
In Vitro Characterization of a Nuclear Receptor-like Domain of the Xylanase Regulator 1 from Trichoderma reesei

 and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plasmid Construction
2.2. Protein Expression and Purification
2.3. Circular Dichroism Spectroscopy
2.4. Electrophoretic Mobility Shift Assay
2.5. Blue Native Polyacrylamide Gel Electrophoresis
3. Results
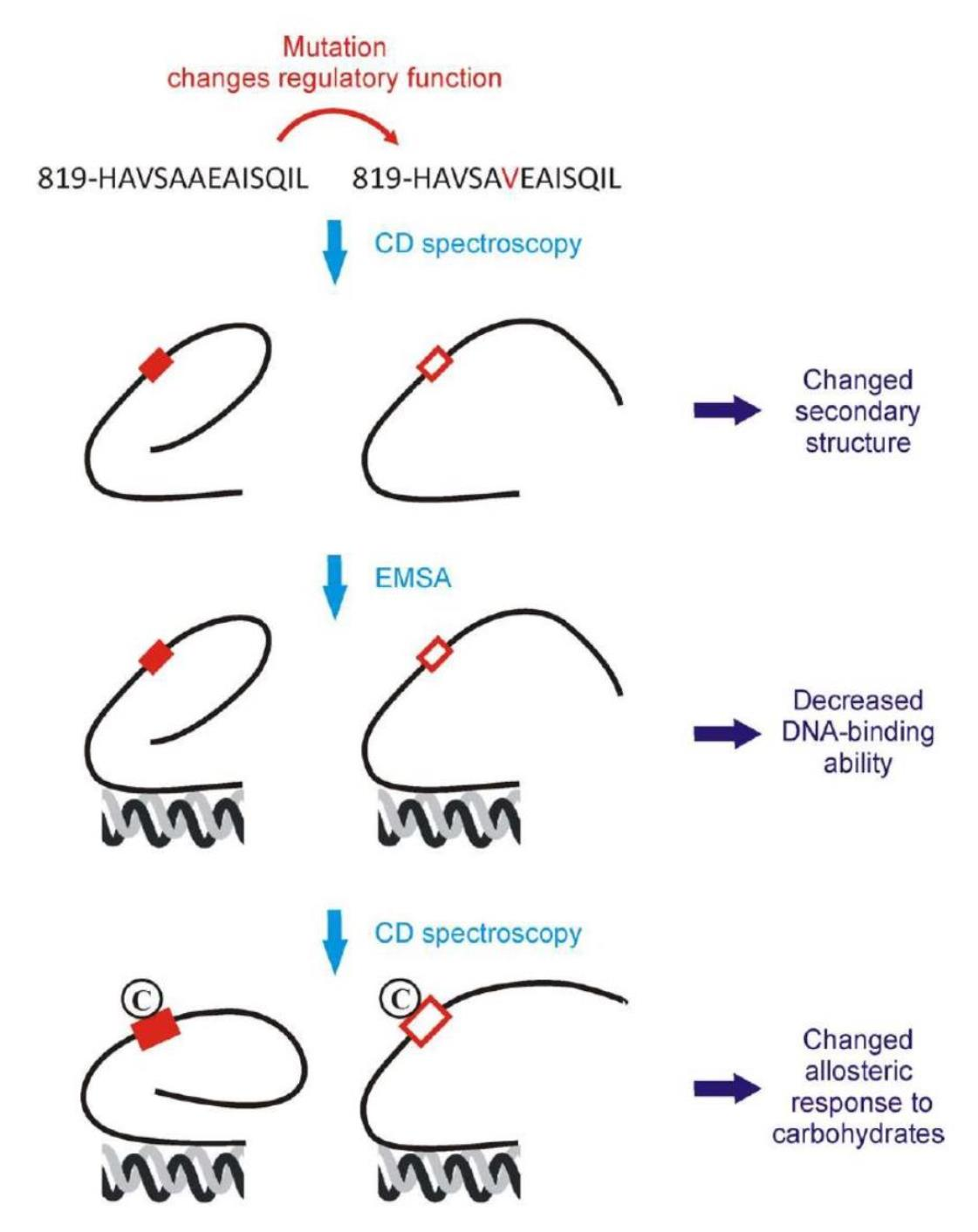
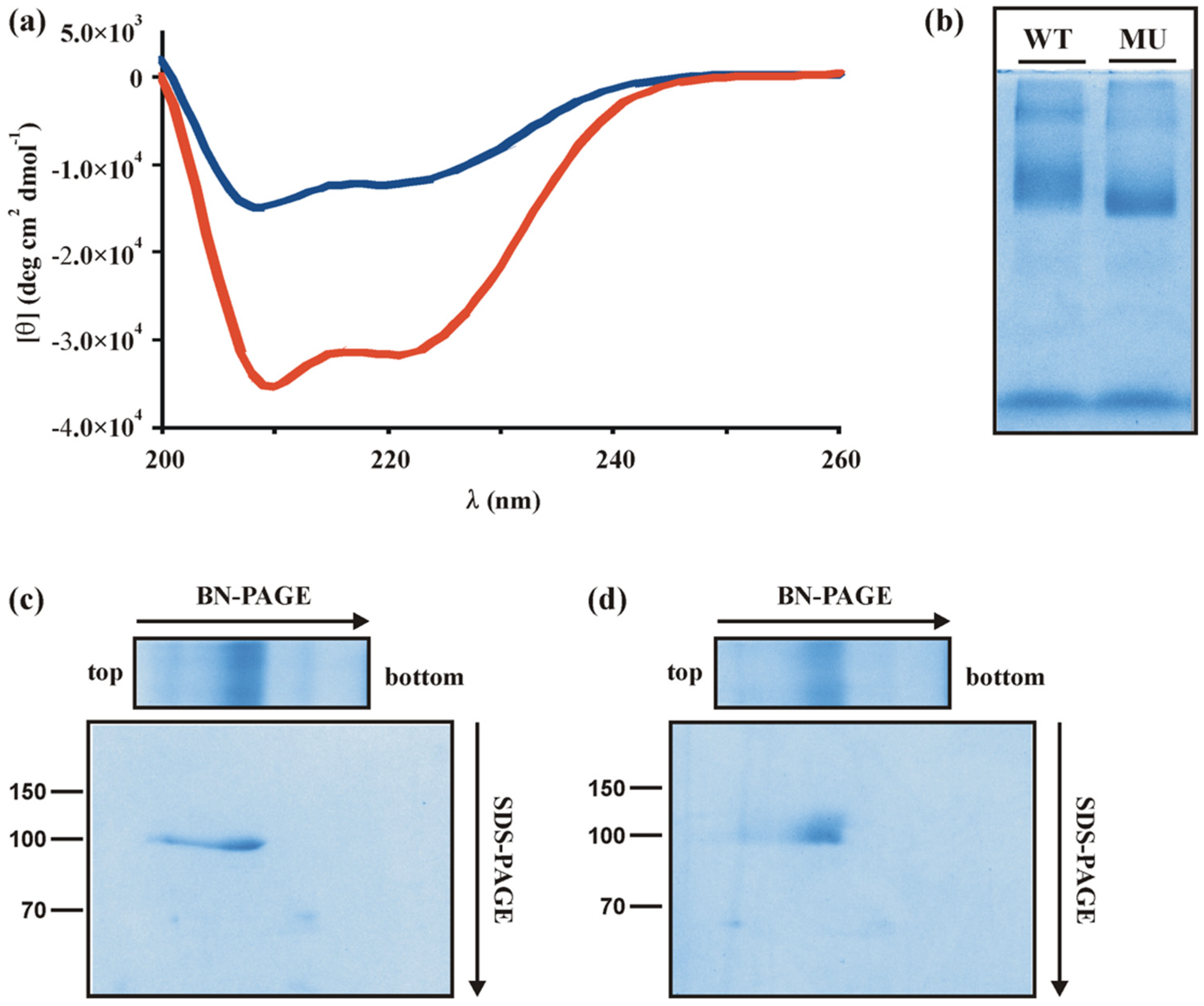
3.1. A Single-Point Mutation in the Zinc Finger Regulatory Protein Xyr1 Changes Its Secondary Structure
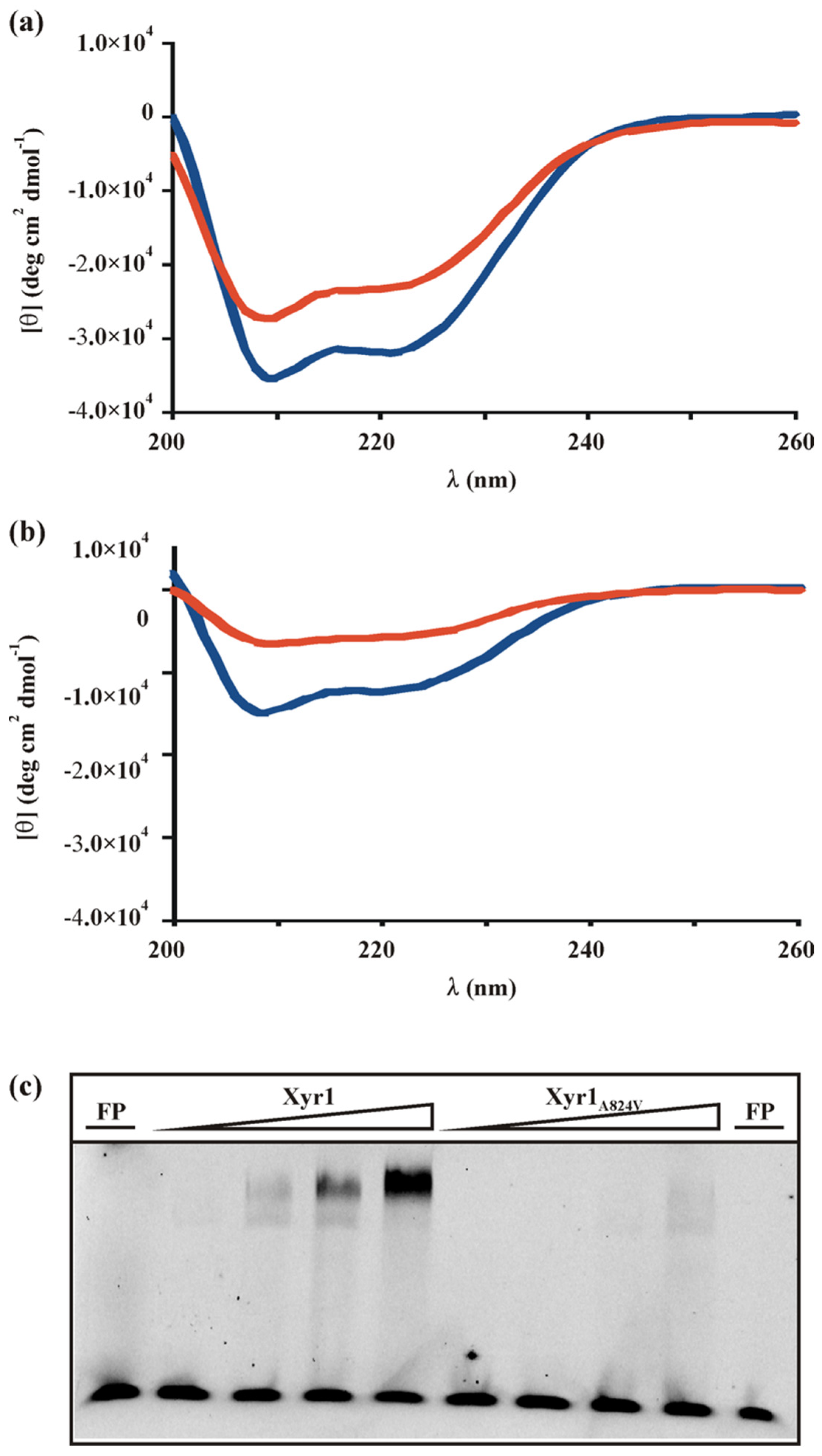
3.2. The A824V Mutation Changes DNA-Binding Properties of Xyr1
3.3. Determination of the Equilibrium Dissociation Constant of the Xyr1 Variants
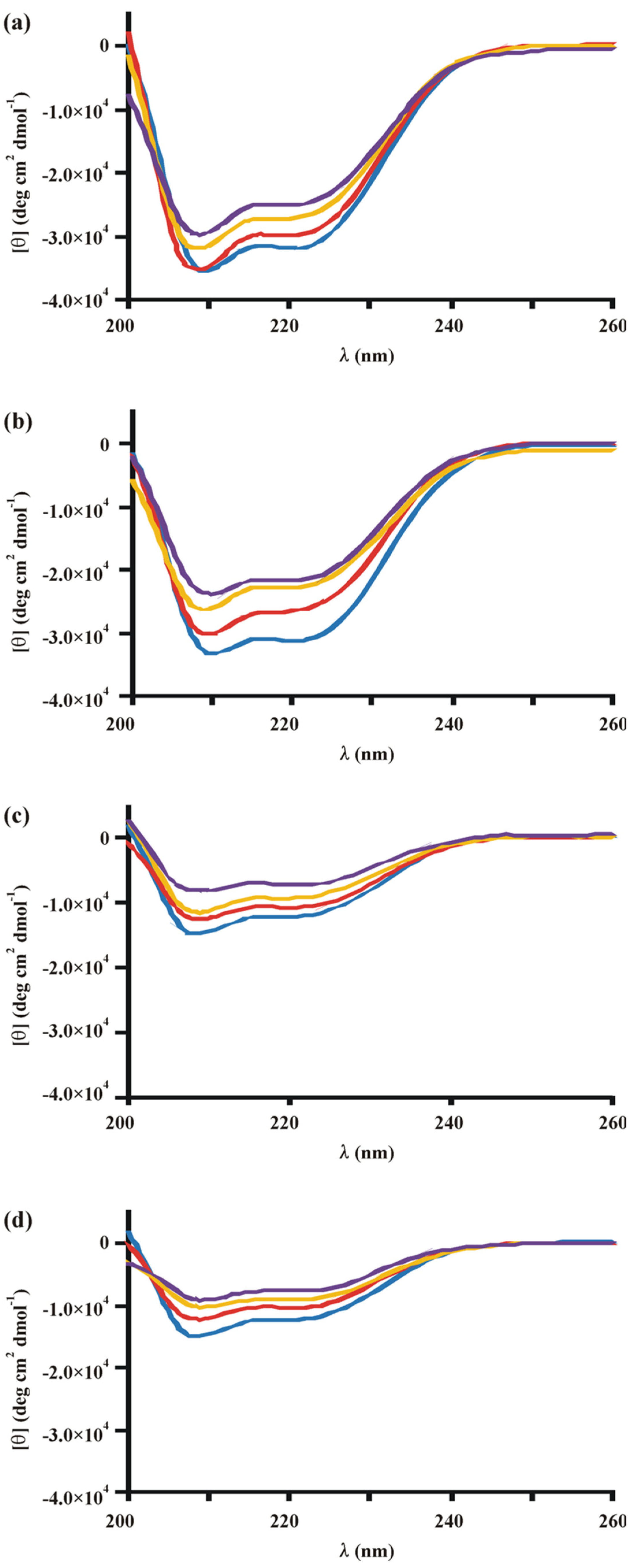
3.4. Conformational Response to Carbohydrates Is Partly Lost in Xyr1A824V
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lieckfeldt, K.K.E.; Samuels, G.J.; Kovacs, W.; Meyer, W.; Petrini, O.; Gams, W.; Borner, T.; Kubicek, C.P. Molecular evidence that the asexual industrial fungus Trichoderma reesei is a clonal derivative of the ascomycete Hypocrea jecorina. Proc. Natl. Acad. Sci. USA 1996, 93, 7755–7760. [Google Scholar]
- Teeri, T.; Salovouri, I.; Knowles, J. The molecular cloning of the major cellulase gene from Trichoderma reesei. Biotechnology 1983, 1, 696–699. [Google Scholar] [CrossRef]
- Törrönen, A.; Harkki, A.; Rouvinen, J. Three-dimensional structure of endo-1,4-beta-xylanase II from Trichoderma reesei: Two conformational states in the active site. EMBO J. 1994, 13, 2493–2501. [Google Scholar] [CrossRef] [PubMed]
- Vaheri, M.P.; Leisola, M.; Kaupinnen, V. Transgycosylation products of the cellulase system of Trichoderma reesei. Biotechnol. Lett. 1979, 1, 41–46. [Google Scholar] [CrossRef]
- Mach, R.L.; Zeilinger, S. Regulation of gene expression in industrial fungi: Trichoderma. Appl. Microbiol. Biotechnol. 2003, 60, 515–522. [Google Scholar] [CrossRef]
- Stricker, A.R.; Mach, R.L.; de Graaff, L.H. Regulation of transcription of cellulases- and hemicellulases-encoding genes in Aspergillus niger and Hypocrea jecorina (Trichoderma reesei). Appl. Microbiol. Biotechnol. 2008, 78, 211–220. [Google Scholar] [CrossRef]
- Stricker, A.R.; Steiger, M.G.; Mach, R.L. Xyr1 receives the lactose induction signal and regulates lactose metabolism in Hypocrea jecorina. FEBS Lett. 2007, 581, 3915–3920. [Google Scholar] [CrossRef] [Green Version]
- Stricker, A.R.; Grosstessner-Hain, K.; Würleitner, E.; Mach, R.L. Xyr1 (xylanase regulator 1) regulates both the hydrolytic enzyme system and D-xylose metabolism in Hypocrea jecorina. Eukaryot. Cell 2006, 5, 2128–2137. [Google Scholar] [CrossRef] [Green Version]
- Portnoy, T.; Margeot, A.; Seidl-Seiboth, V.; le Crom, S.; Chaabane, F.B.; Linke, R.; Seiboth, B.; Kubicek, C.P. Differential regulation of the cellulase transcription factors XYR1, ACE2, and ACE1 in Trichoderma reesei strains producing high and low levels of cellulase. Eukaryot. Cell 2011, 10, 262–271. [Google Scholar] [CrossRef] [Green Version]
- Derntl, C.; Gudynaite-Savitch, L.; Calixte, S.; White, T.; Mach, R.L.; Mach-Aigner, A.R. Mutation of the Xylanase regulator 1 causes a glucose blind hydrolase expressing phenotype in industrially used Trichoderma strains. Biotechnol. Biofuels 2013, 6, 62. [Google Scholar] [CrossRef] [Green Version]
- Mach-Aigner, A.R.; Pucher, M.E.; Steiger, M.G.; Bauer, G.E.; Preis, S.J.; Mach, R.L. Transcriptional regulation of xyr1, encoding the main regulator of the xylanolytic and cellulolytic enzyme system in Hypocrea jecorina. Appl. Environ. Microbiol. 2008, 74, 6554–6562. [Google Scholar] [CrossRef] [PubMed]
- Mach, R.L.; Strauss, J.; Zeilinger, S.; Schindler, M.; Kubicek, C.P. Carbon catabolite repression of xylanase I (xyn1) gene expression in Trichoderma reesei. Mol. Microbiol. 1996, 21, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Mach-Aigner, A.R.; Pucher, M.E.; Mach, R.L. D-Xylose as a repressor or inducer of xylanase expression in Hypocrea jecorina (Trichoderma reesei). Appl. Environ. Microbiol. 2010, 76, 1770–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gielkens, M.M.; Dekkers, E.; Visser, J.; de Graaff, L.H. Two cellobiohydrolase-encoding genes from Aspergillus niger require D- xylose and the xylanolytic transcriptional activator XlnR for their expression. Appl. Environ. Microbiol. 1999, 65, 4340–4345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasper, A.A.; Visser, J.; de Graaff, L.H. The Aspergillus niger transcriptional activator XlnR, which is involved in the degradation of the polysaccharides xylan and cellulose, also regulates D-xylose reductase gene expression. Mol. Microbiol. 2000, 36, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Hasper, A.A.; Trindade, L.M.; van der Veen, D.; van Ooyen, A.J.; de Graaff, L.H. Functional analysis of the transcriptional activator XlnR from Aspergillus niger. Microbiology 2004, 150, 1367–1375. [Google Scholar] [CrossRef] [Green Version]
- Kelly, S.M.; Jess, T.J.; Price, N.C. How to study proteins by circular dichroism. Biochim. Biophys. Acta 2005, 1751, 119–139. [Google Scholar] [CrossRef]
- Ranjbar, B.; Gill, P. Circular dichroism techniques: Biomolecular and nanostructural analyses—A review. Chem. Biol. Drug Des. 2009, 74, 101–120. [Google Scholar] [CrossRef]
- Matthews, J.R.; Nicholson, J.; Jaffray, E.; Kelly, S.M.; Price, N.C.; Hay, R.T. Conformational changes induced by DNA binding of NF-kappa B. Nucleic Acids Res. 1995, 23, 3393–3402. [Google Scholar] [CrossRef]
- Engel, G. Estimation of binding parameters of enzyme-ligand complex from fluorometric data by a curve fitting procedure: Seryl-tRNA synthetase-tRNA Ser complex. Anal. Biochem. 1974, 61, 184–191. [Google Scholar] [CrossRef]
- Stangl, H.; Gruber, F.; Kubicek, C.P. Characterization of the Trichoderma reesei cbh2 promoter. Curr. Genet. 1993, 23, 115–122. [Google Scholar] [CrossRef] [PubMed]
- da Cunha, E.S.; Domingues, C.C.; de Paula, E. Modified native electrophoresis protocol for the solubilization and separation of mitochondrial protein complexes. Anal. Biochem. 2011, 418, 158–160. [Google Scholar] [CrossRef] [PubMed]
- Fiala, G.J.; Schamel, W.W.; Blumenthal, B. Blue native polyacrylamide gel electrophoresis (BN-PAGE) for analysis of multiprotein complexes from cellular lysates. J. Vis. Exp. 2011, 48, e2164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauscher, R.; Würleitner, E.; Wacenovsky, C.; Aro, N.; Stricker, A.R.; Zeilinger, S.; Kubicek, C.P.; Penttilä, M.; Mach, R.L. Transcriptional regulation of xyn1, encoding xylanase I, in Hypocrea jecorina. Eukaryot. Cell 2006, 5, 447–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friden, P.; Schimmel, P. LEU3 of Saccharomyces cerevisiae encodes a factor for control of RNA levels of a group of leucine-specific genes. Mol. Cell. Biol. 1987, 7, 2708–2717. [Google Scholar]
- Friden, P.; Schimmel, P. LEU3 of Saccharomyces cerevisiae activates multiple genes for branched-chain amino acid biosynthesis by binding to a common decanucleotide core sequence. Mol. Cell. Biol. 1988, 8, 2690–2697. [Google Scholar]
- Wang, D.; Hu, Y.; Zheng, F.; Zhou, K.; Kohlhaw, G.B. Evidence that intramolecular interactions are involved in masking the activation domain of transcriptional activator Leu3p. J. Biol. Chem. 1997, 272, 19383–19392. [Google Scholar] [CrossRef] [Green Version]
- Traven, A.; Jelicic, B.; Sopta, M. Yeast Gal4: A transcriptional paradigm revisited. EMBO Rep. 2006, 7, 496–499. [Google Scholar] [CrossRef] [Green Version]
- Stone, G.; Sadowski, I. GAL4 is regulated by a glucose-responsive functional domain. EMBO J. 1993, 12, 1375–1385. [Google Scholar] [CrossRef]
- Ellilä, S.; Fonseca, L.; Uchima, C.; Cota, J.; Goldman, G.H.; Saloheimo, M.; Sacon, V.; Siika-aho, M. Development of a low-cost cellulase production process using Trichoderma reesei for Brazilian biorefineries. Biotechnol. Biofuels 2017, 10, 30. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, L.M.; Parreiras, L.S.; Murakami, M.T. Rational engineering of the Trichoderma reesei RUT-C30 strain into an industrially relevant platform for cellulase production. Biotechnol. Biofuels 2020, 13, 93. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Valkonen, M.; Jackson, R.E.; Palmer, J.M.; Bhalla, A.; Nikolaev, I.; Saloheimo, M.; Ward, M. Modification of transcriptional factor ACE3 enhances protein production in Trichoderma reesei in the absence of cellulase gene inducer. Biotechnol. Biofuels 2020, 13, 137. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Li, Z.; Xu, Y.; Yang, P.; Gao, L.; Yan, Q.; Li, S.; Wang, Y.; Qu, Y.; Song, X. Introduction of heterologous transcription factors and their target genes into Penicillium oxalicum leads to increased lignocellulolytic enzyme production. Appl. Microbiol. Biotechnol. 2019, 103, 2675–2687. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′–3′) | Employment |
|---|---|---|
| Pxyn1f_FAM | [FAM]- TTGGCAGGCTAAATGCGACATCTTAGCCGGATGCA | EMSA |
| Pxyn1f | TTGGCAGGCTAAATGCGACATCTTAGCCGGATGCA | CD |
| Pxyn1r | TGCATCCGGCTAAGATGTCGCATTTAGCCTGCCAA | EMSA/CD |
| CKT067 | CACTCCACATGTTAAAGGCGCATTCAACCAGCTTC | EMSA |
| CKT068 | GAAGCTGGTTGAATGCGCCTTTAACATGTGGAGTG | EMSA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mello-de-Sousa, T.M.; Gorsche, R.; Jovanović, B.; Mach, R.L.; Mach-Aigner, A.R. In Vitro Characterization of a Nuclear Receptor-like Domain of the Xylanase Regulator 1 from Trichoderma reesei. J. Fungi 2022, 8, 1254. https://doi.org/10.3390/jof8121254
Mello-de-Sousa TM, Gorsche R, Jovanović B, Mach RL, Mach-Aigner AR. In Vitro Characterization of a Nuclear Receptor-like Domain of the Xylanase Regulator 1 from Trichoderma reesei. Journal of Fungi. 2022; 8(12):1254. https://doi.org/10.3390/jof8121254
Chicago/Turabian StyleMello-de-Sousa, Thiago M., Rita Gorsche, Birgit Jovanović, Robert L. Mach, and Astrid R. Mach-Aigner. 2022. "In Vitro Characterization of a Nuclear Receptor-like Domain of the Xylanase Regulator 1 from Trichoderma reesei" Journal of Fungi 8, no. 12: 1254. https://doi.org/10.3390/jof8121254