
Development of Carbazole Derivatives Compounds against Candida albicans: Candidates to Prevent Hyphal Formation via the Ras1-MAPK Pathway


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Culture Media, and Chemicals

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Relevant Genotype | Source |
|---|---|---|
| SC5314 | Clinical isolate from the London Mycological Reference Laboratory | [23] |
| BWP17 | ura3::imm434/ura3::imm434 his1::hisG/his1::hisG arg4::hisG/arg4::hisG | [24] |
| Ume6-myc | As BWP17 except for ume6::HIS1/UME6-9myc-NAT1 | In this study |
| Nrg1-myc | As BWP17 except for nrg1::HIS1/NRG1-9myc-NAT1 | In this study |
| tetO-UME6 | As BWP17 except for ADH1/adh1::Ptet-UME6 | In this study |
| 12–99 | Fluconazole resistant strain | [25] |
| 89 | Caspofungin resistant strain | [26] |
| 177 | Caspofungin resistant strain | [26] |
2.2. Antifungal Susceptibility Test
2.3. Hyphae Formation
2.4. Determination of the Minimum Inhibitory Concentrations of C. albicans Biofilm Formation
2.5. Cell Cytotoxicity Test
2.6. Real-Time PCR
2.7. Immunoblotting
2.8. Antifungal Activity Test in the Murine Candidiasis Model
2.9. Ethics Statement
3. Results
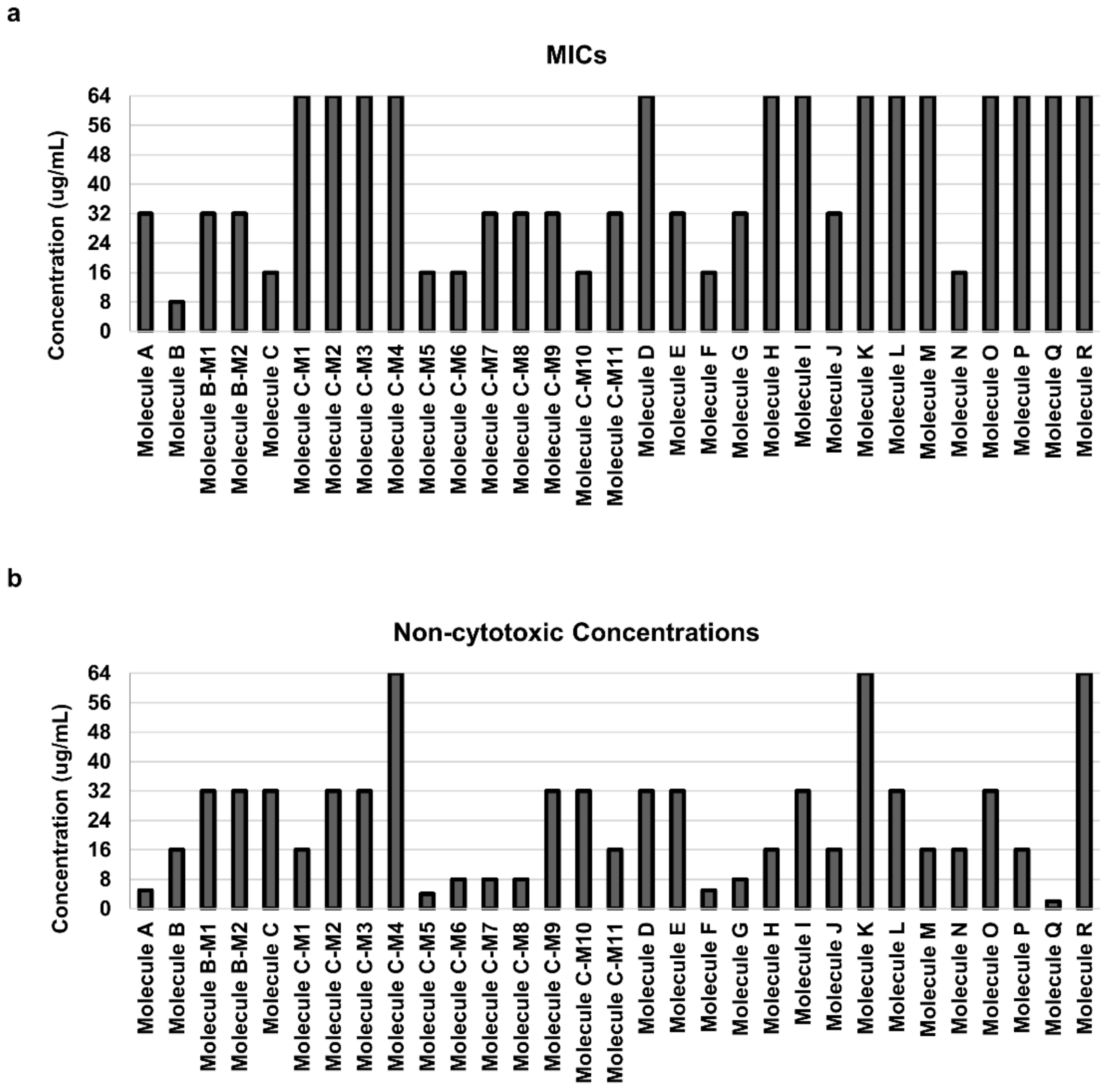
3.1. Identification of Carbazole Derivatives That Inhibit the Growth of Candida Albicans
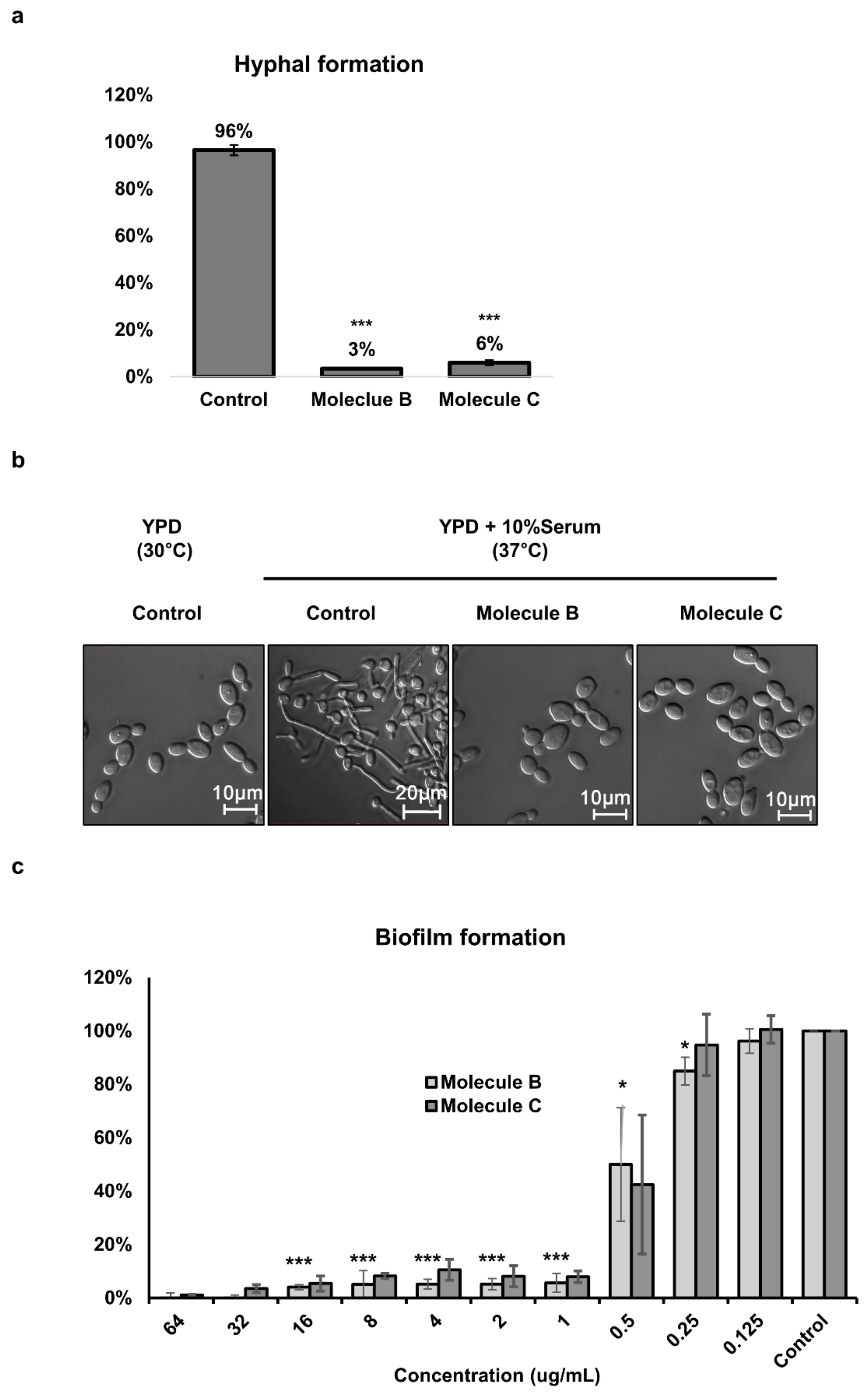
3.2. Inhibitory Effects of Molecule B and Molecule C on Morphogenesis and Biofilm Formation in Candida Albicans
3.3. Virulence Diminishes in the Candidiasis Murine Model Treated with Molecule B and Molecule C via Oral Intake and Vein Injection
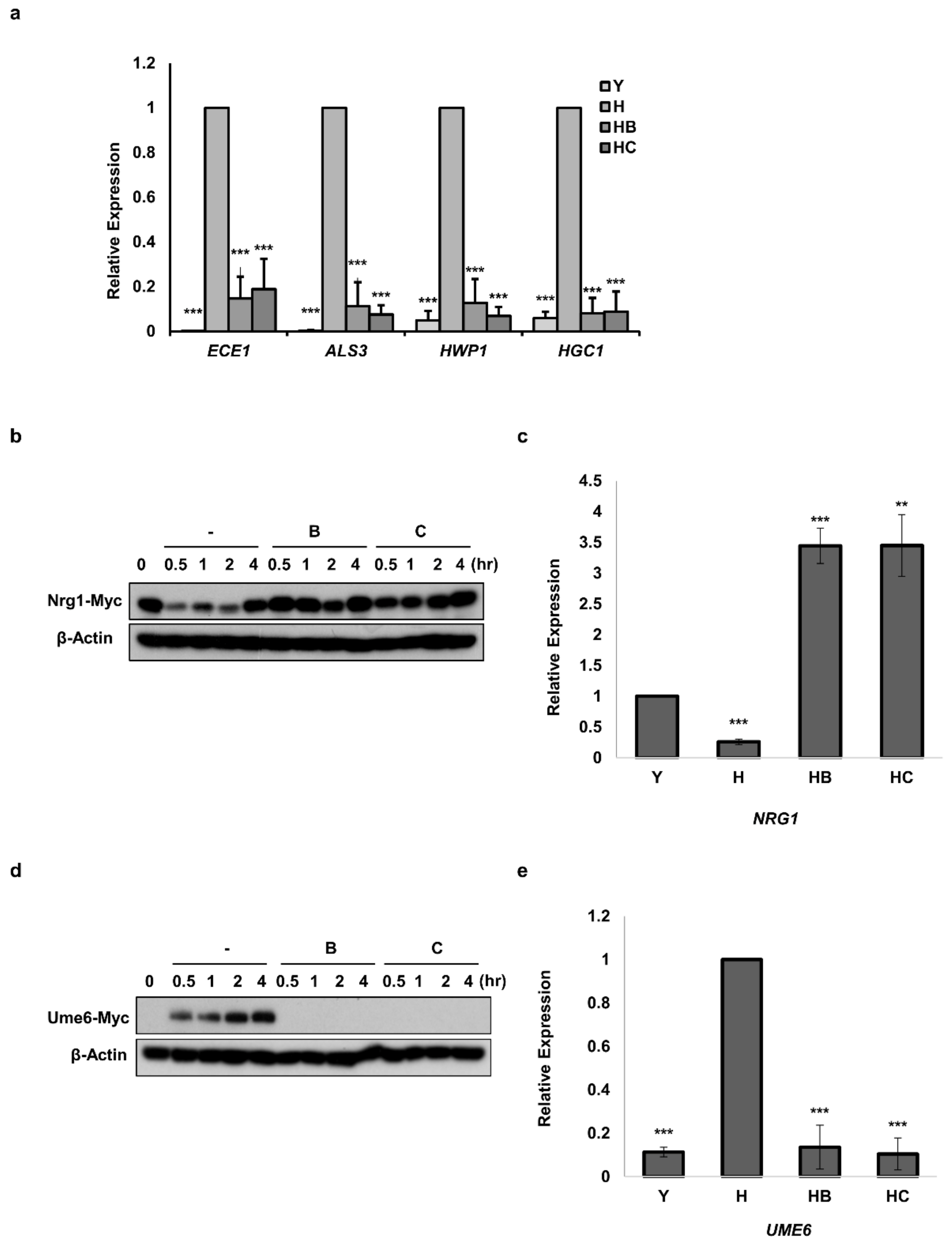
3.4. Molecule B and Molecule C Inhibit the Morphogenesis of C. albicans by Regulating the Protein Levels of Nrg1 and Ume6
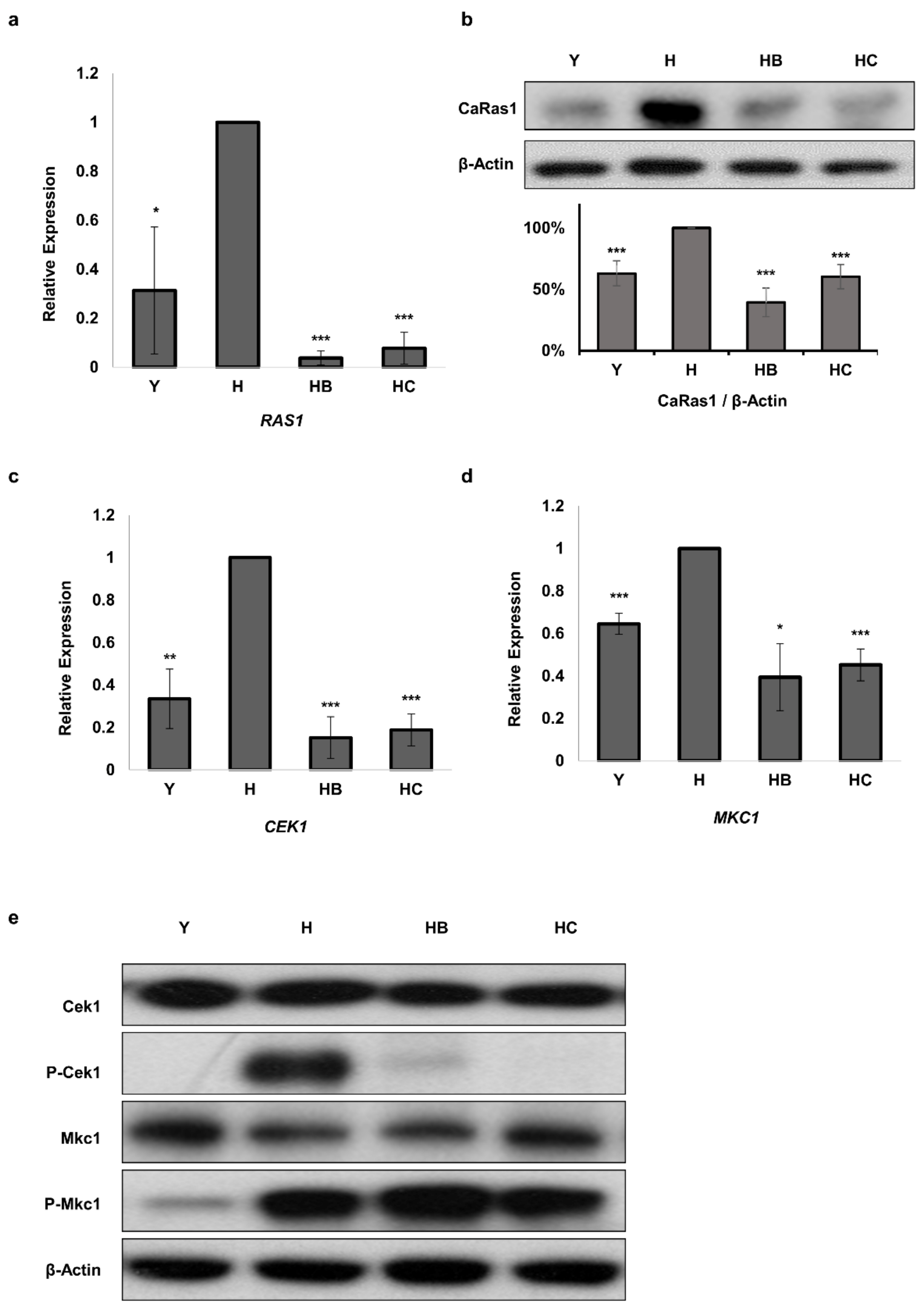
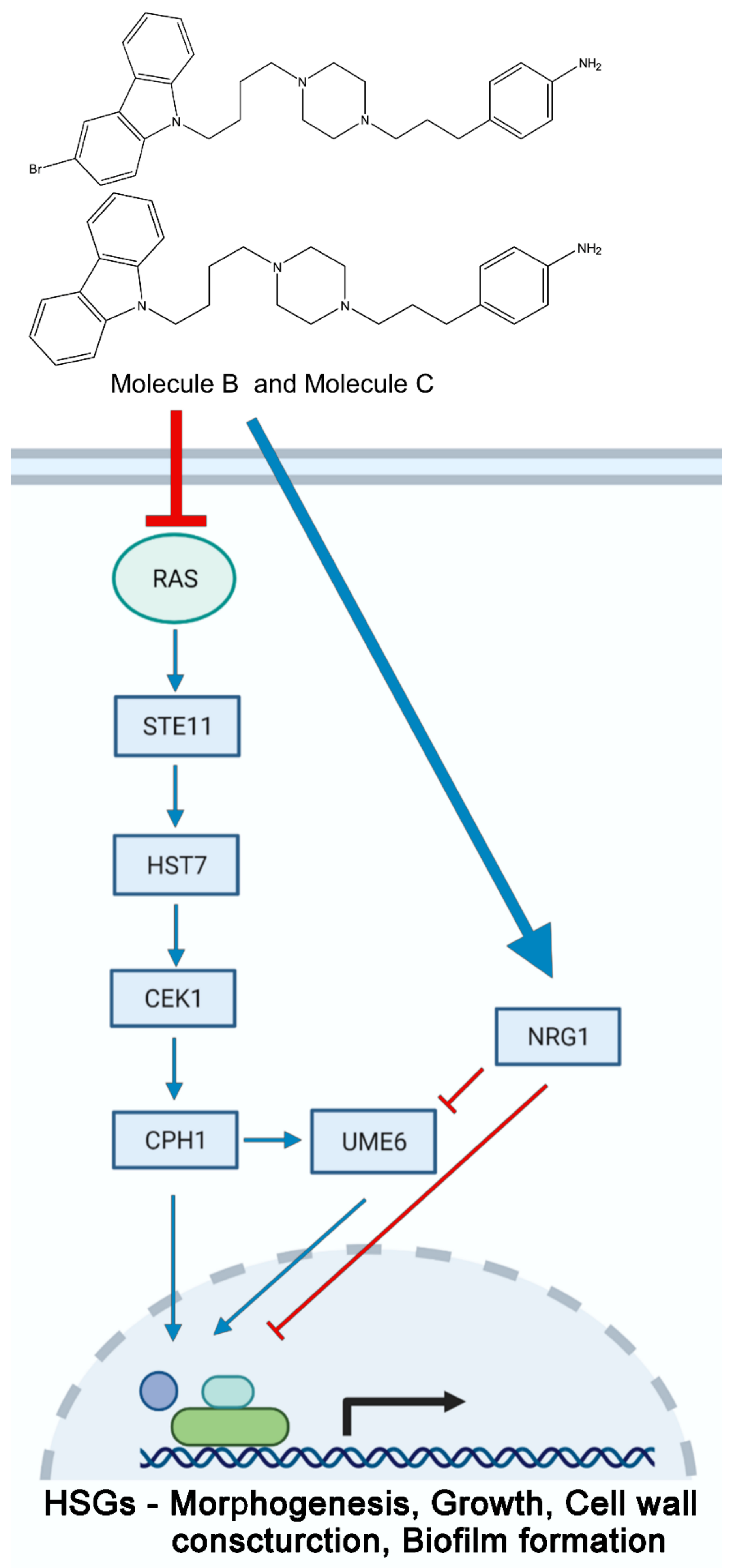
3.5. Molecule B and Molecule C Regulate the Ras1 and MAPK Pathways
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al-Hatmi, A.M.S.; Mohsin, J.; Al-Huraizi, A.; Khamis, F. COVID-19 associated invasive candidiasis. J. Infect. 2021, 82, e45–e46. [Google Scholar] [CrossRef] [PubMed]
- Govender, N.P.; Todd, J.; Nel, J.; Mer, M.; Karstaedt, A.; Cohen, C.; for GERMS-SA. HIV Infection as Risk Factor for Death among Hospitalized Persons with Candidemia, South Africa, 2012–2017. Emerg. Infect. Dis. 2021, 27, 1607–1615. [Google Scholar] [CrossRef]
- San-Juan, R.; Aguado, J.M.; Lumbreras, C.; Fortun, J.; Len, O.; Munoz, P.; Montejo, M.; Moreno, A.; Cordero, E.; Blanes, M.; et al. Universal prophylaxis with fluconazole for the prevention of early invasive fungal infection in low-risk liver transplant recipients. Transplantation 2011, 92, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Klein, B.S.; Tebbets, B. Dimorphism and virulence in fungi. Curr. Opin. Microbiol. 2007, 10, 314–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.W.; Joo, Y.J.; Chun, Y.J.; Park, Y.K.; Kim, J. Cross-talk between Tor1 and Sch9 regulates hyphae-specific genes or ribosomal protein genes in a mutually exclusive manner in Candida albicans. Mol. Microbiol. 2019, 112, 1041–1057. [Google Scholar] [CrossRef]
- Gow, N.A.; Brown, A.J.; Odds, F.C. Fungal morphogenesis and host invasion. Curr. Opin. Microbiol. 2002, 5, 366–371. [Google Scholar] [CrossRef]
- Sudbery, P.E. Growth of Candida albicans hyphae. Nat. Rev. Microbiol. 2011, 9, 737–748. [Google Scholar] [CrossRef]
- Kim, J.; Sudbery, P. Candida albicans, a major human fungal pathogen. J. Microbiol. 2011, 49, 171–177. [Google Scholar] [CrossRef]
- Hall, R.A.; De Sordi, L.; Maccallum, D.M.; Topal, H.; Eaton, R.; Bloor, J.W.; Robinson, G.K.; Levin, L.R.; Buck, J.; Wang, Y.; et al. CO(2) acts as a signalling molecule in populations of the fungal pathogen Candida albicans. PLoS Pathog. 2010, 6, e1001193. [Google Scholar] [CrossRef] [Green Version]
- Kohler, J.R.; Fink, G.R. Candida albicans strains heterozygous and homozygous for mutations in mitogen-activated protein kinase signaling components have defects in hyphal development. Proc. Natl. Acad. Sci. USA 1996, 93, 13223–13228. [Google Scholar] [CrossRef] [Green Version]
- Feng, Q.; Summers, E.; Guo, B.; Fink, G. Ras signaling is required for serum-induced hyphal differentiation in Candida albicans. J. Bacteriol. 1999, 181, 6339–6346. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, M.; Thompson, D.S.; Lazzell, A.; Carlisle, P.L.; Pierce, C.; Monteagudo, C.; Lopez-Ribot, J.L.; Kadosh, D. UME6, a novel filament-specific regulator of Candida albicans hyphal extension and virulence. Mol. Biol. Cell 2008, 19, 1354–1365. [Google Scholar] [CrossRef] [Green Version]
- Murad, A.M.; Leng, P.; Straffon, M.; Wishart, J.; Macaskill, S.; MacCallum, D.; Schnell, N.; Talibi, D.; Marechal, D.; Tekaia, F.; et al. NRG1 represses yeast-hypha morphogenesis and hypha-specific gene expression in Candida albicans. EMBO J. 2001, 20, 4742–4752. [Google Scholar] [CrossRef] [Green Version]
- Braun, B.R.; Kadosh, D.; Johnson, A.D. NRG1, a repressor of filamentous growth in C.albicans, is down-regulated during filament induction. EMBO J. 2001, 20, 4753–4761. [Google Scholar] [CrossRef] [PubMed]
- Hitchcock, C.A. Cytochrome P-450-dependent 14 alpha-sterol demethylase of Candida albicans and its interaction with azole antifungals. Biochem. Soc. Trans. 1991, 19, 782–787. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.J.; Li, M.Y.; Jin, X.J.; Zheng, C.J.; Piao, H.R. Design, synthesis and evaluation of carbazole derivatives as potential antimicrobial agents. J. Enzym. Inhib. Med. Chem. 2021, 36, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Gluszynska, A. Biological potential of carbazole derivatives. Eur. J. Med. Chem. 2015, 94, 405–426. [Google Scholar] [CrossRef]
- Lee, H.Y.; Jung, Y.; Kim, W.; Kim, J.H.; Suh, M.S.; Shin, S.K.; Yoon, H.J. Structure-activity relationship studies of the chromosome segregation inhibitor, Incentrom A. Bioorganic Med. Chem. Lett. 2008, 18, 4670–4674. [Google Scholar] [CrossRef]
- Kim, S.W.; Park, Y.K.; Joo, Y.J.; Chun, Y.J.; Hwang, J.Y.; Baek, J.H.; Kim, J. Subunits of the vacuolar H+-ATPase complex, Vma4 and Vma10, are essential for virulence and represent potential drug targets in Candida albicans. Fungal Biol. 2019, 123, 709–722. [Google Scholar] [CrossRef]
- Jung, Y.; Kim, H.D.; Yang, H.W.; Kim, H.J.; Jang, C.Y.; Kim, J. Modulating cellular balance of Rps3 mono-ubiquitination by both Hel2 E3 ligase and Ubp3 deubiquitinase regulates protein quality control. Exp. Mol. Med. 2017, 49, e390. [Google Scholar] [CrossRef] [Green Version]
- Seong, K.M.; Jung, S.O.; Kim, H.D.; Kim, H.J.; Jung, Y.J.; Choi, S.Y.; Kim, J. Yeast ribosomal protein S3 possesses a beta-lyase activity on damaged DNA. FEBS Lett. 2012, 586, 356–361. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved Standard-Third Edition; CLSI document M27-A3; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008; ISBN 1-56238-666-2. [Google Scholar]
- Fonzi, W.A.; Irwin, M.Y. Isogenic strain construction and gene mapping in Candida albicans. Genetics 1993, 134, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.B.; Davis, D.; Mitchell, A.P. Rapid hypothesis testing with Candida albicans through gene disruption with short homology regions. J. Bacteriol. 1999, 181, 1868–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.C.; Pfaller, M.A.; Rinaldi, M.G.; Smith, J.; Redding, S.W. Stable azole drug resistance associated with a substrain of Candida albicans from an HIV-infected patient. Oral. Dis. 1997, 3 (Suppl. 1), S102–S109. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Effron, G.; Park, S.; Perlin, D.S. Correlating echinocandin MIC and kinetic inhibition of fks1 mutant glucan synthases for Candida albicans: Implications for interpretive breakpoints. Antimicrob. Agents Chemother. 2009, 53, 112–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Kim, H.D.; Kim, J. Cytoplasmic ribosomal protein S3 (rpS3) plays a pivotal role in mitochondrial DNA damage surveillance. Biochim. Biophys Acta 2013, 1833, 2943–2952. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.S.; Kim, H.D.; Park, Y.J.; Kong, E.; Yang, H.W.; Jung, Y.; Kim, Y.; Kim, J. JNK activation induced by ribotoxic stress is initiated from 80S monosomes but not polysomes. BMB Rep. 2019, 52, 502–507. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.; Seong, K.M.; Baek, J.H.; Kim, J. Ssb2 is a novel factor in regulating synthesis and degradation of Gcn4 in Saccharomyces cerevisiae. Mol. Microbiol. 2018, 110, 728–740. [Google Scholar] [CrossRef]
- Yang, H.W.; Kim, H.D.; Kim, T.S.; Kim, J. Senescent cells differentially translate senescence-related mRNAs via ribosome heterogeneity. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2019, 74, 1015–1024. [Google Scholar] [CrossRef]
- Kong, E.; Kim, H.D.; Kim, J. Deleting key autophagy elongation proteins induces acquirement of tumor-associated phenotypes via ISG15. Cell Death Differ. 2020, 27, 2517–2530. [Google Scholar] [CrossRef]
- Joo, Y.J.; Kim, J.H.; Kang, U.B.; Yu, M.H.; Kim, J. Gcn4p-mediated transcriptional repression of ribosomal protein genes under amino-acid starvation. EMBO J. 2011, 30, 859–872. [Google Scholar] [CrossRef]
- McCall, A.D.; Pathirana, R.U.; Prabhakar, A.; Cullen, P.J.; Edgerton, M. Candida albicans biofilm development is governed by cooperative attachment and adhesion maintenance proteins. NPJ Biofilms Microbiomes 2019, 5, 21. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Joo, Y.J.; Kim, J. Asc1p, a ribosomal protein, plays a pivotal role in cellular adhesion and virulence in Candida albicans. J. Microbiol. 2010, 48, 842–848. [Google Scholar] [CrossRef]
- Navarathna, D.H.; Nickerson, K.W.; Duhamel, G.E.; Jerrels, T.R.; Petro, T.M. Exogenous farnesol interferes with the normal progression of cytokine expression during candidiasis in a mouse model. Infect. Immun. 2007, 75, 4006–4011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoyer, L.L. The ALS gene family of Candida albicans. Trends Microbiol. 2001, 9, 176–180. [Google Scholar] [CrossRef]
- Moyes, D.L.; Wilson, D.; Richardson, J.P.; Mogavero, S.; Tang, S.X.; Wernecke, J.; Hofs, S.; Gratacap, R.L.; Robbins, J.; Runglall, M.; et al. Candidalysin is a fungal peptide toxin critical for mucosal infection. Nature 2016, 532, 64–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkel, J.S.; Mitchell, A.P. Genetic control of Candida albicans biofilm development. Nat. Rev. Microbiol. 2011, 9, 109–118. [Google Scholar] [CrossRef]
- Han, T.L.; Cannon, R.D.; Villas-Boas, S.G. The metabolic basis of Candida albicans morphogenesis and quorum sensing. Fungal Genet. Biol. 2011, 48, 747–763. [Google Scholar] [CrossRef]
- Sharkey, L.L.; McNemar, M.D.; Saporito-Irwin, S.M.; Sypherd, P.S.; Fonzi, W.A. HWP1 functions in the morphological development of Candida albicans downstream of EFG1, TUP1, and RBF1. J. Bacteriol. 1999, 181, 5273–5279. [Google Scholar] [CrossRef] [Green Version]
- Carlisle, P.L.; Kadosh, D. Candida albicans Ume6, a filament-specific transcriptional regulator, directs hyphal growth via a pathway involving Hgc1 cyclin-related protein. Eukaryot. Cell 2010, 9, 1320–1328. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.N.; Morschhauser, J. Tetracycline-inducible gene expression and gene deletion in Candida albicans. Eukaryot. Cell 2005, 4, 1328–1342. [Google Scholar] [CrossRef] [Green Version]
- Berman, J.; Sudbery, P.E. Candida Albicans: A molecular revolution built on lessons from budding yeast. Nat. Rev. Genet. 2002, 3, 918–930. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.J.; Kohler, J.R.; DiDomenico, B.; Loebenberg, D.; Cacciapuoti, A.; Fink, G.R. Nonfilamentous C. albicans mutants are avirulent. Cell 1997, 90, 939–949. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Daniels, K.J.; Oh, S.H.; Green, C.B.; Yeater, K.M.; Soll, D.R.; Hoyer, L.L. Candida albicans Als3p is required for wild-type biofilm formation on silicone elastomer surfaces. Microbiology 2006, 152, 2287–2299. [Google Scholar] [CrossRef] [PubMed] [Green Version]






| Fluconazole | Caspofungin | Molecule B | Molecule C | |
|---|---|---|---|---|
| SC5314 a | 1 | 1 | 8 | 16 |
| 12–99 b | >128 | 1 | 8 | 16 |
| 89 c | >64 | 4 | 8 | 16 |
| 177 c | 1 | 4 | 8 | 16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, Y.-K.; Shin, J.; Lee, H.-Y.; Kim, H.-D.; Kim, J. Development of Carbazole Derivatives Compounds against Candida albicans: Candidates to Prevent Hyphal Formation via the Ras1-MAPK Pathway. J. Fungi 2021, 7, 688. https://doi.org/10.3390/jof7090688
Park Y-K, Shin J, Lee H-Y, Kim H-D, Kim J. Development of Carbazole Derivatives Compounds against Candida albicans: Candidates to Prevent Hyphal Formation via the Ras1-MAPK Pathway. Journal of Fungi. 2021; 7(9):688. https://doi.org/10.3390/jof7090688
Chicago/Turabian StylePark, Young-Kwang, Jisoo Shin, Hee-Yoon Lee, Hag-Dong Kim, and Joon Kim. 2021. "Development of Carbazole Derivatives Compounds against Candida albicans: Candidates to Prevent Hyphal Formation via the Ras1-MAPK Pathway" Journal of Fungi 7, no. 9: 688. https://doi.org/10.3390/jof7090688