; Steinbauer, P.; Culas, A.; Hoffmann, R.W. Detection of Aphanomyces astaci in North American crayfish by polymerase chain reaction. Dis. Aquat. Org. 2006, 72, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Oidtmann, B.; Schaefers, N.; Cerenius, L.; Söderhäll, K.; Hoffmann, R.W. Detection of genomic DNA of the crayfish plague fungus Aphanomyces astaci (Oomycete) in clinical samples by PCR. Vet. Microbiol. 2004, 100, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Makkonen, J.; Jussila, J.; Panteleit, J.; Keller, N.S.; Schrimpf, A.; Thessinger, K.; Kortet, R.; Martín-Torrijos, L.; Diéguez-Uribeondo, J.; Kokko, H. MtDNA allows the sensitive detection and haplotyping of the crayfish plague disease agent Aphanomyces astaci showing clues about its origin and migration. Parasitology 2018, 145, 1210–1218. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Jussila, J.; Kokko, H.; Kortet, R.; Makkonen, J. Aphanomyces astaci PsI-genotype isolates from different Finnish signal crayfish stocks show variation in their virulence but still kill fast. Knowl. Manag. Aquat. Ecosyst. 2013, 411, 10. [Google Scholar] [CrossRef] [Green Version]
- Woodhams, D.C.; Ardipradja, K.; Alford, R.A.; Marantelli, G.; Reinert, L.K.; Rollins-Smith, L.A. Resistance to chytridiomycosis varies among amphibian species and is correlated with skin peptide defenses. Anim. Conserv. 2007, 10, 409–417. [Google Scholar] [CrossRef]
- Krams, I.; Daukšte, J.; Kivleniece, I.; Krama, T.; Rantala, M.J. Overwinter survival depends on immune defence and body length in male Aquarius najas water striders. Entomol. Exp. Appl. 2011, 140, 45–51. [Google Scholar] [CrossRef]
- Cerenius, L.; Söderhäll, K. The prophenoloxidase-activating system in invertebrates. Immunol. Rev. 2004, 198, 116–126. [Google Scholar] [CrossRef]
- Brown, J.; Ogle, H.; Dale, M. Disease management: General concepts. In Plant Pathogens and Plant Diseases; Brown, J., Ogle, H., Eds.; Rockvale Publications: Armidale, Australia, 1997; pp. 43–358. [Google Scholar]
- Makkonen, J.; Strand, D.; Vrålstad, T.; Kokko, H.; Jussila, J. Quantity and timing of Aphanomyces astaci zoospore release from the noble crayfish suffering from crayfish plague. Vet. Microbiol. 2012, 162, 750–755. [Google Scholar] [CrossRef]
- Svoboda, J.; Kozubíková-Balcarová, E.; Kouba, A.; Buřič, M.; Kozák, P.; Diéguez-Uribeondo, J.; Petrusek, A. Temporal dynamics of spore release of the crayfish plague pathogen from its natural host, American spiny-cheek crayfish (Orconectes limosus), evaluated by transmission experiments. Parasitology 2013, 140, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Råberg, L.; Sim, D.; Read, A.F. Disentangling genetic variation for resistance and tolerance to infectious diseases in animals. Science 2007, 318, 812–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kause, A.; Ødegård, J. The genetic analysis of tolerance to infections: A review. Front. Genet. 2012, 3, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population Name | Location | Experiment | Replicate 1 | Replicate 2 | Replicate 3 | Control | Total Individuals |
|---|---|---|---|---|---|---|---|
| P1 | Cuenca, Castilla-La Mancha | 1 | 4 | 4 | 4 | 4 | 16 |
| P2 | Cuenca, Castilla-La Mancha | 1 | 2 | 2 | 2 | 2 | 8 |
| P3 | Cuenca, Castilla-La Mancha | 1 | 4 | 4 | 4 | 4 | 16 |
| P4 | Cuenca, Castilla-La Mancha | 1 | 6 | 6 | 6 | 6 | 24 |
| P5 | Cuenca, Castilla-La Mancha | 1 | 3 | 3 | 3 | 3 | 12 |
| P6 | Cuenca, Castilla-La Mancha | 1 | 5 | 5 | 5 | 5 | 20 |
| P7 | Guadalajara, Castilla-La Mancha | 1 | 9 | 9 | 9 | 4 | 31 |
| P8 | Ciudad Real, Castilla-La Mancha | 1 | 10 | 10 | 10 | 7 | 37 |
| P9 | Navarra | 1 | 4 | 4 | 4 | 2 | 14 |
| P10 | Navarra | 1 | 10 | 10 | 10 | 3 | 33 |
| P11 | Navarra | 1 | 4 | 4 | 4 | 2 | 14 |
| P12 | Navarra | 1 | 7 | 7 | 7 | 4 | 25 |
| P13 | Navarra | 1 | 3 | 3 | 3 | 2 | 11 |
| P14 | Huesca, Aragón | 1 | 3 | 3 | 3 | 2 | 11 |
| P15 | Huesca, Aragón | 1 | 3 | 3 | 3 | 1 | 10 |
| P16 | Granada, Andalucía | 2 | 3 | 3 | 3 | 3 | 12 |
| P17 | Granada, Andalucía | 2 | 3 | 3 | 3 | 3 | 12 |
| P18 | Granada, Andalucía | 2 | 2 | 2 | 2 | 1 | 7 |
| P19 | Granada, Andalucía | 2 | 3 | 3 | 3 | 3 | 12 |
| P20 | Granada, Andalucía | 2 | 2 | 2 | 2 | 2 | 8 |
| P21 | Granada, Andalucía | 2 | 3 | 3 | 3 | 3 | 12 |
| P22 | Jaén, Andalucía | 2 | 3 | 3 | 3 | 3 | 12 |
| P23 | Jaén, Andalucía | 2 | 3 | 3 | 3 | 3 | 12 |
| P24 | Jaén, Andalucía | 2 | 3 | 3 | 3 | 3 | 12 |
| P25 | Jaén, Andalucía | 2 | 3 | 3 | 3 | 2 | 11 |
| P26 | Jaén, Andalucía | 2 | 3 | 3 | 3 | 1 | 10 |
| P27 | Málaga, Andalucía | 2 | 3 | 3 | 3 | 3 | 12 |
| P28 | Málaga, Andalucía | 2 | 2 | 2 | 2 | 2 | 8 |
| P29 | Málaga, Andalucía | 2 | 2 | 2 | 2 | 3 | 9 |
| Population Name | Experiment | Total Cumulative Mortality | Mean Survival Time (Standard Deviation) (Days) |
|---|---|---|---|
| P1 | 1 | 100% | 36.25 (sd ± 13.95) |
| P2 | 1 | 100% | 34.33 (sd ± 16.46) |
| P3 | 1 | 100% | 28.83 (sd ± 12.44) |
| P4 | 1 | 100% | 25.50 (sd ± 14.75) |
| P5 | 1 | 100% | 27.44 (sd ± 11.85) |
| P6 | 1 | 100% | 24.60 (sd ± 15.76) |
| P7 | 1 | 100% | 38.00 (sd ± 9.05) |
| P8 | 1 | 100% | 33.7 (sd ± 13.46) |
| P9 | 1 | 100% | 27.17 (sd ± 15.91) |
| P10 | 1 | 100% | 33.53 (sd ± 9.92) |
| P11 | 1 | 100% | 24.33 (sd ± 14.38) |
| P12 | 1 | 100% | 35.52 (sd ± 11.68) |
| P13 | 1 | 100% | 33.25 (sd ± 7.15) |
| P14 | 1 | 100% | 35.89 (sd ± 10.40) |
| P15 | 1 | 100% | 33 (sd ± 15.29) |
| Control | 1 | 10.9% | 47.84 (sd ± 19.05) |
| P16 | 2 | 100% | 13.44 (sd ± 8.85) |
| P17 | 2 | 100% | 23.22 (sd ± 13.28) |
| P18 | 2 | 100% | 24.00 (sd ± 11.73) |
| P19 | 2 | 100% | 9.89 (sd ± 5.86) |
| P20 | 2 | 100% | 20.50 (sd ± 9.99) |
| P21 | 2 | 100% | 12.22 (sd ± 9.51) |
| P22 | 2 | 100% | 7.22 (sd ± 2.73) |
| P23 | 2 | 100% | 9.78 (sd ± 2.99) |
| P24 | 2 | 100% | 10.11 (sd ± 3.79) |
| P25 | 2 | 100% | 13.33 (sd ± 3.14) |
| P26 | 2 | 100% | 13.83 (sd ± 0.97) |
| P27 | 2 | 100% | 10.78 (sd ± 2.05) |
| P28 | 2 | 100% | 15.50 (sd ± 1.00) |
| P29 | 2 | 100% | 11.56 (sd ± 2.19) |
| Control | 2 | 7.89% | 51.17 (sd ± 17.60) |
| p-Value | P16 | P17 | P18 | P19 | P20 | P21 | P22 | P23 | P24 | P25 | P26 | P27 | P28 | P29 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P16 | ||||||||||||||
| P17 | >0.05 | |||||||||||||
| P18 | 0.0084433 | >0.05 | ||||||||||||
| P19 | >0.05 | 0.0051506 | 0.0002135 | |||||||||||
| P20 | >0.05 | 0.0180352 | 0.0009389 | >0.05 | ||||||||||
| P21 | >0.05 | 0.0516023 | 0.0025667 | >0.05 | >0.05 | |||||||||
| P22 | >0.05 | 0.0006579 | 0.0000276 | >0.05 | >0.05 | >0.05 | ||||||||
| P23 | >0.05 | 0.0045652 | 0.0001885 | >0.05 | >0.05 | >0.05 | >0.05 | |||||||
| P24 | >0.05 | 0.0208085 | 0.0009735 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | ||||||
| P25 | >0.05 | 0.0379016 | 0.0023556 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | |||||
| P26 | >0.05 | 0.0098631 | 0.0004890 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | ||||
| P27 | >0.05 | 0.0130939 | 0.0005671 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | |||
| P28 | >0.05 | 0.0241095 | 0.0012926 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | ||
| P29 | >0.05 | 0.0280708 | 0.0012905 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 | >0.05 |
| Cumulative Mortality (%) | ||
|---|---|---|
| Week | Experiment 1 | Experiment 2 |
| 1 | 3.14 | 16.04 |
| 2 | 8.52 | 64.15 |
| 3 | 12.1 | 11.32 |
| 4 | 9.87 | 1.89 |
| 5 | 14.8 | 2.83 |
| 6 | 32.29 | 1.89 |
| 7 | 13 | 0.94 |
| 8 | 6.28 | 0.94 |
| 9 | 0 | 0 |
| GLM | AIC Value |
|---|---|
| Data set 1 | 2270.599 |
| Data set 2 | 2261.121 |
| Data set 3 | 2256.364 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Ríos, M.; Lapesa-Lázaro, S.; Larumbe-Arricibita, J.; Alonso-Gutiérrez, F.; Galindo-Parrila, F.J.; Martín-Torrijos, L.; Diéguez-Uribeondo, J. Resistance to Crayfish Plague: Assessing the Response of Native Iberian Populations of the White-Clawed Freshwater Crayfish. J. Fungi 2022, 8, 342. https://doi.org/10.3390/jof8040342
Martínez-Ríos M, Lapesa-Lázaro S, Larumbe-Arricibita J, Alonso-Gutiérrez F, Galindo-Parrila FJ, Martín-Torrijos L, Diéguez-Uribeondo J. Resistance to Crayfish Plague: Assessing the Response of Native Iberian Populations of the White-Clawed Freshwater Crayfish. Journal of Fungi. 2022; 8(4):342. https://doi.org/10.3390/jof8040342
Chicago/Turabian StyleMartínez-Ríos, María, Sara Lapesa-Lázaro, Jokin Larumbe-Arricibita, Fernando Alonso-Gutiérrez, Francisco Javier Galindo-Parrila, Laura Martín-Torrijos, and Javier Diéguez-Uribeondo. 2022. "Resistance to Crayfish Plague: Assessing the Response of Native Iberian Populations of the White-Clawed Freshwater Crayfish" Journal of Fungi 8, no. 4: 342. https://doi.org/10.3390/jof8040342