

Fungal Hal3 (and Its Close Relative Cab3) as Moonlighting Proteins


Abstract
:1. Introduction
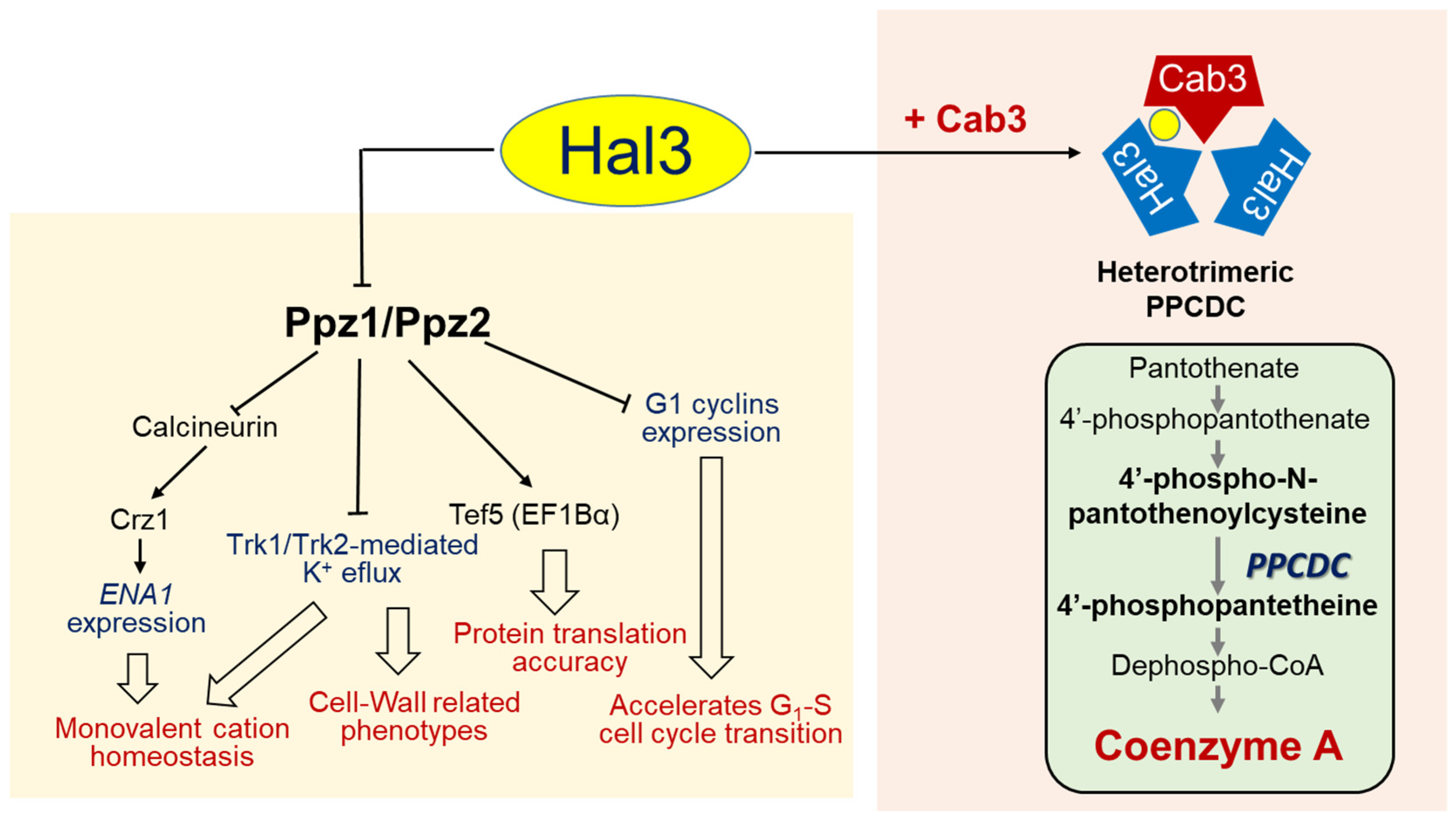
2. Identification of Hal3 as an Inhibitor of the Ser/Thr Protein Phosphatase Ppz1
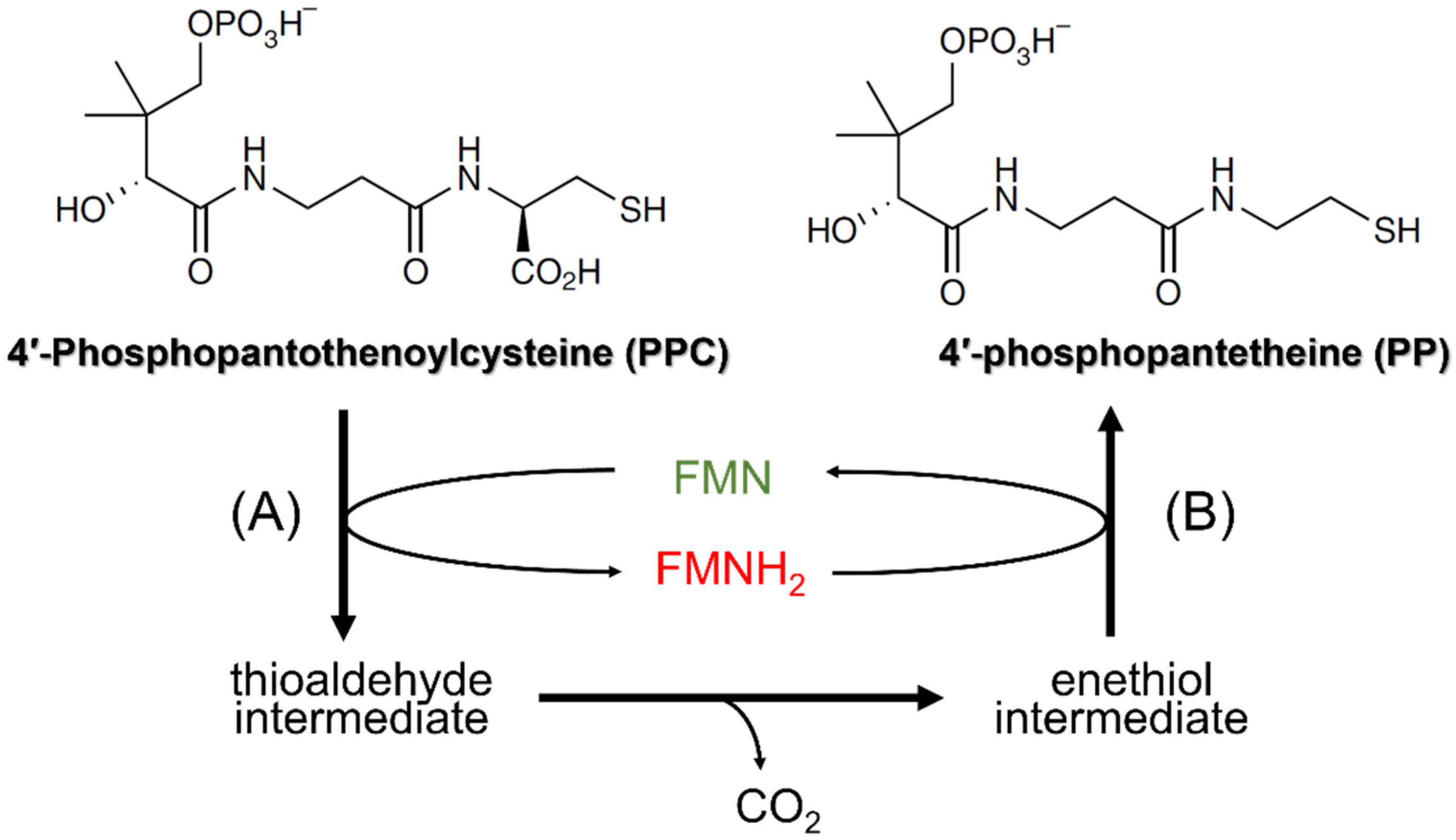
3. Hal3 (and Vhs3) as Moonlighting Proteins: Their Involvement in CoA Biosynthesis
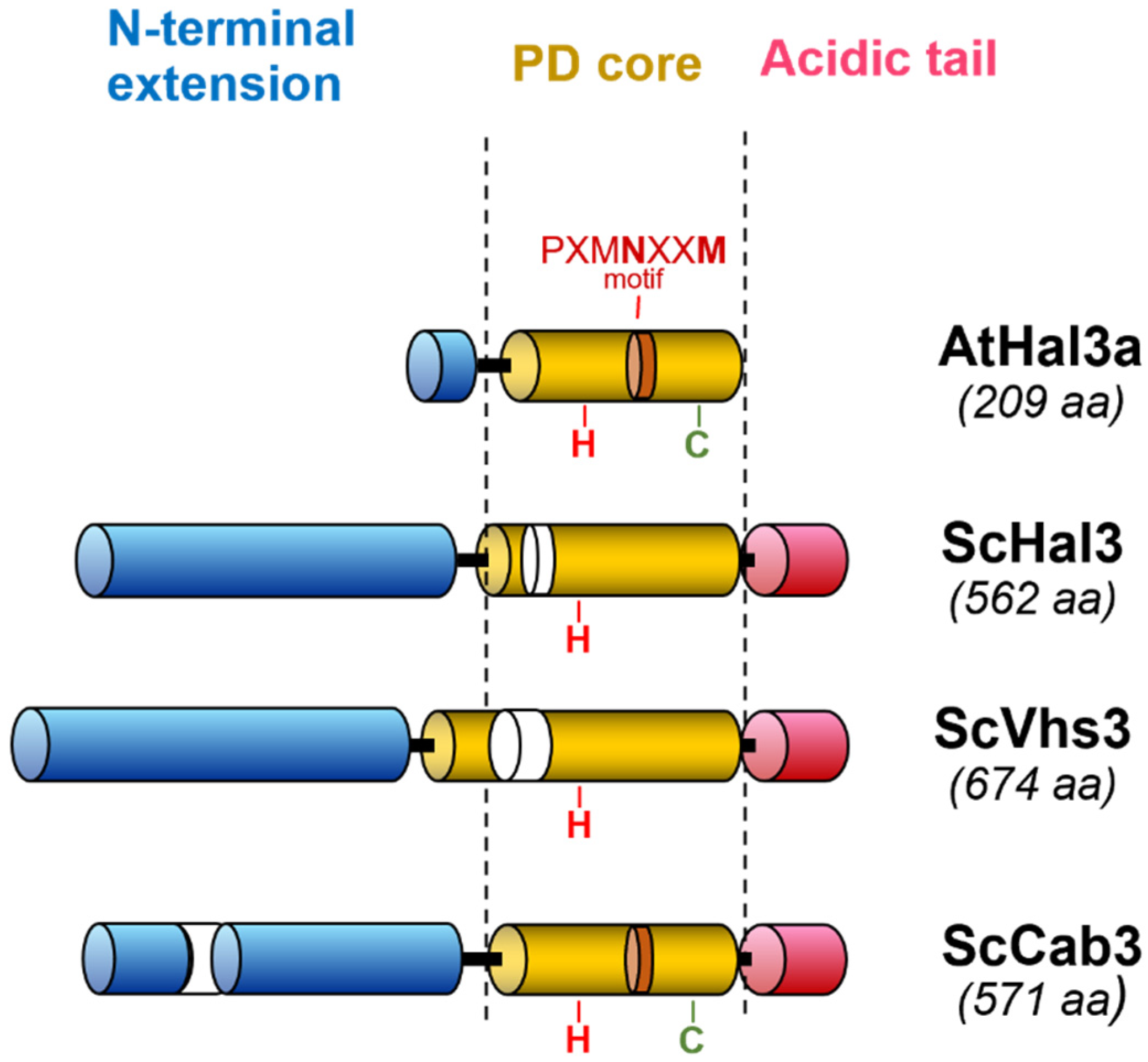
4. Structural Insights into the Moonlighting Functions of Hal3
5. The Moonlighting Capacity of Hal3 (and Cab3) across Fungi
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jeffery, C.J. Why study moonlighting proteins? Front. Genet. 2015, 6, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Como, C.J.; Bose, R.; Arndt, K.T. Overexpression of SIS2, which contains an extremely acidic region, increases the expression of SWI4, CLN1 and CLN2 in sit4 mutants. Genetics 1995, 139, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Ferrando, A.; Kron, S.J.; Rios, G.; Fink, G.R.; Serrano, R. Regulation of cation transport in Saccharomyces cerevisiae the salt tolerance gene HAL3. Mol. Cell Biol. 1995, 15, 5470–5481. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, P.L.; Ali, R.; Serrano, R. CtCdc55p and CtHa13p: Two putative regulatory proteins from Candida tropicalis with long acidic domains. Yeast 1996, 12, 1321–1329. [Google Scholar] [CrossRef]
- Posas, F.; Casamayor, A.; Morral, N.; Ariño, J. Molecular cloning and analysis of a yeast protein phosphatase with an unusual amino-terminal region. J. Biol. Chem. 1992, 267, 11734–11740. [Google Scholar] [CrossRef]
- Lee, K.S.; Hines, L.K.; Levin, D.E. A pair of functionally redundant yeast genes (PPZ1 and PPZ2) encoding type 1-related protein phosphatases function within the PKC1-mediated pathway. Mol. Cell Biol. 1993, 13, 5843–5853. [Google Scholar] [PubMed] [Green Version]
- Posas, F.; Casamayor, A.; Ariño, J. The PPZ protein phosphatases are involved in the maintenance of osmotic stability of yeast cells. FEBS Lett. 1993, 318, 282–286. [Google Scholar] [CrossRef] [Green Version]
- Posas, F.; Camps, M.; Ariño, J. The PPZ protein phosphatases are important determinants of salt tolerance in yeast cells. J. Biol. Chem. 1995, 270, 13036–13041. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, A.; Yenush, L.; Arino, J. Regulation of ENA1 Na(+)-ATPase gene expression by the Ppz1 protein phosphatase is mediated by the calcineurin pathway. Eukaryot. Cell 2003, 2, 937–948. [Google Scholar] [CrossRef] [Green Version]
- De Nadal, E.; Clotet, J.; Posas, F.; Serrano, R.; Gomez, N.; Ariño, J. The yeast halotolerance determinant Hal3p is an inhibitory subunit of the Ppz1p Ser/Thr protein phosphatase. Proc. Natl. Acad. Sci. USA 1998, 95, 7357–7362. [Google Scholar] [CrossRef]
- Clotet, J.; Posas, F.; De Nadal, E.; Arino, J. The NH2-terminal extension of protein phosphatase PPZ1 has an essential functional role. J. Biol. Chem. 1996, 271, 26349–26355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yenush, L.; Mulet, J.M.; Ariño, J.; Serrano, R. The Ppz protein phosphatases are key regulators of K+ and pH homeostasis: Implications for salt tolerance, cell wall integrity and cell cycle progression. EMBO J. 2002, 21, 920–929. [Google Scholar] [CrossRef] [PubMed]
- Merchan, S.; Bernal, D.; Serrano, R.; Yenush, L. Response of the Saccharomyces cerevisiae Mpk1 mitogen-activated protein kinase pathway to increases in internal turgor pressure caused by loss of Ppz protein phosphatases. Eukaryot Cell 2004, 3, 100–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Nadal, E.; Fadden, R.P.P.; Ruiz, A.; Haystead, T.; Ariño, J. A role for the Ppz Ser/Thr protein phosphatases in the regulation of translation elongation factor 1Balpha. J. Biol. Chem. 2001, 276, 14829–14834. [Google Scholar] [CrossRef] [Green Version]
- Clotet, J.; Garí, E.; Aldea, M.; Ariño, J. The yeast ser/thr phosphatases sit4 and ppz1 play opposite roles in regulation of the cell cycle. Mol. Cell Biol. 1999, 19, 2408–2415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, I.; Simón, E.; Casals, N.; Clotet, J.; Ariño, J. Identification of multicopy suppressors of cell cycle arrest at the G1-S transition in Saccharomyces cerevisiae. Yeast 2003, 20, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, A.; Muñoz, I.; Serrano, R.; Gonzalez, A.; Simon, E.; Arino, J. Functional characterization of the Saccharomyces cerevisiae VHS3 gene: A regulatory subunit of the Ppz1 protein phosphatase with novel, phosphatase-unrelated functions. J. Biol. Chem. 2004, 279, 34421–34430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinosa-Ruiz, A.; Belles, J.M.; Serrano, R.; Culianez-Macia, F.A. Arabidopsis thaliana AtHAL3: A flavoprotein related to salt and osmotic tolerance and plant growth. Plant J. 1999, 20, 529–539. [Google Scholar] [CrossRef] [Green Version]
- Albert, A.; Martinez-Ripoll, M.; Espinosa-Ruiz, A.; Yenush, L.; Culianez-Macia, F.A.; Serrano, R. The X-ray structure of the FMN-binding protein AtHal3 provides the structural basis for the activity of a regulatory subunit involved in signal transduction. Struct. Fold. Des. 2000, 8, 961–969. [Google Scholar] [CrossRef] [Green Version]
- Kupke, T.; Hernandez-Acosta, P.; Steinbacher, S.; Culianez-Macia, F.A. Arabidopsis thaliana flavoprotein AtHAL3a catalyzes the decarboxylation of 4’-Phosphopantothenoylcysteine to 4’-phosphopantetheine, a key step in coenzyme A biosynthesis. J. Biol. Chem. 2001, 276, 19190–19196. [Google Scholar] [CrossRef]
- Steinbacher, S.; Hernandez-Acosta, P.; Bieseler, B.; Blaesse, M.; Huber, R.; Culianez-Macia, F.A.; Kupke, T. Crystal structure of the plant PPC decarboxylase AtHAL3a complexed with an ene-thiol reaction intermediate. J. Mol. Biol. 2003, 327, 193–202. [Google Scholar] [CrossRef]
- Hernandez-Acosta, P.; Schmid, D.G.; Jung, G.; Culianez-Macia, F.A.; Kupke, T. Molecular characterization of the Arabidopsis thaliana flavoprotein AtHAL3a reveals the general reaction mechanism of 4’-phosphopantothenoylcysteine decarboxylases. J. Biol. Chem. 2002, 277, 20490–20498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kupke, T.; Uebele, M.; Schmid, D.; Jung, G.; Blaesse, M.; Steinbacher, S. Molecular characterization of lantibiotic-synthesizing enzyme EpiD reveals a function for bacterial Dfp proteins in coenzyme A biosynthesis. J. Biol. Chem. 2000, 275, 31838–31846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giaever, G.; Chu, A.M.; Ni, L.; Connelly, C.; Riles, L.; Veronneau, S.; Dow, S.; Lucau-Danila, A.; Anderson, K.; Andre, B.; et al. Functional profiling of the Saccharomyces cerevisiae genome. Nature 2002, 418, 387–391. [Google Scholar] [CrossRef]
- Olzhausen, J.; Schubbe, S.; Schuller, H.J. Genetic analysis of coenzyme A biosynthesis in the yeast Saccharomyces cerevisiae: Identification of a conditional mutation in the pantothenate kinase gene CAB1. Curr. Genet. 2009, 55, 163–173. [Google Scholar] [CrossRef]
- Ruiz, A.; Gonzalez, A.; Munoz, I.; Serrano, R.; Abrie, J.A.; Strauss, E.; Arino, J. Moonlighting proteins Hal3 and Vhs3 form a heteromeric PPCDC with Ykl088w in yeast CoA biosynthesis. Nat. Chem. Biol. 2009, 5, 920–928. [Google Scholar] [CrossRef] [Green Version]
- Munoz, I.; Ruiz, A.; Marquina, M.; Barcelo, A.; Albert, A.; Arino, J. Functional characterization of the yeast Ppz1 phosphatase inhibitory subunit Hal3: A mutagenesis study. J. Biol. Chem. 2004, 279, 42619–42627. [Google Scholar] [CrossRef] [Green Version]
- Manoj, N.; Ealick, S.E. Unusual space-group pseudosymmetry in crystals of human phosphopantothenoylcysteine decarboxylase. Acta Crystallogr. D Biol. Crystallogr. 2003, 59, 1762–1766. [Google Scholar] [CrossRef] [Green Version]
- Abrie, J.A.A.; González, A.; Strauss, E.; Ariño, J. Functional mapping of the disparate activities of the yeast moonlighting protein Hal3. Biochem. J. 2012, 442, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Abrie, J.A.; Molero, C.; Ariño, J.; Strauss, E. Complex stability and dynamic subunit interchange modulates the disparate activities of the yeast moonlighting proteins Hal3 and Vhs3. Sci. Rep. 2015, 5, 15774. [Google Scholar] [CrossRef]
- Wang, M.; Herrmann, C.J.; Simonovic, M.; Szklarczyk, D.; von, M.C. Version 4.0 of PaxDb: Protein abundance data, integrated across model organisms, tissues, and cell-lines. Proteomics 2015, 15, 3163–3168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, B.; Baryshnikova, A.; Brown, G.W. Unification of Protein Abundance Datasets Yields a Quantitative Saccharomyces cerevisiae Proteome. Cell Syst. 2018, 6, 192–205.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santolaria, C.; Velázquez, D.; Strauss, E.; Ariño, J. Mutations at the hydrophobic core affect Hal3 trimer stability, reducing its Ppz1 inhibitory capacity but not its PPCDC moonlighting function. Sci. Rep. 2018, 8, 14701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olzhausen, J.; Moritz, T.; Neetz, T.; Schüller, H.-J. Molecular characterization of the heteromeric coenzyme A-synthesizing protein complex (CoA-SPC) in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2013, 13, 565–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Offley, S.R.; Schmidt, M.C. Protein phosphatases of Saccharomyces cerevisiae. Curr. Genet. 2019, 65, 41–55. [Google Scholar] [CrossRef] [PubMed]
- Ariño, J.; Velázquez, D.; Casamayor, A. Ser/thr protein phosphatases in fungi: Structure, regulation and function. Microb. Cell 2019, 6, 217–256. [Google Scholar] [CrossRef] [Green Version]
- Casamayor, A.; Ariño, J. Controlling Ser/Thr protein phosphatase PP1 activity and function through interaction with regulatory subunits. Adv. Protein Chem. Struct. Biol. 2020, 122, 231–288. [Google Scholar] [CrossRef]
- Garcia-Gimeno, M.A.; Munoz, I.; Arino, J.; Sanz, P. Molecular characterization of Ypi1, a novel Saccharomyces cerevisiae type 1 protein phosphatase inhibitor. J. Biol. Chem. 2003, 278, 47744–47752. [Google Scholar] [CrossRef] [Green Version]
- Molero, C.; Casado, C.; Ariño, J. The inhibitory mechanism of Hal3 on the yeast Ppz1 phosphatase: A mutagenesis analysis. Sci. Rep. 2017, 7, 8819. [Google Scholar] [CrossRef]
- Zhang, C.; García-Rodas, R.; Molero, C.; de Oliveira, H.C.; Tabernero, L.; Reverter, D.; Zaragoza, O.; Ariño, J. Characterization of the atypical Ppz/Hal3 phosphatase system from the pathogenic fungus Cryptococcus neoformans. Mol. Microbiol. 2019, 111, 898–917. [Google Scholar] [CrossRef]
- Casamayor, A.; Velázquez, D.; Santolaria, C.; Albacar, M.; Rasmussen, M.I.; Højrup, P.; Ariño, J. Comparative Analysis of Type 1 and Type Z Protein Phosphatases Reveals D615 as a Key Residue for Ppz1 Regulation. Int. J. Mol. Sci. 2022, 23, 1327. [Google Scholar] [CrossRef] [PubMed]
- Santolaria, C.; Velázquez, D.; Albacar, M.; Casamayor, A.; Ariño, J. Functional mapping of the N-terminal region of the yeast moonlighting protein Sis2/Hal3 reveals crucial residues for Ppz1 regulation. FEBS J. 2022. [Google Scholar] [CrossRef] [PubMed]
- Makanae, K.; Kintaka, R.; Makino, T.; Kitano, H.; Moriya, H. Identification of dosage-sensitive genes in Saccharomyces cerevisiae using the genetic tug-of-war method. Genome Res. 2013, 23, 300–311. [Google Scholar] [CrossRef] [Green Version]
- Calafí, C.; López-Malo, M.; Velázquez, D.; Zhang, C.; Fernández-Fernández, J.; Rodríguez-Galán, O.; de la Cruz, J.; Ariño, J.; Casamayor, A. Overexpression of budding yeast protein phosphatase Ppz1 impairs translation. Biochim. Biophys. Acta-Mol. Cell Res. 2020, 1867, 118727. [Google Scholar] [CrossRef] [PubMed]
- Velázquez, D.; Albacar, M.; Zhang, C.; Calafí, C.; López-Malo, M.; Torres-Torronteras, J.; Martí, R.; Kovalchuk, S.I.; Pinson, B.; Jensen, O.N.; et al. Yeast Ppz1 protein phosphatase toxicity involves the alteration of multiple cellular targets. Sci. Rep. 2020, 10, 15613. [Google Scholar] [CrossRef]
- Albacar, M.; Sacka, L.; Calafí, C.; Velázquez, D.; Casamayor, A.; Ariño, J.; Zimmermannova, O. The Toxic Effects of Ppz1 Overexpression Involve Nha1-Mediated Deregulation of K+ and H+ Homeostasis. J. Fungi 2021, 7, 1010. [Google Scholar] [CrossRef] [PubMed]
- Casamayor, A.; Ariño, J. When Phosphatases Go Mad: The Molecular Basis for Toxicity of Yeast Ppz1. Int. J. Mol. Sci. 2022, 23, 4304. [Google Scholar] [CrossRef] [PubMed]
- Albacar, M.; Velázquez, D.; Casamayor, A.; Ariño, J. The toxic effects of yeast Ppz1 phosphatase are counteracted by subcellular relocalization mediated by its regulatory subunit Hal3. FEBS Lett. 2022, 596, 1556–1566. [Google Scholar] [CrossRef]
- Balcells, L.; Gomez, N.; Casamayor, A.; Clotet, J.; Arino, J. Regulation of salt tolerance in fission yeast by a protein-phosphatase-Z-like Ser/Thr protein phosphatase. Eur. J. Biochem. 1997, 250, 476–483. [Google Scholar] [CrossRef] [Green Version]
- Vissi, E.; Clotet, J.; de Nadal, E.; Barcelo, A.; Bako, E.; Gergely, P.; Dombradi, V.; Arino, J. Functional analysis of the Neurospora crassa PZL-1 protein phosphatase by expression in budding and fission yeast. Yeast 2001, 18, 115–124. [Google Scholar] [CrossRef]
- Ádám, C.; Erdei, É.; Casado, C.; Kovács, L.; González, A.; Majoros, L.; Petrényi, K.; Bagossi, P.; Farkas, I.; Molnar, M.; et al. Protein phosphatase CaPpz1 is involved in cation homeostasis, cell wall integrity and virulence of Candida albicans. Microbiology 2012, 158, 1258–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacs, L.; Farkas, I.; Majoros, L.; Miskei, M.; Pocsi, I.; Dombradi, V. The polymorphism of protein phosphatase Z1 gene in Candida albicans. J. Basic Microbiol. 2010, 50 (Suppl. S1), S74–S82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minhas, A.; Sharma, A.; Kaur, H.; Fnu, Y.; Ganesan, K.; Mondal, A.K. A conserved Ser/Arg rich motif in PPZ orthologs from fungi is important for its role in cation tolerance. J. Biol. Chem. 2012, 287, 7301–7312. [Google Scholar] [CrossRef] [Green Version]
- Leiter, É.; González, A.; Erdei, É.; Casado, C.; Kovács, L.; Ádám, C.; Oláh, J.; Miskei, M.; Molnar, M.; Farkas, I.; et al. Protein phosphatase Z modulates oxidative stress response in fungi. Fungal Genet. Biol. 2012, 49, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; de la Torre, A.; Pérez-Martín, J.; Ariño, J. Protein Phosphatase Ppz1 Is Not Regulated by a Hal3-Like Protein in Plant Pathogen Ustilago maydis. Int. J. Mol. Sci. 2019, 20, 3817. [Google Scholar] [CrossRef] [Green Version]
- Molero, C.; Petrényi, K.; González, A.; Carmona, M.; Gelis, S.; Abrie, J.A.; Strauss, E.; Ramos, J.; Dombradi, V.; Hidalgo, E.; et al. The Schizosaccharomyces pombe fusion gene hal3 encodes three distinct activities. Mol. Microbiol. 2013, 90, 367–382. [Google Scholar] [CrossRef]
- Petrényi, K.; Molero, C.; Kónya, Z.; Erdődi, F.; Ariño, J.; Dombrádi, V. Analysis of Two Putative Candida albicans Phosphopantothenoylcysteine Decarboxylase / Protein Phosphatase Z Regulatory Subunits Reveals an Unexpected Distribution of Functional Roles. PLoS ONE 2016, 11, e0160965. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.J.; Kim, S.H. A genome Tree of Life for the Fungi kingdom. Proc. Natl. Acad. Sci. USA 2017, 114, 9391–9396. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Mean N-Term (Hal3 & Cab3) | Consensus Sequence at the N-Terminal Extension | Acidic C-Tail (% aa) | |
|---|---|---|---|---|
| Hal3 | Cab3 | Hal3/Cab3 | ||
| Saccharomycetaceae | 356 | KRxxxVTF (1) | --xxxVSF (2) | 65.8/67.6 |
| Saccharomycodaceae | 578 | BBxxxVTF | - | 49.5/46.8 |
| Phaffomycetaceae | 208 | - | K-xxxVSF | 51.3/51.4 |
| KRxxxISF (3) | ||||
| --xxxVKF (4) | ||||
| Pichiaceae | 384 | - | - | 50.6/52.7 |
| RKxxxVTF (5) | ||||
| CUG-Ser1 clade | 252 | - | --xxxVSF | 46.1/48.0 |
| -RxxxVSF (6) | ||||
| Dipodascaceae | 173 | - | - | 39.5/39.7 |
| SRxxxVSF (7) | ||||
| Saccharomycetales inc. sed. | 180 | - | K-xxxVSF (8) | 44.4/46.4 |
| Trichosporonaceae | 48 | - | - | 9.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casamayor, A.; Ariño, J. Fungal Hal3 (and Its Close Relative Cab3) as Moonlighting Proteins. J. Fungi 2022, 8, 1066. https://doi.org/10.3390/jof8101066
Casamayor A, Ariño J. Fungal Hal3 (and Its Close Relative Cab3) as Moonlighting Proteins. Journal of Fungi. 2022; 8(10):1066. https://doi.org/10.3390/jof8101066
Chicago/Turabian StyleCasamayor, Antonio, and Joaquín Ariño. 2022. "Fungal Hal3 (and Its Close Relative Cab3) as Moonlighting Proteins" Journal of Fungi 8, no. 10: 1066. https://doi.org/10.3390/jof8101066