
Geo-Distribution Patterns of Soil Fungal Community of Pennisetum flaccidum in Tibet


Abstract
:1. Introduction
2. Materials and Methods
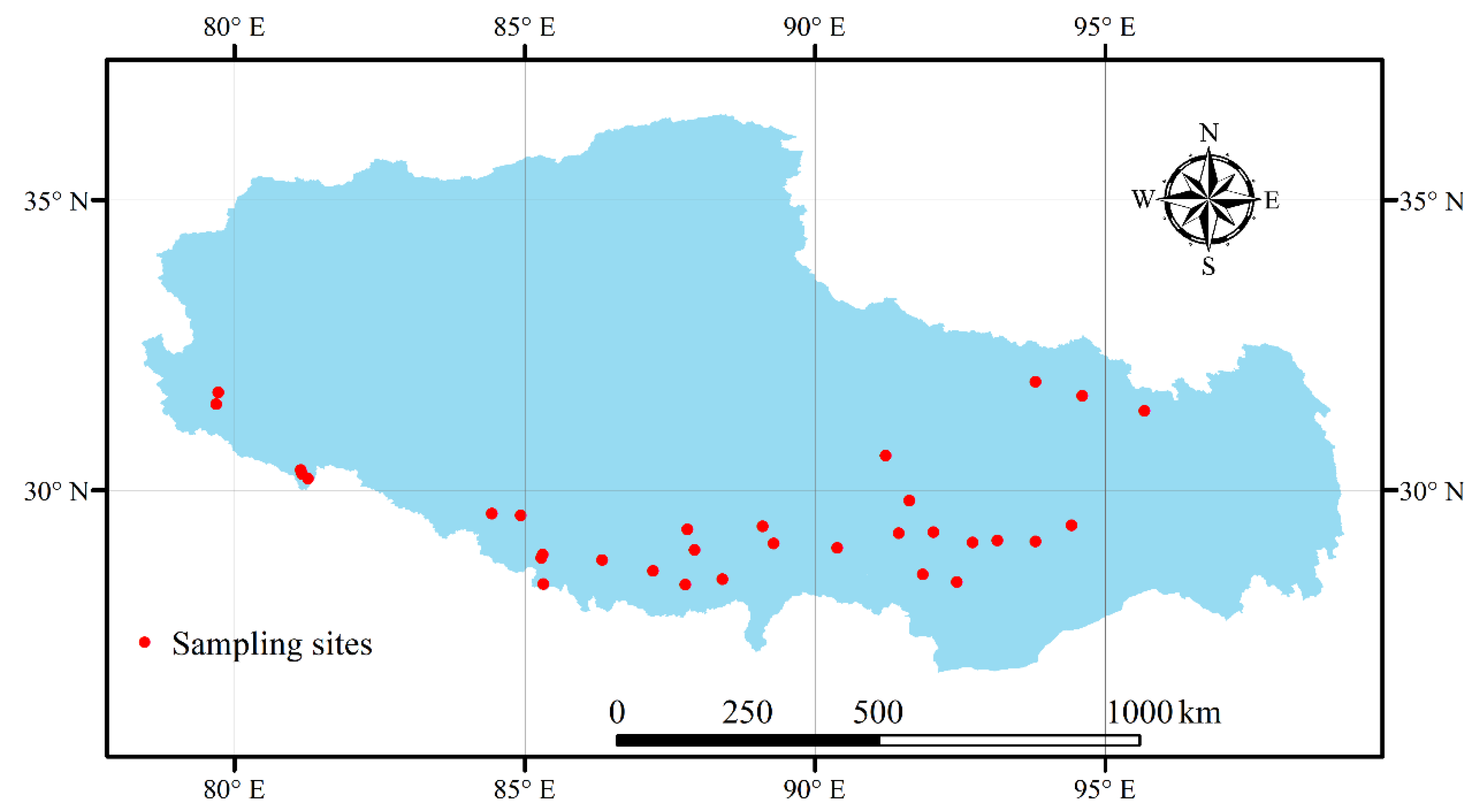
2.1. Study Area
2.2. Plant Sampling and Observation, Soil Sampling and Analyses
2.3. Statistical Analyses
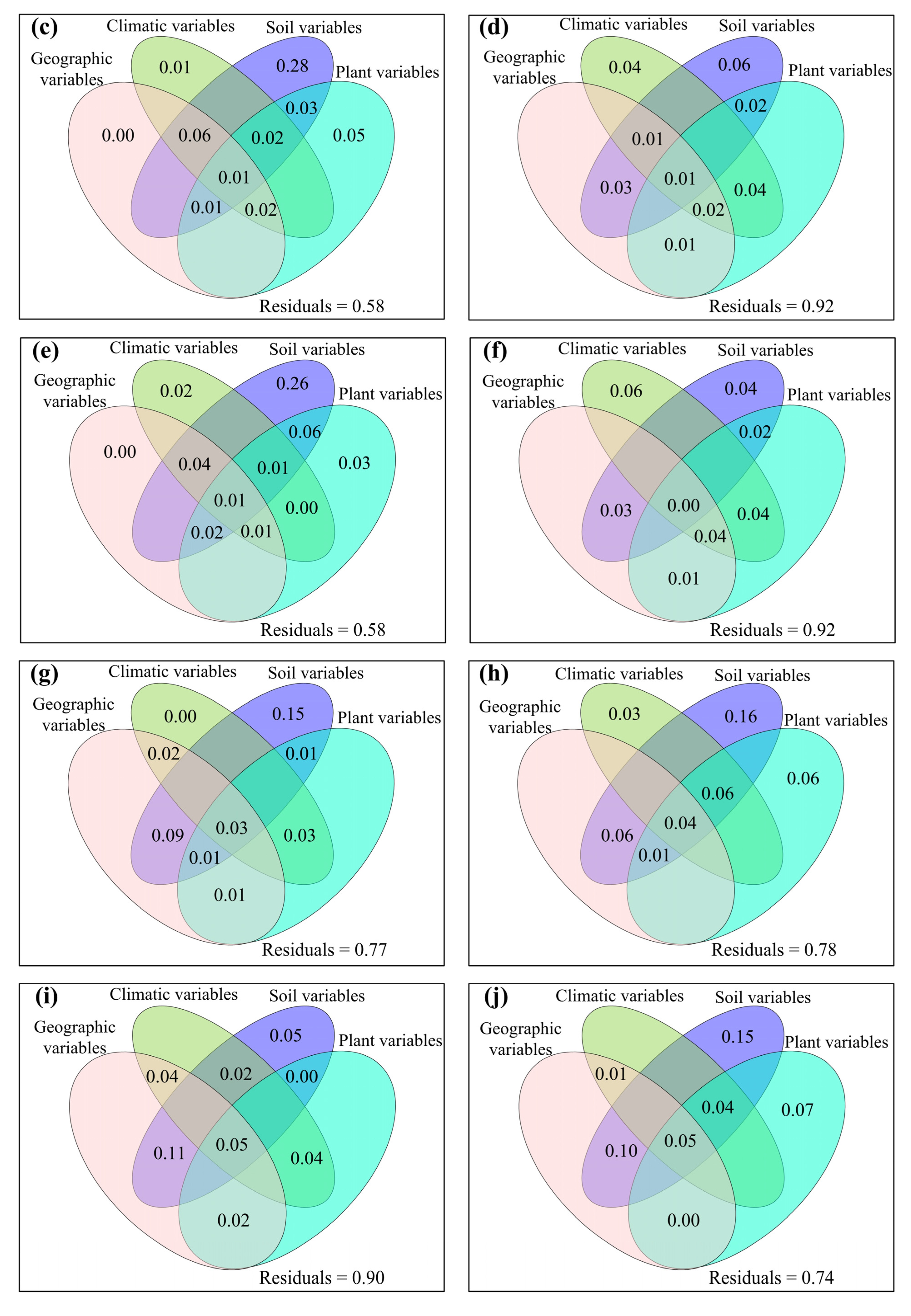
3. Results
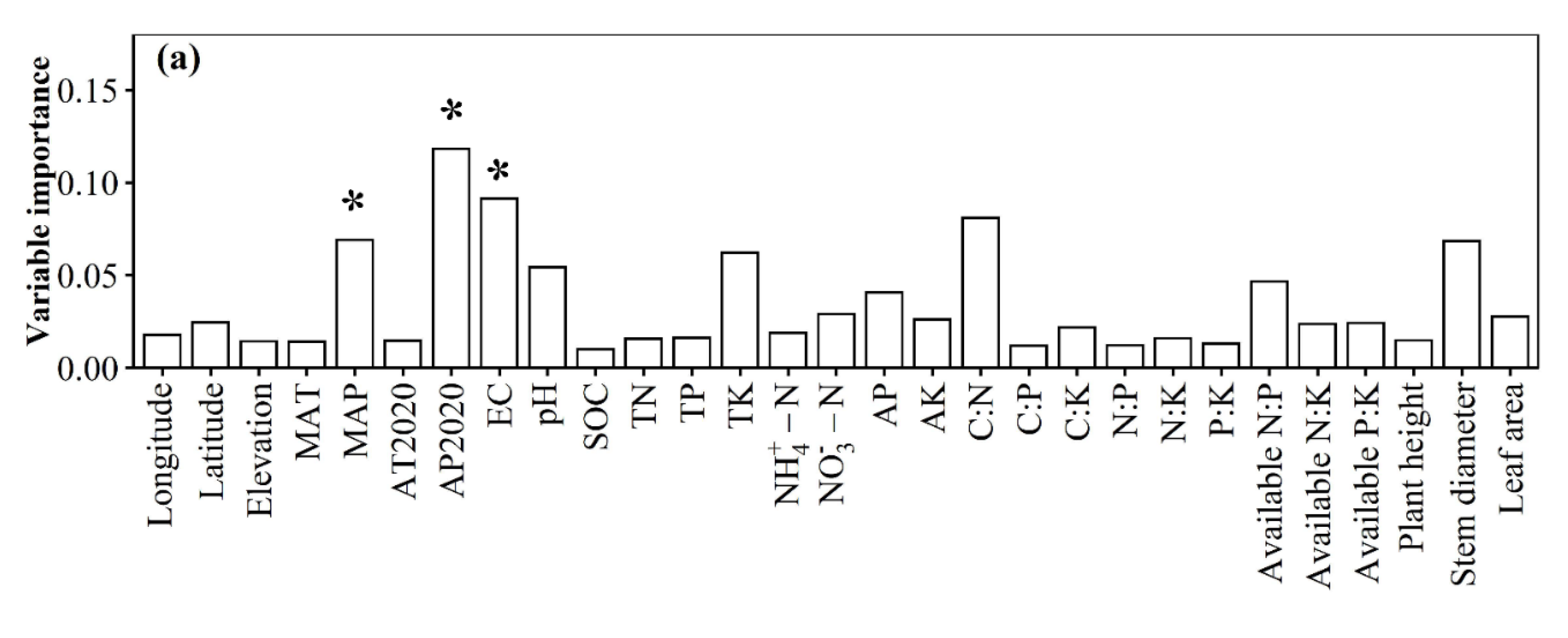
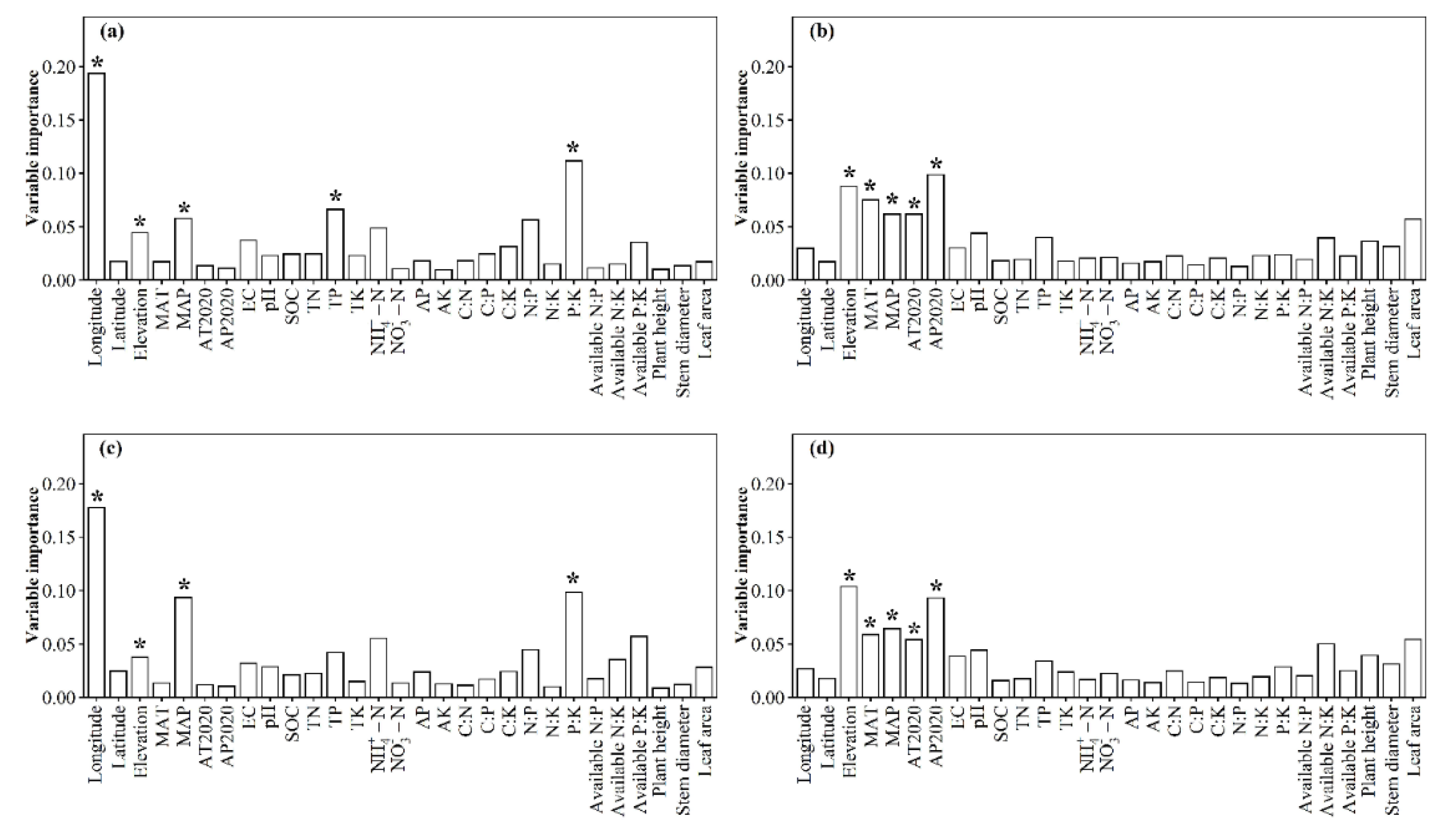
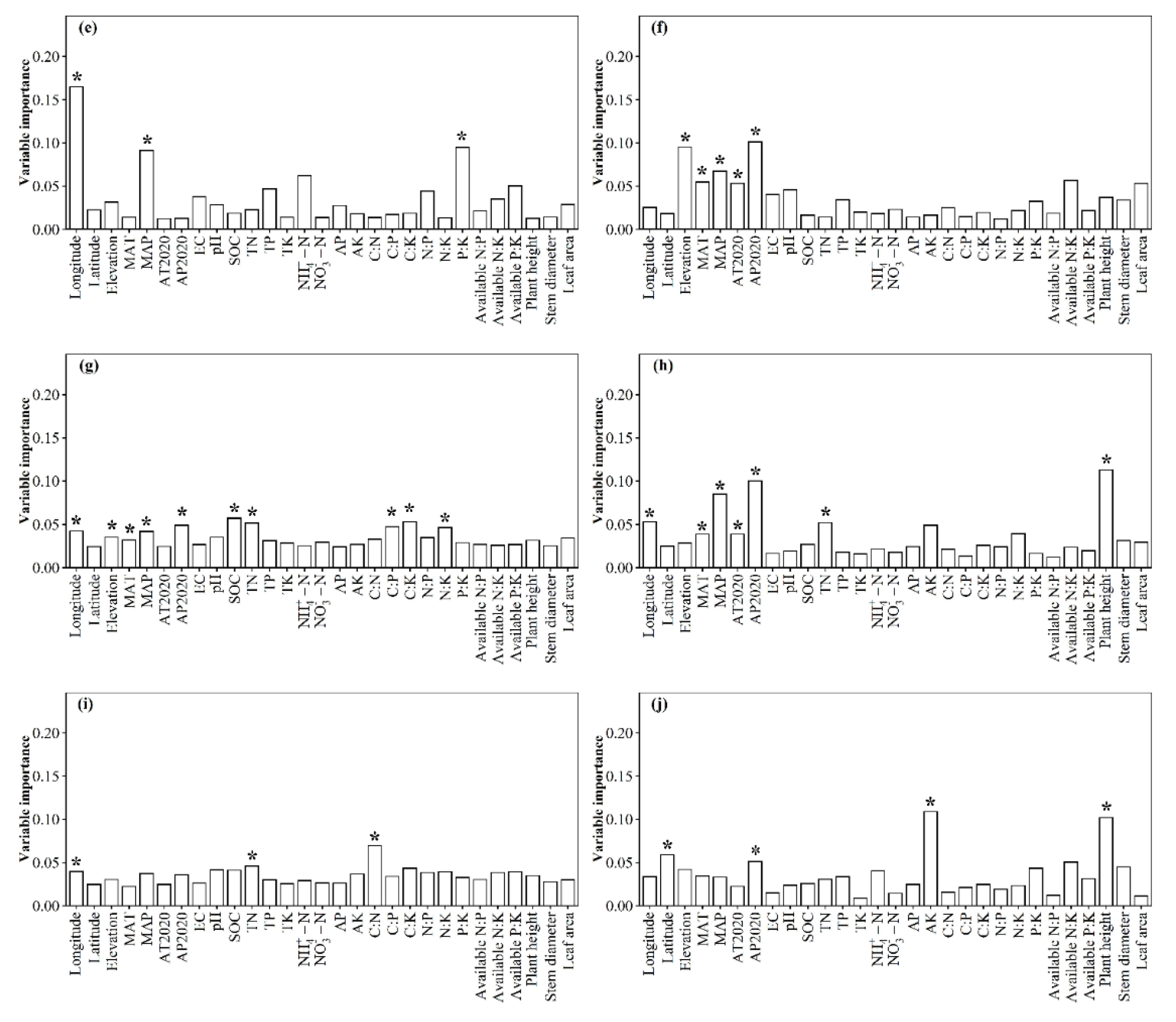
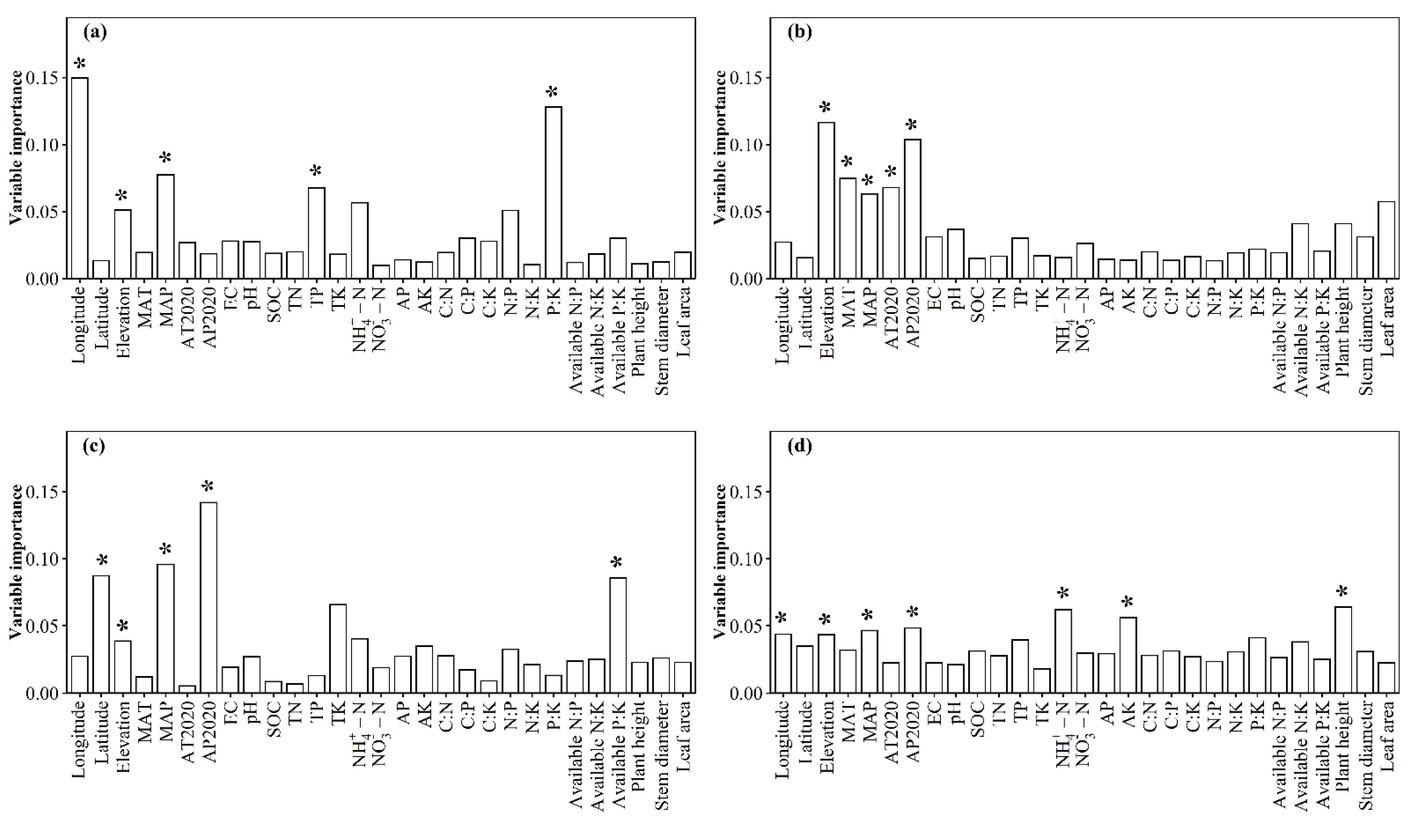
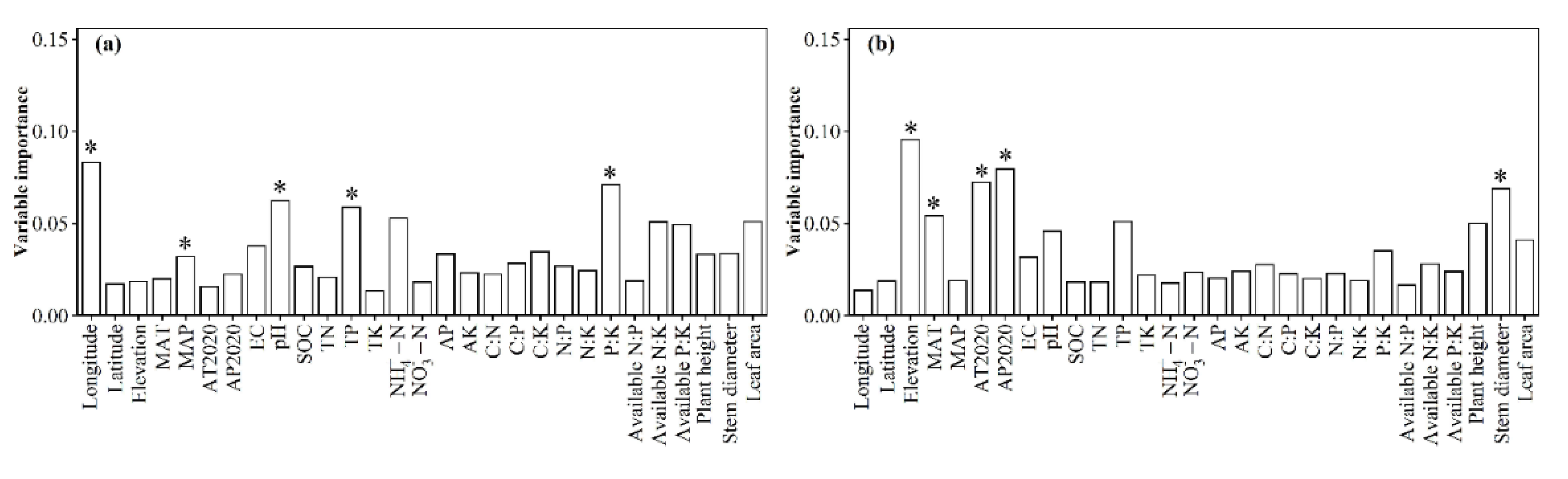
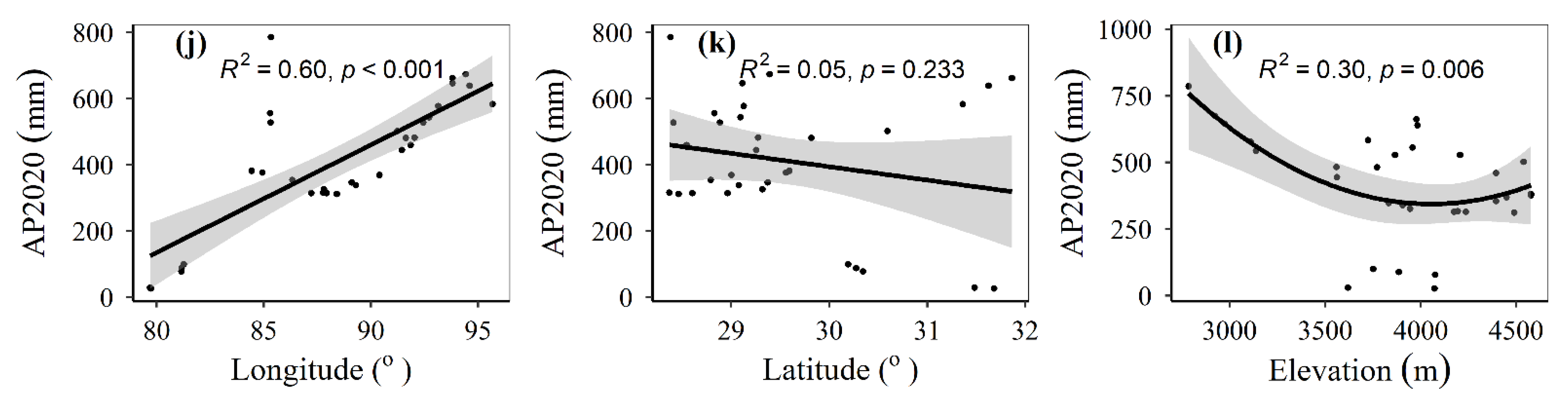
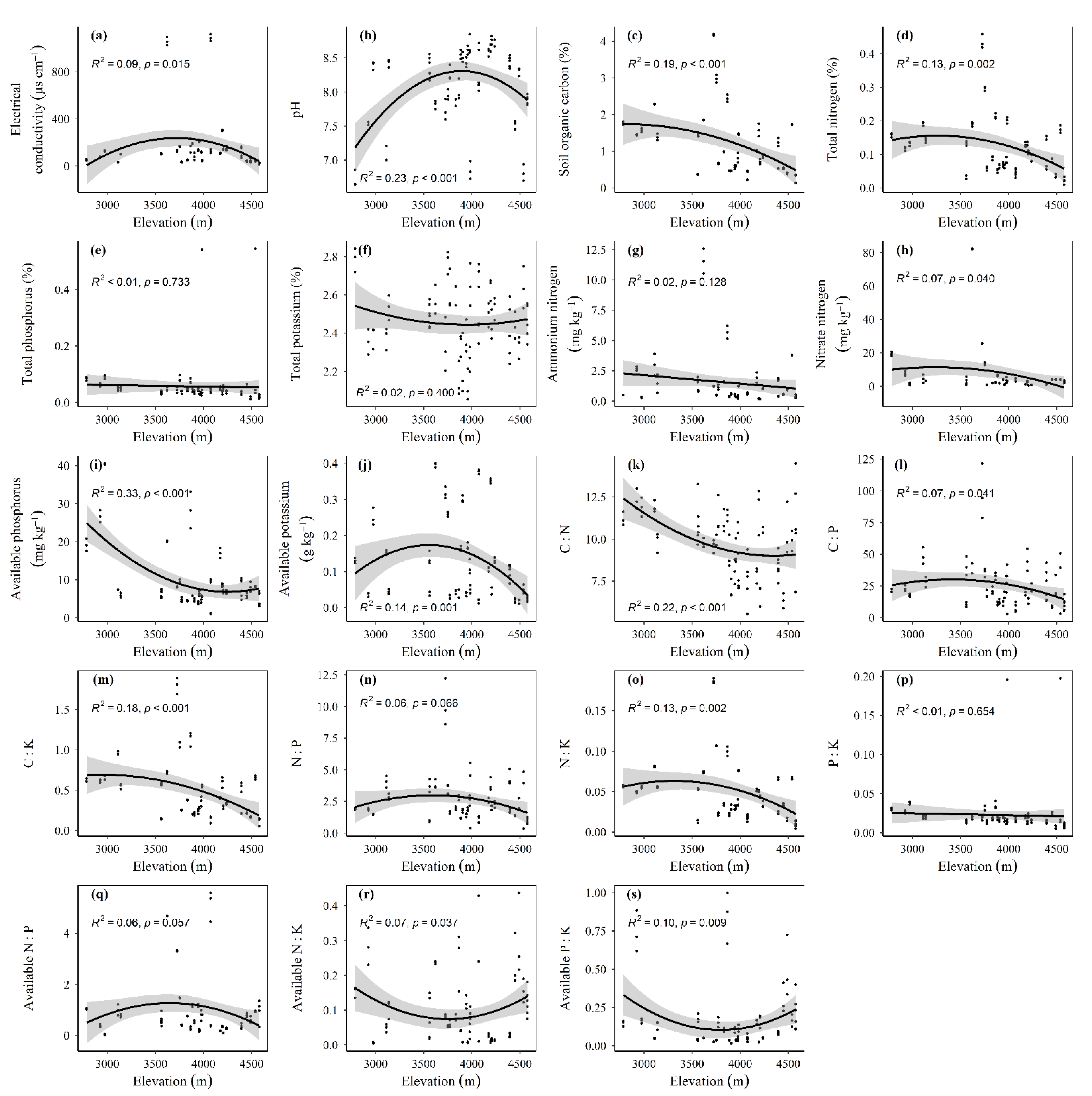
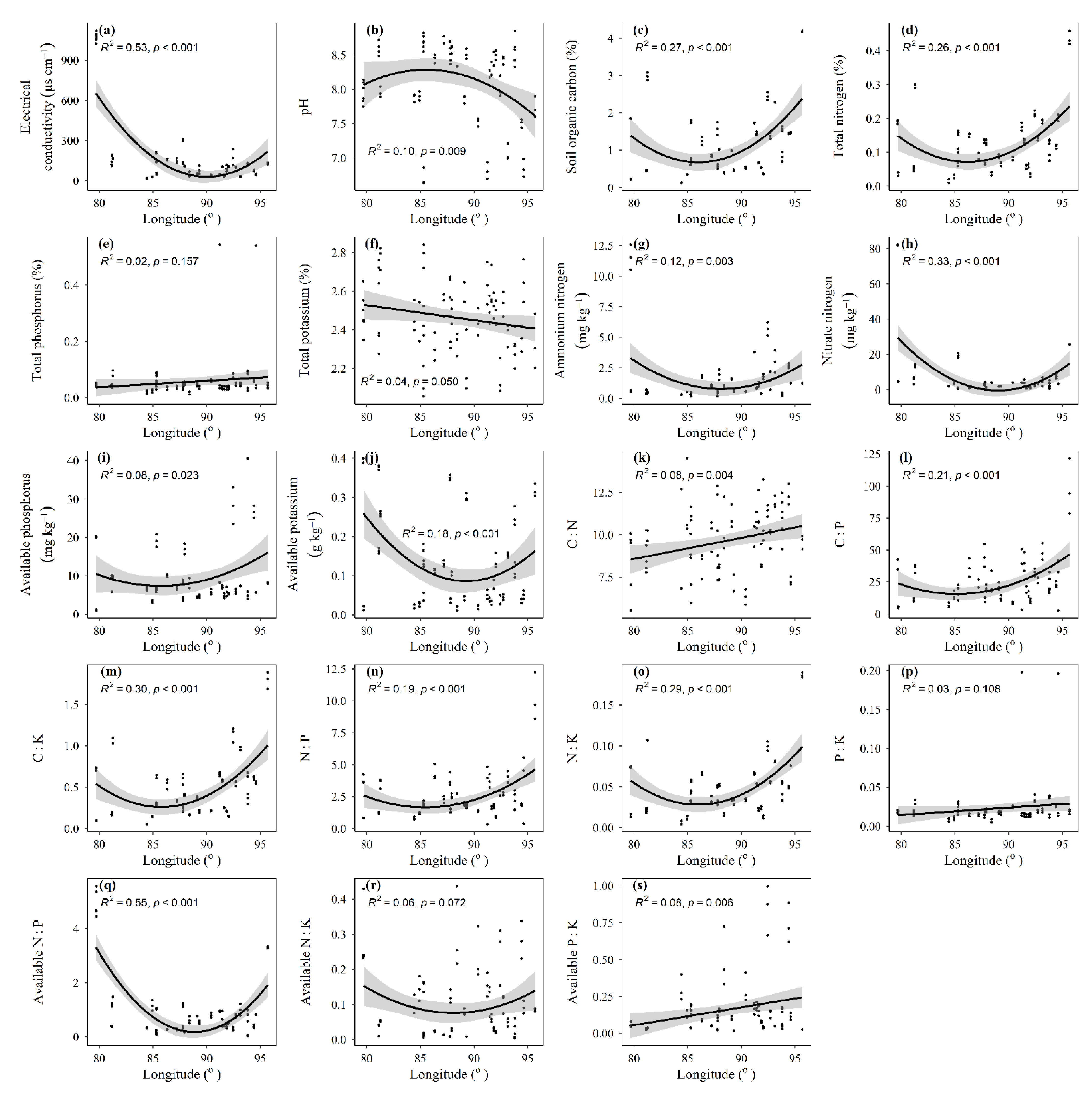
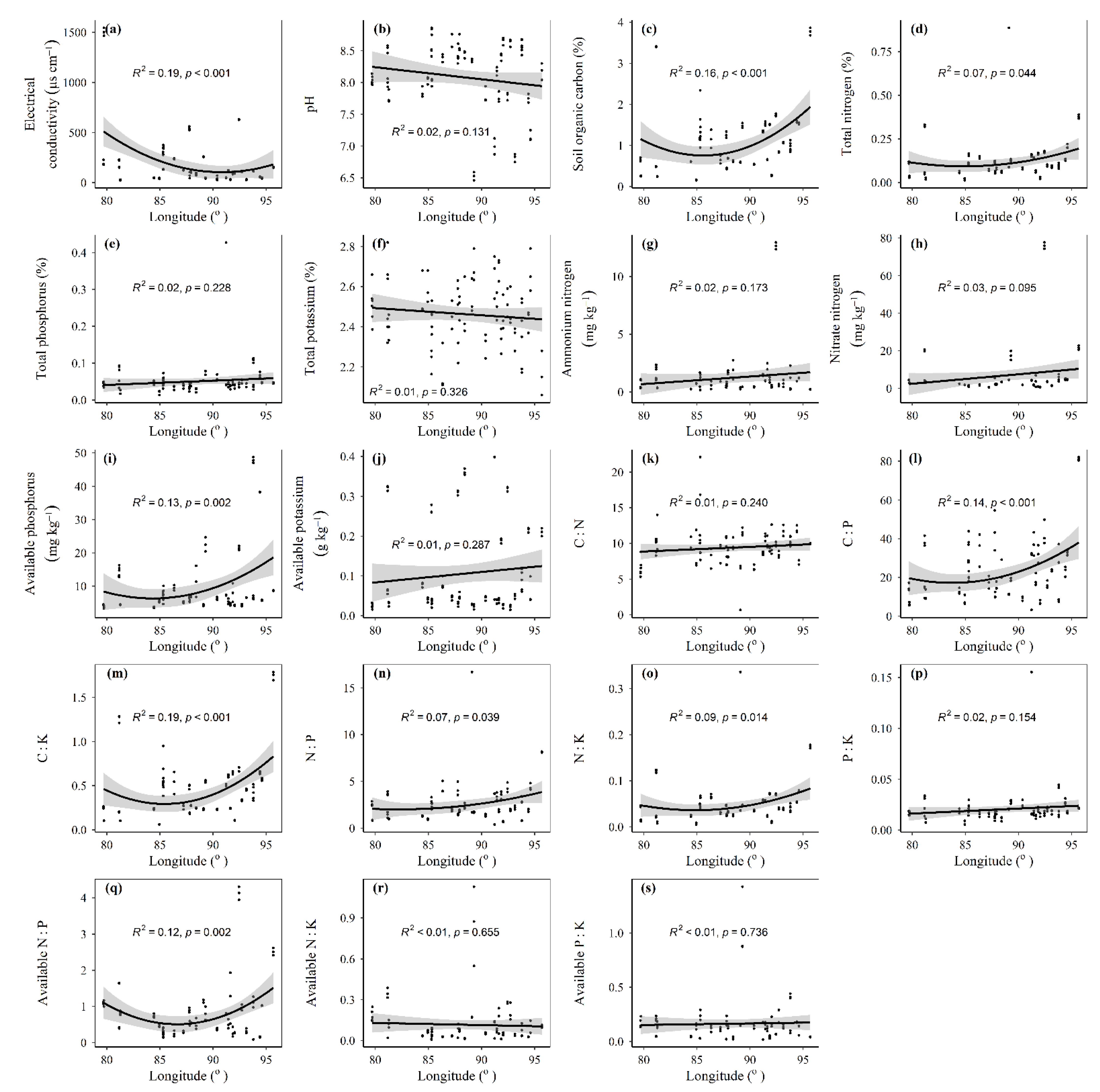
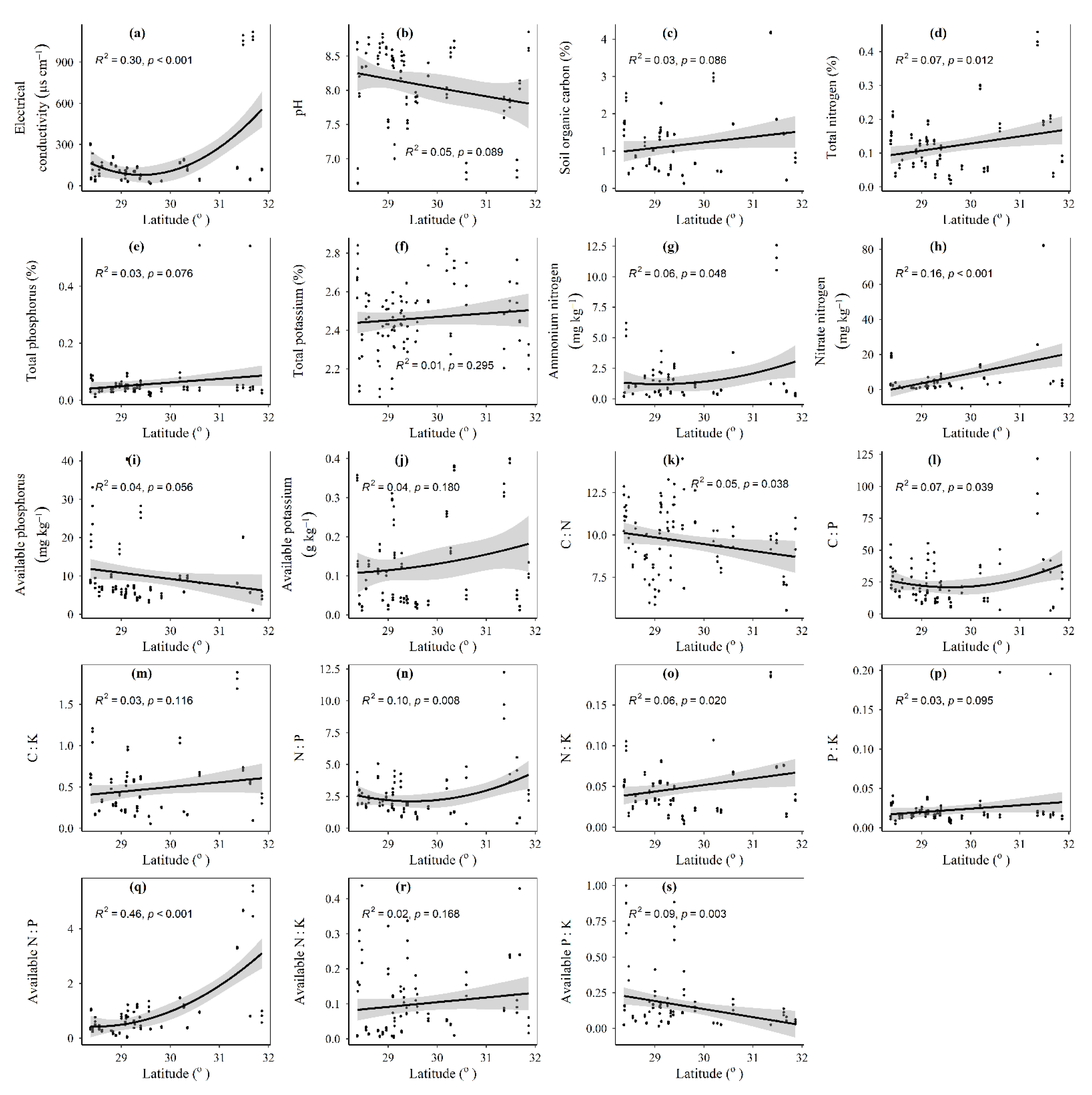
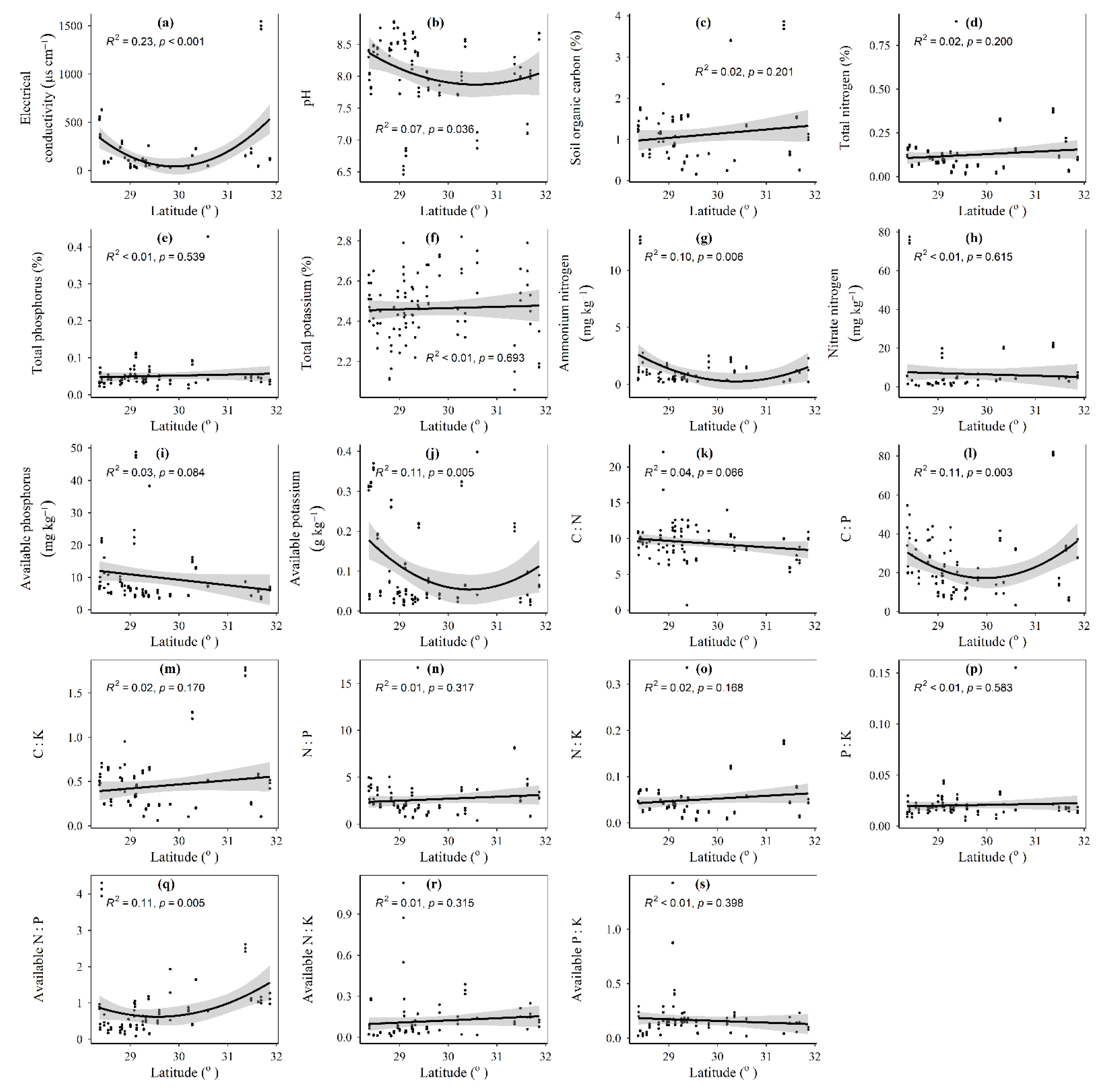
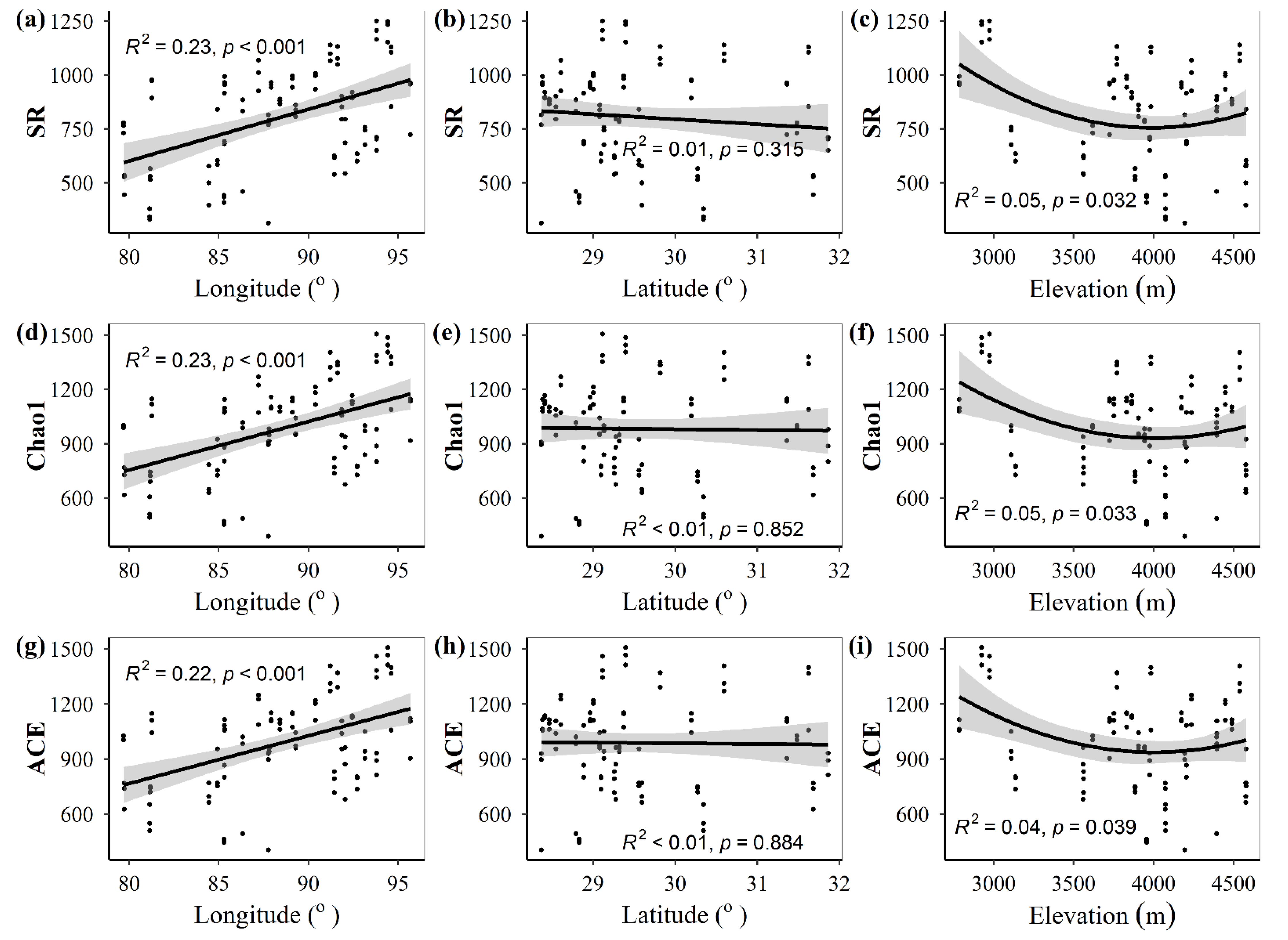
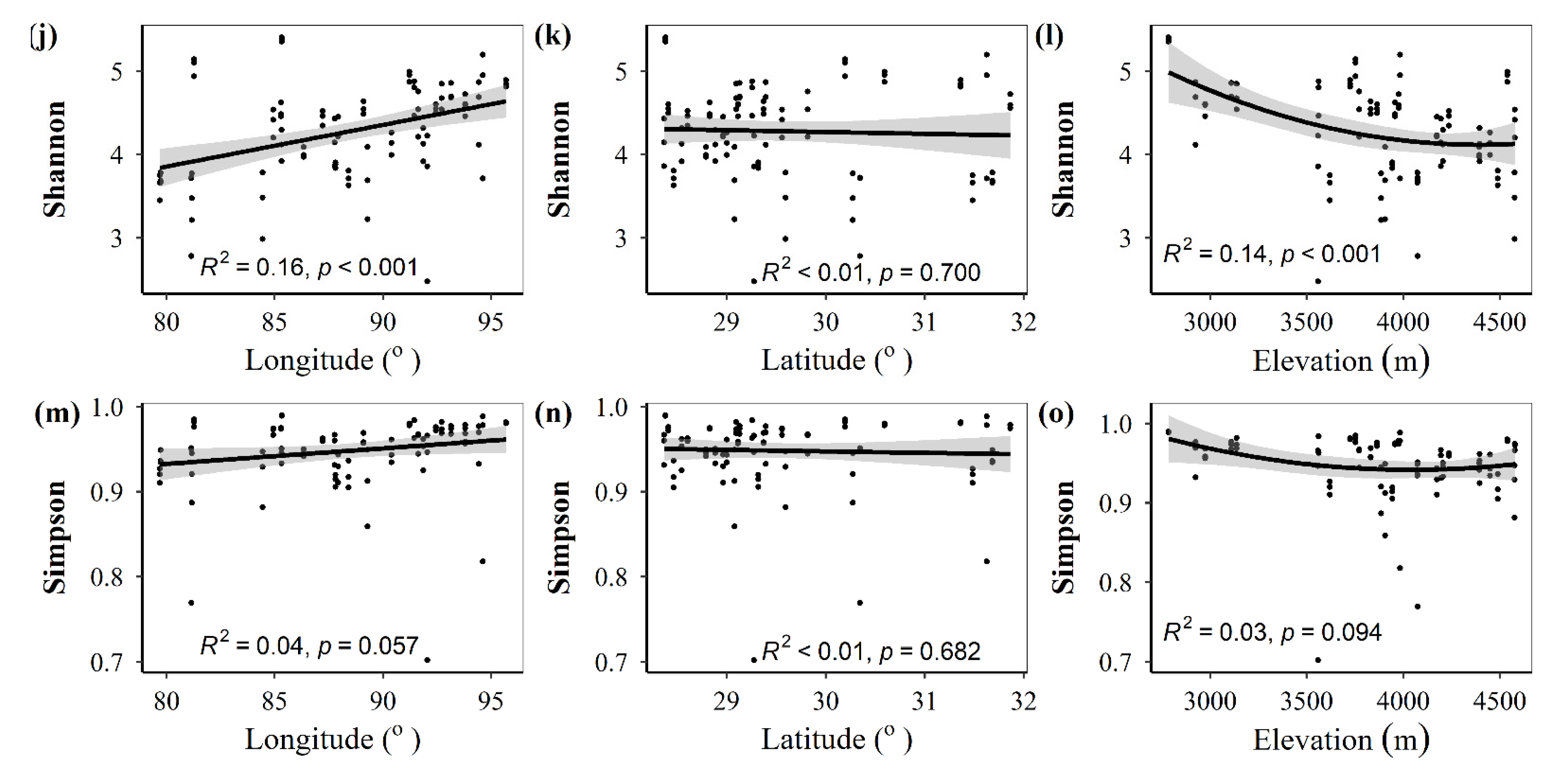
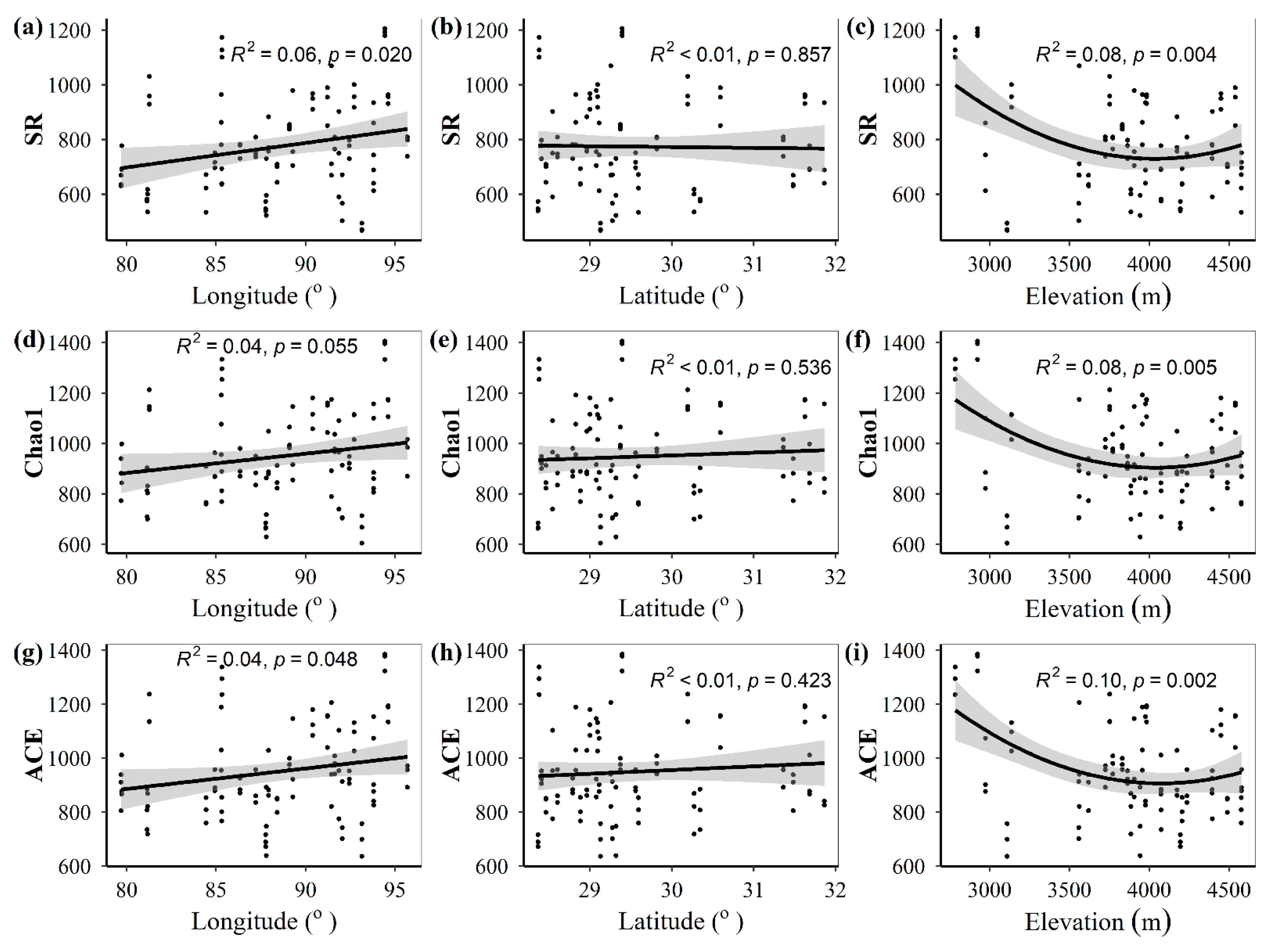
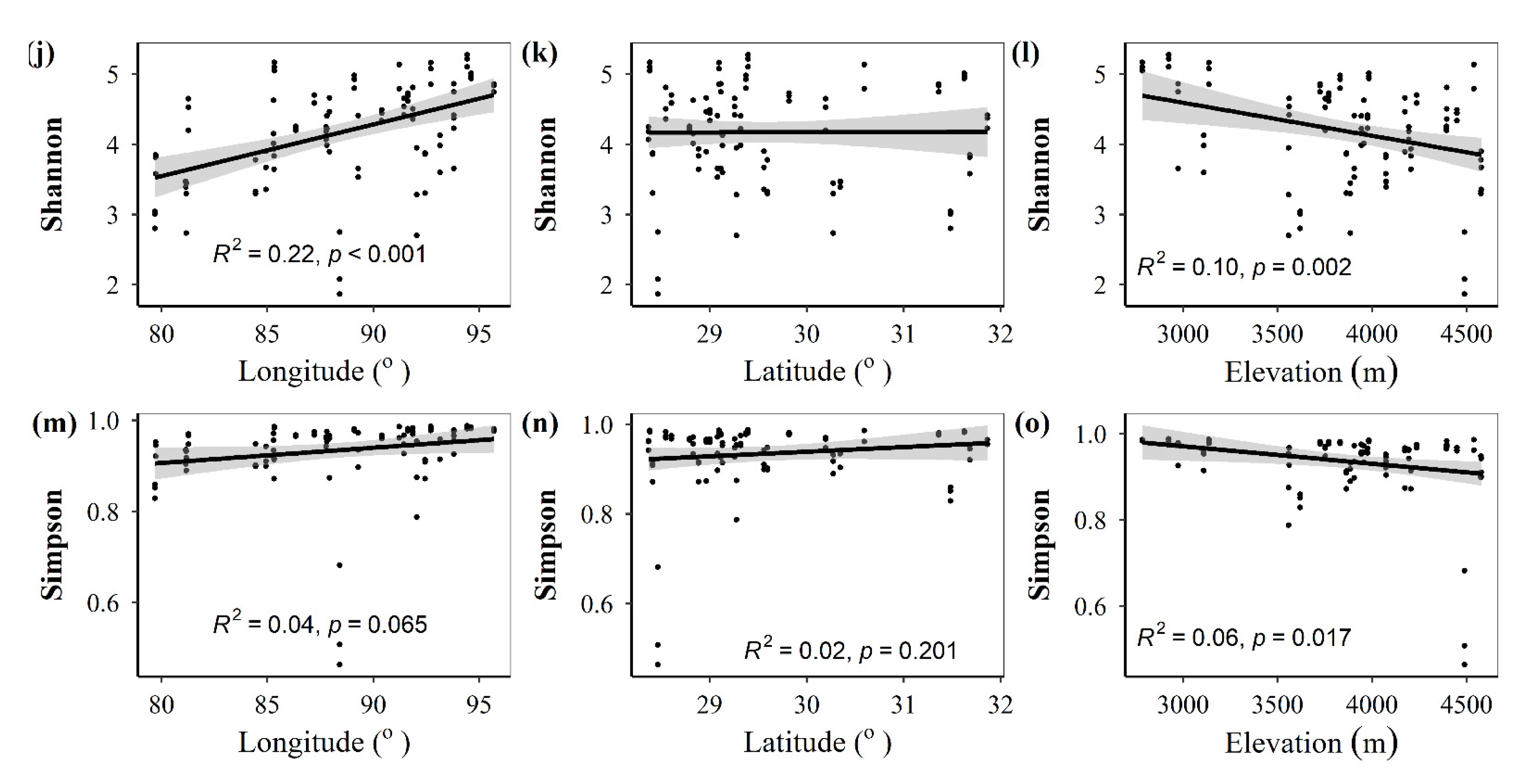
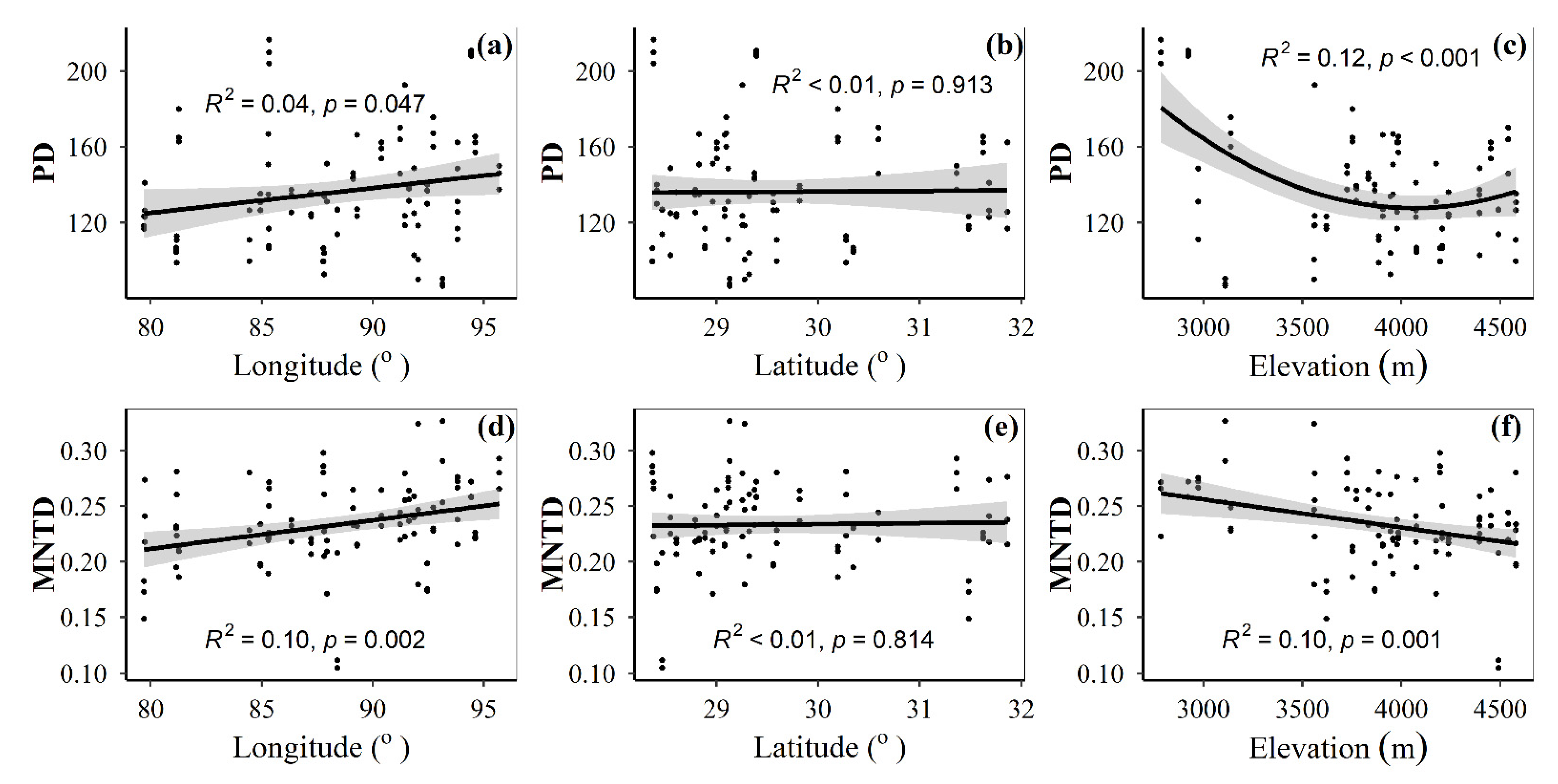
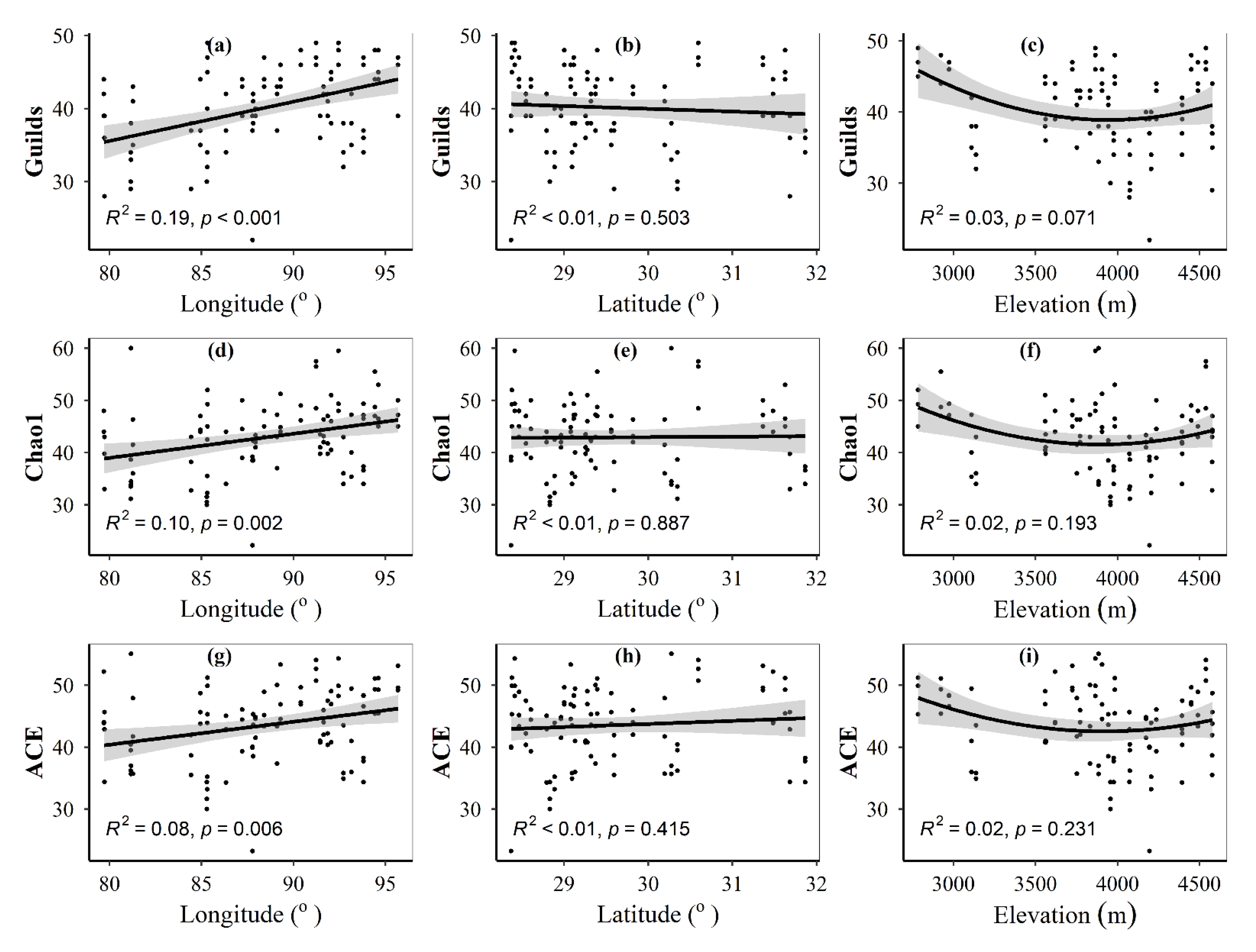
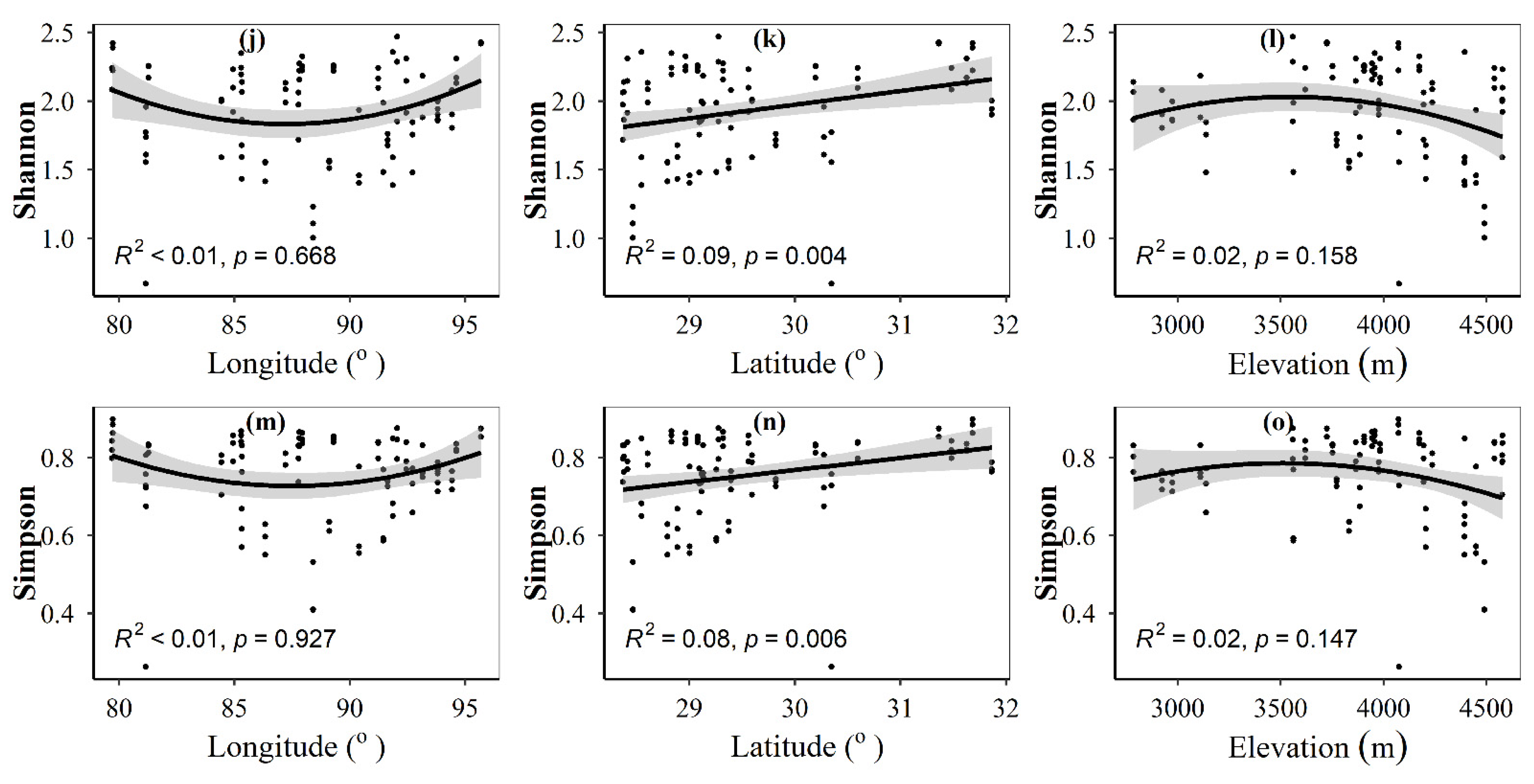
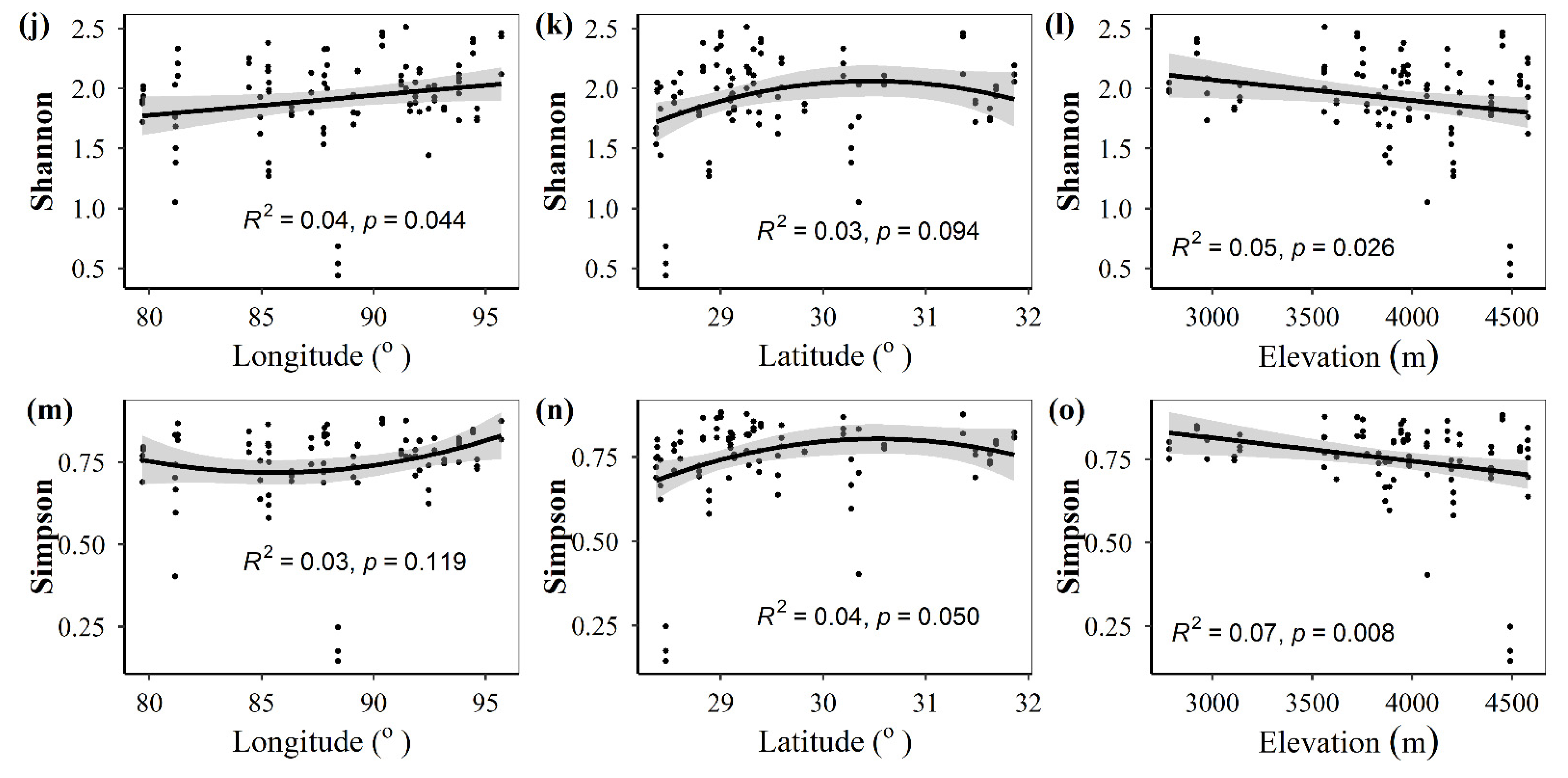
3.1. Spatial Distributions of Environmental Variables, and Total Abundance and Diversity of Soil Fungal Community
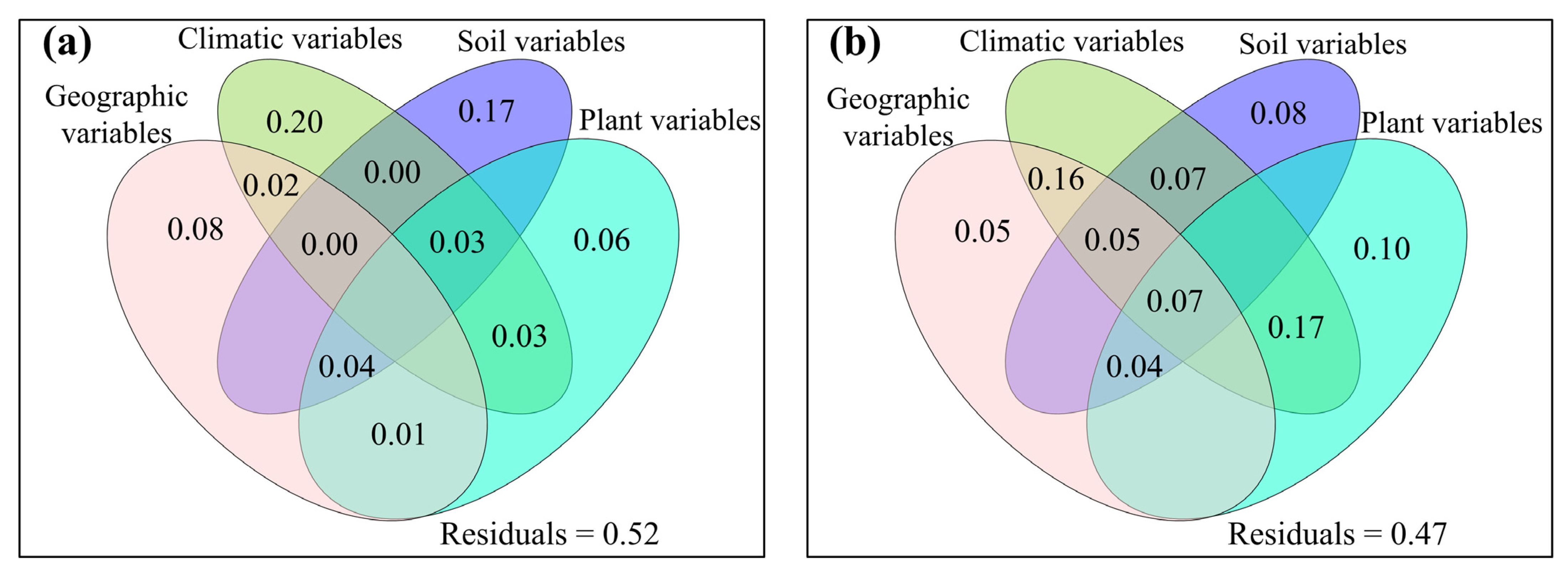
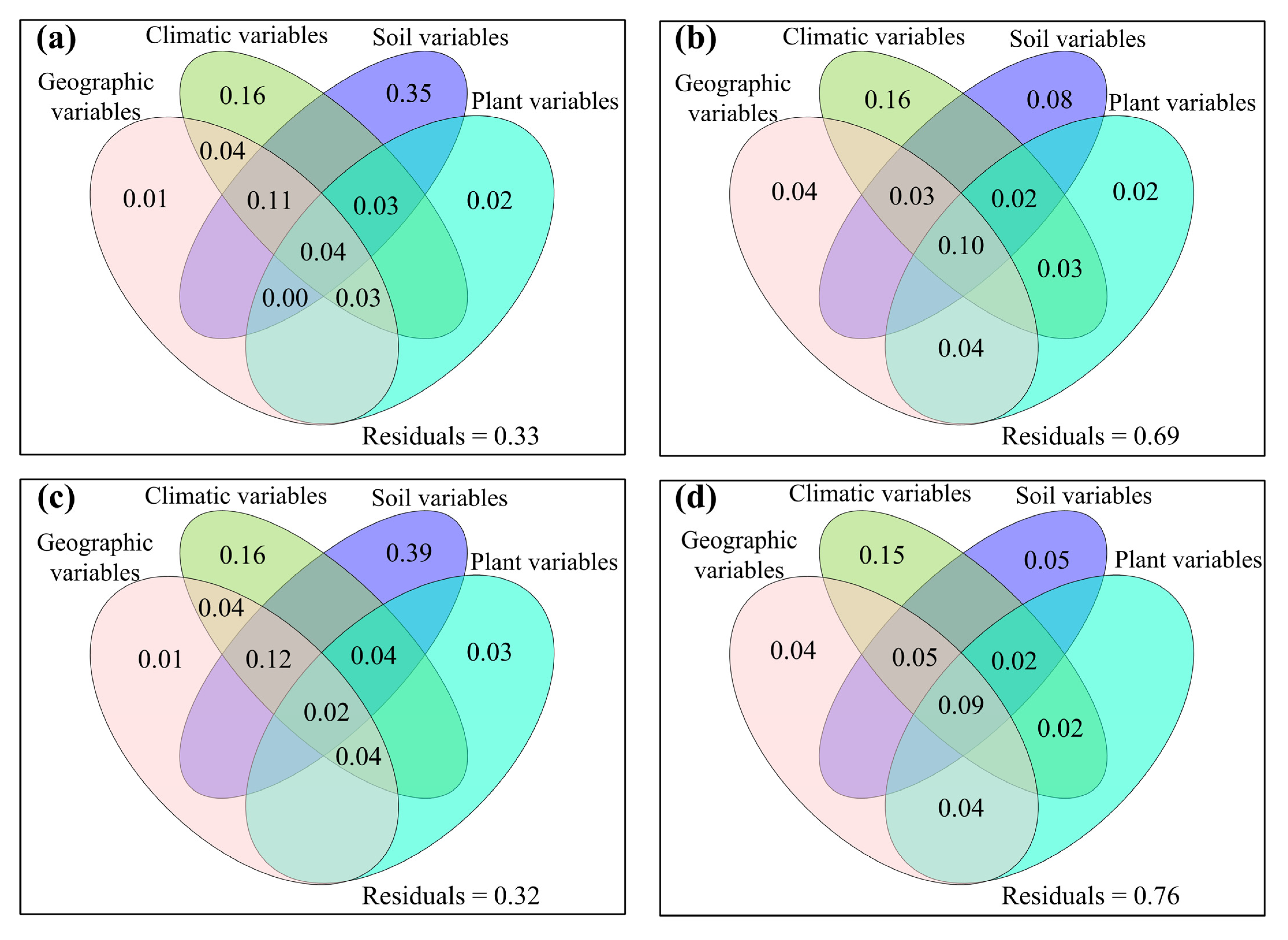
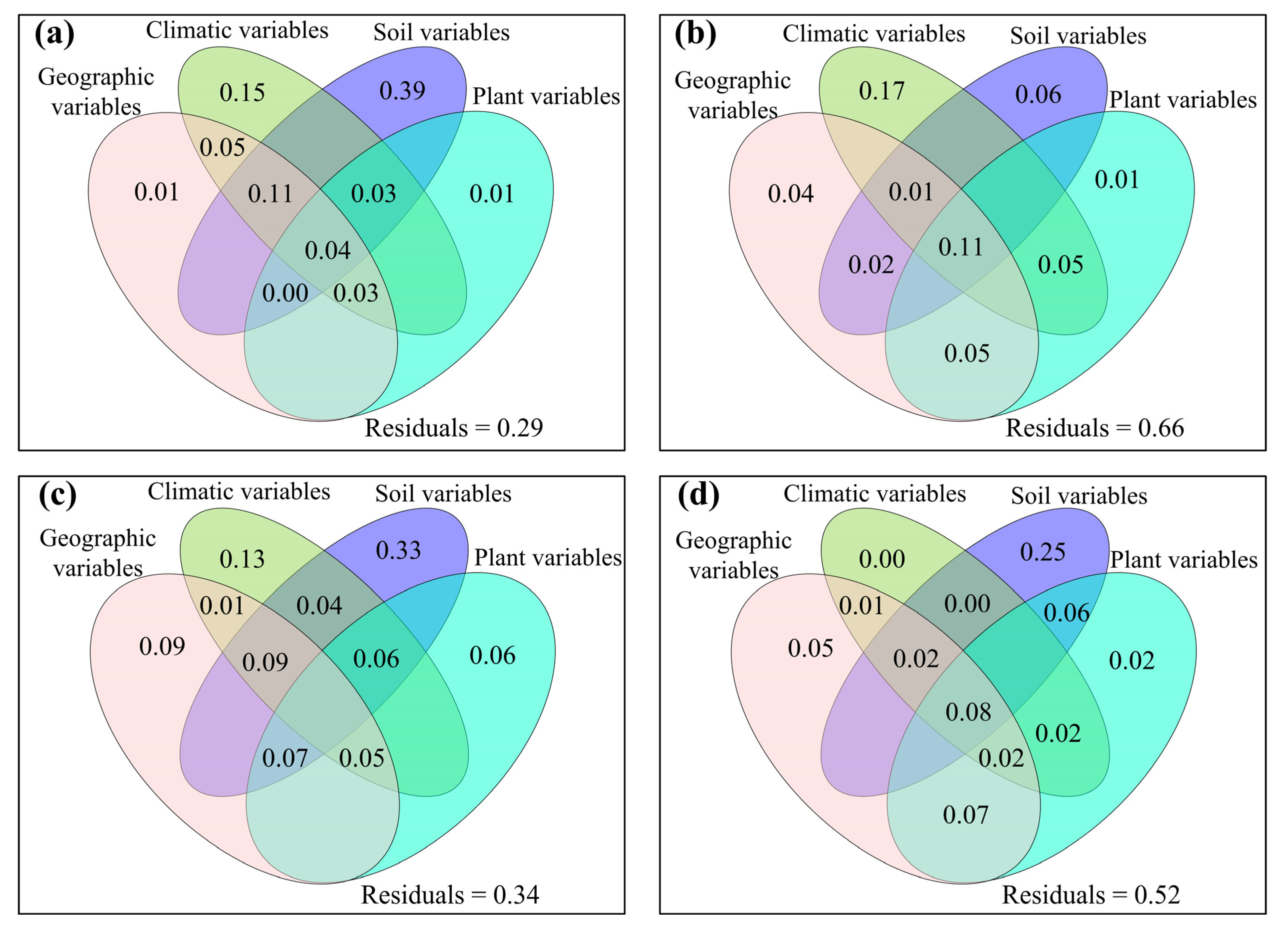
3.2. Correlations between Fungal Community and Environmental Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A





















References
- Chen, B.; Zhang, X.; Tao, J.; Wu, J.; Wang, J.; Shi, P.; Zhang, Y.; Yu, C. The impact of climate change and anthropogenic activities on alpine grassland over the Qinghai-Tibet Plateau. Agric. For. Meteorol. 2014, 189, 11–18. [Google Scholar] [CrossRef]
- Wang, J.; Li, M.; Yu, C.; Fu, G. The change in environmental variables linked to climate change has a stronger effect on aboveground net primary productivity than does phenological change in alpine grasslands. Front. Plant Sci. 2021, 12, 798633. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Fu, G.; Yu, C.; Wang, S. Modeling Nutrition Quality and Storage of Forage Using Climate Data and Normalized-Difference Vegetation Index in Alpine Grasslands. Remote Sens. 2022, 14, 3410. [Google Scholar] [CrossRef]
- Fu, G.; Wang, J.; Li, S. Response of forage nutritional quality to climate change and human activities in alpine grasslands. Sci. Total Environ. 2022, 845, 157552. [Google Scholar] [CrossRef]
- Tian, Y.; Fu, G. Quantifying plant species α-diversity using normalized difference vegetation index and climate data in alpine grasslands. Remote Sens. 2022, 14, 5007. [Google Scholar] [CrossRef]
- Yu, C.Q.; Wang, J.W.; Shen, Z.X.; Fu, G. Effects of experimental warming and increased precipitation on soil respiration in an alpine meadow in the Northern Tibetan Plateau. Sci. Total Environ. 2019, 647, 1490–1497. [Google Scholar] [CrossRef]
- Fu, G.; Zhang, H.R.; Sun, W. Response of plant production to growing/non-growing season asymmetric warming in an alpine meadow of the Northern Tibetan Plateau. Sci. Total Environ. 2019, 650, 2666–2673. [Google Scholar] [CrossRef]
- Sun, J.; Hou, G.; Liu, M.; Fu, G.; Zhan, T.; Zhou, H.; Tsunekawa, A.; Haregeweyn, N. Effects of climatic and grazing changes on desertification of alpine grasslands, Northern Tibet. Ecol. Indic. 2019, 107, 105647. [Google Scholar] [CrossRef]
- Han, F.S.; Yu, C.Q.; Fu, G. Warming alters elevation distributions of soil bacterial and fungal communities in alpine grasslands. Glob. Ecol. Conserv. 2022, 39, e02306. [Google Scholar] [CrossRef]
- Zha, X.; Tian, Y.; Ou, Z.; Fu, G. Response of forage nutrient storages to grazing in alpine grasslands. Front. Plant Sci. 2022, 13, 991287. [Google Scholar] [CrossRef]
- Mao, W.; Zhang, T.; Li, Y. Allometric response of perennial Pennisetum centrasiaticum Tzvel to nutrient and water limitation in the Horqin Sand Land of China. J. Arid Land 2012, 4, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Shen, Z.; Fu, G. Function diversity of soil fungal community has little exclusive effects on the response of aboveground plant production to experimental warming in alpine grasslands. Appl. Soil Ecol. 2021, 168, 104153. [Google Scholar] [CrossRef]
- Zhong, Z.; Fu, G. Response of soil fungal species, phylogenetic and functional diversity to diurnal asymmetric warming in an alpine agricultural ecosystem. Agr. Ecosyst. Environ. 2022, 335, 107993. [Google Scholar] [CrossRef]
- Yu, C.Q.; Han, F.S.; Fu, G. Effects of 7 years experimental warming on soil bacterial and fungal community structure in the Northern Tibet alpine meadow at three elevations. Sci. Total Environ. 2019, 655, 814–822. [Google Scholar] [CrossRef]
- Zhang, H.; Fu, G. Responses of plant, soil bacterial and fungal communities to grazing vary with pasture seasons and grassland types, northern Tibet. Land Degrad. Dev. 2021, 32, 1821–1832. [Google Scholar] [CrossRef]
- Zong, N.; Fu, G. Variations in species and function diversity of soil fungal community along a desertification gradient in an alpine steppe. Ecol. Indic. 2021, 131, 108197. [Google Scholar] [CrossRef]
- Liu, C.; Cui, Y.; Li, X.; Yao, M. microeco: An R package for data mining in microbial community ecology. FEMS Microbiol. Ecol. 2021, 97, fiaa255. [Google Scholar] [CrossRef]
- Sun, W.; Li, S.; Wang, J.; Fu, G. Effects of grazing on plant species and phylogenetic diversity in alpine grasslands, Northern Tibet. Ecol. Eng. 2021, 170, 106331. [Google Scholar] [CrossRef]
- Fu, G.; Shen, Z. Asymmetrical warming of growing/non-growing season increases soil respiration during growing season in an alpine meadow. Sci. Total Environ. 2022, 812, 152591. [Google Scholar] [CrossRef]
- Fu, G.; Sun, W. Temperature sensitivities of vegetation indices and aboveground biomass are primarily linked with warming magnitude in high-cold grasslands. Sci. Total Environ. 2022, 843, 157002. [Google Scholar] [CrossRef]
- Wang, J.; Yu, C.; Fu, G. Warming reconstructs the elevation distributions of aboveground net primary production, plant species and phylogenetic diversity in alpine grasslands. Ecol. Indic. 2021, 133, 108355. [Google Scholar] [CrossRef]
- Wang, J.W.; Yu, C.Q.; Fu, G. Asymmetrical warming between elevations may result in similar plant community composition between elevations in alpine grasslands. Front. Ecol. Evol. 2021, 9, 757943. [Google Scholar] [CrossRef]
- Rinnan, R.; Michelsen, A.; Baath, E.; Jonasson, S. Fifteen years of climate change manipulations alter soil microbial communities in a subarctic heath ecosystem. Glob. Chang. Biol. 2007, 13, 28–39. [Google Scholar] [CrossRef]
- Fu, G.; Sun, W.; Li, S.; Zhong, Z. Response of microbial communities in soil to multi-level warming in a highland barley system of the Lhasa River. J. Resour. Ecol. 2019, 10, 373–378. [Google Scholar]
- Xiong, J.; Peng, F.; Sun, H.; Xue, X.; Chu, H. Divergent responses of soil fungi functional groups to short-term warming. Microb. Ecol. 2014, 68, 708–715. [Google Scholar] [CrossRef]
- Xia, J.Y.; Chen, S.P.; Wan, S.G. Impacts of day versus night warming on soil microclimate: Results from a semiarid temperate steppe. Sci. Total Environ. 2010, 408, 2807–2816. [Google Scholar] [CrossRef]
- Fu, G.; Shen, Z.X.; Zhang, X.Z. Increased precipitation has stronger effects on plant production of an alpine meadow than does experimental warming in the Northern Tibetan Plateau. Agr. For. Meteorol. 2018, 249, 11–21. [Google Scholar] [CrossRef]
- Rui, Y.C.; Wang, Y.F.; Chen, C.R.; Zhou, X.Q.; Wang, S.P.; Xu, Z.H.; Duan, J.C.; Kang, X.M.; Lu, S.B.; Luo, C.Y. Warming and grazing increase mineralization of organic P in an alpine meadow ecosystem of Qinghai-Tibet Plateau, China. Plant Soil 2012, 357, 73–87. [Google Scholar] [CrossRef]
- Ogwu, M.C.; Kerfahi, D.; Song, H.; Dong, K.; Seo, H.; Lim, S.; Srinivasan, S.; Kim, M.K.; Waldman, B.; Adams, J.M. Changes in soil taxonomic and functional diversity resulting from gamma irradiation. Sci. Rep. 2019, 9, 7894. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Fu, G. Responses of phylogenetic diversity of soil fungal community to grazing in alpine grasslands of the Northern Tibet. Acta Agrestia Sin. 2022, 30, 21–28. [Google Scholar]
- Zhou, J.Z.; Ning, D.L. Stochastic community assembly: Does it matter in microbial ecology? Microbiol. Mol. Biol. Rev. 2017, 81, e00002-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Violle, C.; Nemergut, D.R.; Pu, Z.C.; Jiang, L. Phylogenetic limiting similarity and competitive exclusion. Ecol. Lett. 2011, 14, 782–787. [Google Scholar] [CrossRef]
- Hahn, P.G.; Buffington, L.; Larkin, B.; LaFlamme, K.; Maron, J.L.; Lekberg, Y. Effects of short- and long-term variation in resource conditions on soil fungal communities and plant responses to soil biota. Front. Plant Sci. 2018, 9, 1605. [Google Scholar] [CrossRef] [PubMed]
- Chesson, P. Mechanisms of maintenance of species diversity. Annu. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef]
- Li, W.H. An overview of ecological research conducted on the Qinghai-Tibetan Plateau. J. Resour. Ecol. 2017, 8, 1–4. [Google Scholar]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Depth | Variables | Species β- Diversity | Phylogenetic β- Diversity | Function β- Diversity |
|---|---|---|---|---|
| 0–10 cm | Longitude | 0.19 *** | 0.06 + | 0.13 ** |
| Latitude | 0.16 *** | −0.01 | 0.03 | |
| Elevation | 0.24 *** | 0.01 | 0.04 | |
| MAT | 0.07 * | 0.08 * | 0.03 | |
| MAP | 0.16 *** | 0.00 | 0.12 * | |
| AT2020 | 0.04 | 0.03 | 0.03 | |
| AP2020 | 0.16 *** | 0.08 + | 0.12 * | |
| EC | 0.22*** | −0.07 | 0.15 ** | |
| pH | 0.19 *** | 0.13** | −0.08 | |
| SOC | 0.19 *** | −0.07 | 0.16 *** | |
| TN | 0.20 *** | −0.10 | 0.20 *** | |
| TP | −0.05 | −0.07 | 0.04 | |
| TK | 0.13 *** | 0.10 * | −0.05 | |
| NH4+-N | 0.05 + | 0.08 * | 0.01 | |
| NO3−-N | 0.04 | 0.08 + | −0.02 | |
| AP | −0.05 | −0.03 | −0.01 | |
| AK | 0.12 *** | −0.05 | 0.09 ** | |
| C:N | 0.04 | −0.04 | 0.06 + | |
| C:P | 0.16 *** | −0.07 | 0.06 + | |
| C:K | 0.19 *** | −0.07 | 0.16 *** | |
| N:P | 0.17 *** | −0.09 | 0.09 * | |
| N:K | 0.20 *** | −0.10 | 0.21 *** | |
| P:K | −0.04 | −0.07 | 0.05 | |
| Available N:P | 0.00 | 0.07 + | 0.00 | |
| Available N:K | 0.13 *** | 0.08 ** | 0.04 | |
| Available P:K | 0.14 *** | −0.03 | 0.07 * | |
| Plant height | 0.08 * | 0.03 | 0.09 * | |
| Leaf area | 0.15 *** | 0.00 | −0.09 | |
| Stem diameter | 0.09 * | 0.02 | 0.06 + | |
| 10–20 cm | Longitude | 0.17 *** | 0.10 * | 0.25 *** |
| Latitude | 0.14 *** | -0.05 | 0.03 | |
| Elevation | 0.19 *** | 0.01 | 0.00 | |
| MAT | 0.11 ** | 0.02 | −0.06 | |
| MAP | 0.08 * | 0.10 + | 0.23 *** | |
| AT2020 | 0.08 ** | 0.01 | −0.11 | |
| AP2020 | 0.05 | 0.19 ** | 0.26 *** | |
| EC | 0.13 *** | 0.02 | 0.08 * | |
| pH | 0.19 *** | 0.01 | 0.00 | |
| SOC | 0.11 ** | −0.01 | 0.09 * | |
| TN | 0.09 * | −0.02 | 0.04 | |
| TP | −0.02 | 0.07 | 0.01 | |
| TK | 0.12 *** | 0.01 | 0.05 | |
| NH4+-N | 0.12 *** | 0.13 ** | 0.13 ** | |
| NO3−-N | 0.15 *** | −0.02 | 0.10 * | |
| AP | −0.04 | 0.02 | 0.04 | |
| AK | 0.09 ** | 0.01 | 0.15 *** | |
| C:N | 0.03 | −0.01 | −0.05 | |
| C:P | 0.13 *** | 0.00 | 0.03 | |
| C:K | 0.11 ** | −0.02 | 0.08 * | |
| N:P | 0.11 ** | 0.00 | −0.03 | |
| N:K | 0.09 * | −0.03 | 0.04 | |
| P:K | 0.00 | 0.05 | 0.00 | |
| Available N:P | 0.08 * | 0.07 + | 0.00 | |
| Available N:K | 0.09 ** | 0.07 + | 0.17 *** | |
| Available P:K | 0.12 ** | −0.01 | 0.17 *** | |
| Plant height | 0.14 *** | 0.00 | 0.18 *** | |
| Stem diameter | 0.10 ** | 0.06 + | 0.06 | |
| Leaf area | 0.18 *** | −0.01 | −0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, G.; Shen, Z.; Fu, G. Geo-Distribution Patterns of Soil Fungal Community of Pennisetum flaccidum in Tibet. J. Fungi 2022, 8, 1230. https://doi.org/10.3390/jof8111230
Zhang G, Shen Z, Fu G. Geo-Distribution Patterns of Soil Fungal Community of Pennisetum flaccidum in Tibet. Journal of Fungi. 2022; 8(11):1230. https://doi.org/10.3390/jof8111230
Chicago/Turabian StyleZhang, Guangyu, Zhenxi Shen, and Gang Fu. 2022. "Geo-Distribution Patterns of Soil Fungal Community of Pennisetum flaccidum in Tibet" Journal of Fungi 8, no. 11: 1230. https://doi.org/10.3390/jof8111230