
The Analysis of the Mycobiota in Plastic Polluted Soil Reveals a Reduction in Metabolic Ability

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Sample Collection
2.2. Evaluation of Soil Plastic Fragments Counts
2.3. Soil Physico-Chemical Analyses
2.4. Evaluation of Soil Fungal Counts
2.5. Soil Microbial Communities Metabolic Profiling
2.6. DNA Metabarcoding and Bioinformatic Data Analysis
2.7. Statistical Analysis
3. Results
3.1. Evaluation of Soil Plastic Fragments Counts
3.2. Soil Physico-Chemical Analyses
3.3. Evaluation of Soil Fungal Counts
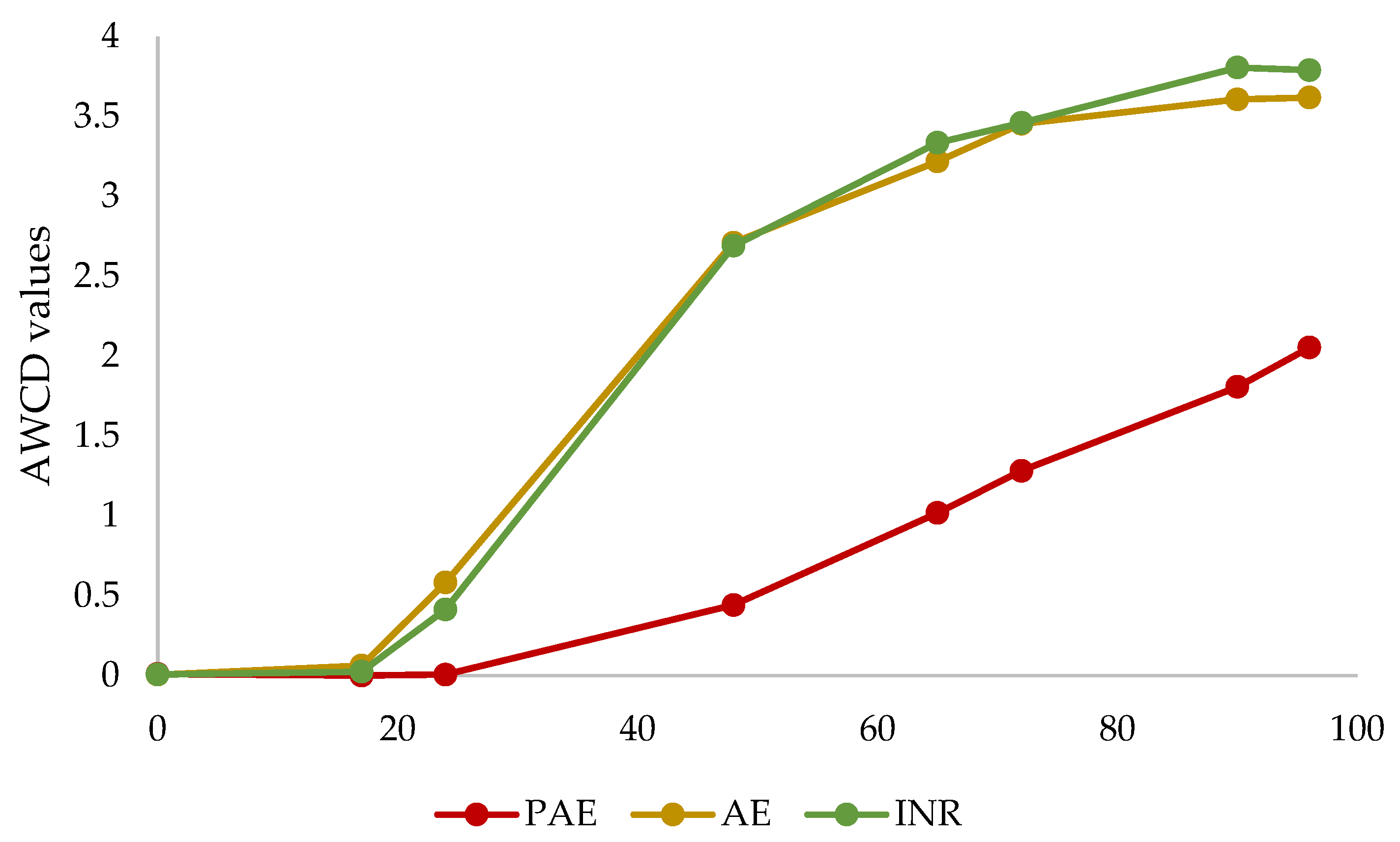
3.4. Soil Microbial Communities Metabolic Profiling
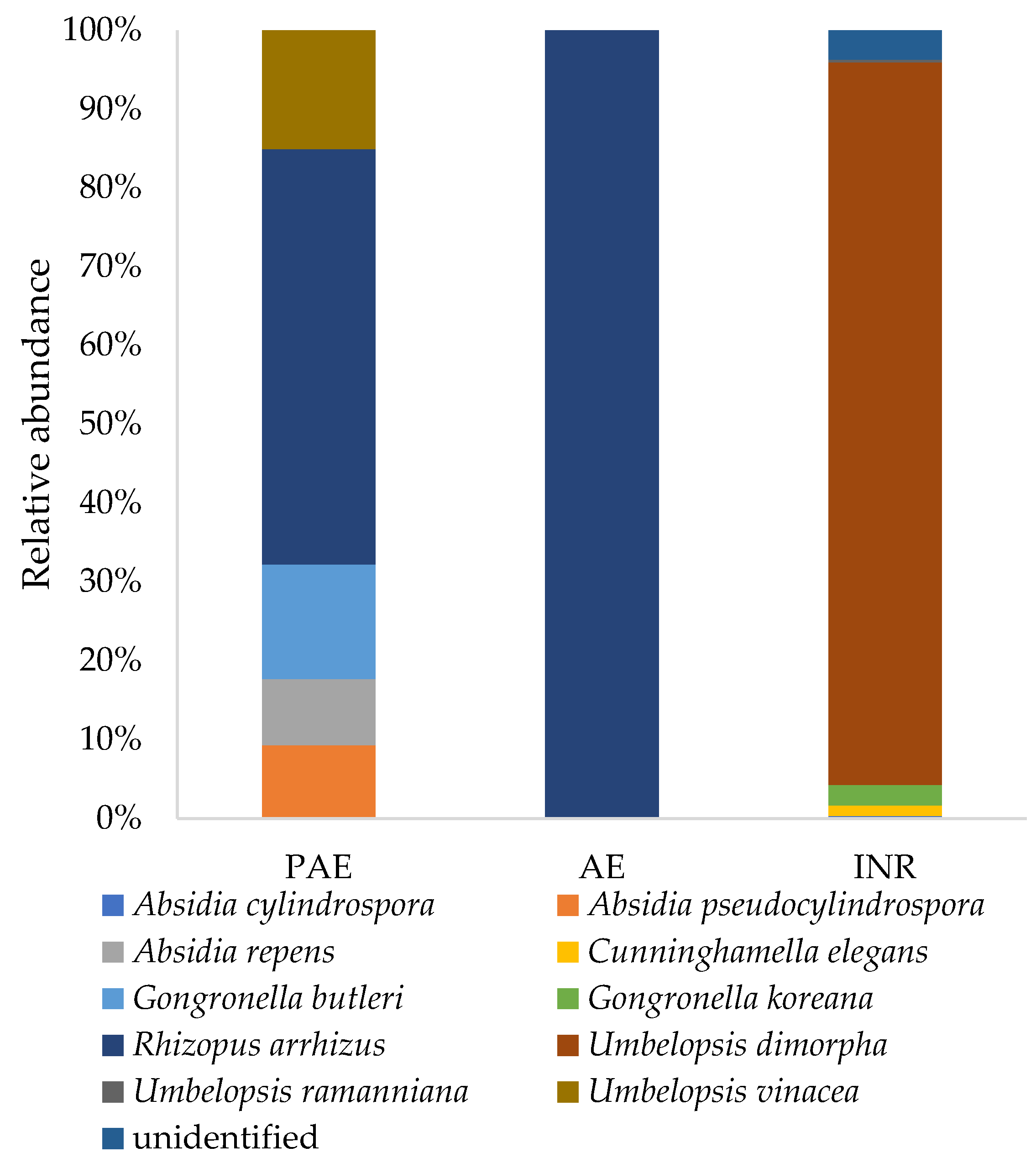
3.5. Soil Fungal Composition
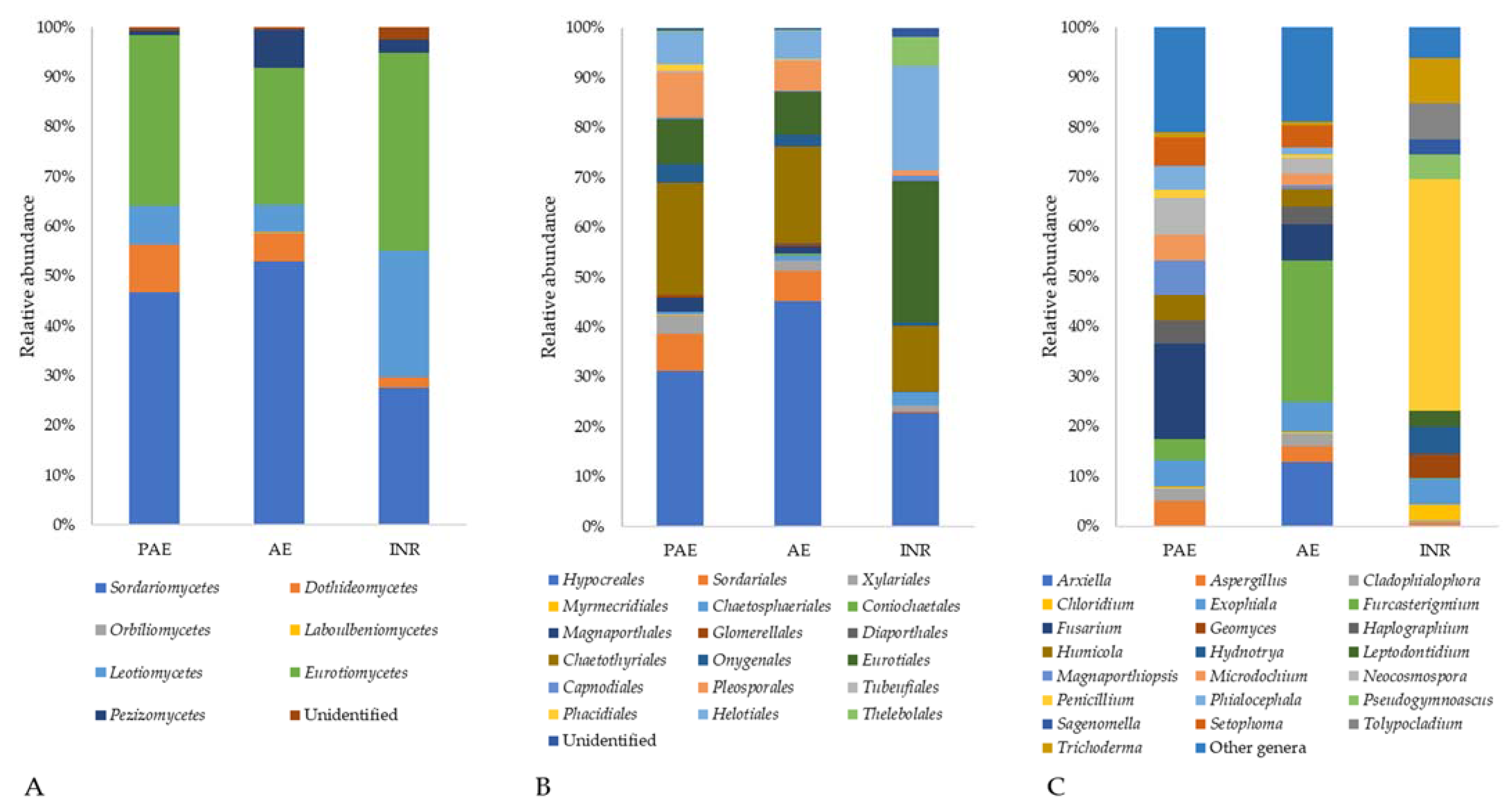
3.5.1. Phylum Ascomycota
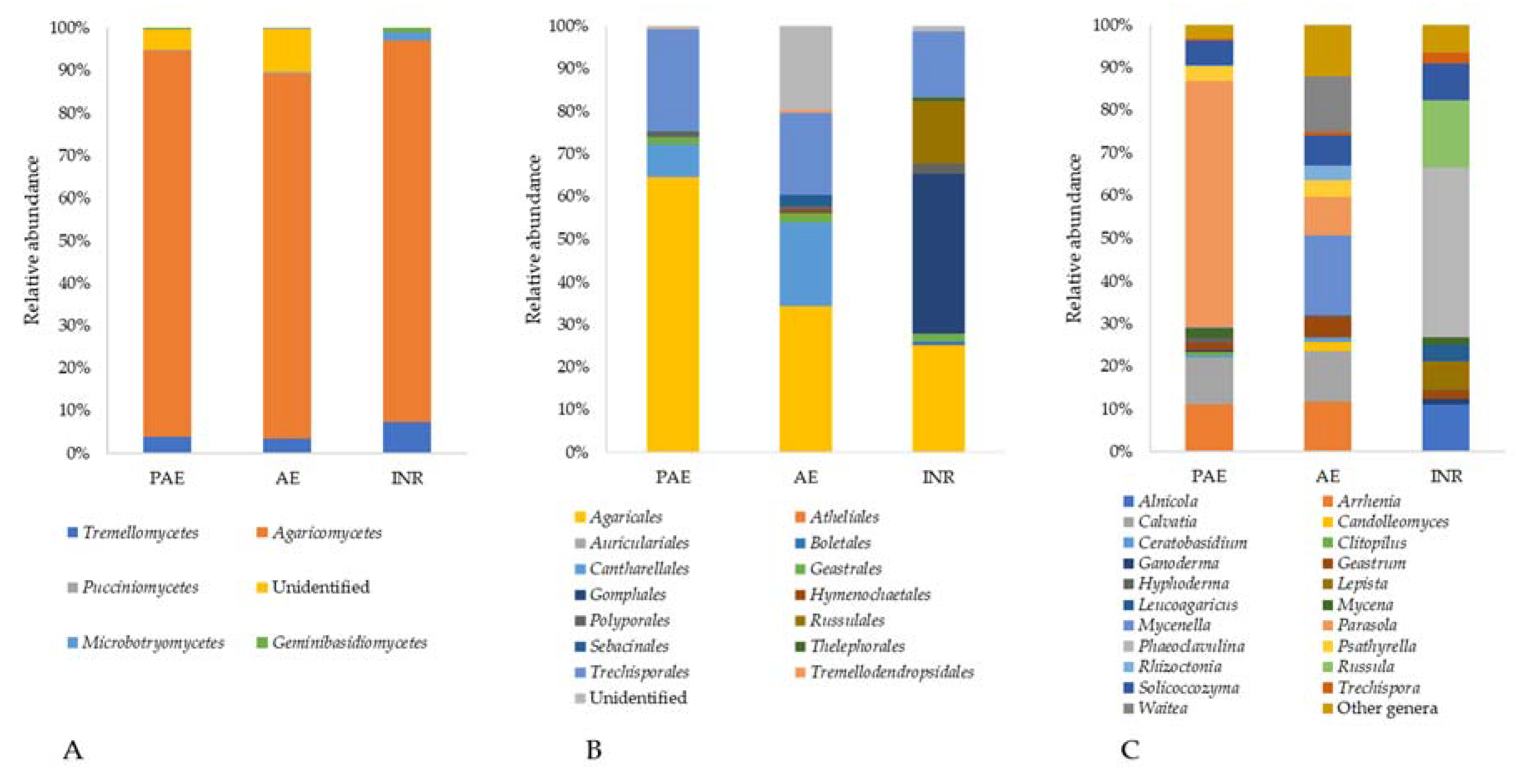
3.5.2. Phylum Basidiomycota
3.5.3. Phylum Mortierellomycota
3.5.4. Phylum Mucoromycota
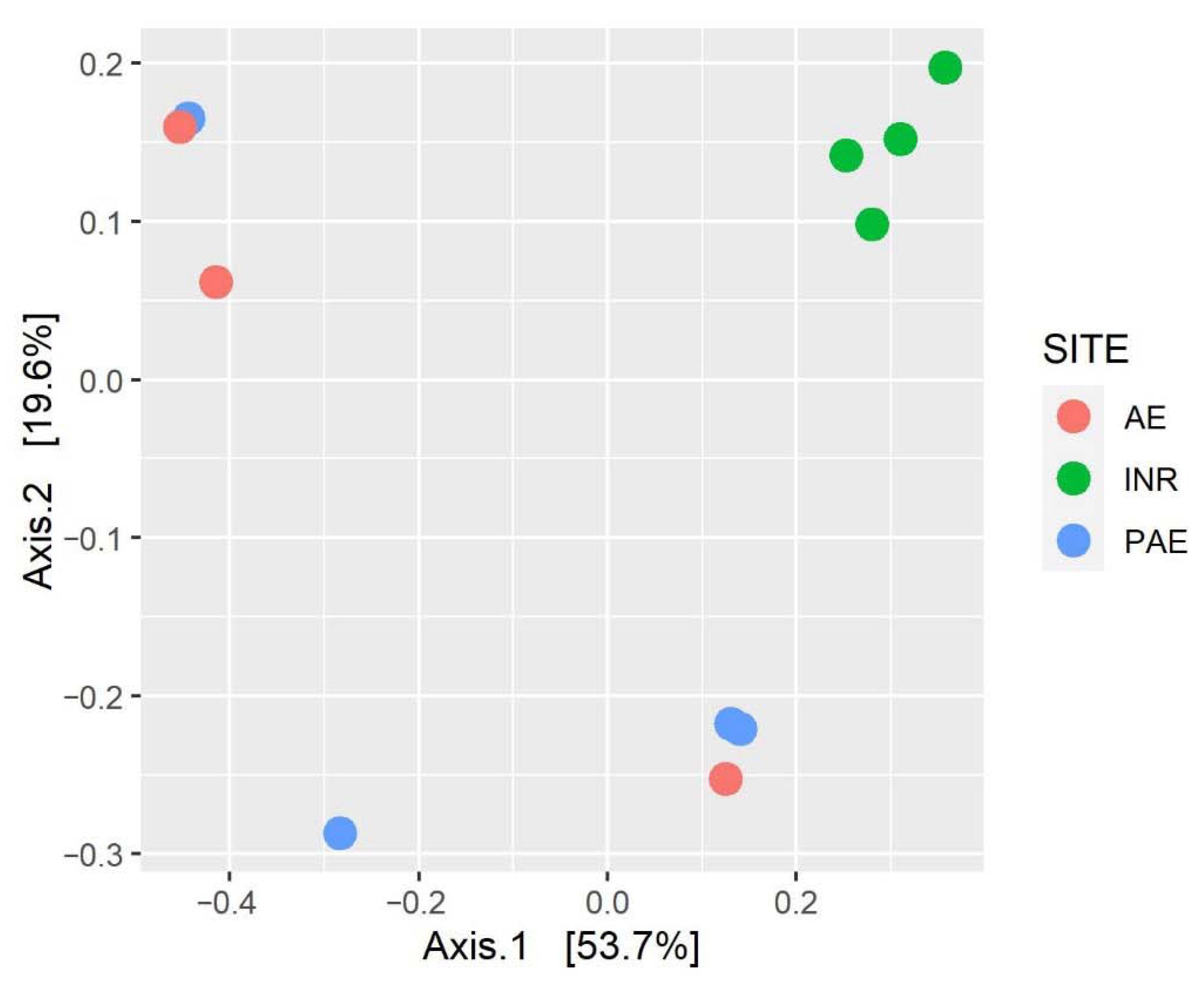
3.6. Diversity of the Soil Mycobiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Plastic Europe. Plastics–The Facts 2021: An Analysis of European Plastics Production, Demand and Waste Data. 2021. Available online: https://plasticseurope.org/knowledge-hub/plastics-the-facts-2021/ (accessed on 14 June 2022).
- Ruimin, Q.; Jones, D.L.; Zhen, L.; Qin, L.; Changrong, Y. Behavior of microplastics and plastic film residues in the soil environment: A critical review. Sci. Total Environ. 2019, 703, 134722. [Google Scholar] [CrossRef]
- Fan, P.; Tan, W.; Yu, H. Effects of different concentrations and types of microplastics on bacteria and fungi in alkaline soil. Ecotoxicol. Environ. Saf. 2022, 229, 113045. [Google Scholar] [CrossRef] [PubMed]
- Mao, R.; Lang, M.; Yu, X.; Wu, R.; Yang, X.; Guo, X. Aging mechanism of microplastics with UV irradiation and its effects on the adsorption of heavy metals. J. Hazard. Mater. 2020, 393, 122515. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.C.; Ylva, O.; Mitchell, R.P.; Anthony, D.; Rowland, S.J.; John, A.W.G.; Daniel, M.G.; Russell, A.E. Lost at sea: Where is all the plastic? Science 2004, 304, 838. [Google Scholar] [CrossRef]
- Huang, B.; Sun, L.; Liu, M.; Huang, H.; He, H.; Han, F.; Wang, X.; Xu, Z.; Li, B.; Pan, X. Abundance and distribution characteristics of microplastic in plateau culti-vated land of Yunnan Province, China. Environ. Sci. Pollut. Res. 2021, 28, 1675–1688. [Google Scholar] [CrossRef]
- Liu, M.; Lu, S.; Song, Y.; Lei, L.; Hu, J.; Lv, W.; Zhou, W.; Cao, C.; Shi, H.; Yang, X.; et al. Microplastic and mesoplastic pollution in farmland soils in suburbs of Shanghai, China. Environ. Pollut. 2018, 242, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Horton, A.A.; Walton, A.; Spurgeon, D.J.; Lahive, E.; Svendsen, C. Microplastics in freshwater and terrestrial environments: Evaluating the current understanding to identify the knowledge gaps and future research priorities. Sci. Total Environ. 2017, 586, 127–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacques, O.; Prosser, R.S. A probabilistic risk assessment of microplastics in soil ecosystems. Sci. Total Environ. 2021, 757, 143987. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, Y.; Ouyang, D.; Lei, J.; Tan, Q.; Xie, L.; Li, Z.; Liu, T.; Xiao, Y.; Farooq, T.H.; et al. Systematical review of interactions between microplastics and microorganisms in the soil environment. J. Hazard. Mater. 2021, 418, 126288. [Google Scholar] [CrossRef] [PubMed]
- Rong, L.; Zhao, L.; Zhao, L.; Cheng, Z.; Yao, Y.; Yuan, C.; Wang, L.; Sun, H. LDPE microplastics affect soil microbial communities and nitrogen cycling. Sci. Total Environ. 2021, 773, 145640. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Liu, W.; Zhao, F.; Zhong, Z.; Deng, J.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Soil bacterial and fungal diversity and compositions respond differently to forest development. Catena 2019, 181, 104071. [Google Scholar] [CrossRef]
- Doran, J.W.; Zeiss, M.R. Soil health and sustainability: Managing the biotic component of soil quality. Appl. Soil Ecol. 2000, 15, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, R.H.; Anslan, S.; Bahram, M.; Wurzbacher, C.; Baldrian, P.; Tedersoo, L. Mycobiome diversity: High-throughput sequencing and identification of fungi. Nat. Rev. Microb. 2019, 17, 95–109. [Google Scholar] [CrossRef]
- Bani, A.; Pioli, S.; Ventura, M.; Panzacchi, P.; Borruso, L.; Tognetti, R.; Tonon, G.; Brusetti, L. The role of microbial community in the decomposition of leaf litter and deadwood. Appl. Soil Ecol. 2018, 126, 75–84. [Google Scholar] [CrossRef]
- van der Wal, A.; Geydan, T.D.; Kuyper, T.W.; De Boer, W. A thready affair: Linking fungal diversity and community dynamics to terrestrial decomposition processes. FEMS Microb. Rev. 2013, 37, 477–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devi, R.; Kaur, T.; Kour, D.; Rana, K.L.; Yadav, A.; Yadav, A.N. Beneficial fungal communities from different habitats and their roles in plant growth promotion and soil health. Microb. Biosyst. 2020, 5, 21–47. [Google Scholar] [CrossRef]
- Du, H.; Wang, J. Characterization and environmental impacts of microplastics. Gondwana Res. 2021, 98, 63–75. [Google Scholar] [CrossRef]
- Yu, H.; Fan, P.; Hou, J.; Dang, Q.; Cui, D.; Xi, B.; Tan, W. Inhibitory effect of microplastics on soil extracellular enzymatic activities by changing soil properties and direct adsorption: An investigation at the aggregate-fraction level. Environ. Pollut. 2020, 267, 115544. [Google Scholar] [CrossRef] [PubMed]
- Fei, Y.; Huang, S.; Zhang, H.; Tong, Y.; Wen, D.; Xia, X.; Wang, H.; Luo, Y.; Barceló, D. Response of soil enzyme activities and bacterial communities to the accumulation of microplastics in an acid cropped soil. Sci. Total Environ. 2020, 707, 135634. [Google Scholar] [CrossRef]
- de Souza Machado, A.A.; Lau, C.W.; Kloas, W.; Bergmann, J.; BachelierI, B.; Faltin, E.; Becker, R.; Görlich, A.S.; Rillig, M.C. Microplastics can change soil properties and affect plant performance. Environ. Sci. Technol. 2019, 53, 6044–6052. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Zhang, M.; Liu, G.; Lu, T.; Qu, Q.; Du, B.; Pan, X. Effects of soil residual plastic film on soil microbial community structure and fertility. Water Air Soil Pollut. 2018, 229, 261. [Google Scholar] [CrossRef]
- Gao, B.; Yao, H.; Li, Y.; Zhu, Y. Microplastic addition alters the microbial community structure and stimulates soil carbon dioxide emissions in vegetable-growing soil. Environ. Toxicol. Chem. 2021, 40, 352–365. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yang, X.; Liu, G.; Liang, C.; Xue, S.; Chen, H.; Ritsema, C.J.; Geissen, V. Response of soil dissolved organic matter to microplastic addition in Chinese loess soil. Chemosphere 2017, 185, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Awet, T.T.; Kohl, Y.; Meier, F.; StrasIba, S.; Grün, A.L.; Ruf, T.; Jost, C.; Drexel, R.; Tunc, E.; Emmerling, C. Effects of polystyrene nanoparticles on the microbiota and functional diversity of enzymes in soil. Environ. Sci. Eur. 2018, 30, 11. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lv, S.; Zhang, M.; Chen, G.; Zhu, T.; Zhang, S.; Christie, P.; Luo, Y. Effects of plastic film residues on occurrence of phthalates and microbial activity in soils. Chemosphere 2016, 151, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Ventorino, V.; Pascale, A.; Adamo, P.; Rocco, C.; Fiorentino, N.; Mori, M.; Faraco, V.; Pepe, O.; Fagnano, M. Comparative assessment of autochthonous bacterial and fungal communities and microbial biomarkers of polluted agricultural soils of the Terra dei Fuochi. Sci. Rep. 2018, 8, 14281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Fan, J.; Zhu, W.; Amombo, E.; Lou, Y.; Chen, L.; Fu, J. Effect of heavy metals pollution on soil microbial diversity and bermudagrass genetic variation. Front. Plant Sci. 2016, 7, 755. [Google Scholar] [CrossRef] [Green Version]
- Galitskaya, P.; Biktasheva, L.; Blagodatsky, S.; Selivanovskaya, S. Response of bacterial and fungal communities to high petroleum pollution in different soils. Sci. Rep. 2021, 11, 164. [Google Scholar] [CrossRef] [PubMed]
- Chikere, C.B.; Mordi, I.J.; Chikere, B.O.; Selvarajan, R.; Ashafa, T.O.; Obieze, C.C. Comparative metagenomics and functional profiling of crude oil-polluted soils in Bodo West Community, Ogoni, with other sites of varying pollution history. Ann. Microb. 2019, 69, 495–513. [Google Scholar] [CrossRef]
- Bosco Siro Negri. 2022. Available online: https://boscosironegri.unipv.it/la-riserva/ (accessed on 14 September 2022).
- Crichton, E.M.; Noël, M.; Gies, E.A.; Ross, P.S.A. Novel, Density-Independent and FTIR-Compatible Approach for the Rapid Extraction of Microplastics from Aquatic Sediments. Anal. Methods 2017, 9, 1419–1428. [Google Scholar] [CrossRef]
- Gams, W.; Hoestra, E.S.; Aptroot, A. CSB Course in Mycology, 4th ed.; CBS: Baarn, The Netherlands, 1998. [Google Scholar]
- Landínez-Torres, A.Y.; Panelli, S.; Picco, A.M.; Comandatore, F.; Tosi, S.; Capelli, E. A meta-barcoding nalysis of soil mycobiota of the upper Andean Colombian agro-environment. Sci. Rep.-UK 2019, 9, 10085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temporiti, M.E.E.; Daccò, C.; Nicola, L. Isolation and Screening from Soil Biodiversity for Fungi Involved in the Degradation of Recalcitrant Materials. J. Vis. Exp. 2022, 183, e63445. [Google Scholar] [CrossRef]
- Zabinski, C.A.; Gannon, J.E. Effects of recreational impacts onsoil microbial communities. Environ. Manag. 1997, 21, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhu, L.; Yang, T.; Qian, Z.; Xu, C.; Tian, D.; Tang, L. Poplar agroforestry systems in eastern China enhance the spatiotemporal stability of soil microbial community structure and metabolism. Land Degrad. Dev. 2022, 33, 916–930. [Google Scholar] [CrossRef]
- Nicola, L.; Landinez-Torres, A.Y.; Zambuto, F.; Capelli, E.; Tosi, S. The mycobiota of high altitude pear orchards soil in Colombia. Biology 2021, 10, 1002. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Mills, D.A. Improved Selection of Internal Transcribed Spacer-Specific Primers Enables Quantitative, Ul-tra-High-Throughput Profiling of Fungal Communities. Appl. Environ. Microb. 2013, 79, 2519–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Caporaso, J.G.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2018, 47, D259–D264. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Sánchez-Ramírez, S.; Kõljalg, U.; Bahram, M.; Döring, M.; Schigel, D.; May, T.; Ryberg, M.; Abarenkov, K. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Divers. 2018, 90, 135–159. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- McMurdie, P.J.; Holmes, S. Phyloseq: An r package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wu, Y.; Ma, J.; An, Y.; Liu, Q.; Yang, S.; Qu, Y.; Chen, H.; Zhao, W.; Tian, Y. Microplastics pollution in the soil mulched by dust-proof nets: A case study in Beijing, China. Environ. Pollut. 2021, 275, 116600. [Google Scholar] [CrossRef] [PubMed]
- Chai, B.; Wei, Q.; She, Y.; Lu, G.; Dang, Z.; Yin, H. Soil microplastic pollution in an e-waste dismantling zone of China. Waste Manag. 2020, 118, 291–301. [Google Scholar] [CrossRef] [PubMed]
- van den Berg, P.; Huerta-Lwanga, E.; Corradini, F.; Geissen, V. Sewage sludge application as a vehicle for microplastics in eastern Spanish agricultural soils. Environ. Pollut. 2020, 261, 114198. [Google Scholar] [CrossRef]
- Huang, Y.; Liu, Q.; Jia, W.; Yan, C.; Wang, J. Agricultural plastic mulching as a source of microplastics in the terrestrial environment. Environ. Pollut. 2020, 260, 114096. [Google Scholar] [CrossRef]
- Corradini, F.; Meza, P.; Eguiluz, R.; Casado, F.; Huerta-Lwanga, E.; Geissen, V. Evidence of microplastic accumulation in agricultural soils from sewage sludge disposal. Sci. Total Environ. 2019, 671, 411–420. [Google Scholar] [CrossRef]
- Zubris, K.A.V.; Richards, B.K. Synthetic fibers as an indicator of land application of sludge. Environ. Pollut. 2005, 138, 201–211. [Google Scholar] [CrossRef]
- Schoenholtz, S.H.; Van Miegroet, H.; Burger, J.A. A review of chemical and physical properties as indicators of forest soil quality: Challenges and opportunities. For. Ecol. Manag. 2000, 138, 335–356. [Google Scholar] [CrossRef]
- Bünemann, E.K.; Bongiorno, G.; Bai, Z.; Creamer, R.E.; De Deyn, G.; de Goede, R.; Fleskens, L.; Geissen, V.; Kuyper, T.W.; Mäder, P.; et al. Soil quality–A critical review. Soil Biol. Biochem. 2018, 120, 105–125. [Google Scholar] [CrossRef]
- Romig, D.E.; Garlynd, M.J.; Harris, R.F. Farmer-based assessment of soil quality: A soil health scorecard. Methods Assess. Soil Qual. 1997, 49, 39–60. [Google Scholar] [CrossRef] [Green Version]
- Quijano, L.; Van Oost, K.; Nadeu, E.; Gaspar, L.; Navas, A. Modelling the effect of land management changes on soil organic carbon stocks in a Mediterranean cultivated field. Land Degrad. Dev. 2017, 28, 515–523. [Google Scholar] [CrossRef]
- Bruun, T.B.; Elberling, B.; de Neergaard, A.; Magid, J. Organic carbon dynamics in different soil types after conversion of forest to agriculture. Land Degrad. Dev. 2015, 26, 272–283. [Google Scholar] [CrossRef]
- Slobodian, N.; Van Rees, K.; Pennock, D. Cultivation-induced effects on belowground biomass and organic carbon. Soil Sci. Soc. Am. J. 2002, 66, 924–930. [Google Scholar] [CrossRef]
- Zhao, T.; Lozano, Y.M.; Rillig, M.C. Microplastics increase soil pH and decrease microbial activities as a function of microplastic shape, polymer type, and exposure time. Front. Environ. Sci. 2021, 9, 675803. [Google Scholar] [CrossRef]
- Lozano, Y.M.; Aguilar-Trigueros, C.A.; Onandia, G.; Maaß, S.; Zhao, T.; Rillig, M.C. Effects of microplastics and drought on soil ecosystem functions and multifunctionality. J. Appl. Ecol. 2021, 58, 988–996. [Google Scholar] [CrossRef]
- You, J.; Liu, X.; Zhang, B.; Xie, Z.; Hou, Z.; Yang, Z. Seasonal changes in soil acidity and related properties in ginseng artificial bed soils under a plastic shade. J. Ginseng Res. 2015, 39, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Stefanowicz, A. The Biolog Plates Technique as a Tool in Ecological Studies of Microbial Communities. Polish J. Environ. Stud. 2006, 15, 669–676. [Google Scholar]
- Gomez, E.; Ferreras, L.; Toresani, S. Soil bacterial functional diversity as influenced by organic amendment application. Bioresour. Technol. 2006, 97, 1484–1489. [Google Scholar] [CrossRef]
- Kraus, T.E.; Dahlgren, R.A.; Zasoski, R.J. Tannins in nutrient dynamics of forest ecosystems-a review. Plant Soil 2003, 256, 41–66. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Vitousek, P.M. The role of polyphenols in terrestrial ecosystem nutrient cycling. Trends Ecol. Evol. 2000, 15, 238–243. [Google Scholar] [CrossRef]
- Min, K.; Freeman, C.; Kang, H.; Choi, S.U. The regulation by phenolic compounds of soil organic matter dynamics under a changing environment. BioMed. Res. Int. 2015, 2015, 825098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wunderlichová, L.; Buňková, L.; Koutný, M.; Jančová, P.; Buňka, F. Formation, degradation, and detoxification of putrescine by foodborne bacteria: A review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 1012–1030. [Google Scholar] [CrossRef]
- Yuebing, S.; Shunan, Z.; Lin, W.; Xuefeng, L.; Yingming, X. Changes of enzymatic activities, substrate utilization pattern, and microbial community diversity in heavy metal-contaminated soils. Water Air Soil Pollut. 2020, 231, 422. [Google Scholar] [CrossRef]
- Zhang, C.; Zhou, T.; Zhu, L.; Du, Z.; Li, B.; Wang, J.; Wang, J.; Sun, Y. Using enzyme activities and soil microbial diversity to understand the effects of fluoxastrobin on microorganisms in fluvo-aquic soil. Sci. Total Environ. 2019, 666, 89–93. [Google Scholar] [CrossRef]
- Gryta, A.; Frąc, M.; Oszust, K. The application of the Biolog EcoPlate approach in ecotoxicological evaluation of dairy sewage sludge. Appl. Biochem. Biotechnol. 2014, 174, 1434–1443. [Google Scholar] [CrossRef] [Green Version]
- Gu, M.; Zhang, Z.; Wang, W.; Tang, Q.; Song, S.; Zhu, J.; Xie, Y.; Zhang, L. The effects of radiation pollution on the population diversities and metabolic characteristics of soil microorganisms. Water Air Soil Pollut. 2014, 225, 2133. [Google Scholar] [CrossRef]
- Saravanan, A.; Kumar, P.S.; Vo, D.V.N.; Jeevanantham, S.; Karishma, S.; Yaashikaa, P.R. A review on catalytic-enzyme degradation of toxic environmental pollutants: Microbial enzymes. J. Hazard. Mater. 2021, 419, 126451. [Google Scholar] [CrossRef]
- Sánchez, C. Fungal potential for the degradation of petroleum-based polymers: An overview of macro-and microplastics biodegradation. Biotechnol. Adv. 2020, 40, 107501. [Google Scholar] [CrossRef]
- Singh, R.K.; Tripathi, R.; Ranjan, A.; Srivastava, A.K. Fungi as potential candidates for bioremediation. In Abatement of Environmental Pollutants; Elsevier: Amsterdam, The Netherlands, 2020; pp. 177–191. [Google Scholar]
- Yu, H.; Zhang, Y.; Tan, W. The “neighbor avoidance effect” of microplastics on bacterial and fungal diversity and communities in different soil horizons. E.S.E. 2021, 8, 100121. [Google Scholar] [CrossRef]
- Li, H.Z.; Zhu, D.; Lindhardt, J.H.; Lin, S.M.; Ke, X.; Cui, L. Long-term fertilization history alters effects of microplastics on soil properties, microbial communities, and functions in diverse farmland ecosystem. Environ. Sci. Technol. 2021, 55, 4658–4668. [Google Scholar] [CrossRef]
- Yi, M.; Zhou, S.; Zhang, L.; Ding, S. The effects of three different microplastics on enzyme activities and microbial communities in soil. Water Environ. Res. 2021, 93, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Tennakoon, D.S.; Kuo, C.H.; Maharachchikumbura, S.S.; Thambugala, K.M.; Gentekaki, E.; Phillips, A.J.; Bhat, D.J.; Wanasinghe, D.N.; de Silva, N.I.; Promputtha, I.; et al. Taxonomic and phylogenetic contributions to Celtis formosana, Ficus ampelas, F. septica, Macaranga tanarius and Morus australis leaf litter inhabiting microfungi. Fungal Divers. 2021, 108, 1–215. [Google Scholar] [CrossRef]
- Monkai, J.; Jayawardena, R.S.; Maharachchikumbura, S.S.; Hyde, K.D. Sexual morph of furcasterigmium furcatum (Plectosphaerellaceae) from magnolia liliifera collected in northern thailand. Phyton 2020, 89, 765. [Google Scholar] [CrossRef]
- Sharma, L.; Marques, G. Fusarium, an entomopathogen—A myth or reality? Pathogens 2018, 7, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.J.; Geiser, D.M.; Proctor, R.H.; Rooney, A.P.; O’Donnell, K.; Trail, F.; Gardiner, D.M.; Manners, J.M.; Kazan, K. Fusarium pathogenomics. Annu. Rev. Microb. 2013, 67, 399–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, A.A.; Hasan, F.; Hameed, A.; Akhter, J.I. Isolation of Fusarium sp. AF4 from sewage sludge, with the ability to adhere the surface of polyethylene. Afr. J. Microb. Res. 2009, 3, 658–663. [Google Scholar]
- Dor, S.; Degani, O. Uncovering the Host Range for Maize Pathogen Magnaporthiopsis maydis. Plants 2019, 8, 259. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Wang, L.; Tang, J.; Sun, H.; Giesy, J.P. Combined effects of degradable film fragments and micro/nanoplastics on growth of wheat seedling and rhizosphere microbes. Environ. Pollut. 2022, 294, 118516. [Google Scholar] [CrossRef]
- Redhead, S.A.; Vilgalys, R.; Moncalvo, J.M.; Johnson, J.; Hopple, J.S., Jr. Coprinus Pers. and the disposition of Coprinus species sensu lato. Taxon 2001, 50, 203–241. [Google Scholar] [CrossRef]
- Schafer, D.J. Keys to sections of Parasola, Coprinellus, Coprinopsis and Coprinus in Britain. Field Mycol. 2010, 11, 44–51. [Google Scholar] [CrossRef]
- Robich, G. Mycena D’Europa; AMB, Fondazione Centro Studi Micologici: Trento, Italy, 2003. [Google Scholar]
- Wang, T.; Su, D.; Wang, X.; He, Z. Adsorption-degradation of polycyclic aromatic hydrocarbons in soil by immobilized mixed bacteria and its effect on microbial communities. J. Agric. Food Chem. 2020, 68, 14907–14916. [Google Scholar] [CrossRef] [PubMed]
- Kubatova, A.; Prasil, K.; Vanova, M. Diversity of soil microscopic fungi on abandoned industrial deposits. Cryptogam. Mycol. 2002, 3, 205–219. [Google Scholar]
- Shilpa, P. Microbial Inoculants for Enhancing Degradation of Biosolid Waste in Aerobic Composting. Master’s Thesis, Kerala Agricultural University, Thrissur, India, 2019. [Google Scholar]
- Valsalan, R.; Mathew, D.; Devaki, G. Draft genome of Gongronella butleri reveals the genes contributing to its biodegradation potential. J. Genet Eng. Biotechnol. 2022, 20, 74. [Google Scholar] [CrossRef]
- Nagy, L.G.; Petkovits, T.; Kovács, G.M.; Voigt, K.; Vágvölgyi, C.; Papp, T. Where is the unseen fungal diversity hidden? A study of Mortierella reveals a large contribution of reference collections to the identification of fungal environmental sequences. New Phytol. 2011, 191, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Daghino, S.; Murat, C.; Sizzano, E.; Girlanda, M.; Perotto, S. Fungal diversity is not determined by mineral and chemical differences in serpentine substrates. PLoS ONE 2012, 7, e44233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grządziel, J.; Gałązka, A. Fungal biodiversity of the most common types of Polish soil in a long-term microplot experiment. Front. Microb. 2019, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Tian, J.; Bai, C.; Xiang, M.; Sun, J.; Liu, X. The biogeography of fungal communities in wetland sediments along the Changjiang River and other sites in China. ISME J. 2013, 7, 1299–1309. [Google Scholar] [CrossRef] [Green Version]
- Delgado, E.F.; Valdez, A.T.; Covarrubias, S.A.; Tosi, S.; Nicola, L. Soil Fungal Diversity of the Aguarongo Andean Forest (Ecuador). Biology 2021, 10, 1289. [Google Scholar] [CrossRef]
- Ozimek, E.; Hanaka, A. Mortierella species as the plant growth-promoting fungi present in the agricultural soils. Agriculture 2020, 11, 7. [Google Scholar] [CrossRef]
- Ning, Q.; Chen, L.; Jia, Z.; Zhang, C.; Ma, D.; Li, F.; Zhang, J.; Li, D.; Han, X.; Cai, Z.; et al. Multiple long-term observations reveal a strategy for soil pH-dependent fertilization and fungal communities in support of agricultural production. Agric Ecosyst. Environ. 2020, 293, 106837. [Google Scholar] [CrossRef]
- Lindahl, B.D.; Ihrmark, K.; Boberg, J.; Trumbore, S.E.; Högberg, P.; Stenlid, J.; Finlay, R.D. Spatial separation of litter decomposition and mycorrhizal nitrogen uptake in a boreal forest. New Phytol. 2007, 173, 611–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mäkipää, R.; Rajala, T.; Schigel, D.; Rinne, K.T.; Pennanen, T.; Abrego, N.; Ovaskainen, O. Interactions between soil- and dead wood-inhabiting fungal communities during the decay of Norway spruce logs. ISME J. 2017, 11, 1964–1974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gawas-Sakhalkar, P.; Singh, S.M. Fungal community associated with Arctic moss, Tetraplodon mimoides and its rhizosphere: Bioprospecting for production of industrially useful enzymes. Curr. Sci. 2011, 100, 1701–1705. [Google Scholar]
- Varnaite, R.; Paškevicius, A.; Raudoniene, V. Cellulose degradation in rye straw by micromycetes and their complexes. Ekologija 2008, 54, 29–31. [Google Scholar] [CrossRef]
- Lisov, A.; Belova, O.; Zavarzina, A.; Konstantinov, A.; Leontievsky, A. The Role of Laccase from Zygomycetous Fungus Mortierella elasson in Humic Acids Degradation. Agronomy 2021, 11, 2169. [Google Scholar] [CrossRef]
- Ellegaard-Jensen, L.; Aamand, J.; Kragelund, B.B.; Johnsen, A.H.; Rosendahl, S. Strains of the soil fungus Mortierella show different degradation potentials for the phenylurea herbicide diuron. Biogeochemistry 2013, 24, 765–774. [Google Scholar] [CrossRef]
- Nakagawa, A.; Osawa, S.; Hirata, T.; Yamagishi, Y.; Hosoda, J.; Horikoshi, T. 2, 4-Dichlorophenol degradation by the soil fungus Mortierella sp. Biosci. Biotechnol. Biochem. 2006, 70, 525–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Chen, L.; Redmile-Gordon, M.; Zhang, J.; Zhang, C.; Ning, Q.; Li, W. Mortierella elongata’s roles in organic agriculture and crop growth promotion in a mineral soil. Land Degrad. Dev. 2018, 29, 1642–1651. [Google Scholar] [CrossRef]
- Kataoka, R.; Takagi, K.; Sakakibara, F. A new endosulfan-degrading fungus, Mortierella species, isolated from a soil contaminated with organochlorine pesticides. J. Pestic. Sci. 2010, 35, 326–332. [Google Scholar] [CrossRef] [Green Version]
- Ceci, A.; Pinzari, F.; Russo, F.; Maggi, O.; Persiani, A.M. Saprotrophic soil fungi to improve phosphorus solubilisation and release: In vitro abilities of several species. Ambio 2018, 47, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Wani, Z.A.; Kumar, A.; Sultan, P.; Bindu, K.; Riyaz-Ul Hassan, S.; Ashraf, N. Mortierella alpina CS10E4, an oleaginous fungal endophyte of Crocus sativus L. enhances apocarotenoid biosynthesis and stress tolerance in the host plant. Sci. Rep. 2017, 7, 8598. [Google Scholar] [CrossRef] [Green Version]
- El Hajj Assaf, C.; Zetina-Serrano, C.; Tahtah, N.; Khoury, A.E.; Atoui, A.; Oswald, I.P.; Puel, O.; Lorber, S. Regulation of secondary metabolism in the penicillium genus. Int. J. Mol. Sci. 2020, 21, 9462. [Google Scholar] [CrossRef]
- Matei, G.M.; Matei, S.; Mocanu, V. Assessing the role of soil microbial communities of natural forest ecosystem. EuroBiotech J. 2020, 4, 01–07. [Google Scholar] [CrossRef] [Green Version]
- Tosi, S.; Baretta, F.; Savini, D.; Sartori, F. Analisi sul microbiota del suolo lungo un gradiente di profondità nella riserva integrale Siro Negri (PV, Italy). Arch. Geobot. 2005, 11, 39–48. [Google Scholar]
- Breitenbach, J.; Kränzlin, F. Fungi of Switzerland; Ascomycetes. Edition Mycologia: Lucerne, Switzerland, 1984; Volume 1. [Google Scholar]
- Tedersoo, L.; Hansen, K.; Perry, B.A.; Kjøller, R. Molecular and morphological diversity of pezizalean ectomycorrhiza. New Phytol. 2006, 170, 581–596. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Dong, C.; Chen, W.; Mou, Q.; Lu, X.; Han, Y.; Huang, J.; Liang, Z. The enigmatic Thelebolaceae (Thelebolales, Leotiomycetes): One new genus Solomyces and five new species. Front. Microb. 2020, 11, 2618. [Google Scholar] [CrossRef] [PubMed]
- Garzoli, L.; Bozzetta, E.; Varello, K.; Cappelleri, A.; Patriarca, E.; Debernardi, P.; Riccucci, M.; Boggero, A.; Girometta, C.; Picco, A.M. White-Nose Syndrome Confirmed in Italy: A Preliminary Assessment of Its Occurrence in Bat Species. J. Fungus 2021, 7, 192. [Google Scholar] [CrossRef] [PubMed]
- Calabon, M.S.; Hyde, K.D.; Jones, E.B.; Luo, Z.L.; Dong, W.; Hurdeal, V.G.; Gentekaki, E.; Rossi, W.; Leonardi, M.; Thiyagaraja, V.; et al. Freshwater fungal numbers. Fungal Divers. 2022, 114, 3–235. [Google Scholar] [CrossRef]
- Hibbett, D.S. A phylogenetic overview of the Agaricomycotina. Mycologia 2006, 98, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Hibbett, D.S.; Bauer, R.; Binder, M.; Giachini, A.J.; Hosaka, K.; Justo, A.; Larsson, E.; Larsson, K.H.; Lawrey, J.D.; Miettinen, O.; et al. 14 Agaricomycetes. In Systematics and Evolution; Springer: Berlin, Heidelberg, 2014; pp. 373–429. [Google Scholar]
- He, M.-Q.; Zhao, R.-L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.C.; Raspé, O.; Kakishima, M.; Sánchez-Ramírez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar] [CrossRef] [Green Version]
- Bernicchia, A.; Gorjón, S.P. Fungi Europaei 12: Corticiaceae sl; Edizioni Candusso: Alassio, Italy, 2010; pp. 731–744. [Google Scholar]
- Della Rocca, F.; Stefanelli, S.; Pasquaretta, C.; Campanaro, A.; Bogliani, G. Effect of deadwood management on saproxylic beetle richness in the floodplain forests of northern Italy: Some measures for deadwood sustainable use. J. Insect Conserv. 2014, 18, 121–136. [Google Scholar] [CrossRef]
- Girometta, C.E.; Bernicchia, A.; Baiguera, R.M.; Bracco, F.; Buratti, S.; Cartabia, M.; Picco, A.M.; Savino, E. An italian research culture collection of wood decay fungi. Diversity 2020, 12, 58. [Google Scholar] [CrossRef] [Green Version]
- Brusa, G.; Gheza, G.; Bracco, F.; Assini, S. Bryophytes from a residual oak-elm forest: The case study of the “Bosco Siro Negri” reserve in the Ticino River valley (N Italy). Borziana 2021, 2, 125–138. [Google Scholar] [CrossRef]
- Wagner, L.; Stielow, B.; Hoffmann, K.; Petkovits, T.; Papp, T.; Vágvölgyi, C.; Voigt, K. A comprehensive molecular phylogeny of the Mortierellales (Mortierellomycotina) based on nuclear ribosomal DNA. Pers. Mol. Phylogeny Evol. Fungi 2013, 30, 77–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grantina-Ievina, L.; Kasparinskis, R.; Tabors, G.; Nikolajeva, V. Features of saprophytic soil microorganism communities in conifer stands with or without Heterobasidion annosum sensu lato infection: A special emphasis on Penicillium spp. Environ. Exp. Biol. 2013, 11, 23–28. [Google Scholar]
- Nguyen, T.T.; Park, S.W.; Pangging, M.; Lee, H.B. Molecular and morphological confirmation of three undescribed species of Mortierella from Korea. Mycobiology 2019, 47, 31–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanchette, R.A. A review of microbial deterioration found in archaeological wood from different environments. Int. Biodeterior. Biodegrad. 2000, 46, 189–204. [Google Scholar] [CrossRef]
- Vieyra, H.; San Martín-Martínez, E.; Juárez, E.; Figueroa-López, U.; Aguilar-Méndez, M.A. Biodegradation process of a blend of thermoplastic unripe banana flour—Polyethylene under composting: Identification of the biodegrading agent. J. Appl. Polym. Sci. 2015, 132, 42258. [Google Scholar] [CrossRef]
- Abbas, H.K.; Ocamb, C.M. First report of production of fumonisin B-1 by Fusarium polyphialidicum collected from seeds of Pinus strobus. Plant Dis. 1995, 79, 642. [Google Scholar] [CrossRef]
- Aoki, T.; O’Donnell, K.; Geiser, D.M. Systematics of key phytopathogenic Fusarium species: Current status and future challenges. J. Gen. Plant Pathol. 2014, 80, 189–201. [Google Scholar] [CrossRef]
- Boccardo, F.; Traverso, M.; Vizzini, A.; Zotti, M. Funghi d’Italia; Zanichelli: Bologna, Italy, 2008. [Google Scholar]
- Grantina-Ievina, L.; Berzina, A.; Nikolajeva, V.; Mekss, P.; Muiznieks, I. Production of fatty acids by Mortierella and Umbelopsis species isolated from temperate climate soils. Environ. Exp. Biol. 2014, 12, 15–27. [Google Scholar]
- Purahong, W.; Wubet, T.; Lentendu, G.; Schloter, M.; Pecyna, M.J.; Kapturska, D.; Hofrichter, M.; Krüger, D.; Buscot, F. Life in leaf litter: Novel insights into community dynamics of bacteria and fungi during litter decomposition. Mol. Ecol. 2016, 25, 4059–4074. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
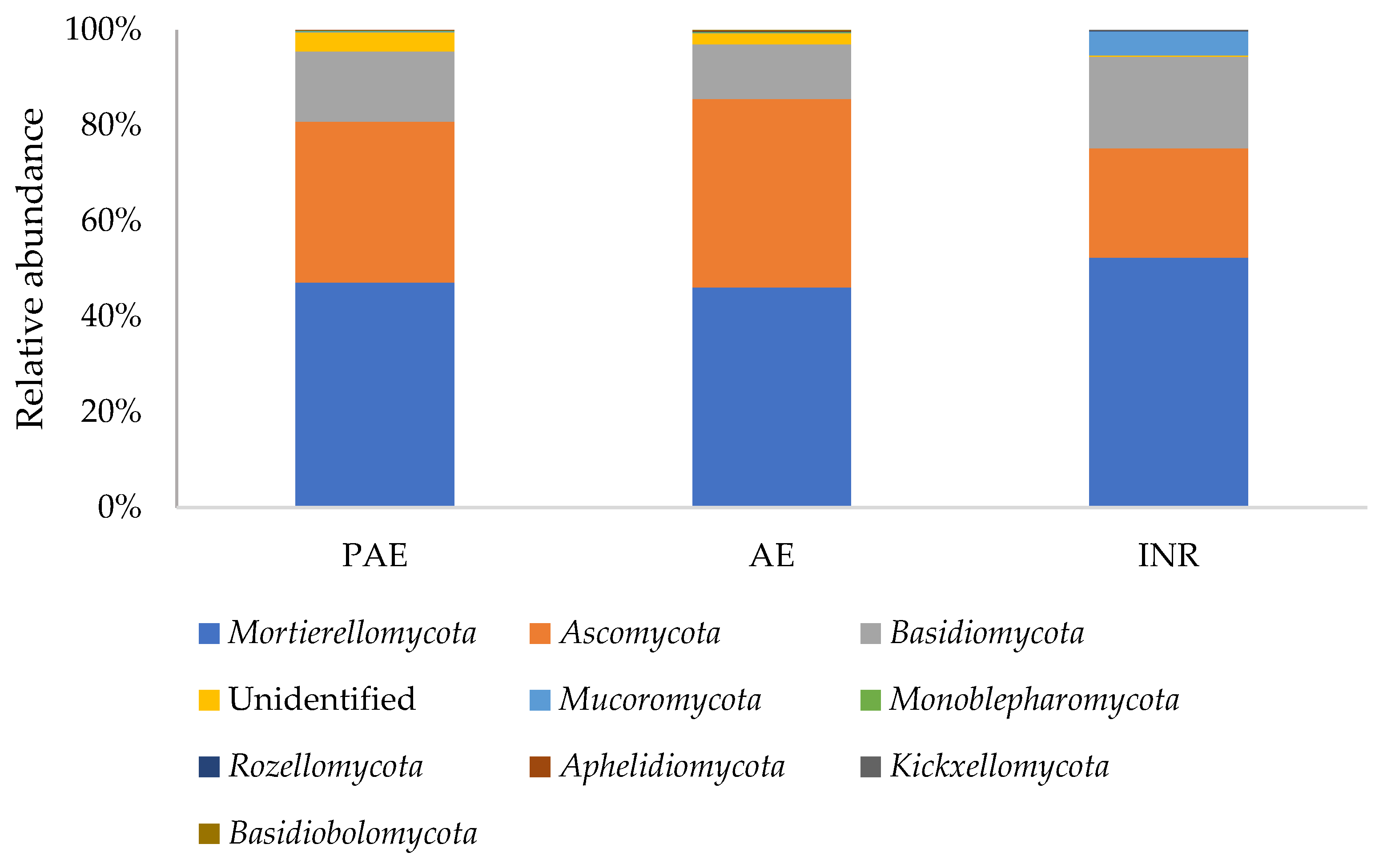{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microplastics | Macroplastics | Total Plastic Amount | |
|---|---|---|---|
| PAE | 13,200 ± 2466 a | 5202 ± 230 a | 18,402 ± 2696 a |
| AE | 160 ± 33 b | 9 ± 11 b | 169 ± 44 b |
| INR | 60 ± 52 b | 0 ± 0 b | 60 ± 52 b |
| Site | pH | SOC | Organic Matter | TN | C/N | P | Ca | Mg | K |
|---|---|---|---|---|---|---|---|---|---|
| % | % | g/kg | mg/kg | mg/kg | mg/kg | mg/kg | |||
| PAE | 5.3 ± 0.0 ab | 3.1 ± 0.1 ab | 5.3 ± 0.2 ab | 2.97 ± 0.16 ab | 10.4 ± 0.2 ab | 24.0 ± 0.0 ab | 1423 ± 141 a | 137.6 ± 10.8 ab | 96.8 ± 10.2 a |
| AE | 6.0 ± 0.0 a | 1.4 ± 0.04 a | 2.5 ± 0.01 a | 1.43 ± 0.03 a | 10.1 ± 0.1 a | 30.0 ± 0.3 a | 857 ± 45 b | 143.7 ± 3.2 a | 272.5 ± 26.3 b |
| INR | 4.4 ± 0.0 b | 4.7 ± 0.1 b | 8.1 ± 0.2 b | 3.64 ± 0.08 b | 12.9 ± 0.4 b | 15.0 ± 0.5 b | 1041 ± 70 ab | 101.6 ± 10.0 b | 104.0 ± 16.1 ab |
| MEA | Humic Acids | Lignocellulose | |
|---|---|---|---|
| PAE | 5.0 × 105 a | 3.4 × 105 a | 3.7 × 105 a |
| AE | 5.2 × 105 a | 2.3 × 105 b | 1.5 × 105 b |
| INR | 5.8 × 105 b | 2.0 × 105 b | 1.8 × 105 b |
| Observed Species | Shannon | Simpson | |
|---|---|---|---|
| PAE | 173.25 ± 38.78 | 3.98 ± 0.49 | 0.96 ± 0.02 |
| AE | 177.00 ± 73.82 | 3.91 ± 0.90 | 0.95 ± 0.04 |
| INR | 122.75 ± 9.11 | 3.29 ± 0.21 | 0.92 ± 0.03 |
| Fungal Species | Relative Abundance INR | Relative Abundance PAE | Relative Abundance AE |
|---|---|---|---|
| Podila humilis | 33.51% a | 0.00% b | 0.00% b |
| Phaeoclavulina decurrens | 5.82% a | 0.00% b | 0.00% b |
| Penicillium bilaiae | 4.70% a | 0.00% b | 0.00% b |
| Russula sp. | 2.06% a | 0.00% b | 0.00% b |
| Lepista flaccida | 1.08% a | 0.00% b | 0.00% b |
| Tolypocladium album | 0.78% a | 0.00% b | 0.00% b |
| Hydnotrya tulasnei | 0.60% a | 0.00% b | 0.00% b |
| Mortierella simplex | 0.58% a | 0.00% b | 0.00% b |
| Phaeoclavulina decurrens | 0.55% a | 0.00% b | 0.00% b |
| Pseudogymnoascus roseus | 0.54% a | 0.00% b | 0.00% b |
| Geomyces auratus | 0.52% a | 0.00% b | 0.00% b |
| Mortierella alpina | 0% a | 15.22% b | 9.43% b |
| Linnemannia elongata | 0% a | 5.44% b | 3.42% b |
| Fusarium concolor | 0% a | 2.47% b | 1.44% b |
| Calvatia cyathiformis | 0% a | 1.06% b | 0.57% b |
| Neocosmospora solani | 0% a | 1.03% b | 0.71% b |
| Magnaporthiopsis incrustans | 0% a | 0.95% b | 0.15% b |
| Setophoma terrestris | 0% a | 0.79% b | 0.98% b |
| Tetracladium sp. | 0% a | 0.75% b | 0.2% ab |
| Microdochium novae-zelandiae | 0% a | 0.72% b | 0.52% b |
| Humicola olivacea | 0% a | 0.72% b | 0.78% b |
| Phialocephala bamuru | 0% a | 0.66% b | 0.32% b |
| Arrhenia sp. | 0% a | 0.64% b | 0.18% b |
| Haplographium debellae-marengoi var. equinum | 0% a | 0.63% b | 0.8% b |
| Furcasterigmium furcatum | 0.02% a | 0.59% ab | 6.31% b |
| Geastrum morganii | 0.15% a | 0% b | 0.19% a |
| Dissophora globulifera | 0.14% a | 0.04% a | 0.00% b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Temporiti, M.E.E.; Nicola, L.; Girometta, C.E.; Roversi, A.; Daccò, C.; Tosi, S. The Analysis of the Mycobiota in Plastic Polluted Soil Reveals a Reduction in Metabolic Ability. J. Fungi 2022, 8, 1247. https://doi.org/10.3390/jof8121247
Temporiti MEE, Nicola L, Girometta CE, Roversi A, Daccò C, Tosi S. The Analysis of the Mycobiota in Plastic Polluted Soil Reveals a Reduction in Metabolic Ability. Journal of Fungi. 2022; 8(12):1247. https://doi.org/10.3390/jof8121247
Chicago/Turabian StyleTemporiti, Marta Elisabetta Eleonora, Lidia Nicola, Carolina Elena Girometta, Anna Roversi, Chiara Daccò, and Solveig Tosi. 2022. "The Analysis of the Mycobiota in Plastic Polluted Soil Reveals a Reduction in Metabolic Ability" Journal of Fungi 8, no. 12: 1247. https://doi.org/10.3390/jof8121247